Introduction
Previous clinical studies have found that the
primary cause of femoral head necrosis is steroid hormones, which
is the main reason for the disease (1). The occurrence of femoral head necrosis
is associated with the administration method and dose of the
hormones, and is affected by individual differences and
sensitivities (2). It is not
possible to make an early diagnosis for the majority of patients
with certainty, and effective procedures for prevention and
treatment are lacking as the pathogenesis remains unknown. During
the previous decade, the apoptosis theory has received increased
research attention along with the development of molecular biology;
apoptosis of osteocytes and osteoblasts is considered to play an
important role in steroid-induced avascular necrosis of the femoral
head (SANFH) (3). SANFH and the
apoptosis of osteocytes and osteoblasts are considered to be
closely associated (4). Chan and Mok
(5) found that the excessive use of
hormones influenced the formation mechanisms of osteoblasts and
osteoclasts in bone marrow, and decreased the rate of skeleton
formation and bone mineral density (BMD), in addition to inducing
the apoptosis of mature osteoblasts and osteoclasts. The authors
hypothesized that the primary effects of the hormones included
inducing the apoptosis of mature osteoblasts and osteoclasts,
inhibiting osteoclastogenesis and reducing the bone turnover rate,
leading to the development of apoptosis and resulting in femoral
head necrosis. The majority of the target molecules of caspase-3,
which regulates the differentiation of numerous cells, including
skeletal muscle cells, neurocytes and lymphocytes, have been
identified. The target molecules include certain cytoskeletal
proteins, cyclins, kinases and transcription regulators (6). Previous studies have demonstrated that
the expression of caspase-3 is upregulated, along with an increase
in apoptosis rate, in various types of cells, and the association
between caspase-3 and apoptosis can also be observed in osteocytes,
osteoblasts and chondrocytes (7–9).
Therefore, cell apoptosis and the apoptotic pathway in SANFH are
the primary focus of studies concerning the pathogenesis of the
disease.
In the present study, a rabbit model of SANFH was
established and blood samples were collected for centrifugation to
detect changes in nitric oxide (NO) content in serum; the
osteoblast apoptosis index was also detected in the bilateral
femoral head. The aim of the study was to investigate the
association between SANFH and NO content variation, as well as the
association between this variation and osteocyte apoptosis.
Materials and methods
Experimental animal
A total of 40 healthy adult Japanese white rabbits
(male and female), 5 months old and weighing 2.5±0.5 kg, were
provided by the Beijing Fuhao Animal Breeding Center (Beijing,
China) with a certificate issued by the National Institute of
Medical Experiments (no. 20090132). The rabbits were fed with a
standard diet and housed individually in cages. The Ethics
Committee of The First Affiliated Hospital of Chongqing Medical
University (Chongqing, China) approved of this study.
Experimental instruments
A H-700H electron microscope (Hitachi, Tokyo,
Japan), LKB 2800 ultramicrotome (LKB, Bromma, Sweden), T22N visible
spectrophotometer (Shanghai Qinghua Scientific Education Equipment
Co., Ltd., Shanghai, China), paraffin embedding machine (EG1150 C;
Leica Biosystems Co. Ltd., Shanghai, China), an ultra-thin slicing
machine (Sweden LKB2800; Leica Biosystems), JD-801 medical image
analysis system (Jiangsu Xunjie Co. Ltd., Jiangsu, China),
Millipore RU water purification system (EMD Millipore, Billerica,
MA, USA), and a CX41 microscope (Olympus Corporation, Tokyo, Japan)
were used in the study. The above instruments were provided by the
Molecular Biology Research Center and Electron Microscopy Center of
Inner Mongolia Medical University (Hohhot, China).
Reagents
Escherichia coli (E. coli) endotoxin
(provided by the National Institute for the Control of
Pharmaceutical and Biological Products, Beijing, China),
methylprednisolone (Pharmacia & Upjohn Co., Brussels, Belgium),
NO detection kit using a nitrate reductase assay (Beijing 4A
Biotech Co., Ltd., Beijing, China) and the terminal
deoxynucleotidyl transferase dUTP nick end labeling (TUNEL) assay
apoptosis detection kit (Wuhan Boster Company, Wuhan, China) were
used. Other conventional reagents, including hematoxylin and eosin
(H&E) staining and ethylenediaminetetraacetic acid (EDTA)
decalcification solutions, were provided by the Molecular Biology
Research Center of the Inner Mongolia Medical University.
Grouping and establishment of the
experimental animal model
Experimental animals were divided into 4 groups by a
random number table method. Group A, with 10 rabbits, was the model
group; rabbits were intravenously injected with the E. coli
endotoxin, twice at 24 h intervals with 100 µg/kg each time.
Following the second injection of the E. coli endotoxin, the
rabbits were intramuscularly injected with methylprednisolone, 3
times at 24 h intervals with 20 mg/kg each time. Group B, with 10
rabbits, was the endotoxin group; rabbits were intravenously
injected with E. coli endotoxin, twice at 24 h intervals
with 100 µg/kg each time. Following the second injection of E.
coli endotoxin, rabbits were intramuscularly injected with
normal saline, 3 times at 24 h intervals with 20 mg/kg each time.
