Introduction
Bone fracture can initiate a series of
bio-physiological and pathological reactions, and a variety of
biomechanical and biological factors that determine the outcome of
fracture healing have been identified during past decades (1,2).
Numerous studies have indicated that autophagy plays an important
role in nutrient and energy regulation, as well as in the removal
of dysfunctional or damaged organelles and molecules (3,4). Bone
fracture can disrupt the cellular homeostasis and place the bone
cells under considerable stress, which can result in the activation
of autophagy. The involvement of autophagy in neurodegeneration,
cardiomyopathies and abnormal skeletal development has been
investigated in previous studies (5–7);
however, the role of autophagy in the various stages of bone
fracture remains to be studied.
Autophagy is a common physiological mechanism that
targets altered and dysfunctional cytosolic macromolecules,
membranes and organelles for delivery to lysosomes for degradation
and recycling (8,9). The process of autophagy is activated
when there is a shortage of energy or nutrients. At the beginning
of the process, an autophagosome is created via a double membrane
that forms around cellular substances, which then fuses with a
lysosome. The organelles and proteins in cells are degraded to
amino and fatty acids by the formed lysosome, to enable cell
survival (10). Previous studies
have demonstrated that autophagy is associated with the progression
of numerous diseases, such as heart disease (11), and is involved in tumor suppression
(12) and the removal of toxic
agents (13). The role of autophagy
has additionally been investigated in the field of bone science,
including in the pathogenesis of osteoporosis (14) and Paget's disease (15).
Microtubule-associated protein II light chain 3
(LC3-II) is one of the major regulators of the autophagy pathway.
The protein binds to the membrane of the autophagosome, and the
extent of autophagosome formation is correlated with the level of
LC3-II formation (16). Thus, the
upregulation of LC3-II is often used to illustrate the activation
of autophagy in disease models (17).
In a regenerating tissue, cell proliferation plays
an important role in tissue amplification, particularly in
physiological and pathological processes (18). In a previous study, Lee et al
(19) quantified the proliferating
cells in each of the cellular events occurring during bone fracture
healing. The degree of cell proliferation varied according to the
length of time that had passed since the fracture, suggesting the
existence of local regulatory factors, such as growth factors and
cytokines (20). Proliferating cell
nuclear antigen (PCNA) is a crucial protein for the proliferation
of osteoblasts, its expression and cell proliferation cycle are
closely associated, and thus accurately reflect the cell
proliferation. The aim of the present study, therefore, was to
investigate the role of autophagy in the bone following fracture
and describe the association between cell proliferation and
autophagy, in order to identify a potential therapeutic target to
improve fracture healing. The protein expression of PCNA was used
to investigate the cell proliferation in bone tissue following
fracture via immunohistochemical analysis.
Materials and methods
Experimental groups and surgical
procedure
The present study was performed according to
protocols approved by the local governmental Animal Care Committee
and the institutional Animal Care and Use Committee at Xiamen
University (Xiamen, China). Every effort was made to minimize
animal suffering and to reduce the number of animals used. In
total, 36 adult male Wistar rats weighing 230–260 g were obtained
from the Experimental Animal Center, the Affiliated Southeast
Hospital of Xiamen University (SCZZ(min)2012-0203). The rats were
randomly separated into six groups (n=6/group): Five experimental
groups (for examination at different time-points) and one
sham-surgery group (as a control group). The rats were housed with
3 or 4 rats to a cage, 1 week before the experiments began. For the
experimental procedure, in brief, the rats were anesthetized using
an intraperitoneal injection of ketamine (75 mg/kg) and xylazine
(25 mg/kg), and a lateral incision was then made through the shaved
