Introduction
Diabetes mellitus is a serious social health
problem, with diabetes-related vascular complications representing
a major public health burden. According to statistics, >40% of
patients hospitalized with acute myocardial infarction have
clinical diabetes, and another 35% exhibit impaired glucose
tolerance (1). Excessive production
of reactive oxygen species (ROS) and the subsequent increase in
oxidative stress under high-glucose conditions play a critical role
in this pathology (2). The
overproduction of ROS, including superoxide anion radicals,
hydrogen peroxide and hydroxyl radicals, can prominently damage
nucleic acids, lipids and proteins and result in cellular necrosis,
tissue structural damage and functional disorder (3). In the setting of hypoxia,
hypoxia-inducible factor-1α (HIF-1α) increases in response to
ischemia. HIF-1α is a key regulator of oxygen homeostasis, which
restores blood flow to ischemic regions (4); however, sustained and prolonged
activation of the HIF-1α pathway induces cell death due to the
subsequent activation of p53 and other associated genes (5).
Among the currently available medications for
diabetes, natural products have drawn increasing attention. As a
traditional Chinese herbal medicine, garlic has been used to treat
a range of diseases for thousands of years. Allicin is one of the
main active components in garlic and exerts several therapeutic
effects, such as promoting insulin sensitivity (6,7),
decreasing blood glucose levels (8,9),
regulating lipid metabolism (10,11),
reducing homocysteine levels (12),
attenuating superoxide production (13) and limiting inflammation and
fibrogenesis. Given the role of oxidative stress within the
etiology and pathogenesis of diabetic complications, the present
study utilized an in vitro model of diabetes-associated
oxidative stress using aortic endothelial cells cultured under
high-glucose/hypoxic conditions to clarify the possible mechanism
of allicin on diabetic macrovascular complications.
Materials and methods
Materials
The experiment was carried out in the Institute of
Basic Medical Sciences, Qilu Hospital of Shandong University
(Jinan, China) between February and September, 2014. The murine
aortic endothelial cells were obtained from CHI Scientific, Inc.
(Jiangyin, China). Xanthine (X), xanthine oxidase (XO), MTT,
trypsin, and GF109203x [an inhibitor of protein kinase C (PKC)]
were purchased from Sigma-Aldrich (St. Louis, MO, USA). Allicin was
provided by Xuzhou Laien Pharmaceutical Co. Ltd. (Xuzhou, China).
Endothelial cell medium (ECM) was provided by Sciencell Research
Laboratories (Carlsbad, CA, USA) and TRIzol® was obtained from
Gibco-BRL (Grand Island, NY, USA). Culture flasks and 6-, 12-, 24-
and 96-well culture plates were purchased from Sigma-Aldrich.
8-Hydroxydeoxyguanosine (8-OHdG), nuclear factor-κB (NF-κB), NADPH
oxidase 4 (Nox4), HIF-1α and PKC test kits were obtained from
Nanjing Jiancheng Bioengineering Institute (Nanjing, China). Taq
DNA polymerase and oligo (dt) were provided by Fermentas (Thermo
Fisher Scientific, Pittsburgh, PA, USA). The primers were
synthesized by Sangon Biotech, Co., Ltd. (Shanghai, China), and all
other reagents were analytically pure.
Cell cultures
Murine aortic endothelial cells of the third passage
were used in this study. Cells were grown in ECM at 37°C with 5%
CO2 and 95% air. The medium was changed every 2 days,
and the cells were checked every day using an inverted
phase-contrast microscope (CKX41; Olympus Corp., Tokyo, Japan).
When the cells had predominantly mixed together and reached ~80%
confluence, the medium was abandoned and the cells were gently
washed twice with phosphate-buffered saline (PBS), prior to
detachment from the culture flask, with the aid of 0.2% (w/v)
trypsin for 2–3 min. A single cell suspension was generated, and
the cells were plated in 6-, 12-, 24- or 96-well plates.
Grouping and treatments
Allicin injection, X/XO and GF109203x were prepared
in serum-free medium at 10 µg/ml, 1 mmol/l/20 U/l and 10 µmol/l,
respectively. The murine aortic endothelial cells in the
exponential growth phase were randomly divided into five groups:
The normal group (NG), the mannitol group (MG), the
high-glucose/hypoxia group (HG), the allicin group (AG) and the PKC
inhibitor (GF109203x) group (GG). The cells of the NG and MG were
respectively incubated with serum-free ECM and mannitol (25 mmol/l)
for 24 h. The cells of the HG, AG and GG were respectively
incubated with X/XO (1 mmol/l/20 U/l), allicin (10 µg/ml) plus X/XO
(1 mmol/l/20 U/l), and GF109203x (10 µmol/l) plus X/XO (1 mmol/l/20
U/l) for 40 min, prior to the supernatant solution being abandoned
and the cells being treated with glucose (25 mmol/l), allicin (10
µg/ml) plus glucose (25 mmol/l), and GF109203x (10 µmol/l) plus
glucose (25 mmol/l), respectively, for 24 h. Following each of the
above treatment regimens, the supernatant and culture cells were
collected. The collected cells were resuspended in PBS. In order to
measure the components within cells, the freeze-thawing method was
adopted to break up the cells.
Cell viability assay
Following the different treatments, the
morphological changes were observed under a phase-contrast
microscope (Olympus Corp.). In addition, MTT assays were performed
to assess the cell viability. First, cell viability was assessed
using MTT when the cells were cultured in different allicin
concentrations of 50, 20, 10, 5 and 2.5 µg/ml (all diluted in
serum-free ECM). In brief, the cells of the six groups (the normal
control group and the five groups of different allicin
concentrations) were seeded into 96-well plates at a density of
1×104 cells per well. Upon reaching ~80% confluence, the
cells were incubated with different concentrations of allicin for
24 h. The supernatant was then abandoned, and 200 µl serum-free
medium, as well as 20 µl MTT dye solution (5 mg/ml), was added to
each well. Following the incubation of the samples at 37°C for 4 h,
the MTT/medium solution was removed and 150 µl dimethylsulfoxide
was added to dissolve the formazan product in each well on a
concentrating table for 15 min. The optical density of each well
was measured using an ELISA microplate reader (Bio-Rad
Laboratories, Inc., Hercules, CA, USA) at a wavelength of 290 nm.
In addition, the cell viability in the NG, MG, HG, AG and GG was
assessed using MTT following treatment with the different
interventions for 24 h.
Dihydroethidium (DHE) staining to
measure ROS
DHE, which is used as a relatively specific
measurement for the superoxide anion, is an oxidative fluorescent
dye that undergoes a two-electron oxidation to form the DNA-binding
fluorophore ethidium bromide. The DHE (Vigorous Biotechnology
Beijing Co., Ltd., Beijing, China) staining for superoxide was
carried out as previously described (14). Briefly, the cells were treated with
the aforementioned interventions for 24 h. Following the removal of
the supernatant solution, the cells were cultured in DHE (10
µmol/l) diluted with PBS in a light-protected, humidified chamber
at 37°C for 30 min. Once the cells had been washed twice with PBS
to remove the uncombined fluorescence probe, fluorescent images
were obtained using a fluorescence microscope. The mean
fluorescence intensity was measured using Image-Pro Plus 6.0
software (Media Cybernetics, Inc., Rockville, MD, USA) for
quantification. The generation of superoxide was demonstrated by
red fluorescent labeling. Non-stained cells were used as a
background control. The average of three DHE-stained images was
taken as the value for each group.
