Introduction
Human embryonic stem cells (hESCs) were first
isolated and cultured in 1998 (1). A
characteristic feature of hESCs is their capacity to self-renew and
differentiate into numerous types of cells. Therefore, hESCs may
potentially be used in the therapeutic management of major human
diseases, such as neurodegeneration, diabetes and cardiovascular
diseases (2). In order to improve
the application of stem cell-based strategies for the treatment of
destructive diseases, research investigating new territories in the
context of hESC maintenance and differentiation is undergoing
(3). The prerequisites for the
clinical application of hESCs are as follows. Firstly, the
derivation, maintenance and differentiation of hESCs should be
accomplished under strict aseptic culture conditions. Secondly, the
protocol for hESC differentiation into a desired functional cell
type should be investigated and validated, in order to ensure the
differentiated cells are homogeneous and do not form teratomas or
cause cancer. Finally, the transplantation of hESCs or their
differentiated derivatives should not induce immune responses or
rejection (4–6).
Mature endometrium cells have a limited potential to
proliferate in vitro; however, endometrium-like cells
derived from hESCs are able to overcome this limitation. Thus,
endometrium-like cells derived from hESCs can aid the study of
signaling pathways critical to endometrial regeneration, and may
ultimately lead to the development of stem cell-based therapies.
Although conventional methods are insufficient to induce the
differentiation of hESCs into endometrium-like cells, a previous
study demonstrated successful induction of the formation of
endometrium de novo from bone marrow-derived cells (7). However, the mechanisms underlying the
induction of hESC differentiation are not fully understood.
The coordinated regulation of gene expression and
the precise interactions between neighboring cells are critical for
the specification and proper arrangement of new cell types during
tissue differentiation. This process occurs via at least three
types of cell-based interactions, namely cell-cell,
cell-extracellular matrix and cell-growth factors/signaling
molecules, within the tissues (8).
Numerous morphogenetic changes are induced by the engagement of
extracellular ligands with their respective receptors (9–11).
Soluble and insoluble signaling molecules, along with
physiochemical factors, form a tissue niche that promotes cell
differentiation toward specific lineages (12). To date, various attempts have been
made worldwide to define the optimal culture conditions for hESC
growth and differentiation, and numerous cytokines and growth
factors, such as Wnt proteins, fibroblast growth factor (FGF),
heparin, transforming growth factor (TGF)-β, insulin-like growth
factor (IGF)-II, activin A, platelet-derived growth factor (PDGF)
and neurotrophins have been identified (13–15). The
Wnt signaling pathway plays an important role in the development of
the endometrium (16). Furthermore,
Wnt4, Wnt5a and Wnt7a have been demonstrated to participate in the
early development of the reproductive system in females (17,18).
However, inducing the differentiation of hESCs into a specific cell
type, such as endometrium cells, remains a challenge.
In the present study, the differentiation potential
of hESCs into endometrium-like cells was compared under different
culture conditions. In addition, the expression of Wnt members
during differentiation was determined.
Materials and methods
Reagents
Recombinant human epidermal growth factor (EGF),
type I collagenase, TGF-α, FGF and PDGF-BB were purchased from
Gibco Life Technologies (Grand Island, NY, USA). Recombinant human
17β-E2 and medroxyprogesterone acetate were purchased from
Sigma-Aldrich (St. Louis, MO, USA). This study was approved by the
Ethics Committee of the First Affiliated Hospital of Zhengzhou
University (Zhengzhou, China). Written consent was obtained from
the subjects who donated the blastocysts for the isolation of hESCs
and those whose endometrium tissue was used.
Preparation of human endometrial
stromal cells
Human endometrium tissues were obtained from nine
women (age, 32.6±0.8 years) who had undergone a hysteroscopy. The
female subjects had regular menstrual cycles and had not received
exogenous hormones in the three months prior to surgery. Full
thickness endometrium (~5 mm) was scraped from the myometrium and
washed in phosphate-buffered saline (PBS) containing 1%
penicillin/streptomycin (Gibco Life Technologies). The samples were
subsequently cut into small pieces and digested in medium
containing 2 mg/ml type I collagenase (Sigma-Aldrich) for 1–2 h at
37°C. The endometrial cells were cultured in medium at 37°C, under
5% CO2 in air. The homogeneity of the stromal cells and
epithelial cells was evaluated by immunostaining using specific
markers for epithelial (cytokeratin) and stromal cells (vimentin),
as described previously (19).
Following two passages, the purity of the stromal cells was
determined to be ~95%.
hESC culture and differentiation
hESCs were isolated from the inner cell mass of
in vitro fertilized blastocysts, which were superfluous to
in vitro fertilization cycles (20). hESC lines, ZZU-hESCs-2 and
ZZU-hESCs-3 (Zhengzhou University), were cultured in knockout
Dulbecco's modified Eagle's medium (DMEM; Gibco Life Technologies),
containing 20% knockout serum replacement, 1% non-essential amino
acids, 2 mM L-glutamine, 0.1 mM β-mercaptoethanol and 8 ng/ml FGF.
The medium was changed daily and the hESCs were passaged using a
mechanical method (a syringe tip cut the hESC clusters into
uniform-sized cell clumps for seeding on a new feeder layer) every
4–5 days, as previously described (21).
