Introduction
Allergic rhinitis (AR) is an atopic disease
characterized by the clinical symptoms of sneezing, itching,
rhinorrhea and nasal blockage. Antigen provocation tests have
indicated that a biphasic reaction occurs in the respiratory tract.
The early phase of the immediate reaction starts minutes after
allergen provocation. The nasal symptoms reappear 5–10 h after
allergen provocation, which is known as the late-phase reaction
(LPR) (1–4). In previous studies, the majority of
patients suffering from AR showed the above symptoms immediately,
while >50% of them developed an LPR following antigen challenge
(1–4). It has been reported that sneezing and
rhinorrhea are the strongest symptoms during the allergic LPR
(5–7), and studies have found that >90% of
patients showed nasal blockage in the LPR (8,9). There
are numerous animal models that can be used to study the LPR of AR
(10,11); however, these only show nasal
blockage in the LPR. To date, no animal models have been reported
to show AR with early- and late-phase sneezing.
Staphylococcal enterotoxin B (SEB), produced by
Staphylococcus aureus, is an exotoxin that may act as
allergen or superantigen to induce allergic disease. We have
previously described an AR model developed by repeated intranasal
instillation with SEB in guinea pigs (12). The model, which exhibited the typical
symptoms of AR, showed sneezing and nasal scratching, thereby
demonstrating that SEB has the potential to act as a conventional
allergen to induce AR. The nasal cavity and skin of humans are the
most common sites for Staphylococcus aureus colonization
(13,14). In excess of 50% of pathogenic
isolates of Staphylococcus aureus produce one or more
superantigen exotoxins (15). This
increases the risk that the atopic individual suffers from allergic
disease. In the present study, the aim was to describe a novel
method for establishing experimental AR with early and late
sneezing with SEB in guinea pigs.
Materials and methods
Animals and materials
Male, eight-week-old, healthy Hartley guinea pigs
(250–300 g) were purchased from the Laboratory Animal Center of
Chongqing Medical University (Chongqing, China). The animals were
housed in an air-conditioned room in which allergen-free conditions
were maintained. The animals were fed according to the
Institutional Guidelines for the Care and Use of Laboratory
Animals. All experimental procedures on animals were approved by
the Ethics Committee of Chongqing Medical University. SEB and
terfenadine were purchased from Sigma (St. Louis, MO, USA).
Sensitization and challenge
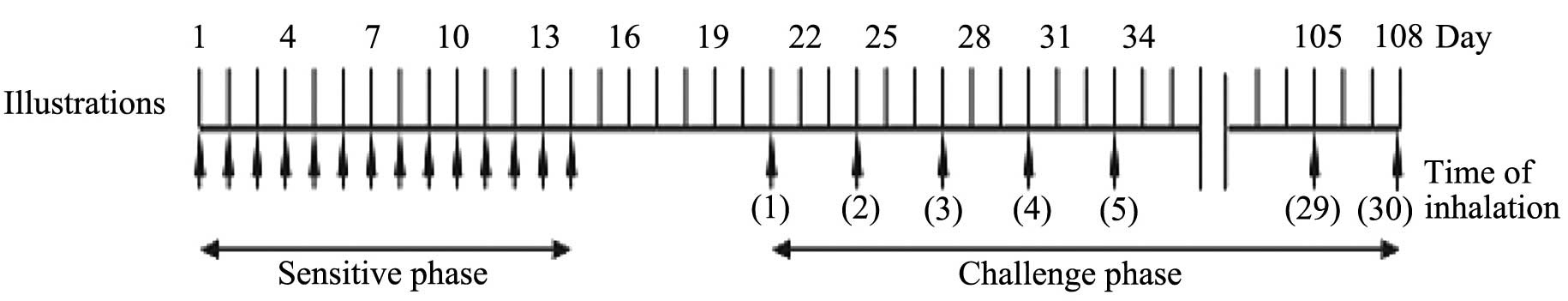
As shown in Fig. 1,
guinea pigs (n=10 per group) were intranasally sensitized with 1 µg
SEB dissolved in 40 µl saline in the absence of adjuvant once every
day for 14 days. Prior to each sensitization, the upper airway
mucosal surface was anesthetized by intranasal instillation of 4%
lidocaine hydrochloride solution (China Otsuka Pharmaceutical Co.,
Ltd., Tianjin, China) to prevent the rapid elimination of antigen
by ciliary movement (8). One week
after the final sensitization, the same treatment was applied
intranasally once every four days for a total of 30 times. For the
control group, the animals were treated with the same volume of
saline instead of SEB. In addition, the effects of histamine in AR
were evaluated. In the treatment group, terfenadine (20 mg/kg) was
administered orally 70 min before the 4th, 14th and 24th challenge.
Terfenadine is an H1 receptor antagonist. It has been reported that
terfenadine at the above doses significantly inhibits allergic
inflammation of the nasal cavity induced by histamine in guinea
pigs (16).
Observation of symptoms
The number of sneezes in the intervals 0–10 min, 10
min-2 h, 2–4 h, 4–6 h, 6–8 h and 8–10 h after the SEB intranasal
challenge was counted. The nasal scratching number was measured
simultaneously. The symptom observation was performed manually in
each animal.