ELISA
The levels of 8-OHdG, NF-κB, Nox4, HIF-1α and PKC in
the cells were determined by ELISA, according to instructions of
each assay kit. Aortic endothelial cells were treated as previously
described for 24 h. Following the removal of the supernatant
solution, the cells were washed twice with ice-cold PBS, scraped
from the plate with trypsin and centrifuged at 1,301 rpm for 5 min
at 4°C. The cells were resuspended in PBS. The freeze-thawing
method was performed to break up the cells.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
RT-qPCR was performed in accordance with the method
described in our previous study (15). Briefly, total RNA was extracted using
TRIzol reagent, following the manufacturer's instructions
(Gibco-BRL). cDNA was synthesized using a commercial reverse
transcription kit (Fermentas; Thermo Fisher Scientific, Waltham,
MA, USA). The sequences of the primers and cycle conditions were as
follows: i) HIF-1α sense, 5′-TCA AGT CAG CAA CGT GGA AG-3′ and
antisense, 5′-TAT CGA GGC TGT GTC GAC TG-3′ (amplification product,
198 bp; 94°C for 30 sec, 59°C for 30 sec and 72°C for 1 min, for 35
cycles); ii) Nox4 sense, 5′-TAG CTG CCC ACT TGG TGA ACG-3′ and
antisense, 5′-TGT AAC CAT GAG GAA CAA TAC CACC-3′ (amplification
product, 170 bp; 94°C for 30 sec, 59°C for 30 sec and 72°C for 1
min, for 30 cycles); iii) NF-κB sense, 5′-GTA TTG CTGTGC CTA CCC
GAA AC-3′ and antisense, 5′-GTT TGA GAT CTG CCC TGA TGG TAA-3′
(amplification product, 134 bp; 94°C for 30 sec, 55°C for 30 sec and
72°C for 1 min, for 30 cycles); iv) β-actin sense, 5′-TGG CAC CCA
GCA CAA TGAA-3′ and antisense, 5′-CTA AGT CAT AGT CCG CCT AGA
AGCA-3′ (amplification product, 188 bp; 94°C for 30 sec, 59°C for 30
sec and 72°C for 1 min, for 30 cycles). The mean value of the
replicates for each sample was calculated and expressed as the
cycle threshold (Ct). The gene expression was then calculated as
the difference (ΔCt) between the Ct value of the target gene and
the Ct value of β-actin.
Statistical analysis
Data are presented as the mean ± standard deviation.
Analysis of variance was used to compare the mean values of more
than two groups. Differences were considered significant at
P<0.05. All statistical calculations were performed using SPSS
13.0 software (SPSS, Inc., Chicago, IL, USA).
Results
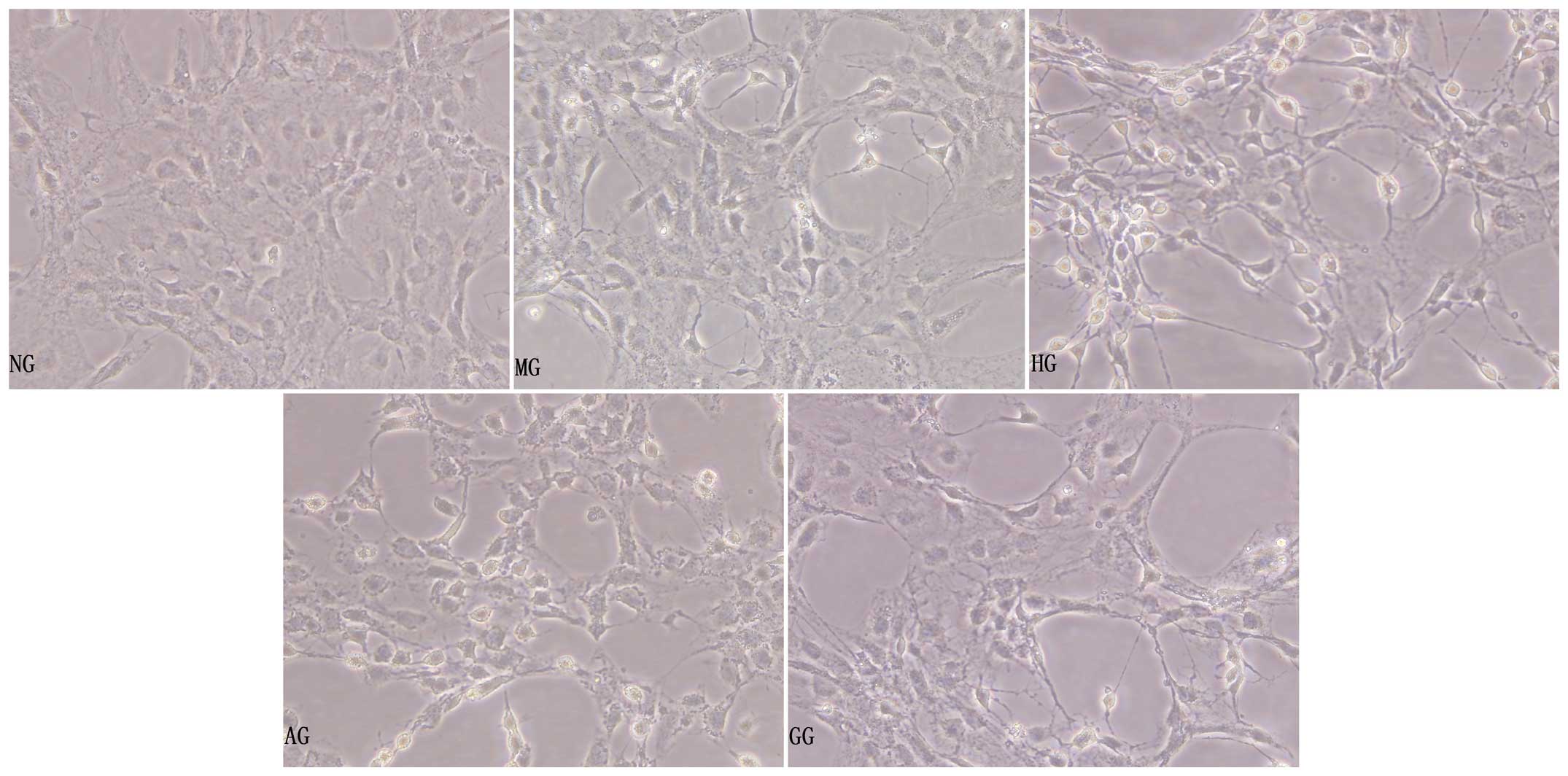
General morphological observation
under the inverted phase-contrast microscope
Under high-glucose/hypoxic conditions for 24 h, the
aortic endothelial cells became shrunken, the intercellular
connection was lessened, some of the cells became exfoliated and a
few of the cells were found to be floating in the supernate
(Fig. 1, HG). The morphology of the
MG cells was not obviously changed compared with that of the NG
cells (Fig. 1, MG). Compared with
the HG cells, the cells in the AG had more complete cell bodies
with visible processes, indicating that allicin played a protective
role in the injured cells (Fig. 1,
AG).