Differentiation of hESCs into endometrium-like cells
was performed using three methods, including induction by feeder
cells, co-culture with endometrial stromal cells and embryoid body
(EB) induction. The feeder cells method included the
differentiation of hESCs via the culture of the cells on a feeder
layer of mouse embryonic fibroblasts (MEFs) for 4 days. In this
strategy, the hESC clone was grown on the feeder cells for 4 days,
after which the culture medium (CM) was changed to differentiation
medium (DM). After 7 days, the cells were cultured in DM,
containing 5% serum, for one week. From day 14, the cells were
cultured in serum-free DM for 7 days, after which the cells were
collected for the assessment of differentiation. Differentiation of
hESCs was also induced using a co-culture method, which involved
culturing hESCs on a feeder layer of MEFs in the lower compartment
of a Transwell system (Corning Life Sciences, Beijing, China),
while endometrial stromal cells were cultured in the upper
compartment. After 4 days, the CM was replaced with DM (10% serum,
1% non-essential amino acids, 10 ng/ml PDGF-BB, 10 ng/ml EGF, 10
ng/ml TGF-α and 10−8 mol/l 17β-E2 in DMEM/nutrient
mixture F12) and cultured for 7 days. An additional differentiation
strategy investigated was the initiation of EB formation. In this
procedure, the hESC clone was isolated and an EB was formed, which
was subsequently cultured in a 0.1% gelatin-coated dish with DM for
7 days. Following differentiation, the cells were decidualized by
culture with DM containing medroxyprogesterone acetate
(10−6 mol/l) for 10 days, after which the expression of
prolactin (PRL) was detected.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was isolated using an RNA mini kit
(Qiagen, Valencia, CA, USA). First-strand cDNA was generated using
a PrimeScriptTM RT reagent kit with a gDNA Eraser
(Perfect Real Time; Takara Bio, Inc., Otsu, Japan). The PCR
reaction was carried out using 2 µl cDNA, 0.2 µl sense and
antisense primers and SYBR Select Master Mix on a 7500 FAST Real
Time PCR System (Applied Biosystems Life Technologies, Foster City,
CA, USA). The PCR parameters were set at 50°C for 2 min, 95°C for 2
min, and 40 cycles of 95°C for 15 sec and 60°C for 1 min. GAPDH was
used as the internal control. Gene expression levels were
determined using the 2−ΔΔCt
method, where ΔCt = (Ctgene - CtGAPDH) and Ct
was the threshold cycle. The specific primer pairs are listed in
Table I.
 | Table I.Sequences for specific primers. |
Table I.
Sequences for specific primers.
| Gene | Primer sequence
(5′-3′) | GenBank accession
number | Annealing temperature
(°C) |
|---|
| Wnt4 | F:
GCTGGGCTCCAAGTACACC | NM_030761.4 | 60 |
|
| R:
GGCTATCCTGACACACATGC |
|
|
| Wnt5a | F:
TTACCACTGCAACTATTGCACC | NM_003392.4 | 62 |
|
| R:
CACAATGAACCTTTAGTTTCCA |
|
|
| Wnt7a | F:
CCTGGAGGAGAACATGAAGC | NM_004625.3 | 63 |
|
| R:
CAGTAATTGGGTGACTTCTCG |
|
|
| CK-18 | F:
GGAAGATGGCGAGGACTTTA | NM_199187.1 | 59 |
|
| R:
AACTTTGGTGTCATTGGTCTC |
|
|
| EPCAM | F:
TGCTGTTATTGTGGTTGTGGTG | NM_002354.2 | 61 |
|
| R:
TACTTTGCCATTCTCTTCTTTCTGG |
|
|
| ER-α | F:
TGCCAAGGAGACTCGCTA | NM_001122742.1 | 60 |
|
| R:
TCAACATTCTCCCTCCTC |
|
|
| PR | F:
ACACAAAACCTGACACCTCC | NM_001271161.2 | 60α |
|
| R:
TACAGCATCTGCCCACTGAC |
|
|
| PRL | F:
GGTGGCGACGACTCCTGGAGCCC | NM_000948.5 | 61 |
|
| R:
GACACCAGACCAACTGGTAATG |
|
|
| GAPDH | F:
AGAAGGCTGGGGCTCATTTG | NM_002046.4 | 59 |
|
| R:
AGGGGCCATCCACAGTCTTC |
|
|
Immunofluorescence (IF)
On days 7, 14 and 21, the differentiated cells were
washed with 1X PBS and fixed with 4% paraformaldehyde for 10 min at
room temperature. Following three washes with 1X PBS, the cells
were permeabilized with 0.1% Triton X-100 in 1X PBS for 15 min, and
blocked with 10% horse serum (Gibco Life Technologies) for 30 min
at room temperature. Subsequently, the cell samples were incubated
with mouse anti-cytokeratin (1:50; cat. no. P2871; Sigma-Aldrich)
and rabbit anti-vimentin (1:50; Santa Cruz Biotechnology, Inc.,
Dallas, TX, USA) antibodies, or mouse anti-epithelial cell adhesion
molecule (EPCAM; 1:50; cat. no. 20160; Abcam, Cambridge, MA, USA)
and rabbit anti-vimentin (1:50; cat. no. sc-5565; Santa Cruz
Biotechnology, Inc.) antibodies overnight at 4°C. Following three
further washes with 1X PBS, the cells were incubated with an
appropriate secondary antibody (iFluor 488 goat anti-mouse IgG,
monoclonal, cat. no. AAT-16448; or Cy3 AffiniPure goat anti-mouse
IgG, monoclonal, cat. no. A22210; 1:800; Jackson Immunoresearch
Laboratories, Inc., West Grove, PA, USA) for 1 h at room
temperature, and the nuclei were stained with
4′,6-diamidino-2-phenylindole (1:5,000, Invitrogen, Life
Technologies). The sections were mounted with Prolong medium (Life
Technologies), and the images were captured using an LSM 700
inverted confocal microscope (Carl Zeiss Shanghai Co. Ltd.,
Shanghai, China).