Group C, with 10 rabbits, was the hormone group; rabbits were
intravenously injected with normal saline, twice at 24 h intervals
with 100 µg/kg each time. Following the second injection of normal
saline, the rabbits were intramuscularly injected with
methylprednisolone, 3 times at 24 h intervals with 20 mg/kg each
time. Group D, with 10 rabbits, was the control group; rabbits were
intravenously injected with normal saline, twice at 24 h intervals
with 100 µg/kg each time. Following the second injection of normal
saline, the rabbits were intramuscularly injected with normal
saline, 3 times at 24 h intervals with 20 mg/kg each time. Four
weeks after the final injection, a 5 ml blood sample was taken from
the heart of each animal and the animals were subsequently
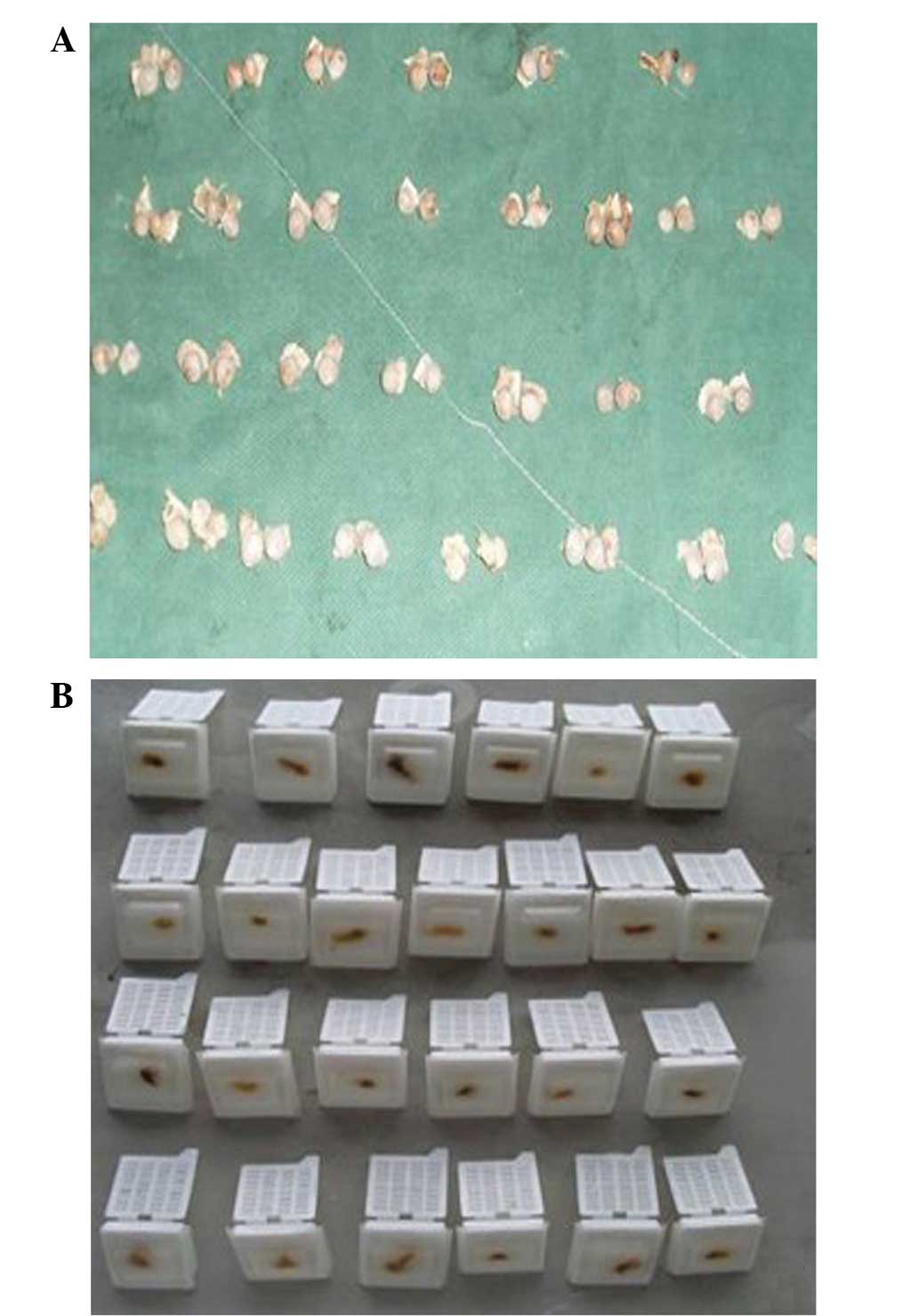
sacrificed by air embolism. The bilateral femoral head samples were
collected under sterile conditions (Fig.
1A).
Specimen preparation
Blood samples were centrifuged for 10 min at 1,989 ×
g at 20°C and the upper serum was stored at −20°C for testing. One
femoral head was fixed in 5% glutaraldehyde solution and the other
in 10% formaldehyde solution, for one week. Bone tissue was
decalcified for 40 days using 15% EDTA and embedded in paraffin
(Fig. 1B).
Histomorphological observation under
an optical microscope
The paraffin-embedded tissue blocks were cut into
slices with a thickness of 4 µm and stained with H&E. The
histopathological changes were observed under an optical microscope
at ×100 and ×400 magnifications. The histopathological changes were
classified according to the Matsui method (10) as follows: Grade 0, intact trabecular
bone and bone marrow without pathological changes; grade 1, simple
bone marrow necrosis without trabecular bone necrosis, manifested
as cell lysis, nuclear fragmentation, karyolysis in bone marrow
cells and hypochromatosis, blurred nuclear boundaries as well as
fused boundaries in fat cells; and grade 2, trabecular necrosis and
bone marrow necrosis; in addition to the pathological changes of
grade 1, there are also empty lacunae accompanied by newly formed
adherent bone. The degree of pathological change associated with
bone cell necrosis was indicated by the percentage of empty bone
lacunae. At ×100 magnification, 5 fields were randomly selected to
determine the empty bone lacunae and calculate the percentage of
empty bone lacunae. The ratio of empty lacunae in the subchondral
femoral region has been widely accepted as a histological indicator
of SANFH. The normal proportion of empty lacunae in the subchondral
femoral region of an adult rabbit is 8–12% (11). Therefore, with the same sample
preparation procedure, an empty lacunae ratio in the subchondral
femoral region greater than that number indicates the presence of
osteonecrosis.