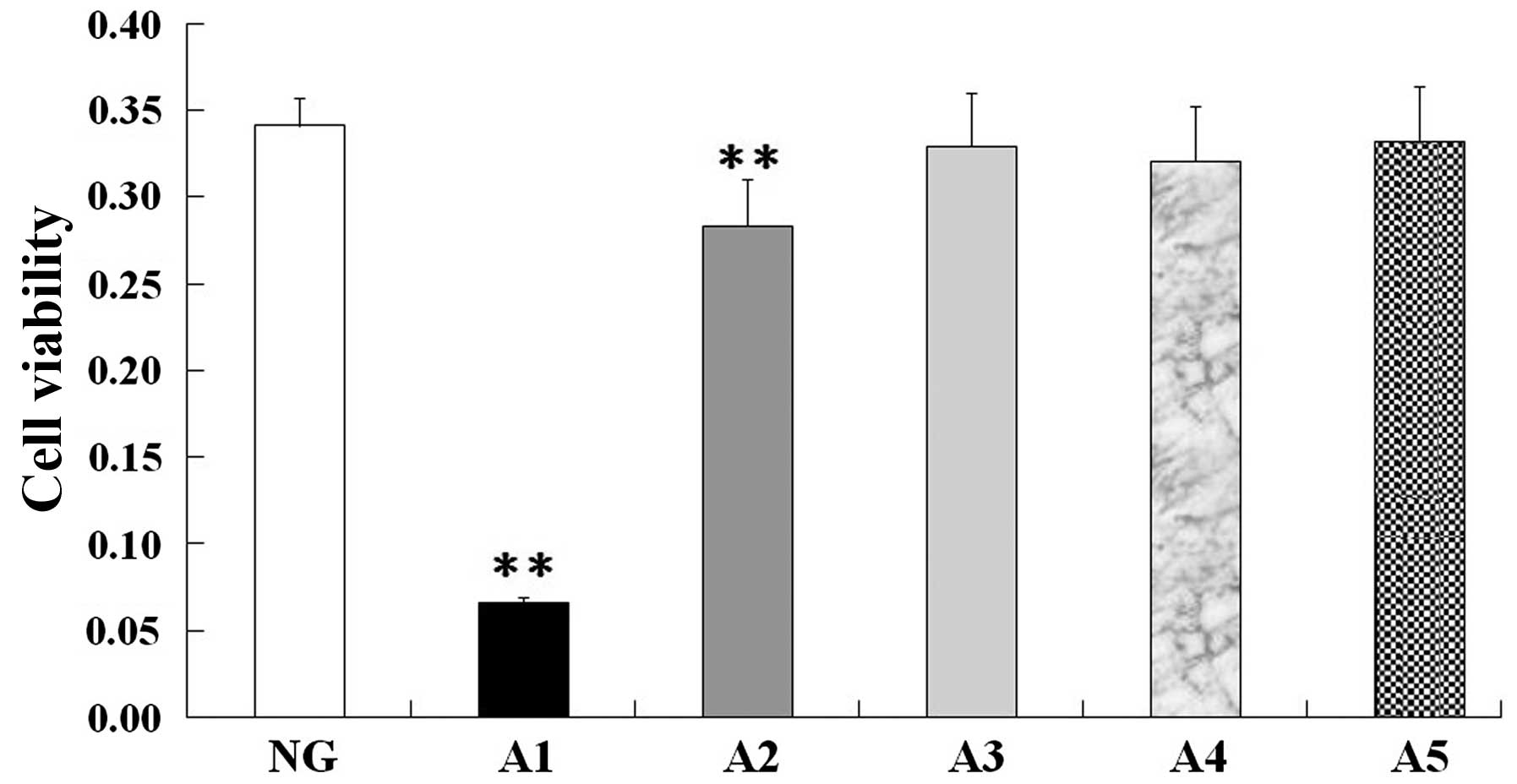
Viability of aortic endothelial
cells
MTT assay revealed no significant differences in
cell viability between the normal control group and the 10, 5 and
2.5 µg/ml allicin groups (Fig. 2, A3, A4
and A5); however, the cell viability of the 50 and 20 µg/ml
allicin groups was significantly decreased compared with that of
the normal control group (P<0.01) (Fig. 2, A1 and A2). Based on these findings,
10 µg/ml allicin was selected as the final treatment concentration.
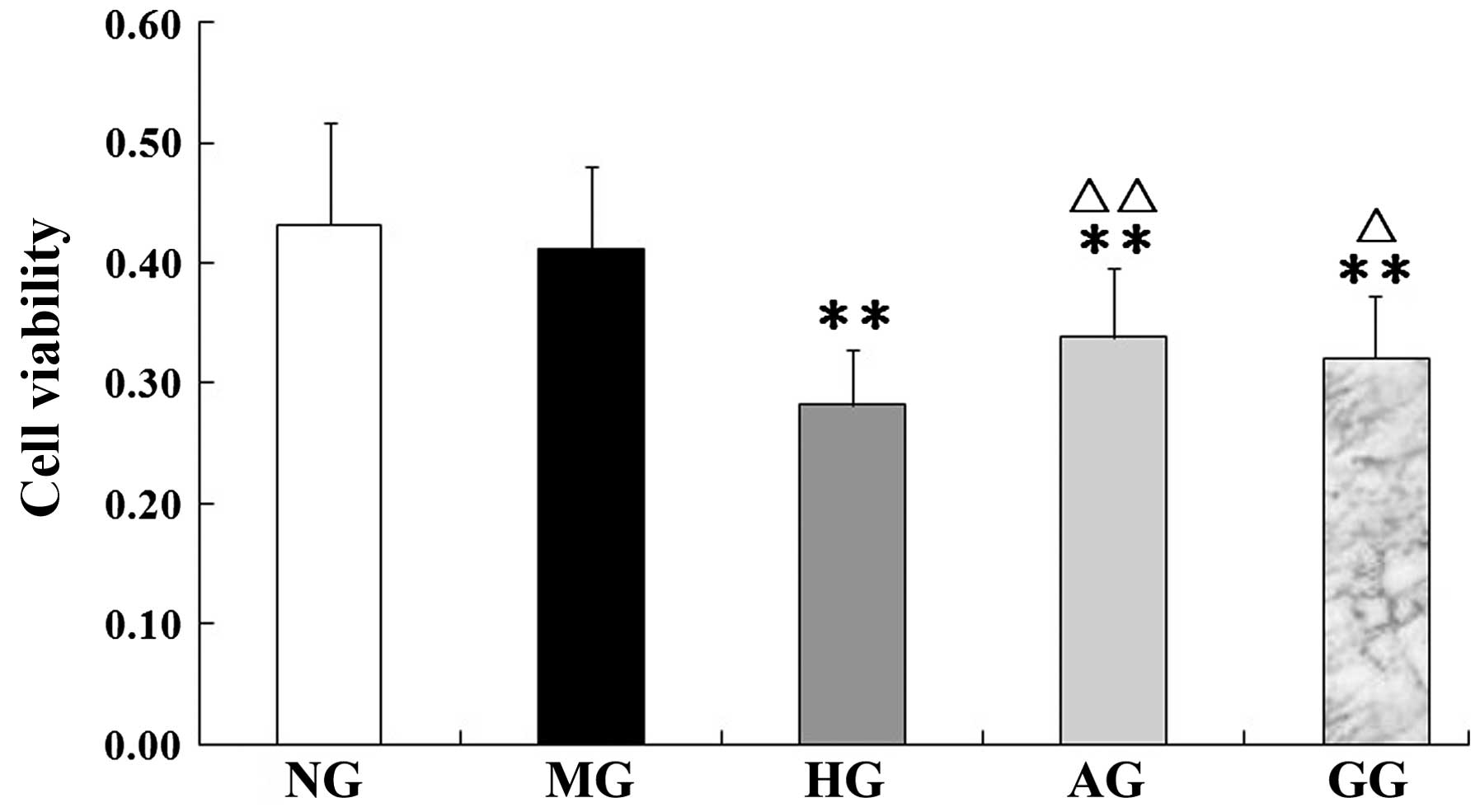
In addition, as shown in Fig. 3, an
evident decrease in viability was observed in the cells under
high-glucose/hypoxic conditions compared with the cells of the NG
(P<0.01); however, in experimental conditions with allicin or
GF109203x the cell viability was significantly increased compared
with that of the HG (P<0.01 or P<0.05). Allicin and GF109203x
exhibited similar effects on cell viability.