Flow cytometry
The differentiated phenotype of endometrium-like
cells was determined using flow cytometry (BD FACS Aria Flow
Cytometer; BD Biosciences, San Jose, CA, USA). Differentiated cells
were trypsinized and stained with fluorescein
isothiocyanate-labeled cytokeratin (BD Biosciences) or Alexa Fluor®
488 labeled-EPCAM (Abcam), and analyzed by flow cytometry.
Statistical analysis
Statistical analyses were conducted by SPSS software
(version 13.0; SPSS Inc., Chicago, IL, USA). A paired, two-tailed
Student's t-test and analyses of variance were performed to
determine the statistical significance of the differences between
conditions. P<0.05 was considered to indicate a statistically
significant difference.
Results
Co-culture with endometrial stromal
cells exhibits the highest efficiency for hESC differentiation into
endometrium-like cells
Cell-cell interactions play an essential role in the
differentiation of hESCs into specific cell types. In the present
study, the differentiation potential of hESCs into endometrium-like
cells was investigated using three methods, namely induction by
feeder cells, co-culture of hESCs with endometrial stromal cells
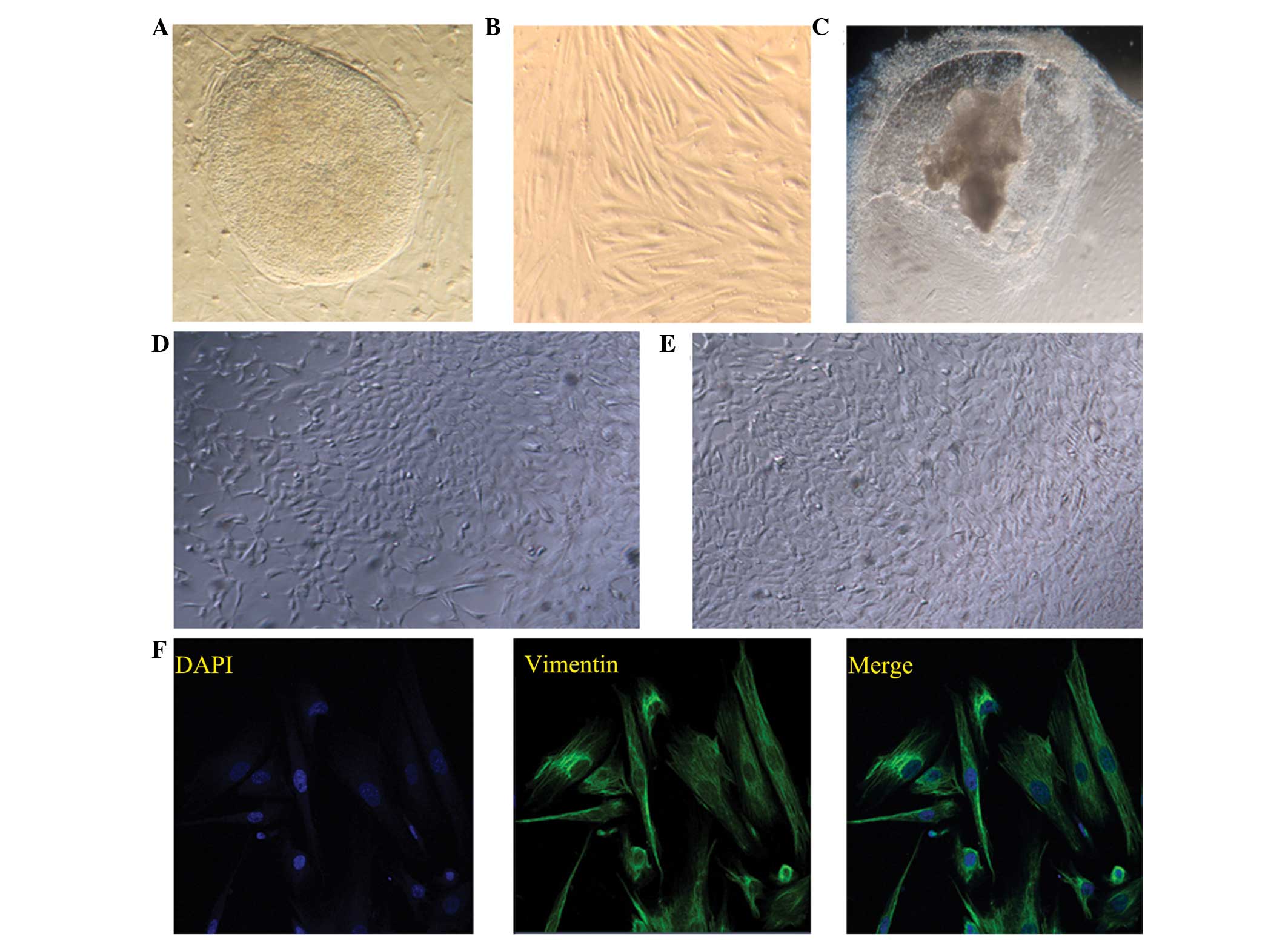
and EB induced-differentiation. The typical morphology of a hESC
clone and endometrial stromal cells is shown in Fig. 1A and B, respectively. The
morphologies of the differentiated cells at day 21 following the
various methods of differentiation induction are shown in Fig. 1C–E.