Observation under a transmission
electron microscope
The femoral heads were fixed with 5% glutaraldehyde
and decalcified. The bone mass 0.2–0.3 cm under the femoral head
cartilage was cut into a 1-mm3 section, decalcified with
5% nitric acid for 2–3 h, fixed in 1% osmium tetroxide for 1 h,
dehydrated in graded alcohol solutions and embedded using epoxy
resin (Epon 812; Beijing Dalike Technology Co. Ltd., Beijing,
China). The anhydrous ethanol was gradually replaced by mixing the
embedding medium with anhydrous ethanol at a ratio of 1:1, 2:1 and
3:1. Subsequently, the sample was saturated with embedding medium
by incubation in a 37°C thermostat incubator (DC-0506; FDL
Technology Information Co. Ltd., Nanjing, China) for 24 h,
aggregated at 60°C in an oven for 48 h, cut into 50-nm ultrathin
slices using an ultramicrotome and stained (double staining with
uranyl acetate and lead citrate). The ultrathin slices were
observed under the H-700H electron microscope.
Determination of NO content in the
rabbit serum by a nitrate reductase assay
NO is chemically active and reacts with molecular
oxygen to generate NO2. This transforms into
NO3− and NO2−, and 95%
NO2− transforms into
NO3− within 1 h. Thus, the determination of
the serum level of NO2− is inaccurate for
determining the level of NO. However, the sum of serum
NO2− and NO3− levels is
an accurate indicator of NO level. Thus, in the present assay,
NO3− was specifically reduced to
NO2− by nitrate reductase and the NO content
was indicated by color intensity; that is, the optical density (OD)
value indicated the NO level. The reagents used for the test were
as follows: Reagent 1, 6.0 ml phosphate-buffered saline (PBS);
reagent 2, 6.0 ml nitrate reductase; reagent 3, 30 ml; reagent 4,
30 ml; reagent 5, 1 ml 100 µmol/ml potassium nitrate solution. The
use of the reagents in the process is detailed in Table I. The mixed solution was maintained
at room temperature for 10 min. A 0.5-cm cuvette was used; the
blank solution was set as the zero point and the OD values of the
measuring and standard solutions were calculated. The OD values
were calculated as follows: Nitric oxide (µmol/l) = measuring
solution OD/standard solution OD ×100.
 | Table I.Determination of nitric oxide (NO)
content in rabbit serum by the nitrate reductase method. |
Table I.
Determination of nitric oxide (NO)
content in rabbit serum by the nitrate reductase method.
| Material | Measuring tube
(ml) | Standard tube
(ml) | Blank tube (ml) |
|---|
| Sample | 0.1 | - | - |
| Potassium
nitrate | - | 0.1 | - |
| Double distilled
water | - | - | 0.1 |
| Reagent 1 | 0.1 | 0.1 | 0.1 |
| Reagent 2 | 0.1 | 0.1 | 0.1 |
| Reagent
3a | 0.5 | 0.5 | 0.5 |
| Reagent 4 | 0.5 | 0.5 | 0.5 |
Detection of osteoblast apoptosis by
TUNEL assay
The assay was performed according to kit
instructions (TUNEL Apoptosis Assay kit; Wuhan Boster Company). The
prepared paraffinized tissue slices were placed in a dyeing tank,
washed twice with dimethyl benzene and anhydrous ethanol and once
with 95 and 75% ethanol, respectively. The slices were subsquently
washed for 5 min with PBS. Proteinase K solution (20 µg/ml) was
added and the slices were incubated for 15 min at room temperature.
The hydrolyzed tissue proteins were removed and the slices were
washed 4 times with distilled water. PBS containing 2% hydrogen
peroxide was added, left for 5 min and the slices were washed twice
with PBS. Residual liquid was removed using filter paper and 2
drops terminal deoxynucleotidyl transferase (TdT) were immediately
added to each slice. The residual liquid was again removed using
filter paper and 54 µl TdT enzyme buffer was then titrated onto
each slice for 1 h. Washing and reaction terminating solutions
preheated to 37°C were added and the slices were maintained at 37°C
for 30 min. The slices were washed 3 times with PBS, and 2 drops
peroxidase-labeled anti-digoxin antibody (1:16) were added for 30
min. After washing 4 times with PBS, freshly prepared 0.05%
3,3′-diaminobenzidine was added to each slice, left for 5 min and
the slices were washed 4 times using distilled water. The slices
were counterstained with methyl green at room temperature for 10
min. They were subsequently washed 3 times with distilled water and
then 3 times with 100% butanol. The slices were dehydrated 3 times
using dimethylbenzene, mounted, dried and observed under an optical
microscope. TUNEL-positive staining, which appeared tan or brown,
was identified in the nucleus with a few positive granules located
in the cytoplasm. Five visual fields were randomly selected at
high-magnification for each slice, 50 osteocytes were counted in
each field and the apoptotic index was calculated (apoptotic cell
number/total cell number in the fields).
Statistical analysis
All data are presented as mean ± standard deviation.
Data were statistically analyzed by analysis of variance (ANOVA)
using SPSS 13.0 software (SPSS, Inc., Chicago, IL, USA). Pairwise
comparisons of multiple samples were carried out with Fisher's
least significant difference test. The linear correlation between
NO content and the apoptosis ratio in group A was analyzed. −1≤r≤+1
indicated that the two variables linearly correlated with one
other; positive and negative values of r indicate a positive and
negative correlation, respectively. P<0.05 was considered to
indicate a statistically significant difference.