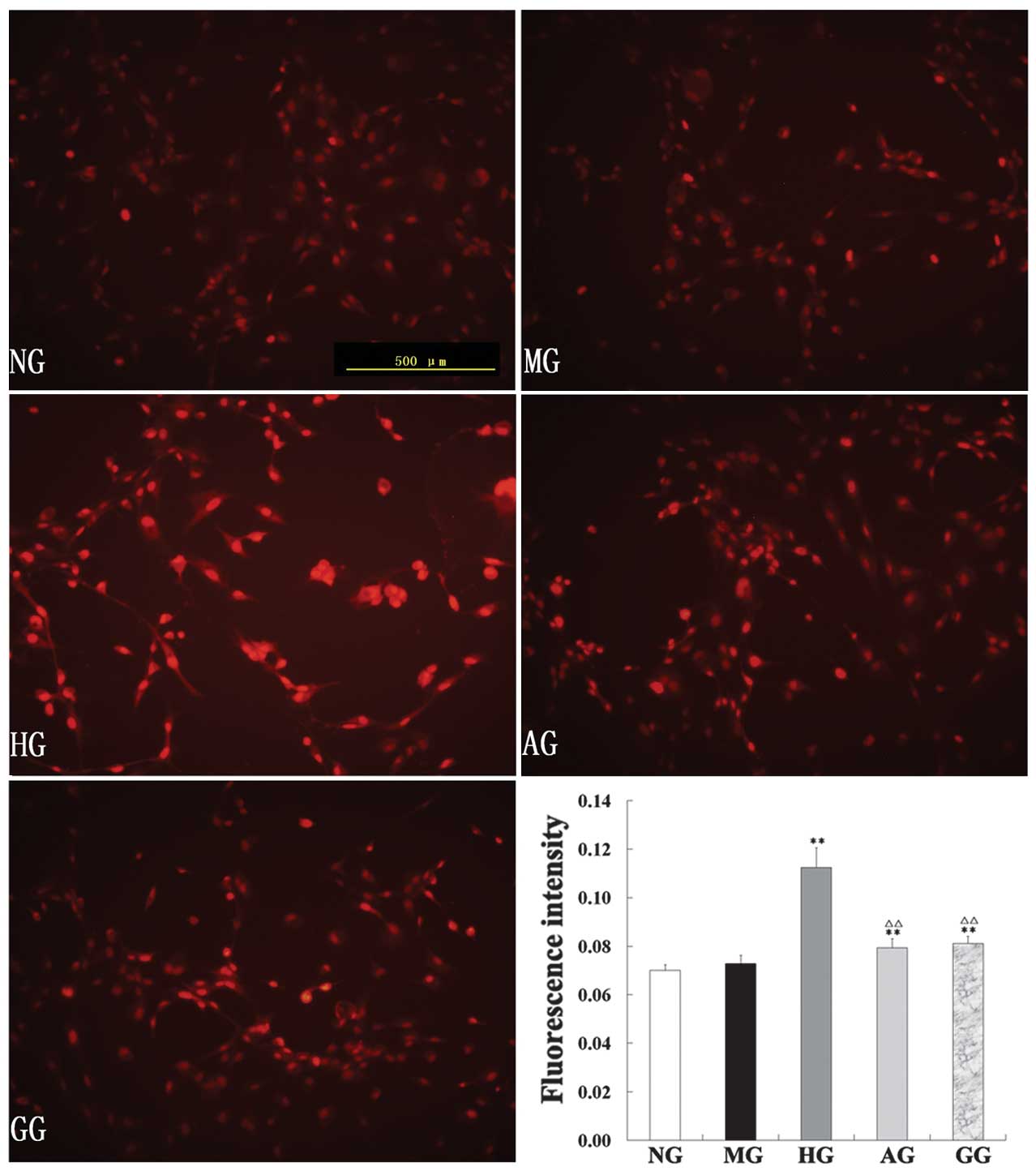
Effect on ROS
ROS production in the endothelial cells of the five
groups was assessed by DHE staining. DHE is a fluorescent dye that
specifically reacts with intracellular ROS and is converted to the
red fluorescent compound ethidium, which then binds irreversibly to
double-stranded DNA and appears as nuclear staining. As shown in
Fig. 4, the intensity of DHE
fluorescence in the endothelial cells in the HG was significantly
enhanced compared with that in the NG; allicin significantly
downregulated the levels of ROS (P<0.01).
Protein levels of 8-OHdG, NF-κB, Nox4
and HIF-1α and activity of PKC
In the HG, the protein levels of 8-OHdG, NF-κB, Nox4
and HIF-1α and the activity of PKC were significantly increased
compared with those of the NG (all P<0.01). In the AG, however,
the five parameters were notably restored, showing a significant
decrease compared with the levels in the HG (P<0.05 or
P<0.01) (Fig. 5).