Serum immunoglobulin (Ig) levels
Ten hours after the final challenge, the guinea pigs
were anesthetized intramuscularly with urethane (1.35 g/kg;
Sinopharm Chemical Reagent Co., Ltd., Shanghai, China). The animals
were then sacrificed by cardiac puncture and the serum was
collected. Serum SEB-specific IgG1 levels were measured
with indirect ELISA, as previously described (12). In brief, the plates were coated (100
µl/well) with 0.1 µg/ml SEB in 0.05 M NaHCO3 (pH 9.6) at
4°C overnight. The plates were washed with phosphate-buffered
saline-Tween 20 and blocked (200 µl/well) with 2% bovine serum
albumin (BSA) for 2 h at 20°C and re-washed. Sera diluted 1:100 in
0.1% BSA were incubated (100 µl/well) at 4°C overnight. Following a
washing step, 1:500 diluted horseradish peroxidase (HRP)-goat
anti-guinea pig IgG1 monoclonal antibody (mAb)
(#AHP861P; AbD SeroTec, Oxford, UK) in 0.1% BSA was incubated (100
µl/well) at 20°C for 2 h, prior to a second washing step.
Tetramethylbenzidine chromogenic reagent (100 µl; Boster Biological
Technology, Ltd., Wuhan, China) was incubated at 20°C for 10 min
and Stop Solution (100 µl/well; Boster Biological Technology, Ltd.)
was added. Titers for SEB-specific IgG1 were estimated
as mean optical density (OD) at 450 nm. SEB-specific
IgG2 and IgE levels were measured by the same
aforementioned method. Specific IgG2 was detected with
HRP-goat anti-guinea pig IgG2 mAb (1:500; #AHP862P; AbD
SeroTec), and specific IgE was detected with HRP-goat anti-mouse
IgE mAb (1:500; #1110-05; Southern Biotech, Inc., Birmingham, AL,
USA).
Histological examination
Samples of nose, trachea, bronchi and lungs were
collected, fixed in formalin and embedded in paraffin. Transverse
sections were cut and stained with hematoxylin and eosin.