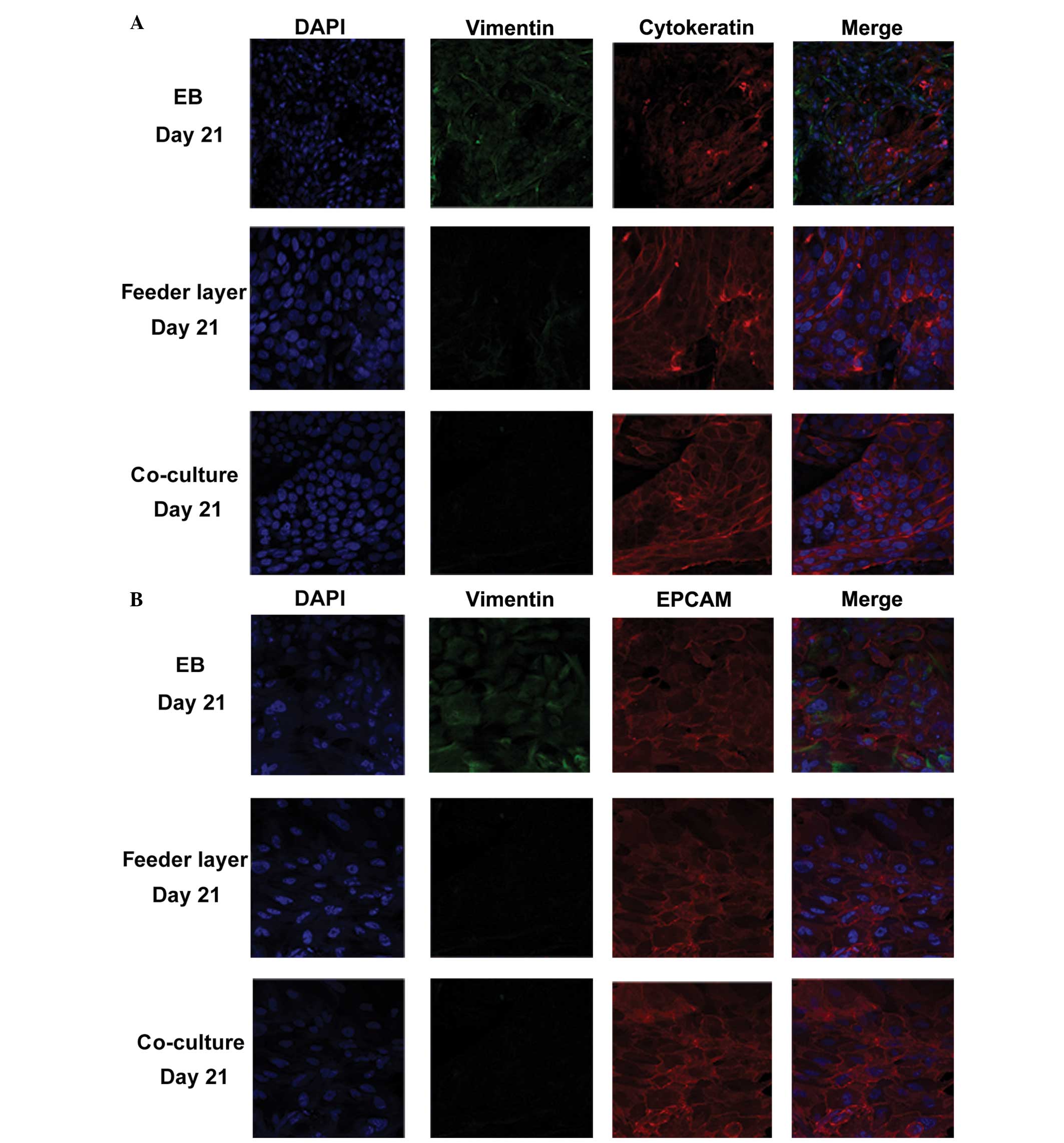
To evaluate the differentiation potential of hESCs
into endometrium-like cells, the protein expression levels of
vimentin, EPCAM and cytokeratin were determined using IF staining.