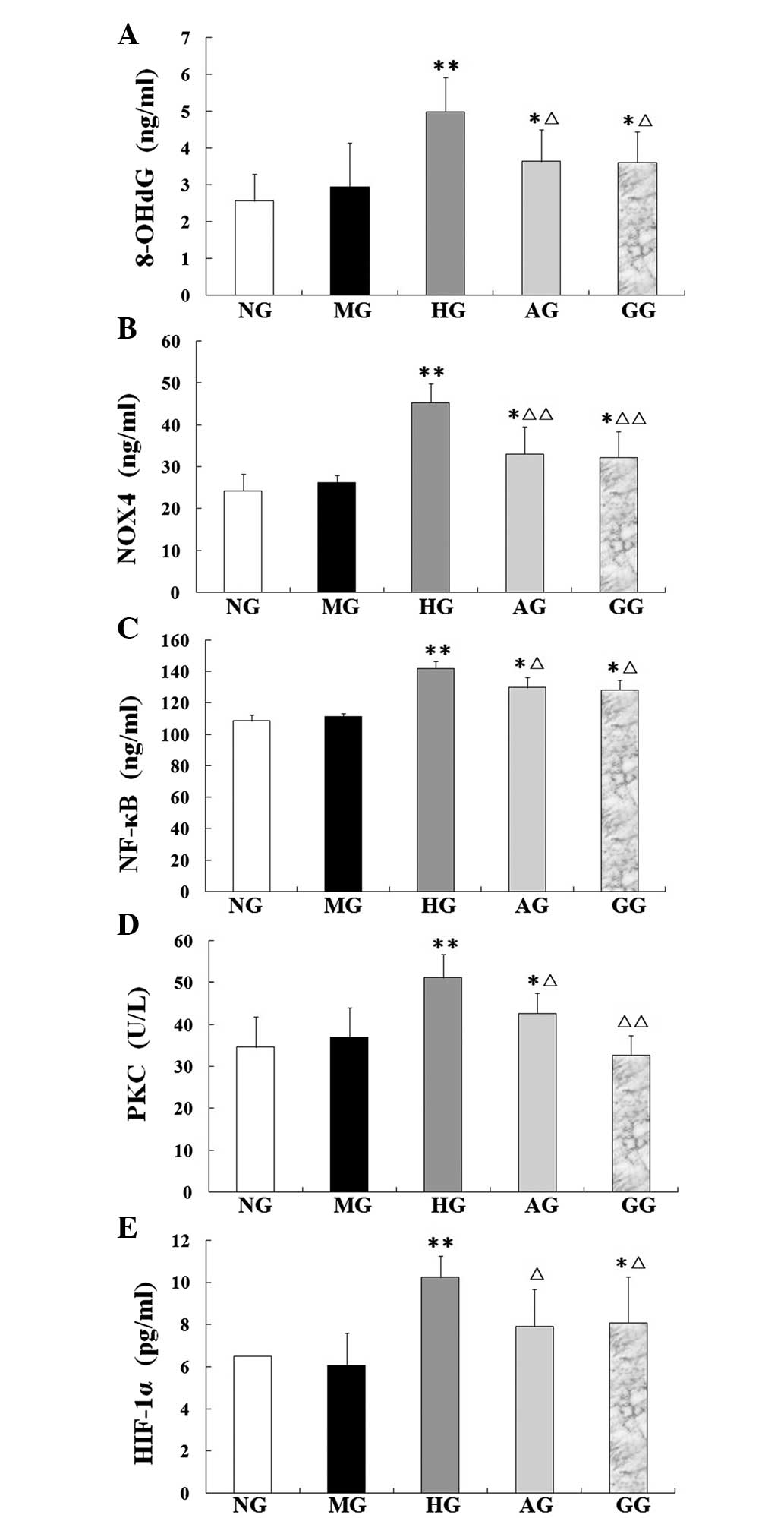 | Figure 5.Levels of (A) 8-OHdG, (B) Nox4, (C)
NF-κB, (D) PKC and (E) HIF-1α in aortic endothelial cells treated
with different interventions. Data are presented as the mean ±
standard deviation (n=10 per group). *P<0.05 and **P<0.01 vs.
NG; ∆P<0.05 and ∆∆P<0.01 vs. HG. NG,
normal group; MG, mannitol group; HG, high glucose/hypoxia group;
AG, allicin group; GG, PKC inhibitor (GF109203x) group; 8-OHdG,
8-hydroxydeoxyguanosine; Nox4, NADPH oxidase 4; NF-κB, nuclear
factor-κB; PKC, protein kinase C; HIF-1α, hypoxia-inducible
factor-1α. |
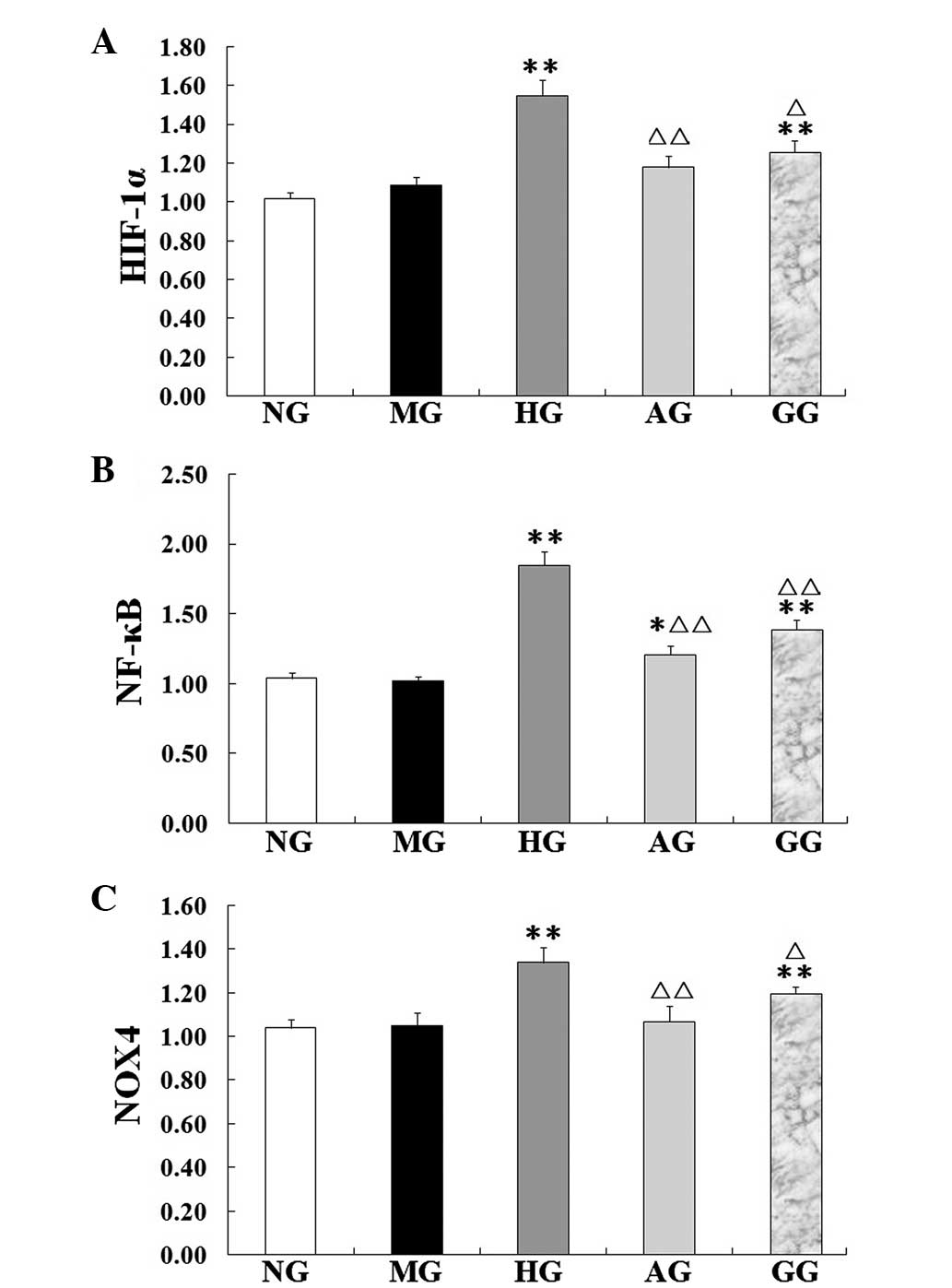
mRNA expression of NF-κB, Nox4 and
HIF-1α
In the HG, the NF-κB, Nox4 and HIF-1α mRNA
expression was found to be significantly increased compared with
the expression in the NG (P<0.01), which suggested that high
glucose/hypoxia could induce the expression of these mRNAs
(Fig. 6). Compared with the HG,
however, the mRNA expression of NF-κB, Nox4 and HIF-1α in the AG
was significantly decreased (P<0.05 or P<0.01).