Results
Histomorphological observation
No significant difference in the general morphology
of the femoral heads in the 4 groups was identified by observation
with the naked eye. One femoral head was partially separated from
the femoral neck in group A.
Changes observed under the optical
microscope
Typical osteonecrosis was observed in the femoral
head slices of group A; it was mainly located under the cartilage.
Bone trabeculae became scattered and thin, or even fractured, with
structural disorder and fragmentation. The number of hematopoietic
cells was reduced. Furthermore, the osteoblasts were arranged
irregularly and the number of empty bone lacunae increased
significantly. The volume of fat cells in the bone marrow
increased, with certain fat cells becoming fused into vacuoles
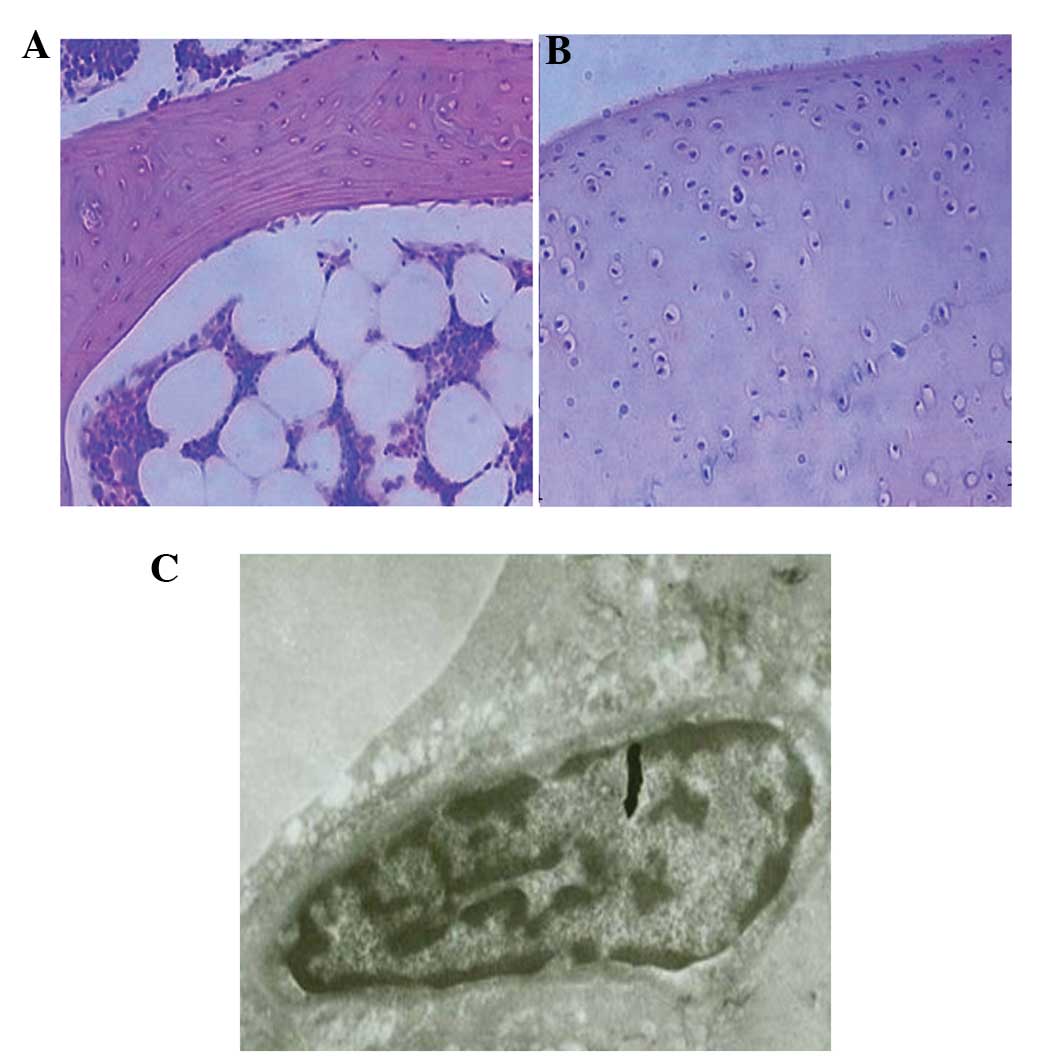
(Fig. 2A and B). In the femoral head
slices in group B, the bone trabeculae in the femoral head biopsy
were intact, regularly arranged and compacted. Bone cells with
centrally located large nuclei were visible and there were numerous
bone marrow hematopoietic cells. There were relatively few fat
cells with normal morphology. In the group C slices, there was no
visible osteonecrosis; bone trabeculae became scattered and thin,
with mild structural disorders. There was an increase in bone
marrow cell lysis and the number and volume of bone marrow fat
cells. In the femoral head slices of group D, the bone trabeculae
were intact, regularly arranged and compacted. Bone cells with
centrally located large nuclei were visible; there were numerous
bone marrow hematopoietic cells and relatively normal fat cells
(Fig. 3A and B). The classification
of the histopathological changes in the bone samples by the Matsui
method is detailed in Table II.