Cytokeratin and EPCAM are markers for epithelial cells, while
vimentin is a marker for stromal cells. In the co-culture group,
>85% of the cells expressed epithelial cell markers and ~10% of
the cells were positive for vimentin (n=100), indicating that
although the majority of these differentiated cells were epithelial
cells, stromal cells were also present (Fig. 2A and B). In the feeder layer group,
almost 70% of the cells were cytokeratin-positive (n=100 cells);
however, in the EB group, only ~40% of the cells were
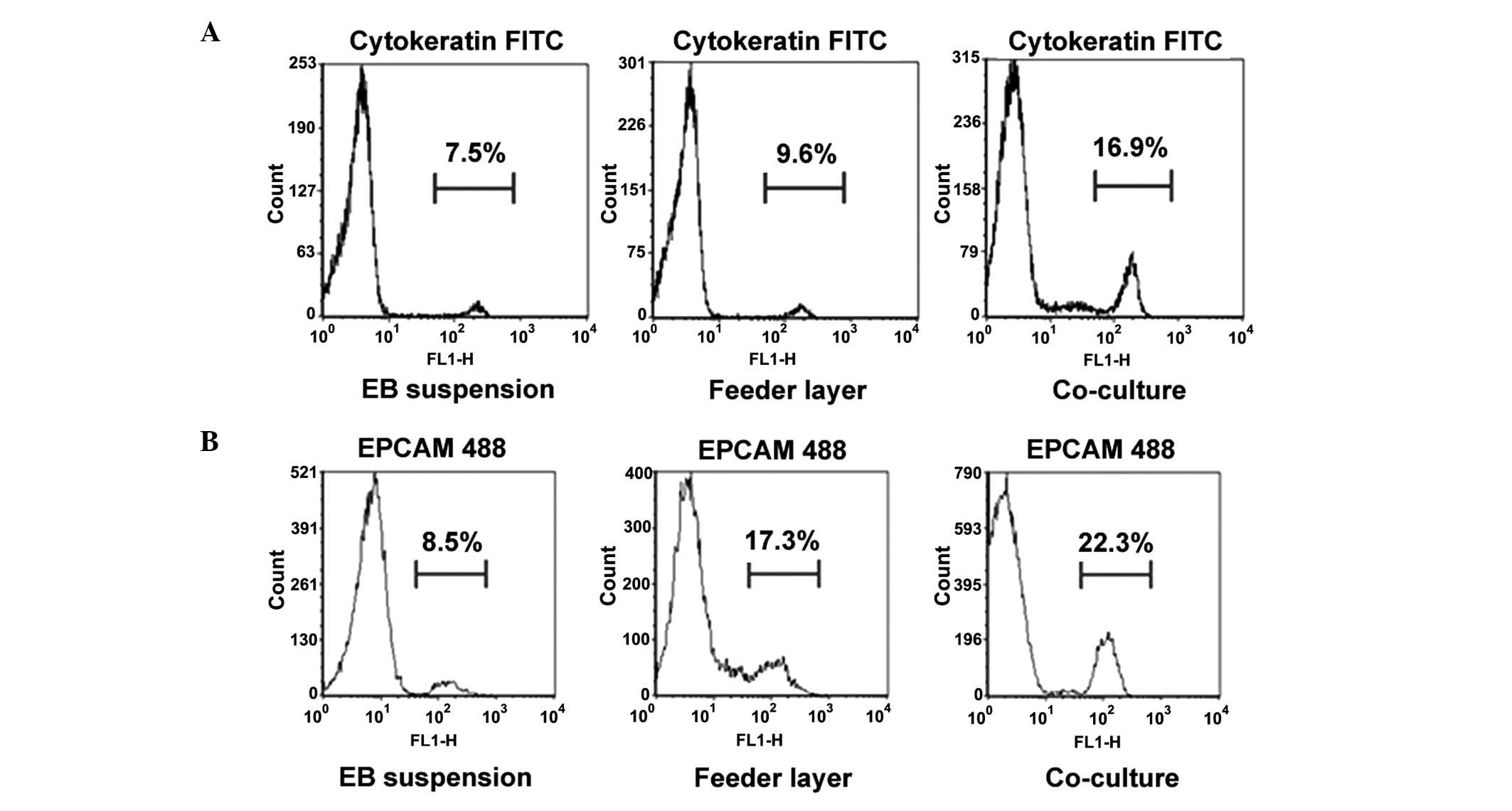
cytokeratin-positive (n=100 cells). These observations were further
confirmed through flow cytometry (Fig.
3). The numbers of cytokeratin- and EPCAM-positive cells were
highest in the co-culture group, when compared with those in the
other two groups (P<0.05).
RT-qPCR analysis was further performed to detect the
mRNA expression levels of cytokeratin, EPCAM, estrogen receptor
(ER) and progesterone receptor (PR). The expression levels of these
genes were significantly increased at day 21 following
differentiation induction using the co-culture method, as compared
with the other two strategies (P<0.05; Fig. 4). The positive expression of ER and
PR indicated that the differentiated cells in the co-culture group
may be mediated by estrogen and progesterone hormones. Moreover,
the differentiated cells were decidualized by culture with
medroxyprogesterone acetate for 10 days, after which the mRNA
expression of PRL was detected. The expression of PRL was highest
in the co-culture group (P<0.05; Fig.
4), indicating that the differentiated cells in the co-culture
group exhibited a superior tendency to decidualization.
 | Figure 4.Human embryonic stem cell
differentiation into endometrium-like cells. mRNA expression levels
of CK18, EPCAM, ER, PR and PRL were analyzed in the differentiated
cells on day 21 following induction by EB, mouse embryonic
fibroblast feeder cells or co-culture with endometrial stromal
cells. Data are presented as the mean ± standard deviation (n=3).