Discussion
In diabetes mellitus, the risk of atherosclerosis is
enhanced, which results in an increase in the incidence of both
cardiovascular and cerebrovascular diseases, such as myocardial
infarction and cerebral ischemic attacks. Cardiovascular disease is
the major cause of morbidity and mortality in diabetic patients
(16). The exact mechanisms
responsible for this accelerated development of atherosclerosis in
diabetes have remained elusive, but oxidative stress appears to
play a major role. Several studies have reported that the
overproduction of ROS by the mitochondrial electron transport chain
is responsible for hyperglycemia-induced oxidative damage and the
pathogenesis of diabetic complications (17–21).
Hyperglycemia promotes glucose oxidation and protein glycation,
impairs DNA repair, with resultant DNA cleavage, and generates ROS,
thereby leading to increased oxidative stress (22). Oxidative stress is considered to be
the final common pathway through which the hyperglycemia-related
pathways (PKC and polyol) can trigger the chronic complications of
diabetes. Cumulative oxidant-mediated damage and cellular
dysfunction are a result of an imbalance between ROS generation and
antioxidant capacity. Furthermore, studies have found that there is
a hypoxic microenvironment in diabetes, which is closely associated
with the oxidative stress induced by hyperglycemia and plays an
important role in diabetic complications (Amandine, Chavez). The
development of therapeutic strategies aimed at the removal of free
radicals or the prevention of their formation is therefore
necessary.
To determine the injury elicited in aortic
endothelial cells by high glucose/hypoxia and investigate whether
allicin would effectively ameliorate the damage, the expression of
oxidative stress-related markers was examined in endothelial cells
cultured in high glucose (25 mmol/l for 24 h) and hypoxia [X/XO (1
mmol/l/20 U/l) for 40 min], simulating a diabetic microenvironment
in vivo. The possible associated mechanisms were also
explored. Since GF109203x is a highly specific PKC inhibitor, it
was selected for use as a control. The study showed that the cells
in the HG became shrunken and intercellular connections were
lessened; furthermore, some of the cells became exfoliated and a
few of the cells were observed to be floating. The MG was
established to counteract the possible cellular injury by
hyperosmosis, and it was found that the cell morphology of the MG
was not obviously changed compared with that of the NG. Compared
with the HG, the cells in the AG had more complete cell bodies with
visible processes, which indicated that allicin played a protective
role in the injured cells. Consistent with the MTT results, it was
observed that allicin significantly inhibited cell death.
Furthermore, a relatively abundant generation of ROS was measured
in the cells of the HG by the DHE fluorescence probe. As shown in
Fig. 4, allicin significantly
downregulated the levels of ROS, showing its effective attenuation
of the oxidative stress. The increase in ROS production observed in
the endothelial cells of the HG was associated with an increase in
the generation of Nox4 and 8-OHdG. NADPH oxidases of the Nox family
are major sources of ROS, and the level of 8-OHdG is considered to
be a marker of oxidative DNA damage (23). Our preliminary results confirmed the
enhancement of oxidative damage to DNA in the HG; however, the
damage was markedly attenuated when the cells were treated with
allicin. These data suggested that allicin had significantly
protective antioxidative effects against high
glucose/hypoxia-induced injury in aortic endothelial cells.
To the best of our knowledge, the persistent
upregulation of PKC is recognized as an initial event leading to
insulin resistance, cardiac disease and nephropathy in diabetes.
Numerous studies (24–26) have previously reported that multiple
PKC isoforms are activated in the vascular tissue of diabetic
animal models, and the overactivation of the PKC pathway is a key
mediator of diabetic vascular complications. The understanding of
how hyperglycemia-induced oxidative stress ultimately leads to
tissue damage has been advanced considerably (27), and strategies to reduce oxidative
stress may exert favorable effects on the progression of diabetes;
however, effective therapy to prevent or delay the development of
this damage remains limited (28).
The present results revealed that the level of PKC protein
significantly increased in the murine aortic endothelial cells
under high-glucose/hypoxia conditions, and the increase was
mitigated by allicin, which demonstrated that the protective
effects of allicin against the injury induced by high
glucose/hypoxia involved the inhibition of the PKC pathway. PKC has
been noted to contribute to the activation of NADPH oxidases in
multiple cells (29). The Nox system
is considered to be a key contributor to the generation of ROS in
numerous cell types and tissues (30). Increased generation of ROS and
impaired antioxidant defenses contribute to oxidative stress in
diabetes (21). Previous studies
have shown increased levels of NADPH oxidase subunits in the
vasculature and kidney tissue of diabetic rodents (31,32).
Nox4 was initially identified as a kidney NADPH oxidase, but it has
since been shown that Nox4 is also abundant in vascular cells
(33). There is additionally
evidence that Nox4 contributes to oxidative stress in
cardiovascular diseases (34,35). The
present study showed a significant increase in Nox4 levels in the
HG compared with those in the NG. Allicin significantly reduced the
Nox4 mRNA and protein expression. These results suggest that high
glucose/hypoxia increased the Nox4 levels, which caused an
imbalance between the production of free radicals and the
antioxidant defense system. Collectively, these data indicate that
high-glucose conditions induce increased oxidative stress by
activating the PKC pathway, which in turn mediates the increase in
NADPH oxidase activity and Nox4 upregulation. Allicin can inhibit
the PKC pathway, decrease Nox4-derived ROS and the endothelial cell
injury induced by oxidative stress. The effects of allicin in the
present study were similar to those of the PKC inhibitor
GF109203x.