Initially, eosinophil infiltration in the nasal mucosa was observed
microscopically in a high-power field. A total of 200 leukocytes
were then counted microscopically (magnification, 10×100) in total.
The percentage of eosinophils among the total leukocytes was
counted.
Statistical analysis
All results with a normal distribution are expressed
as the mean ± standard deviation. Comparisons of means between
different groups were performed with a Student's t-test (two
groups). Data presenting in a non-normal distribution are expressed
as median (Q). Statistical analyses were performed with the
Wilcoxon test. P<0.05 was considered to indicate a statistically
significant difference.
Results
Changes in the sneezing and nasal
scratching number
Between the first and 12th challenges, the guinea
pigs in the model group produced a high sneezing number within 10
min after each challenge. From the 10th min, little or no sneezing
was observed at any challenge time. The total number of sneezes
increased with increasing challenge number (Fig. 2A-D). Between the 13th and 18th
challenges, no notable increase in sneezing number was observed
among the SEB-sensitized guinea pigs compared with the sneezing
number of the earlier challenges (Fig.
2E). Between the 19th and 30th challenges, the guinea pigs in
the model group experienced a significant biphasic increase in the
number of sneezes, reaching multiple peaks at 10 min-2 h and 4–8 h
after SEB challenge. The first peak in sneezing number shifted from
the first 10 min to the period of 10 min-2 h after SEB challenge.
In addition, the first sneezing number peak was significantly
higher between the 19th and 30th challenges than that between the
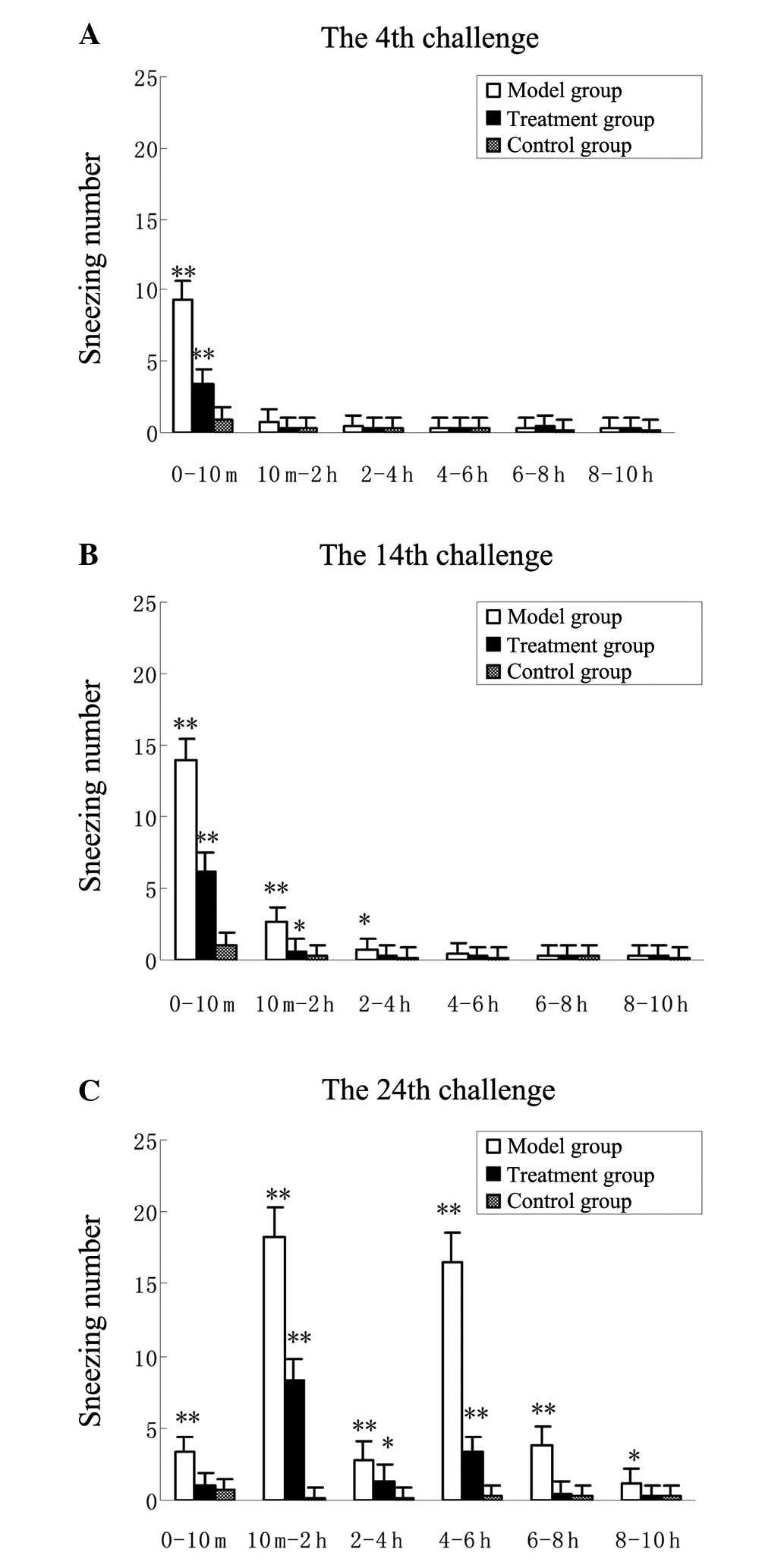