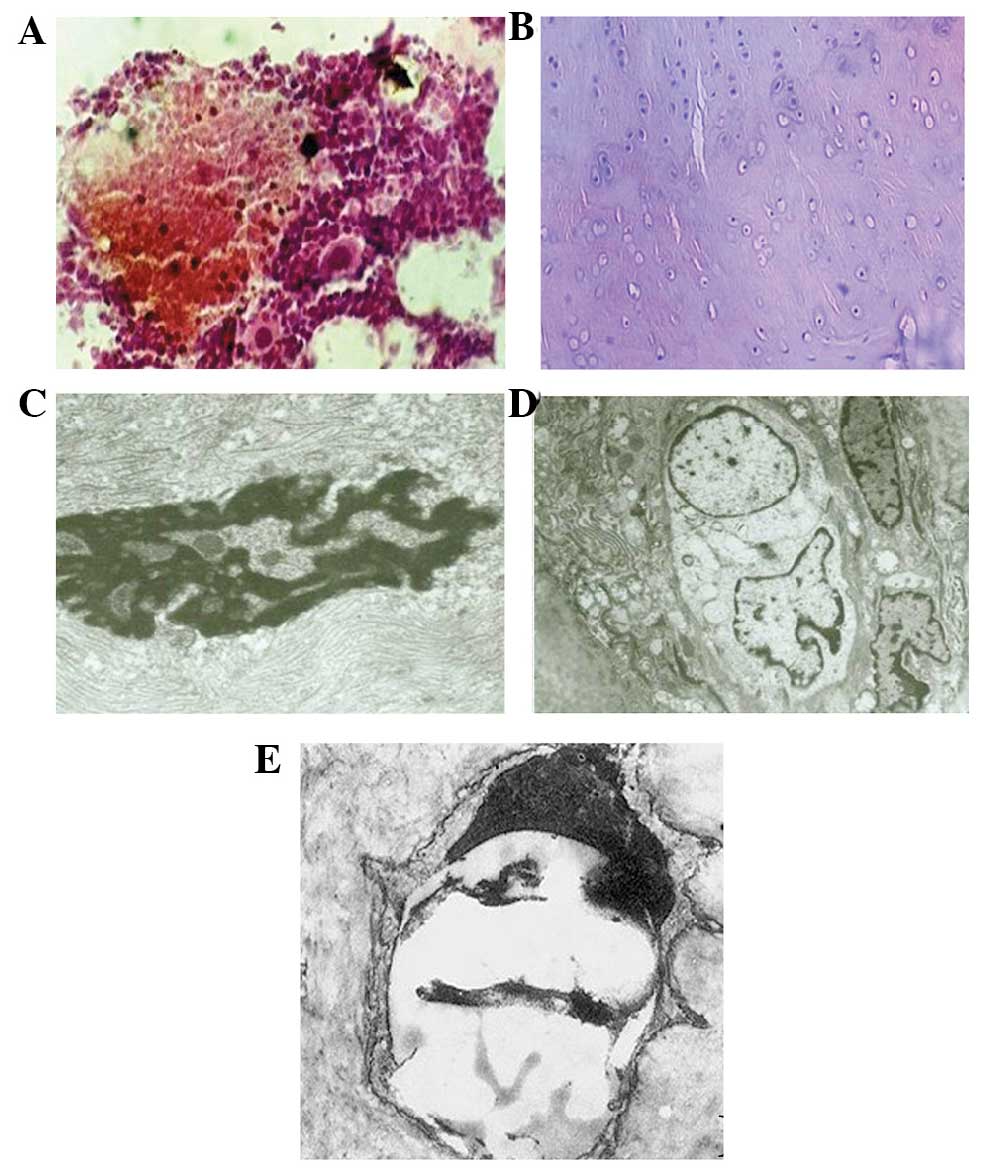 | Figure 2.Bone structure of the rabbits in the
hormone and endotoxin group. (A) The intramedullary structure was
disordered with bone marrow hemorrhagic necrosis (H&E staining;
magnification, ×6,000). (B) Bone cell necrosis resulted in empty
lacunae (H&E staining; magnification, ×100). (C) Bone cell
disrupted by necrosis. The nuclear membrane disappeared, chromatin
was irregular and there were no organelles present (magnification,
×10,000). (D) Bone cell necrosis was demonstrated by large vacuoles
in the cytoplasm. (E) Apoptotic bone cell (TUNEL staining;
magnification, ×10,000). H&E, hematoxylin and eosin; TUNEL,
terminal deoxynucleotidyl transferase dUTO nick end labeling
assay |
| Table II.Classification of the
histopathological changes in the bone samples by the Matsui
method. |
Table II.
Classification of the
histopathological changes in the bone samples by the Matsui
method.
|
|
No. of the
experimental animal |
|---|
|
|
|
|---|
| Group | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 |
|---|
| A | 2 | 1 | 2 | 2 | 1 | 1 | 1 | 0 | 2 | 1 |
| B | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| C | 1 | 0 | 0 | 1 | 0 | 0 | 1 | 0 | 0 | 0 |
| D | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
The empty bone lacuna rate in group A was
significantly higher compared with that in groups B, C and D
(P<0.01); there was no significant difference in the rate among
groups B, C and D. There were numerous empty bone lacunae in group
A and the empty bone lacunae rate was 29.10±3.67%. In groups B, C
and D, the osteocytes in the rabbit femoral bone marrow were
stained homogeneously with normal morphology and were moderately
distributed; the empty bone lacunae rates in these three groups
were 9.10±2.60, 9.50±2.01 and 8.01±2.36%, respectively (F=137.88;
P<0.001).
Changes observed under the electron
microscope
In the femoral head slices in group A, the
osteocytes were disintegrated and necrotic, the nuclei were
fragmented and the nuclear membrane was discontinuous or had
completely disappeared. The majority of the organelles had
partially or completely disappeared. Apoptotic bodies could be
observed and there were large numbers of lysosomes and secretory
granules in the cytoplasm. Bleeding was observed within the bone
marrow (Fig. 2C and D).