*P<0.05, vs. respective EB group; #P<0.05, vs.
respective feeder layer group. EPCAM, epithelial cell adhesion
molecule; ER, estrogen receptor; PR, progesterone receptor; PRL,
prolactin; EB, embryonic body; CK, cytokeratin. |
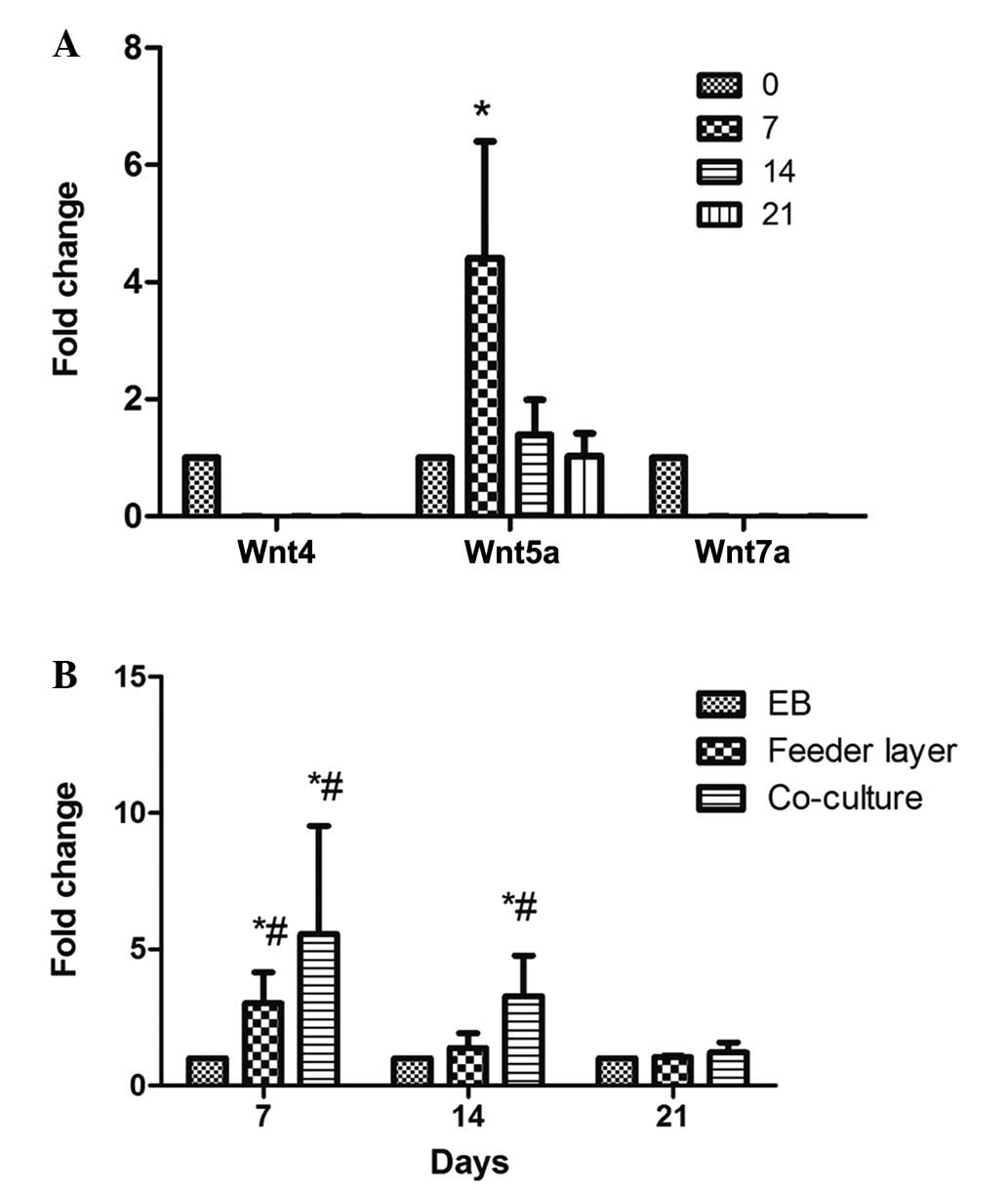
Wnt5a is upregulated following hESC
differentiation into endometrium-like cells
Wnt4, Wnt5a and Wnt7a have been demonstrated to play
important roles in the development of the reproductive system in
females. Wnt4 promotes early gender development, Wnt5a promotes
uterine gland development and Wnt7a maintains the uterus in normal
morphology (22). MEF feeder cells
have been found to secrete Wnt proteins (23). Therefore, in order to prevent
interference by these proteins secreted by feeder cells or
endometrial stromal cells, the mRNA expression levels of Wnt4,
Wnt5a and Wnt7a in the EB-induced differentiated cells were
examined on days 0, 7, 14 and 21. The mRNA expression levels of
Wnt4 and Wnt7a were very low, and no statistically significant
differences were observed during the process of differentiation
(P>0.05; Fig. 5A). However, Wnt5a
was significantly upregulated at days 7 and 14 following
differentiation induction, with the highest level observed on day 7
(P<0.05). This observation indicated that Wnt5a may be
associated with the differentiation of hESCs into endometrium-like
cells. Accordingly, further investigation into the expression of
Wnt5a in the differentiated cells obtained by the various methods
was carried out. When compared with the other two methods, the
co-culture system exhibited a significant upregulation (>30
fold) of Wnt5a expression on day 7 following differentiation
induction, followed by a gradual decrease (P<0.05; Fig. 5B). Furthermore, the mRNA expression
levels of Wnt5a were shown to significantly increase between days 7
and 14 following the induction of differentiation in the feeder
layer group; however, the expression on day 7 was lower compared
with that in the co-culture group (P>0.05; Fig. 5B).