first and 18th challenges (P<0.01) (Fig. 2F-H). Furthermore, terfenadine
significantly reduced the early and the late elevations in sneezing
number at the 24th challenge (P<0.01) (Fig. 3A-C). The guinea pigs in the control
group produced significantly less sneezing than those in the model
group (P<0.01). The changes in nasal scratching number were
similar to the changes in sneezing number (data not shown).
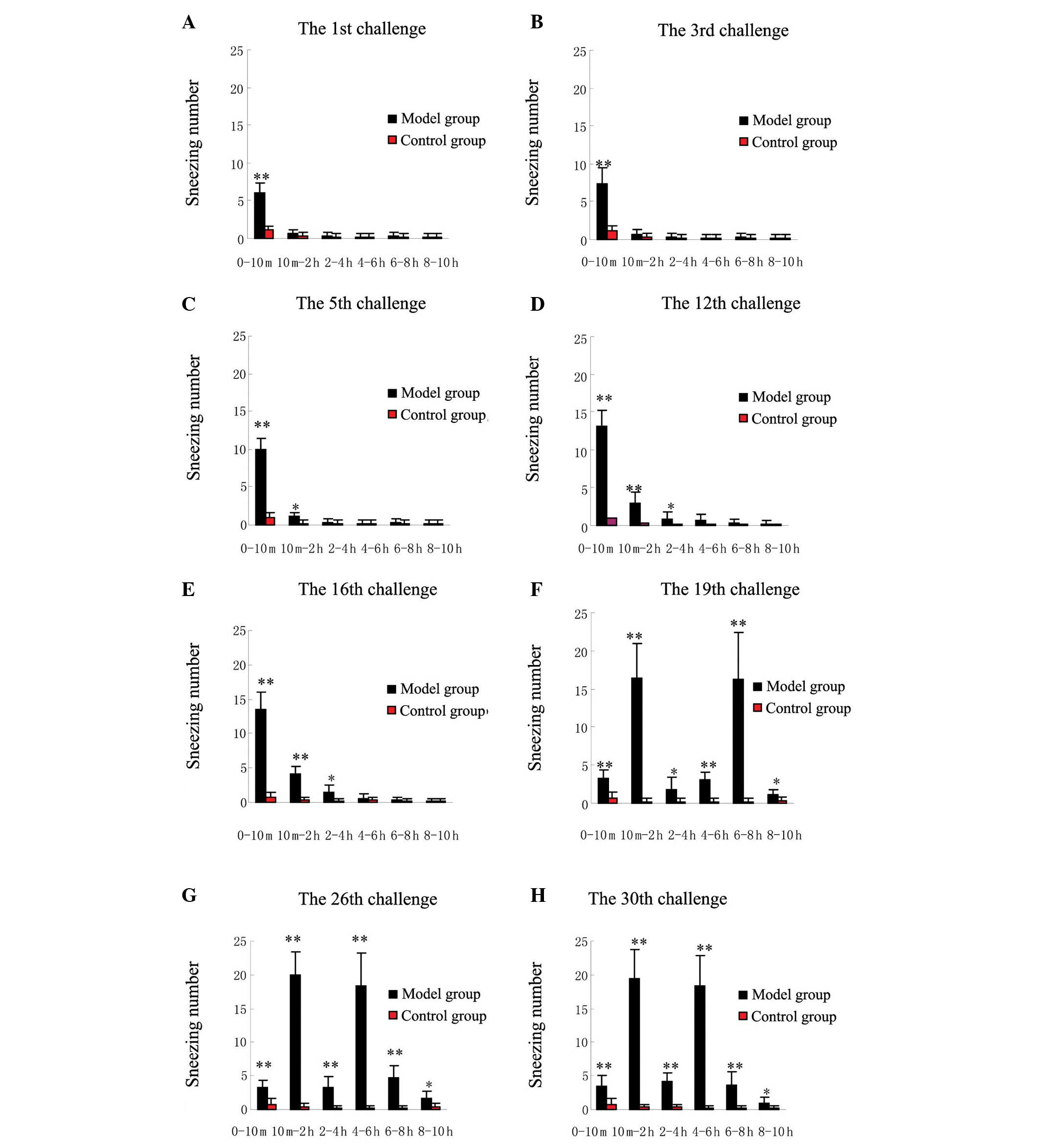 | Figure 2.Changes in sneezing number following
intranasal instillation with SEB and saline at the (A) 1st, (B)
3rd, (C) 5th, (D) 12th, (E) 16th, (F) 19th, (G) 26th and (H) 30th
challenges. Data are presented as the mean ± standard deviation
from 10 guinea pigs. Between the 1st and 18th challenges, the
guinea pigs with SEB challenge produced a significantly higher
sneezing number only within the first 10 min after each challenge.
Between the 19th and 30th challenges, the guinea pigs with SEB
challenge produced significant biphasic elevations in sneezing
number, with peaks appearing 10 min-2 h and 4–8 h after the SEB
challenge. *P<0.05 and **P<0.01 compared with the control
group. SEB, Staphylococcal enterotoxin B. |
Ig production in the serum
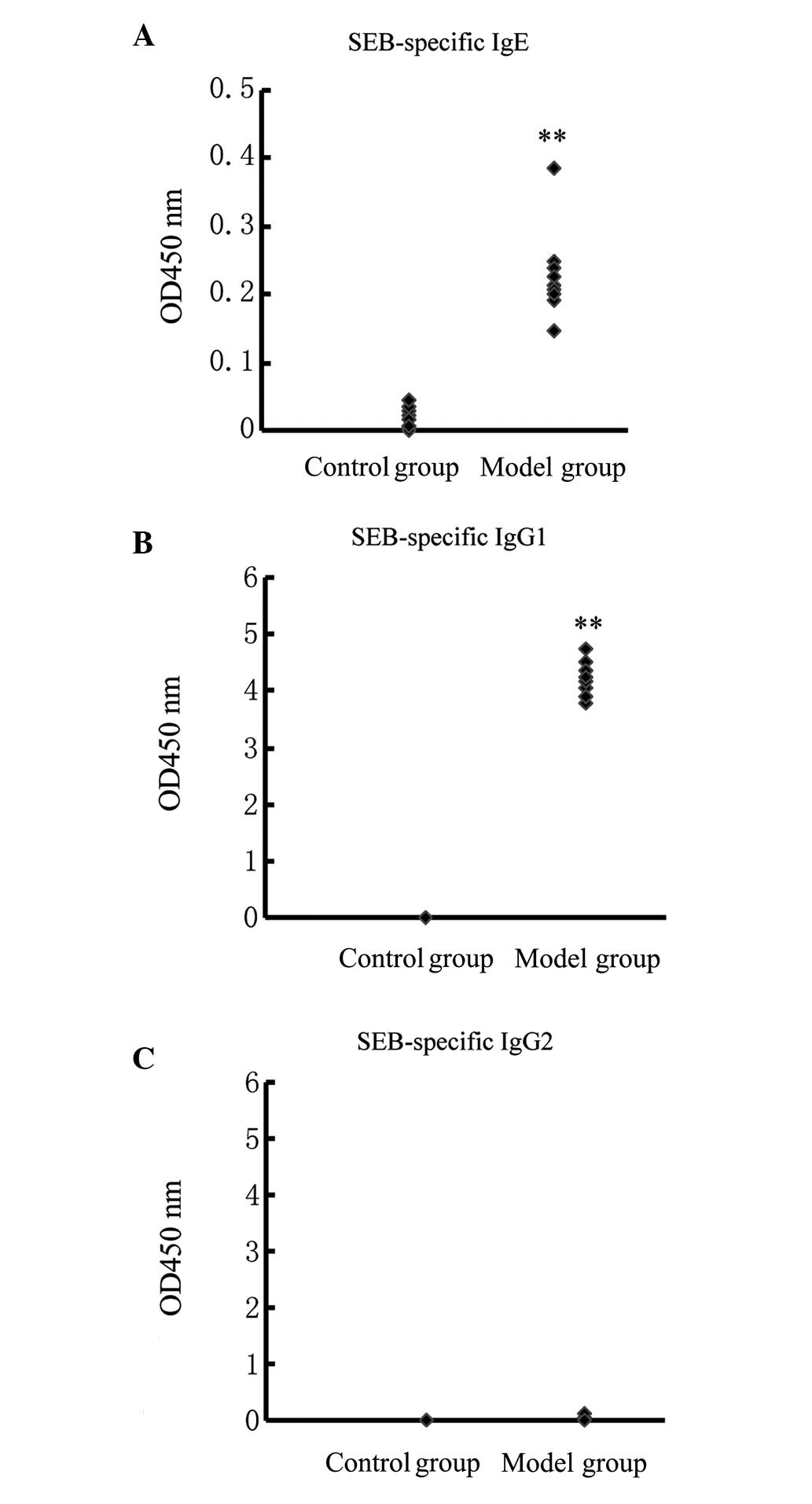
In the model group, the guinea pigs produced
increased levels of SEB-specific IgE and IgG1. The mean
ODs at 450 nm were 0.23±0.06 and 4.21±0.28, respectively. By
contrast, levels of SEB-specific IgE and IgG1 in the
control group could not be detected. Compared with the control
group, the increases observed in the model group were statistically
significant (P<0.01, Wilcoxon test) (Fig. 4A and B). In the two groups, the
guinea pigs did not produce detectable amounts of SEB-specific
IgG2 (Fig. 4C).