The ultrastructure of osteocytes and bone marrow
cells were normal in groups B and D (Fig. 3C).
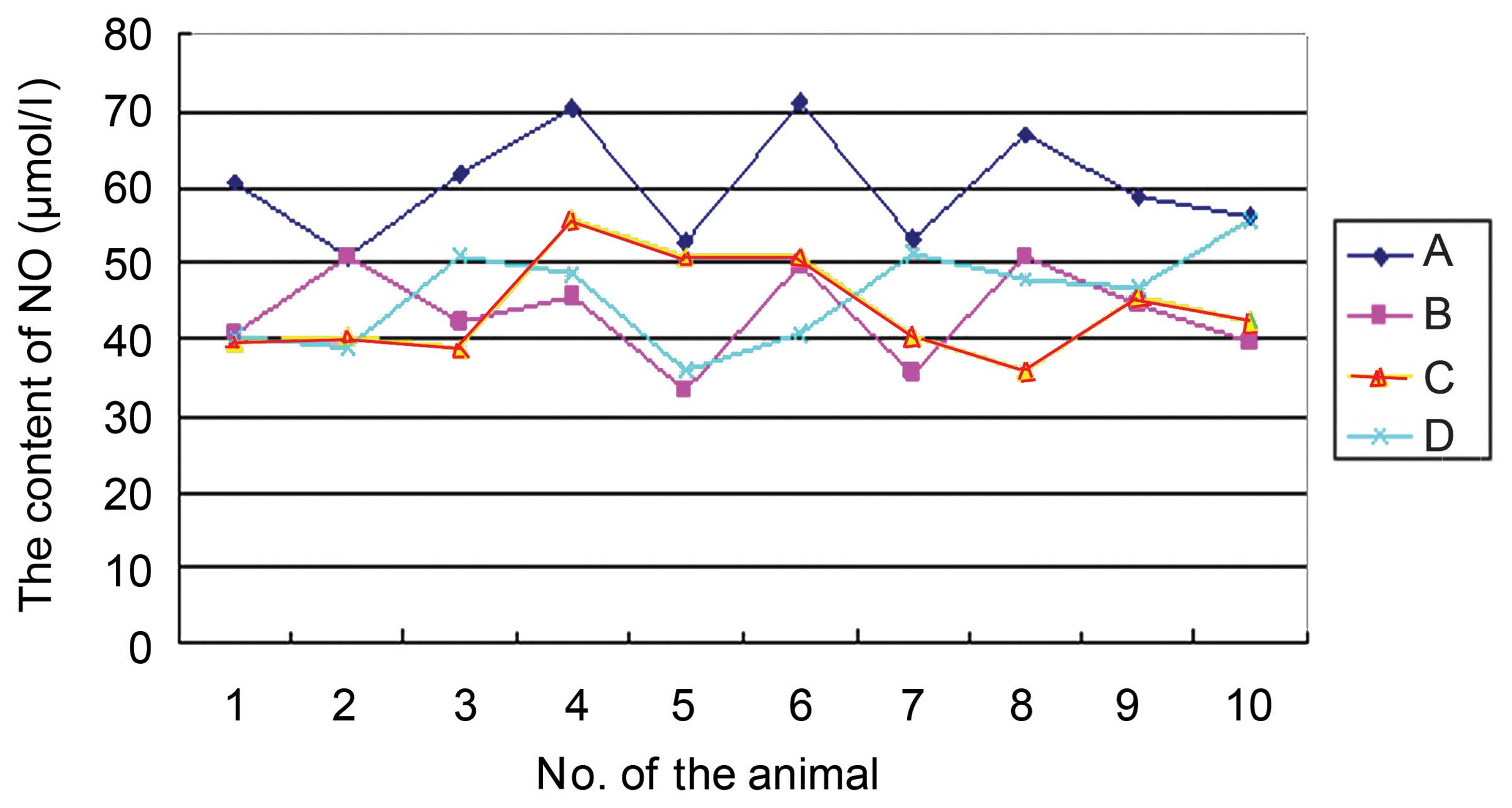
Changes in NO content
The NO content in group A was significantly higher
compared with that in groups B, C and D (P<0.01); there was no
significant difference in NO content among groups B, C and D. The
NO content was 60.19±7.26, 43.25±6.11, 44.04±6.44 and 45.75±6.47
µmol/l in groups A, B, C and D, respectively (Fig. 4; F=14.73; P<0.001).
Osteocyte apoptosis observed with
TUNEL staining
TUNEL-positive staining, which appeared tan or
brown, was located in the nucleus with a few positive granules
located in the cytoplasm. Five randomly-selected visual fields were
observed at high-magnification for each slice and 50 osteocytes in
each field were counted to calculate the apoptotic index. There
were few apoptotic osteocytes in the bone samples from groups B, C
and D. However, there were positive cells with visible brown
particles in the nuclei of the bone samples from group A and
numerous apoptotic osteocytes in the bone marrow tissue (Fig. 3E). The osteocyte apoptosis index in
group A was significantly higher compared with that in groups B, C
and D (P<0.01); there was no significant difference in the
osteocyte apoptosis index among groups B, C and D. The osteocyte
apoptosis indices were 30.80±5.14, 6.90±1.92, 6.60±1.27 and
6.10±1.95 in groups A, B, C and D, respectively (F=14.73;
P=<0.001).