NF-κB is a pleiotropic, oxidant-sensitive
transcription factor that is required for the transcription of the
majority of proinflammatory molecules, including adhesion
molecules, enzymes, cytokines and chemokines, which mediate the
recruitment and retention of monocytes in the subendothelial space;
a key early step in the atherosclerotic process. It has also been
indicated that the activation of PKC is involved in the
hyperglycemia-induced sustained activation of the transcription
factor NF-κB (36). In the present
study, as shown in Figs. 4 and
5, a significant increase was
triggered by high glucose/hypoxia not only in the NF-κB mRNA level,
but also in the NF-κB protein expression. Based on these findings,
it is indicated that high-glucose conditions induced the activation
of the transcription factor NF-κB through the upregulation of PKC.
The upregulation of the PKC pathway leads to the concurrent
upregulation of NADPH oxidase and increase in ROS production, which
can damage endothelial cells and induce the expression of NF-κB,
mediating the recruitment and retention of numerous inflammatory
factors that further aggravate oxidative stress. Allicin can
downregulate both the PKC activation and NF-κB expression. The
inhibition of the PKC pathway should be considered as a target for
the treatment of diabetes and its associated complications, and the
present study showed that allicin inhibited the PKC pathway with a
similar efficacy to that of the PKC inhibitor GF109203x.
Hypoxia has a prominent effect on all diabetic
complications (37). HIF-1α is
highly labile under normal oxygen conditions; however, under
hypoxia it is strongly stabilized by ROS, preventing its
hydroxylation and proteasomal degradation (38,39). In
the present study, X/XO was used to establish the oxygen radical
production system, simulating hypoxic conditions. HIF-1-dependent
gene regulation leads to an adaptation to the hypoxic state,
allowing the cell to survive. Anoxia, however, can induce the
overexpression of HIF-1α, which leads to apoptosis-related gene
expression, resulting in cell injury. In the present study, it was
found that, dependent on the extent of the high glucose and
hypoxia, HIF-1α levels also significantly increased. These results
showed that endothelial cells incubated under high-glucose/hypoxic
conditions expressed significantly high levels of HIF-1α, which
were a risk factor for vascular lesion. In addition, allicin
downregulated the expression of HIF-1α.
A broad range of antioxidant properties have been
attributed to allicin, including direct scavenging of free
radicals, maintenance of glutathione and the endogenous antioxidant
redox balance, as well as the enhanced expression of the
antioxidant enzymes glutathione peroxidase, glutathione reductase,
SOD and CAT (13). Furthermore,
allicin can suppress the production of certain inflammatory
cytokines that have been implicated in the pathogenesis of diabetic
complications, including tumor necrosis factor-α and transforming
growth factor-1 (40). Specifically,
a decrease in oxidative stress has been associated with reduced
diabetes-associated atherosclerotic plaque development. As
oxidative stress is associated with diabetic complications,
antioxidation may be a key factor in the treatment of these
diseases. The results of the present study indicate that allicin
exerted beneficial effects on the treatment of macroangiopathy and
could inhibit endothelial cell apoptosis and promote cellular
survival under high-glucose/hypoxic conditions. These data offer a
plausible explanation for the effects of allicin in vivo,
which appear to be mediated at least partially by the inhibition of
the PKC pathway and the consequent decrease in ROS production. In
conclusion, the results of the present study suggest that allicin
exerts protective effects via the inhibition of the
high-glucose/hypoxia-induced upregulation of the PKC pathway and
the subsequent activation of NADPH oxidase, increased ROS
production, endothelial cell injury, 8-OHdG release and NF-κB
upregulation. Inhibition of the PKC pathway may be the common
protective mechanism of allicin in diabetes.
Acknowledgements
The authors would like to thank Dr Chuanju Liu and
Mr. Brendon Richbourgh at the New York University School of
Medicine for checking the manuscript.
References
|
1
|
Norhammar A, Tenerz A, Nilsson G, Hamsten
A, Efendíc S, Rydén L and Malmberg K: Glucose metabolism in
patients with acute myocardial infarction and no previous diagnosis
of diabetes mellitus: A prospective study. Lancet. 359:2140–2144.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Rolo AP and Palmeira CM: Diabetes and
mitochondrial function: Role of hyperglycemia and oxidative stress.
Toxicol Appl Pharmcol. 212:167–178. 2006. View Article : Google Scholar
|
|
3
|
Olmez I and Ozyurt H: Reactive oxygen
species and ischemic cerebrovascular disease. Neurochem Int.
60:208–212. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Laderoute KR: The interaction between
HIF-1 and AP-1 transcription factors in response to low oxygen.
Semin Cell Dev Biol. 16:502–513. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Nitatori T, Sato N, Waguri S, Waguri S,
Karasawa Y, Araki H, Shibanai K, Kominami E and Uchiyama Y: Delayed
neuronal death in the CA1 pyramidal cell layer of the gerbil
hippocampus following transcient ischemia is apoptosis. J Neurosci.
15:1001–1011. 1995.PubMed/NCBI
|
|
6
|
Patumraj S, Tewit S, Amatyakul S,
Jariyapongskul A, Maneesri S, Kasantikul V and Shepro D:
Comparative effects of garlic and aspirin on diabetic
cardiovascular complications. Drug Deliv. 7:91–96. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Eidi A, Eidi M and Esmaeili E:
Antidiabetic effect of garlic (Allium sativum L.) in normal
and streptozotocin-induced diabetic rats. Phytomedicine.
13:624–629. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ashraf R, Khan RA and Ashraf I: Garlic
(Allium sativum) supplementation with standard antidiabetic
agent provides better diabetic control in type 2 diabetes patients.
Pak J Pharm Sci. 24:565–570. 2011.PubMed/NCBI
|
|
9
|
Nasim SA, Dhir B, Kapoor R, Fatima S,
Mahmooduzzafar and Mujib A: Alliin obtained from leaf extract of
garlic grown under in situ conditions possess higher therapeutic
potency as analyzed in alloxan-induced diabetic rats. Pharm Biol.
49:416–421. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Borek C: Garlic reduces dementia and
heart-disease risk. J Nutr. 136:(3 Suppl). S810–S812. 2006.
|
|
11
|
Ou CC, Tsao SM, Lin MC and Yin MC:
Protective action on human LDL against oxidation and glycation by
four organosulfur compounds derived from garlic. Lipids.