Eosinophils in the nasal mucosa
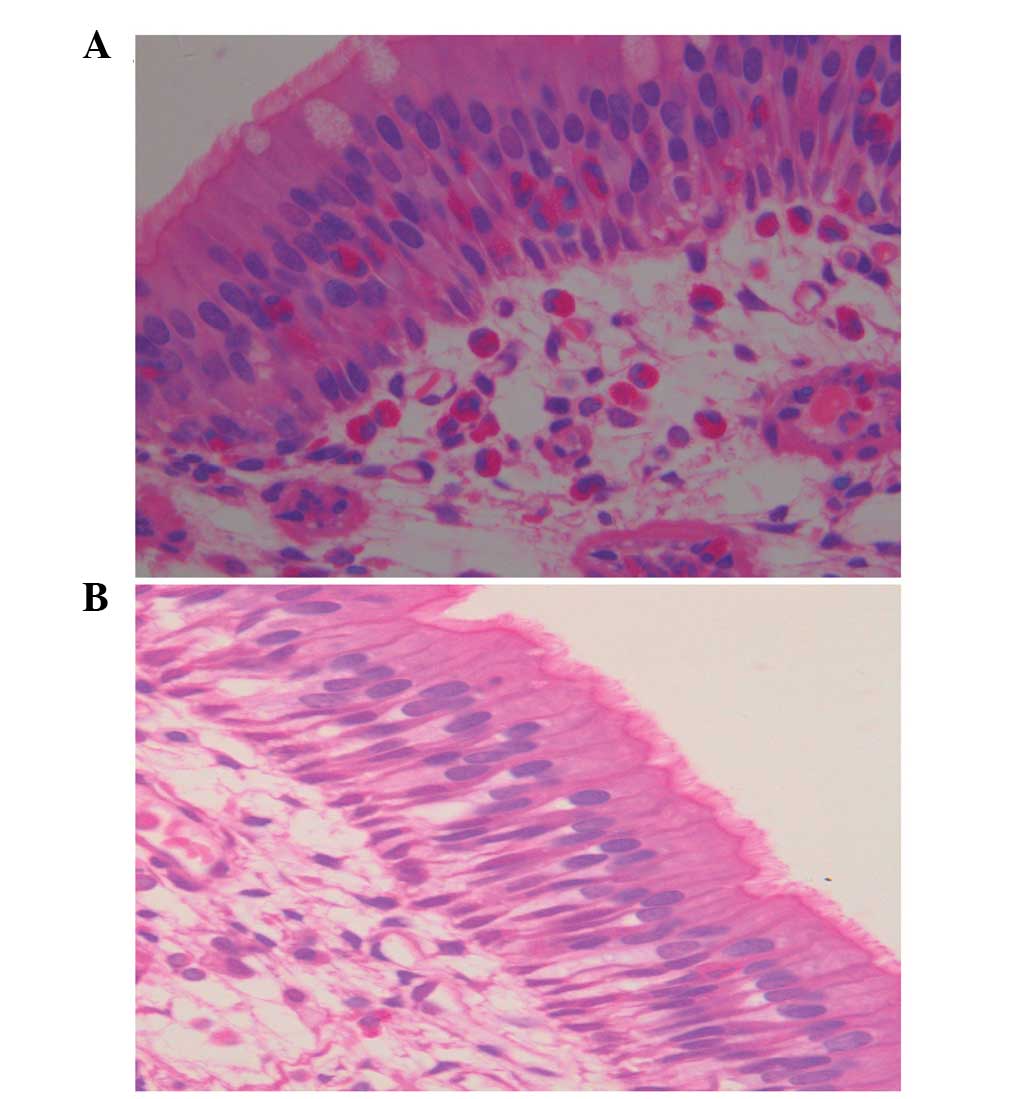
In the model group, the guinea pigs showed apparent
eosinophil infiltration in the nasal mucosa. The eosinophils
infiltrated not only the lamina propria but also the epithelial
layer (Fig. 5A); however, eosinophil
infiltration was not observed in the guinea pigs challenged with
saline (Fig. 5B). In the model
group, eosinophils accounted for 49.5% (median quartile, 0.107) of
the total leukocytes, compared with only 3.1% (median quartile,
0.015) in the control group. Compared with the control group, this
increase was statistically significant (P<0.01, Wilcoxon test).