Discussion
The self-renewal and differentiation of hESCs
requires a large number of endogenous proteins produced by hESCs
and exogenous factors in the culture medium (1–3).
Routinely, hESCs are cultured on feeder layers of MEFs or on
cell-free matrices (laminin or matrigel) supplemented with
MEF-conditioned medium (24). An
additional approach is to culture hESCs in suspension conditions
that enable hESCs to aggregate and form an EB that is capable of
differentiating into three primary germ layers (endoderm, mesoderm
and ectoderm), which may be induced to directly differentiate
toward the lineage of interest by the induction of growth factors
in the culture system (25).
However, previous studies have successfully established co-culture
systems for hESC differentiation (25). For example, Udayashankar et
al (27) successfully generated
a cytotrophoblast cell line through co-culture of hESCs with an
established endometrial cell culture system. To the best of our
knowledge, the present study was the first to compare the
differentiation potential of hESCs into endometrium-like cells
using three methods, namely EB induced-differentiation, feeder
cells and a co-culture system.
In the EB group, the undifferentiated hESCs
gradually grew into small cell aggregates and formed a suitably
sized EB in suspension conditions. These EB-derived hESCs were able
to further differentiate into endometrium-like cells, although the
efficacy was lower compared with the other two methods. In
addition, MEFs (feeder cells) were demonstrated to promote the
differentiation potential of hESCs into endometrium-like cells,
which further supported the hypothesis that MEFs contribute to hESC
differentiation. Previously, the Transwell co-culture system has
been successfully applied to the differentiation of different types
of stem cell, such as hESCs, human bone marrow stem cells and human
mesenchymal stem cells (27–29). In the present study, the co-culture
of hESCs with human endometrial stromal cells was shown to strongly
enhance the differentiation potential of hESCs into
endometrium-like cells, to a greater extent compared with the other
two methods. These observations indicated that the co-culture
system may provide superior niches and an improved microenvironment
for hESC attachment, expansion and differentiation.
Normal endometrial tissue in the luteal phase is
decidualized under the function of progesterone. In addition to
changes in cell morphology, the functions of the cells are also
altered, such as secretory granules appearing on the cell surface
and the secretion of PRL and IGF-binding protein 1. To further
investigate the function of the differentiated cells induced by the
various methods, the cells were cultured with medroxyprogesterone
acetate for 10 days, after which the mRNA expression levels of PRL
were detected by RT-qPCR. Although PRL was found to be expressed in
all three groups, the expression level was highest in the
co-culture group, which is proportional to the efficiency of
induced differentiation, indicating that the differentiated cells
in the co-culture group exhibited a superior function and were the
most sensitive to progesterone.
Yi et al (30)
investigated the role of the Wnt signaling pathway in ESCs and
showed that transcription factor-3, as a downstream target of the
Wnt signaling pathway, may inhibit the promoter activity of Nanog
and reduce its expression, subsequently affecting the self-renewal
capacity of ESCs and promoting their differentiation. As Wnt
signaling is required for embryonic development in invertebrates
and vertebrates (31–33), the focus in the present study was on
three Wnt members associated with cell proliferation, migration and
differentiation. Wnt4, Wnt5a and Wnt7a have also been hypothesized
to play important roles in the development of the female
reproductive system (16–18,22). In
the present study, Wnt5a was revealed to be significantly
upregulated in the differentiated cells induced by feeder cells or
co-cultured with endometrial stromal cells; however, the expression
levels of Wnt4 and Wnt7a were not affected during the
differentiation of hESCs into endometrium-like cells induced by the
three different methods. Furthermore, the expression level of Wnt5a
was consistent with the data regarding the differentiation
efficiency in the three groups. Based on the present findings, the
co-culture system with endometrial stromal cells was demonstrated
to provide the hESCs with a different microenvironment, which lead
to an increased activity of the Wnt signaling pathway in the hESCs,
through which the efficiency of hESC differentiation into
endometrium-like cells was promoted.
In conclusion, the three methods investigated in the
present study are able to induce the differentiation of hESCs into
endometrium-like cells, with the co-culture system exhibiting the
highest efficiency. In addition, Wnt5a expression was found to be
significantly upregulated during hESC differentiation into
endometrial epithelial-like cells. These observations indicate that
the co-culture system is optimal for the induction of hESC
differentiation into endometrial cells, and that the Wnt5a
signaling may be involved in this process.
References
|
1
|
Thomson JA, Itskovitz-Eldor J, Shapiro SS,
et al: Embryonic stem cell lines derived from human blastocysts.
Science. 282:1145–1147. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Hu Q and Rosenfeld MG: Epigenetic
regulation of human embryonic stem cells. Front Genet. 3:2382012.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Gill KP, Hewitt AW, Davidson KC, et al:
Methods of retinal ganglion cell differentiation from pluripotent
stem cells. Transl Vis Sci Technol. 3:72014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Lee JE and Lee DR: Human embryonic stem
cells: Derivation, maintenance and cryopreservation. Int J Stem
Cells. 4:9–17. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Unger C, Skottman H, Blomberg P, Dilber MS
and Hovatta O: Good manufacturing practice and clinical-grade human
embryonic stem cell lines. Hum Mol Genet. 17:R48–R53. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Mountford JC: Human embryonic stem cells:
Origins, characteristics and potential for regenerative therapy.