38:219–224. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Liu DS, Gao W, Liang ES, Wang SL, Lin WW,
Zhang WD, Jia Q, Guo RC and Zhang JD: Effects of allicin on
hyperhomocysteinemia-induced experimental vascular endothelial
dysfunction. Eur J Pharmacol. 714:163–169. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Rajani Kanth V, Uma Maheswara Reddy P and
Raju TN: Attenuation of streptozotocin-induced oxidative stress in
hepatic and intestinal tissues of Wistar rat by methanolic-garlic
extract. Acta Diabetol. 45:243–251. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Guzik TJ, Mussa S, Gastaldi D, Sadowski J,
Ratnatunga C, Pillai R and Channon KM: Mechanisms of increased
vascular superoxide production in human diabetes mellitus: Role of
NAD(P)H oxidase and endothelial nitric oxide synthase. Circulation.
105:1656–1662. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Liu DS, Zhou YH, Liang ES, Li W, Lin WW,
Chen FF and Gao W: Neuroprotective effects of the Chinese
Yi-Qi-Bu-Shen recipe extract on injury of rat hippocampal neurons
induced by hypoxia/reoxygenation. J Ethnopharmacol. 145:168–174.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Bryden KS, Dunger DB, Mayou RA, Peveler RC
and Neil HA: Poor prognosis of young adults with type 1 diabetes: A
longitudinal study. Diabetes Care. 26:1052–1057. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Mäkinen VP, Forsblom C, Thorn LM, Wadén J,
Kaski K, Ala-Korpela M and Groop PH: Network of vascular diseases,
death and biochemical characteristics in a set of 4,197 patients
with type 1 diabetes (the FinnDiane Study). Cardiovasc Diabetol.
8:542009. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Brownlee M: The pathobiology of diabetic
complications: A unifying mechanism. Diabetes. 54:1615–1625. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Duchen MR: Roles of mitochondria in health
and disease. Diabetes. 53:(Suppl 1). S96–S102. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Nishikawa T, Edelstein D, Du XL, Yamagishi
S, Matsumura T, Kaneda Y, Yorek MA, Beebe D, Oates PJ, Hammes HP,
et al: Normalizing mitochondrial superoxide production blocks three
pathways of hyperglycaemic damage. Nature. 404:787–790. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kiritoshi S, Nishikawa T, Sonoda K,
Kukidome D, Senokuchi T, Matsuo T, Matsumura T, Tokunaga H,
Brownlee M and Araki E: Reactive oxygen species from mitochondria
induce cyclooxygenase-2 gene expression in human mesangial cells:
Potential role in diabetic nephropathy. Diabetes. 52:2570–2577.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Retinopathy and nephropathy in patients
with type 1 diabetes four years after a trial of intensive therapy.
The diabetes control and complications trial/epidemiology of
diabetes interventions and complications research group. N Engl J
Med. 342:381–389. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Tuzgen S, Hamnimoglu H, Tanriverdi T,
Kacira T, Sanus G, Atukereny P, Dashti R, Gumustas K, Canbaz B and
Kaynar MY: Relationship between DNA damage and total antioxidant
capacity in patients with glioblastoma multiforme. Clin Oncol (R
Coll Radiol). 19:177–181. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Inoguchi T, Battan R, Handler E, Sportsman
JR, Heath W and King GL: Preferential elevation of protein kinase C
isoform beta II and diacylglycerol levels in the aorta and heart of
diabetic rats: Differential reversibility to glycemic control by
islet cell transplantation. Proc Natl Acad Sci USA. 89:11059–11063.
1992. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Craven PA, Studer RK, Negrete H and
DeRubertis FR: Protein kinase C in diabetic nephropathy. J Diabetes
Complications. 9:241–245. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Koya D, Jirousek MR, Lin YW, Ishii H,
Kuboki K and King GL: Characterization of protein kinase C beta
isoform activation on the gene expression of transforming growth
factor-beta, extracellular matrix components and prostanoids in the
glomeruli of diabetic rats. J Clin Invest. 100:115–126. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Brownlee M: Biochemistry and molecular
cell biology of diabetic complications. Nature. 414:813–820. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Hamilton CA, Miller WH, Al-benna S,
Brosnan J, Drummond RD, McBride MW and Dominiczak AF: Strategies to
reduce oxidative stress in cardiovascular diaease. Clin Sci (Lond).
106:219–234. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Dekker LV, Leitges M, Altschuler G, Mistry
N, McDermott A, Roes J and Segal AW: Protein kinase C-beta
contributes to NADPH oxidase activation in neutrophils. Biochem J.
347:285–289. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Serrander L, Cartier L, Bedard K, Banfi B,
Lardy B, Plastre O, Sienkiewicz A, Fórró L, Schlegel W and Krause
KH: NOX4 activity is determined by mRNA levels and reveals a unique
pattern of ROS generation. Biochem J. 406:105–114. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Gorin Y, Block K, Hernandez J, Bhandari B,
Wagner B, Barnes JL and Abboud HE: Nox4 NAD(P)H oxidase mediates
hypertrophy and fibronectin expression in the diabetic kidney. J
Biol Chem. 280:39616–39626. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Goyal P, Weissmann N, Rose F, Grimminger
F, Schäfers HJ, Seeger W and Hänze J: Identification of novel Nox4
splice variants with impact on ROS levels in A549 cells. Biochem
Biophys Res Commun. 329:32–39. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Griendling KK: Novel NAD(P)H oxidases in
the cardiovascular system. Heart. 90:491–493. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
San Martín A, Du P, Dikalova A, Lassègue
B, Aleman M, Góngora MC, Brown K, Joseph G, Harrison DG, Taylor WR,
et al: Reactive oxygen species-selective regulation of aortic
inflammatory gene expression in Type 2 diabetes. Am J Physiol Heart
Circ Physiol. 292:H2073–H2082. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Thandavarayan RA, Watanabe K, Ma M,
Gurusamy N, Veeraveedu PT, Konishi T, Zhang S, Muslin AJ, Kodama M
and Aizawa Y: Dominant-negative p38 mitogen-activated protein
kinase prevents cardiac apoptosis and remodeling after
streptozotocin-induced diabetes mellitus. Am J Physiol Heart Circ
Physiol. 297:H911–H919. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Yerneni KK, Bai W, Khan BV, Medford RM and
Natarajan R: Hyperglycemia-induced activation of nuclear
transcription factor kappaB in vascular smooth muscle cells.
Diabetes. 48:855–864. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Cameron NE, Eaton SE, Cotter MA and
Tesfaye S: Vascular factors and metabolic interactions in the
pathogenesis of diabetic neuropathy. Diabetologia. 44:1973–1988.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Klimova T and Chandel NS: Mitochondrial
complex III regulates hypoxic activation of HIF. Cell Death Differ.
15:660–666. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Fandrey J, Gorr TA and Gassmann M:
Regulating cellular oxygen sensing by hydroxylation. Cardiovasc
Res. 71:642–651. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Madkor HR, Mansour SW and Ramadan G:
Modulatory effects of garlic, ginger, turmeric and their mixture on
hyperglycaemia, dyslipidaemia and oxidative stress in
streptozotocin-nicotinamide diabetic rats. Br J Nutr.
105:1210–1217. 2011. View Article : Google Scholar : PubMed/NCBI
|