Furthermore, vasodilatation and edema were present in the
submucosal areas among the SEB-challenged guinea pigs. The trachea,
bronchi and lungs in the two groups exhibited normal tissue without
significant pathological changes.
Discussion
AR is a global health problem. The successful study
of the pathogenesis and pathophysiology is dependent upon the
establishment of a successful animal model of AR; however no
unified standards can be used at present. According to the
literature, several sensitization methods, such as intraperitoneal
and intradermal injections and inhalation, have been used to study
AR (17). Animal models induced by
intraperitoneal and intradermal injections of allergens vary
greatly from the natural processes of AR. Furthermore, aerosol
inhalation is likely to cause sensitization of the lower
respiratory tract, which can easily induce asthma (18). In the present study, the intranasal
instillation method was used for the sensitization and challenge
processes; the aim of this was to ensure that the allergic
inflammation was confined to the nasal cavity, consistent with the
natural process of AR. Previous studies concerning animal models of
AR have reported that the time required to build an animal model
varies greatly, from one week to several months (10,11,19–25). In
general, animal models that can be established in a short time can
only reflect the early-phase symptoms; models facilitating the
observation of the LPR can take a long time to build. Nabe et
al (10) have established an
animal model of AR with the biphasic nasal blockage by Japanese
cedar pollen. In their model, biphasic nasal blockage was evoked
6–7 weeks after the first sensitization. In the present study,
biphasic sneezing was exhibited by the guinea pigs following the
19th nasal challenge (model day 75).
To date, investigations of the LPR have focused on
nasal blockage in the animal models of AR (26–28).
These reports describe that nasal blockage is a predominant symptom
in the LPR. Sneezing is a major symptom in the early-phase
reaction; however, in the present study it was demonstrated that
SEB could act as a type of allergen, by repeatedly stimulating
nasal tissue, and induce typical symptoms of AR with a biphasic
response. It was found in this study that frequent sneezing was a
significant feature of the LPR, and exhibited a biphasic response.
The present results are consistent with clinical observations
(5,6), and may suggest that frequent sneezing
is the strongest symptom during the early-phase reaction and LPR of
AR.
There is universal agreement that histamine is the
primary mediator involved in the development of sneezing (29–31).
During the course of AR, inflammatory cells release histamine. The
histamine stimulates H1 receptors of sensory nerve endings in the
nasal mucosa (32). By a nerve
reflex, multiple sneezing is caused exclusively by histamine. In
allergic inflammation, histamine is released by mast cells and
basophils. It is well established that mast cells releasing
histamine are the major cytokine source inducing sneezing in the
early-phase reaction (29). A study
by Schleimer et al (33)
investigated the inflammatory cytokines in the LPR in humans. In
the LPR, histamine and N-α-tosyl-L-arginine methyl esterase were
found in the nasal secretions of patients exhibiting an LPR
following antigen provocation tests. By contrast, prostaglandin D2
(PGD2) was not found. Since basophils do not generate PGD2,
histamine may be released by basophils in the LPR. Basophils
releasing histamine may be the main inducer of frequent sneezing in
the LPR. At the same time, the histamine contributes to the late
phase of AR by exhibiting proinflammatory effects (34–37).
Yamasaki et al (26) reported
that histamine levels had two peaks in nasal lavage fluid in a
guinea pig model of AR, at 20 min and 5 h after challenge, which
coincided with the peak time of frequent sneezing in the present
study. In a previous study, Wagenmann et al (6) demonstrated that histamine levels in the
nasal secretions of patients with AR were significantly increased
in the early and late phases subsequent to nasal allergen
provocation; however, the changes in histamine levels in the nasal
lavage fluid following challenge were not detected in the present
study. It is unknown whether histamine also exhibits a double peak
in the guinea pig model of AR; however, it was shown in the present
study that terfenadine, as an antihistamine drug, inhibited the
development of the biphasic sneezing in this model. This finding
strongly suggests that histamine contributes to the development of
the early and late phases of sneezing.
In the present study, it was found that the guinea
pigs in the model group produced significantly more sneezing in the
first peak between the 19th and 30th challenges than between the
first and 18th challenges (P<0.01). These results suggest that
biphasic responders showed significantly severer symptoms during
the early-phase reaction than single-phase responders, which is
similar to observations in human AR (3,38). The
mechanism of the above clinical phenomenon remains unclear. In a
previous study, Imai et al (39) demonstrated that chronic eosinophil
accumulation was induced by repeated antigen challenges in the
nasal tissue at the early phase. The eosinophils may have been
responsible for the amplification of the early-phase reaction, such
as vascular permeability and mucosal edema (39). Milanese et al (40) subsequently reported that the
eosinophil number was higher during the early phase in AR with LPR
than that in AR with only the early-phase reaction. From these
studies, it can be inferred that eosinophils may have an important
role in the early-phase reaction of allergic inflammation; however,
its participation in LPR has also been reported by numerous studies
(41,42).