Transfus Med. 18:1–12. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Du H and Taylor HS: Contribution of bone
marrow-derived stem cells to endometrium and endometriosis. Stem
Cells. 25:2082–2086. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Cai L, Ye Z, Zhou BY, Mali P, Zhou C and
Cheng L: Promoting human embryonic stem cell renewal or
differentiation by modulating Wnt signal and culture conditions.
Cell Res. 17:62–72. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Brafman DA, Phung C, Kumar N and Willert
K: Regulation of endodermal differentiation of human embryonic stem
cells through integrin-ECM interactions. Cell Death Differ.
20:369–381. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Tran NT, Trinh QM, Lee GM and Han YM:
Efficient differentiation of human pluripotent stem cells into
mesenchymal stem cells by modulating intracellular signaling
pathways in a feeder/serum-free system. Stem Cells Dev.
21:1165–1175. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Brafman DA, Chang CW, Fernandez A, Willert
K, Varghese S and Chien S: Long-term human pluripotent stem cell
self-renewal on synthetic polymer surfaces. Biomaterials.
31:9135–9144. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Jin S, Yao H, Krisanarungson P, Haukas A
and Ye K: Porous membrane substrates offer better niches to enhance
the Wnt signaling and promote human embryonic stem cell growth and
differentiation. Tissue Eng Part A. 18:1419–1430. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Atkinson SP, Lako M and Armstrong L:
Potential for pharmacological manipulation of human embryonic stem
cells. Br J Pharmacol. 169:269–289. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Dravid G, Ye Z, Hammond H, et al: Defining
the role of Wnt/beta-catenin signaling in the survival,
proliferation and self-renewal of human embryonic stem cells. Stem
Cells. 23:1489–1501. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Pyle AD, Lock LF and Donovan PJ:
Neurotrophins mediate human embryonic stem cell survival. Nat
Biotechnol. 24:344–350. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
16
|
Hayashi K and Spencer TE: WNT pathways in
the neonatal ovine uterus: potential specification of endometrial
gland morphogenesis by SFRP2. Biol Reprod. 74:721–733. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Pellegrino M, Maiorino R and Schonauer S:
WNT4 signaling in female gonadal development. Endocr Metab Immune
Disord Drug Targets. 10:168–174. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hayashi K, Yoshioka S, Reardon SN, et al:
WNTs in the neonatal mouse uterus: Potential regulation of
endometrial gland development. Biol Reprod. 84:308–319. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Malayer JR and Woods VM: Expression of
estrogen receptor and maintenance of hormone-responsive phenotype
in bovine fetal uterine cells. Domest Anim Endocrinol. 15:141–154.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Thomson JA, Itskovitz-Eldor J, Shapiro SS,
et al: Embryonic stem cell lines derived from human blastocysts.
Science. 282:1145–1147. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Chen XM, Kan QC, Wang F, et al: Chromosome
dynamic changes in two cultured Chinese human embryonic stem cell
lines: Single nucleotide polymorphism, copy number variation and
loss of heterozygosity. J Cell Biochem. 113:3520–3527. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Heikkila M, Peltoketo H and Vainio S: Wnts
and the female reproductive system. J Exp Zool. 290:616–623. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Xie CQ, Lin G, Luo KL, et al: Newly
expressed proteins of mouse embryonic fibroblasts irradiated to be
inactive. Biochem Biophys Res Commun. 315:581–588. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Sarkar P, Randall SM, Muddiman DC and Rao
BM: Targeted proteomics of the secretory pathway reveals the
secretome of mouse embryonic fibroblasts and human embryonic stem
cells. Mol Cell Proteomics. 11:1829–1839. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Liu Y, Fox V, Lei Y, Hu B, Joo KI and Wang
P: Synthetic niches for differentiation of human embryonic stem
cells bypassing embryoid body formation. J Biomed Mater Res B Appl
Biomater. 102:1101–1112. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Amirpour N, Nasr-Esfahani MH, Esfandiari
E, Razavi S and Karamali F: Comparing three methods of co-culture
of retinal pigment epithelium with progenitor cells derived human
embryonic stem cells. Int J Prev Med. 4:1243–1250. 2013.PubMed/NCBI
|
|
27
|
Udayashankar R, Baker D, Tuckerman E,
Laird S, Li TC and Moore HD: Characterization of invasive
trophoblasts generated from human embryonic stem cells. Hum Reprod.
26:398–406. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Murdoch AD, Grady LM, Ablett MP, Katopodi
T, Meadows RS and Hardingham TE: Chondrogenic differentiation of
human bone marrow stem cells in Transwell cultures: Generation of
scaffold-free cartilage. Stem Cells. 25:2786–2796. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Yang Y, Li J, Pan X, et al: Co-culture
with mesenchymal stem cells enhances metabolic functions of liver
cells in bioartificial liver system. Biotechnol Bioeng.
110:958–968. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Yi F, Pereira L and Merrill BJ: Tcf3
functions as a steady-state limiter of transcriptional programs of
mouse embryonic stem cell self-renewal. Stem Cells. 26:1951–1960.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Clevers H: Wnt/beta-catenin signaling in
development and disease. Cell. 127:469–480. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
van Amerongen R and Nusse R: Towards an
integrated view of Wnt signaling in development. Development.
136:3205–3214. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Angers S and Moon RT: Proximal events in
Wnt signal transduction. Nat Rev Mol Cell Biol. 10:468–477.
2009.PubMed/NCBI
|