Linear association between NO content
and osteocyte apoptosis index
The apoptosis index in group A was r=0.707 which
meant that the NO content and osteocyte apoptosis index positively
correlated with one other. The NO content and osteocyte apoptosis
index in each animal of group A are detailed in Table III.
| Table III.Nitric oxide (NO) content and
osteocyte apoptosis index in the animals of group A. |
Table III.
Nitric oxide (NO) content and
osteocyte apoptosis index in the animals of group A.
|
|
No. of the
experimental animal |
|---|
|
|
|
|---|
| Variable | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 |
|---|
| NO content
(µmol/l) | 60.57 | 50. 68 | 61.72 | 70.34 | 52.77 | 70.90 | 53.24 | 66.75 | 58.77 | 56.25 |
| Apoptotic index
(%) | 27 | 29 | 30 | 35 | 32 | 34 | 26 | 33 | 30 | 30 |
Discussion
SANFH, which is induced by the long-term or
extensive short-term use of adrenocorticotropic hormone, has
received much attention in the medical field. The number of
clinical cases has been increasing since it was first reported by
Pietrograndi in 1957 (12). SANFH,
which is mainly bilateral and involves extensive necrosis,
frequently occurs in young and middle-aged individuals. This
disease is more serious than idiopathic avascular necrosis of the
femoral head, with a higher disability rate (13). A large number of cases have emerged
since ‘atypical pneumonia’ in 2010 (14). Investigating the etiology of SANFH
remains a research hotspot in the field (15).
Steroid hormones have been widely accepted as an
important cause of SANFH and numerous theories have been proposed
concerning the pathogenesis, including fat embolism, high
intraosseous pressure, enlarged fat cells, lipid droplet
aggregation in osteocytes, osteoporosis and cumulative osteocyte
dysfunction (16). However, the
exact mechanism remains incompletely understood. Furthermore, the
above theories do not clearly explain the occurrence of osteocyte
necrosis as no cell swelling and inflammation accompany the
characteristic necrosis of SANFH. Steroidal hormones, such as
glucocorticoids, have been demonstrated to induce apoptosis
(17). They reduce the number of
osteocytes by promoting the apoptosis of osteoblasts and
osteocytes, and they also affect the function of osteoblasts,
retard osteogenesis and induce cell apoptosis, which results in
bone loss. The present study further demonstrated the role of cell
apoptosis in SANFH.
NO, an important bioactive substance, plays an
important role in the regulation of normal body function and
disease occurrence. Its major physiological functions include
dilating vascular smooth muscle, and inhibiting the aggregation of
platelets, platelet and leukocyte adhesion and hyperplasia of the
vascular smooth muscle (18,19). Hormones can damage vascular
endothelial cells in which NO is produced and inhibit the activity
of NO synthase (NOS). The appropriate concentration of NO maintains
normal vasomotor center function to achieve the effective perfusion
of organs. Vascular spasms, hyperplasia of the vascular wall,
stenosis, platelet aggregation, leukocyte adhesion and thrombosis
occur if NO content decreases. Avascular necrosis of femoral head
is induced if these pathological changes occur in the femoral head
and peripheral vasculature. NO is considered to be an important
regulator of osteocyte apoptosis. A certain amount of NO has been
revealed to bond with superoxide anions to reduce the level of
oxygen free radicals and prevent their injuring effects on the
microvasculature and synovium (20).
Another study found that the content of inducible and endothelial
NOS in osteoblasts, osteoclasts and bone marrow cells increased
significantly in patients with osteonecrosis, while it was
extremely low in patients with osteoarthritis (21). It has been suggested that NO may
damage bone tissue in SANFH through the following mechanisms
(22). Firstly, a high concentration
of NO inhibits a variety of enzymes associated with the
mitochondrial electron transport system and the citric acid cycle,
including citrate synthase and aconitase. NO associates with the
iron-sulfur centers of these enzymes to reduce the cellular iron
level and thus inhibit the mitochondrial electron transport chain
to cause cellular damage. Secondly, NO reacts with the superoxide
anion O2•− and generates the peroxynitrite
anion ONOO−, which is relatively stable under alkaline
conditions; however, under acidic conditions it rapidly decomposes
into the free radicals OH• and
NO2• with strong toxicity. These two free
radicals have very strong oxidizing activity and cytotoxicity,
resulting in bone damage. NO has a dual function in bone absorption
and inhibits the growth of osteoblasts or even has toxic effects
during bone formation (1). However,
the effect of NO on the process of SANFH development is unclear.
The opinions of researchers concerning how NO content changes
during SANFH are not consistent. Certain studies have shown that
that the content increases while others have demonstrated the
opposite (21). The dispute may be
due to the different time-points at which the levels are detected.
In the present study, NO content increased during the early period
(week 4) of the SANFH model, which corresponded with osteocyte
apoptosis.
Lee et al (23) studied the bone specimens of patients
with nontraumatic osteonecrosis by immunohistochemistry and found
that the NOS content was increased. The authors hypothesized that
hormones or alcohol had direct cytotoxic effects on osteocytes,
resulting in an increase in the NO content that mediated apoptosis.
Thus, they concluded that osteocyte apoptosis played an important
role in the pathogenesis of femoral head osteonecrosis. The results
of the present study indicated that NO is associated with cell
apoptosis and acts as an important apoptotic modulator; the exact
mechanism for this remains to be further investigated.
In conclusion, in the present study, the
precoagulation status was simulated by intravenously injecting
E. coli endotoxin into rabbits and applying high-dose
hormone to establish animal models of SANFH. Morphological and
ultrastructural characteristics of the femoral head were observed
and a comprehensive analysis was carried out by an
immunohistochemical method. The results showed that NO content and
the osteocyte apoptosis index were increased during the SANFH
process, and indicated that increases in NO content and the
osteocyte apoptosis index play important roles in SANFH.