In conclusion, this study has confirmed that SEB can
be used as an allergen to induce experimental AR with biphasic
sneezing by repeated intranasal instillation. The SEB-sensitized
guinea pigs are characterized by eosinophil infiltration in the
nasal mucosa and detectable levels of allergen-specific IgE and
IgG1 in the serum. The above indicators verify the
successful establishment of the AR animal model with biphasic
sneezing. Histamine may play an important role in the early- and
late-phase sneezing in the model of AR. This model closely reflects
AR and is a useful tool to study the association between SEB, AR
and the pathogenesis of the late phase of AR. Furthermore, this
model can be potentially used for the investigation of new
drugs.
Acknowledgements
This study was supported by the National Natural
Science Foundation Project (no. 81271061) and the National Key
Clinical Specialties Construction Program of China (no.
81271061).
References
|
1
|
Meltzer EO: The pharmacological basis for
the treatment of perennial allergic rhinitis and non-allergic
rhinitis with topical corticosteroids. Allergy. 52 (Suppl):33–40.
1997. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Niedoszytko M, Chełmińska M, Chełmiński K,
et al: Late-phase allergic reaction in nasal provocation with
fungal allergens. Allergy Asthma Proc. 29:35–39. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
de Graaf-in't Veld C, Garrelds IM, van
Toorenenbergen AW and Gerth van Wijk R: Nasal responsiveness to
allergen and histamine in patients with perennial rhinitis with and
without a late phase response. Thorax. 52:143–148. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Wang D and Clement P: Assessment of early-
and late-phase nasal obstruction in atopic patients after nasal
allergen challenge. Clin Otolaryngol Allied Sci. 20:368–373. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Jordan TR, Rasp G, Pfrogner E and Kramer
MF: An approach of immunoneurological aspects in nasal allergic
late phase. Allergy Asthma Proc. 26:382–390. 2005.PubMed/NCBI
|
|
6
|
Wagenmann M, Schumacher L and Bachert C:
The time course of the bilateral release of cytokines and mediators
after unilateral nasal allergen challenge. Allergy. 60:1132–1138.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Oldenbeuving NB, KleinJan A, Mulder PG, et
al: Evaluation of an intranasal house dust mite provocation model
as a tool in clinical research. Allergy. 60:751–759. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Bousquet J, Chanez P and Michel FB:
Pathophysiology and treatment of seasonal allergic rhinitis. Respir
Med. 84 (Suppl A):11–17. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Naclerio RM: Pathophysiology of perennial
allergic rhinitis. Allergy. 52 (Suppl):7–13. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Nabe T, Mizutani N, Shimizu K, et al:
Development of pollen-induced allergic rhinitis with early and late
phase nasal blockage in guinea pigs. Inflamm Res. 47:369–374. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhao Y, van Hasselt CA, Woo KS, et al:
Establishment of a modified intranasally ovalbumin induced animal
model of allergic rhinitis. Zhonghua Er Bi Yan Hou Tou Jing Wai Ke
Za Zhi. 40:176–180. 2005.(In Chinese). PubMed/NCBI
|
|
12
|
Tang X, Sun R, Hong S, et al: Repeated
intranasal instillation with staphylococcal enterotoxin B induces
nasal allergic inflammation in guinea pigs. Am J Rhinol Allergy.
25:176–181. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Breuer K, Haussler S, Kapp A and Werfel T:
Staphylococcus aureus: colonizing features and influence of
an antibacterial treatment in adults with atopic dermatitis. Br J
Dermatol. 147:55–61. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Chambers HF: The changing epidemiology of
Staphylococcus aureus? Emerg Infect Dis. 7:178–182. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Becker K, Friedrich AW, Lubritz G, et al:
Prevalence of genes encoding pyrogenic toxin superantigens and
exfoliative toxins among strains of Staphylococcus aureus
isolated from blood and nasal specimens. J Clin Microbiol.
41:1434–1439. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Sakairi T, Suzuki K, Makita S, et al:
Effects of fexofenadine hydrochloride in a guinea pig model of
antigen-induced rhinitis. Pharmacology. 75:76–86. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Avincsal MO, Ozbal S, Ikiz AO, Pekcetin C
and Güneri EA: Effects of topical intranasal doxycycline treatment
in the rat allergic rhinitis model. Clin Exp Otorhinolaryngol.
7:106–111. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Underwood DC, Osborn RR and Hand JM: Lack
of late-phase airway responses in conscious guinea pigs after a
variety of antigen challenges. Agents Actions. 37:191–194. 1992.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhao XJ: Experimental models of nasal
hypersensitive reaction. Zhonghua Er Bi Yan Hou Ke Za Zhi.