Furthermore, NO content was found to be positively correlated with
the osteocyte apoptosis index, indicating that NO is able to induce
cell apoptosis. Future studies are required to investigate the
exact pathway for this and whether NO has a dual function.
References
|
1
|
Chen S, Li J, Peng H, Zhou J and Fang H:
Administration of erythropoietin exerts protective effects against
glucocorticoid-induced osteonecrosis of the femoral head in rats.
Int J Mol Med. 33:840–848. 2014.PubMed/NCBI
|
|
2
|
Saito M, Ueshima K, Fujioka M, et al:
Corticosteroid administration within 2 weeks after renal
transplantation affects the incidence of femoral head
osteonecrosis. Acta Orthop. 85:266–270. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Nishida K, Yamamoto T, Motomura G,
Jingushi S and Iwamoto Y: Pitavastatin may reduce risk of
steroid-induced osteonecrosis in rabbits: A preliminary
histological study. Clin Orthop Relat Res. 466:1054–1058. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Takano-Murakami R, Tokunaga K, Kondo N,
Ito T, et al: Glucocoriticoid inhibits bone regeneration after
osteonecrosis of the femoral head in aged female rats. Tohoku J Exp
Med. 217:51–58. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Chan KL and Mok CC: Glucocorticoid-induced
avascular bone necrosis: Diagnosis and management. Open Orthop J.
6:449–457. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Chen C, Yang S, Feng Y, et al: Impairment
of two types of circulating endothelial progenitor cells in
patients with glucocorticoid-induced avascular osteonecrosis of the
femoral head. Joint Bone Spine. 80:70–76. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Tan G, Kang PD and Pei FX: Glucocorticoids
affect the metabolism of bone marrow stromal cells and lead to
osteonecrosis of the femoral head: A review. Chin Med J (Engl).
125:134–139. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zou W, Yang S, Zhang T, et al: Hypoxia
enhances glucocorticoid-induced apoptosis and cell cycle arrest via
the PI3K/Akt signaling pathway in osteoblastic cells. J Bone Miner
Metab Sep. 18:2014.(Epub ahead of print).
|
|
9
|
Ye J, Wu G, Li X, et al: Millimeter wave
treatment inhibits apoptosis of chondrocytes via regulation dynamic
equilibrium of intracellular free Ca2+. Evid Based
Complement Alternat Med. 2015:4641612015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ohzono K, Takaoka K, Saito S, Saito M,
Matsui M and Ono K: Intraosseous arterial architecture in
nontraumatic avascular necrosis of the femoral head.
Microangiographic and histologic study. Clin Orthop Relat Res.
277:79–88. 1992.PubMed/NCBI
|
|
11
|
Eberhardt AW, Yeager-Jones A and Blair HC:
Regional trabecular bone matrix degeneration and osteocyte death in
femora of glucocorticoid-treated rabbits. Endocrinology.
142:1333–1340. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Pietrograndi V and Mastromarino R:
Osteopatia da prolungata trattamento cortisonico. Ortop Traum Appar
Mar. 25:791–810. 1957.(In Italian).
|
|
13
|
Fukushima W, Fujioka M, Kubo T, et al:
Nationwide epidemiologic survey of idiopathic osteonecrosis of the
femoral head. Clin Orthop Relat Res. 468:2715–2724. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Qiu NH and Zhang W: Infectious atypical
pneumonia after the etiology and treatment of femoral head
necrosis. Chinese J Tissue Eng Res. 17:5525–5530. 2013.
|
|
15
|
Weinstein RS: Glucocorticoid-induced
osteonecrosis. Endocrine. 41:183–190. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kerachian MA, Cournoyer D, Harvey EJ, et
al: New insights into the pathogenesis of glucocorticoid-induced
avascular necrosis: Microarray analysis of gene expression in a rat
model. Arthritis Res Ther. 12:R1242010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kameda H, Amano K, Nagasawa H, et al:
Notable difference between the development of vertebral fracture
and osteonecrosis of the femoral head in patients treated with
high-dose glucocorticoids for systemic rheumatic diseases. Intern
Med. 48:1931–1938. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Chen RM, Liu HC, Lin YL, Jean WC, Chen JS
and Wang JH: Nitric oxide induces osteoblast apoptosis through the
de novo synthesis of Bax protein. J Orthop Res. 20:295–302. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Buerk DG, Barbee KA and Jaron D: Nitric
oxide signaling in the microcirculation. Crit Rev Biomed Eng.
39:397–433. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Bao F, Wu P, Xiao N, Qiu F and Zeng QP:
Nitric oxide-driven hypoxia initiates synovial angiogenesis,
hyperplasia and inflammatory lesions in mice. PLoS One.
7:e344942012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Calder JD, Buttery L, Revell PA, et al:
Apoptosis - a significant cause of bone cell death in osteonecrosis
of femoral head. J Bone Joint Surg Br. 86:1209–1213. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Hu M, Zhao H, Dong X, Luo D and Zhou X:
General and light microscope observation on histological changes of
femoral heads between SANFH rabbit animal models and it were
intervened by Osteoking Zhongguo Zhong Yao Za Zhi. 35:2912–2916.
2010.(In Chinese). PubMed/NCBI
|
|
23
|
Lee JS, Lee JS, Roh HL, Kim CH, Jung JS
and Suh KT: Alterations in the differentiation ability of
mesenchymal stem cells in patients with nontraumatic osteonecrosis
of the femoral head: Comparative analysis according to the risk
factor. J Orthop Res. 24:604–609. 2006. View Article : Google Scholar : PubMed/NCBI
|