28:17-8–58-9. 1993.(In Chinese).
|
|
20
|
Okano M, Nishizaki K, Abe M, et al:
Strain-dependent induction of allergic rhinitis without adjuvant in
mice. Allergy. 54:593–601. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
van de Rijn M, Mehlhop PD, Judkins A, et
al: A murine model of allergic rhinitis: studies on the role of IgE
in pathogenesis and analysis of the eosinophil influx elicited by
allergen and eotaxin. J Allergy Clin Immunol. 102:65–74. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wang LF, Xu LJ, Guo FH, et al: Effect of
antiallergic herbal agents on chloride channel-3 and immune
microenvironment in nasal mucosal epithelia of allergic rhinitis
rabbits. Chin Med J (Engl). 123:1034–1038. 2010.PubMed/NCBI
|
|
23
|
Tsunematsu M, Yamaji T, Kozutsumi D, et
al: A new murine model of allergic rhinitis by repeated intranasal
Cry j 1 challenge. Biomed Res. 29:119–123. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhao Y, Woo JK, Leung PC, et al:
Symptomatic and pathophysiological observations in a modified
animal model of allergic rhinitis. Rhinology. 43:47–54.
2005.PubMed/NCBI
|
|
25
|
Lei F, Zhu D, Sun J and Dong Z: Effects of
minimal persistent inflammation on nasal mucosa of experimental
allergic rhinitis. Am J Rhinol Allergy. 24:e23–e28. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yamasaki M, Mizutani N, Sasaki K, et al:
Involvement of thromboxane A2 and peptide leukotrienes in early and
late phase nasal blockage in a guinea pig model of allergic
rhinitis. Inflamm Res. 50:466–473. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Brozmanova M, Bartos V, Plank L and Tatar
M: Experimental allergic rhinitis-related cough and airway
eosinophilia in sensitized guinea pigs. J Physiol Pharmacol. 58
(Suppl 5):57–65. 2007.PubMed/NCBI
|
|
28
|
Nabe T, Kubota K, Mizutani N, et al:
Effect of local nasal immunotherapy on nasal blockage in
pollen-induced allergic rhinitis of Guinea pigs. Allergol Int.
57:419–427. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Grønborg H, Bisgaard H, Rømeling F and
Mygind N: Early and late nasal symptom response to allergen
challenge. The effect of pretreatment with a glucocorticosteroid
spray. Allergy. 48:87–93. 1993. View Article : Google Scholar
|
|
30
|
Doyle WJ, Boehm S and Skoner DP:
Physiologic responses to intranasal dose-response challenges with
histamine, methacholine, bradykinin, and prostaglandin in adult
volunteers with and without nasal allergy. J Allergy Clin Immunol.
86:924–935. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Svensson C, Baumgarten CR, Pipkorn U, et
al: Reversibility and reproducibility of histamine induced plasma
leakage in nasal airways. Thorax. 44:13–18. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
White MV and Kaliner MA: Mediators of
allergic rhinitis. J Allergy Clin Immunol. 90:699–704. 1992.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Schleimer RP, Fox CC, Naclerio RM, et al:
Role of human basophils and mast cells in the pathogenesis of
allergic diseases. J Allergy Clin Immunol. 76:369–374. 1985.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Horak F: Clinical advantages of dual
activity in allergic rhinitis. Allergy. 55 (Suppl 64):34–39. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Montoro J, Sastre J, Jáuregui I, et al:
Allergic rhinitis: continuous or on demand antihistamine therapy? J
Investig Allergol Clin Immunol 17 Suppl. 2:21–27. 2007.
|
|
36
|
Pietrzkowicz M and Grzelewska-Rzymowska I:
Histamine as a mediator of allergic inflammation. Pol Merkur
Lekarski. 6:232–235. 1999.(In Polish). PubMed/NCBI
|
|
37
|
Pawankar R, Yamagishi S and Yagi T:
Revisiting the roles of mast cells in allergic rhinitis and its
relation to local IgE synthesis. Am J Rhinol. 14:309–317. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Terada N, Hamano N, Hohki G, et al: Late
phase response in nasal mucosa closely correlated with immediate
phase reaction and hyperreactivity to histamine. Acta Otolaryngol.
118:392–397. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Imai N, Miyahara A, Yamazaki Y, et al:
Involvement of eosinophils in the early-phase allergic reaction in
a guinea pig rhinitis model. Int Arch Allergy Immunol. 122:270–278.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Milanese M, Ricca V, Canonica GW and
Ciprandi G: Eosinophils, specific hyperreactivity and occurrence of
late phase reaction in allergic rhinitis. Eur Ann Allergy Clin
Immunol. 37:7–10. 2005.PubMed/NCBI
|
|
41
|
Elovsson S, Smailagic A, Erjefalt I, et
al: Evaluation of nasal barrier dysfunction at acute- and
late-phase reactions in a guinea pig model of allergic rhinitis.
Vascul Pharmacol. 43:267–276. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Sedgwick JB, Calhoun WJ, Gleich GJ, et al:
Immediate and late airway response of allergic rhinitis patients to
segmental antigen challenge. Characterization of eosinophil and
mast cell mediators. Am Rev Respir Dis. 144:1274–1281. 1991.
View Article : Google Scholar : PubMed/NCBI
|