skin and fascia lata from the right knee to the greater trochanter.
The plane between the vasti and hamstrings was opened through blunt
dissection to expose the femur. The right femur of each animal was
fractured using a 3-point bending device and stabilized using
Kirschner wire (diameter, 1.0 mm; Shanghai Pudong Jinhuan Medical
Products Co., Ltd., Pudong, Shanghai, China), as described
previously (21). The fascia lata
and skin were closed with polyglactin absorbable sutures (Shanghai
Pudong Jinhuan Medical Products Co., Ltd.). The fracture
configuration was a closed, midshaft fracture type A2-A3, according
to AO classification (22), and the
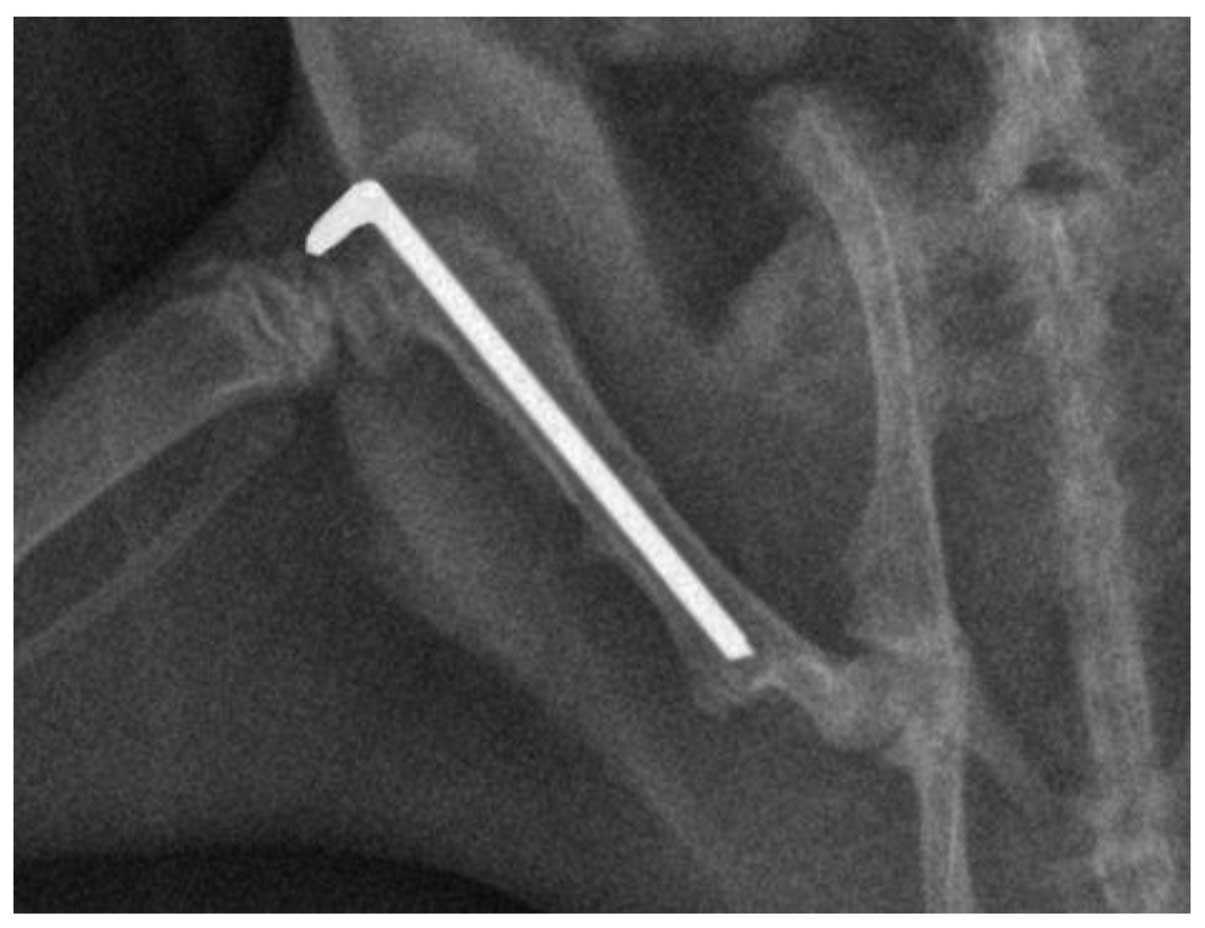
implant positions were documented using X-rays (Fig. 1). The sham-surgery group underwent an
identical procedure without fracture.
The experimental group samples were extracted near
the fracture site at 6 h, 12 h, 24 h, 3 days and 7 days after
fracture. The samples were then fixed in 4% formalin for 24 h and
decalcified in 10% EDTA solution for 5 weeks. A total of 10 slices
per animal (the results from which were averaged) were randomly
selected for paraffin embedding and used for quantitative
immunofluorescence and immunohistochemical analyses.
Immunofluorescence
Paraffin-embedded samples for immunofluorescence
were deparaffinized in the xylene substitute Pro-Par Clearant
(Anatech, Ltd., Battle Creek, MI, USA) and rehydrated in graded
ethanol and water. After washing with phosphate-buffered saline
(PBS), the sections (5-µm) were blocked with 5% goat serum for 1 h
at room temperature and then incubated at 4°C with rabbit
polyclonal anti-LC3II (1:50; NB910-40435; Sigma-Aldrich, St. Louis,
MO, USA) overnight. After further washing with PBS, the sections
were incubated with the Alexa Fluor® 488 anti-rabbit IgG secondary
antibody (Abcam, Cambridge, MA, USA) for 30 min. Finally, the
sections were washed and observed using fluorescence microscopy
Immunohistochemistry
For immunohistochemistry, 6-µm sections from
paraffin-embedded samples were deparaffinized and rehydrated. For
antigen unmasking, the sections were immersed in 10 mM sodium
citrate buffer (pH 6.0), boiled in a microwave oven and kept at
92–95°C for 2 min. Following antigen unmasking, the slides were
cooled for 20 min at room temperature and then washed with PBS. The
sections were subsequently blocked with 0.03% hydrogen peroxide
sodium azide (supplied by the Affiliated Southeast Hospital of
Xiamen University) for 5 min at room temperature. Purified rabbit
anti-rat PCNA monoclonal antibody (1:100; GTX12496; Dako North
America, Inc., Carpinteria, CA, USA) was incubated for 45 min at
room temperature. After washing with PBS, the secondary purified
goat anti-rabbit/mouse biotinylated antibody (1:100; Dako North
America, Inc.) was applied to the sections and incubated for 30 min
at room temperature. The sections were then washed gently with PBS
and incubated with streptavidin-peroxidase (1:50; Dako North
America, Inc.) for 30 min at room temperature. Diaminobenzidine
substrate chromagen was added to the sections, which were
subsequently incubated at room temperature for >5 min and
washed. The tissue sections were counterstained with hematoxylin
(Sigma-Aldrich) for 5 min, washed with distilled water and
dehydrated through a series of graded ethanol solutions to xylene,
mounted in DPX (Olympus Corporation, Tokyo, Japan) and examined
using light microscopy.
Statistical analysis
The percentage of PCNA-positive cells and the number
of the cells with punctuate LC3-II fluorescence were calculated in
10 randomly selected slices per animal and averaged, and the
results were then quantitatively analyzed using Image-Pro Plus
software (Media Cybernetics, Inc., Silver Spring, MD, USA). Results
are presented as the mean ± standard deviation. Significant
differences between animals at each time-point were assessed using
analysis of variance, and correlation analysis was conducted using
linear regression. Statistical analysis was performed using SPSS
software for Windows (version 13.0; SPSS, Inc., Chicago, IL, USA).
P<0.05 was considered to indicate a statistically significant
difference.
Results
LC3-II is upregulated in bone tissue
following fracture injury
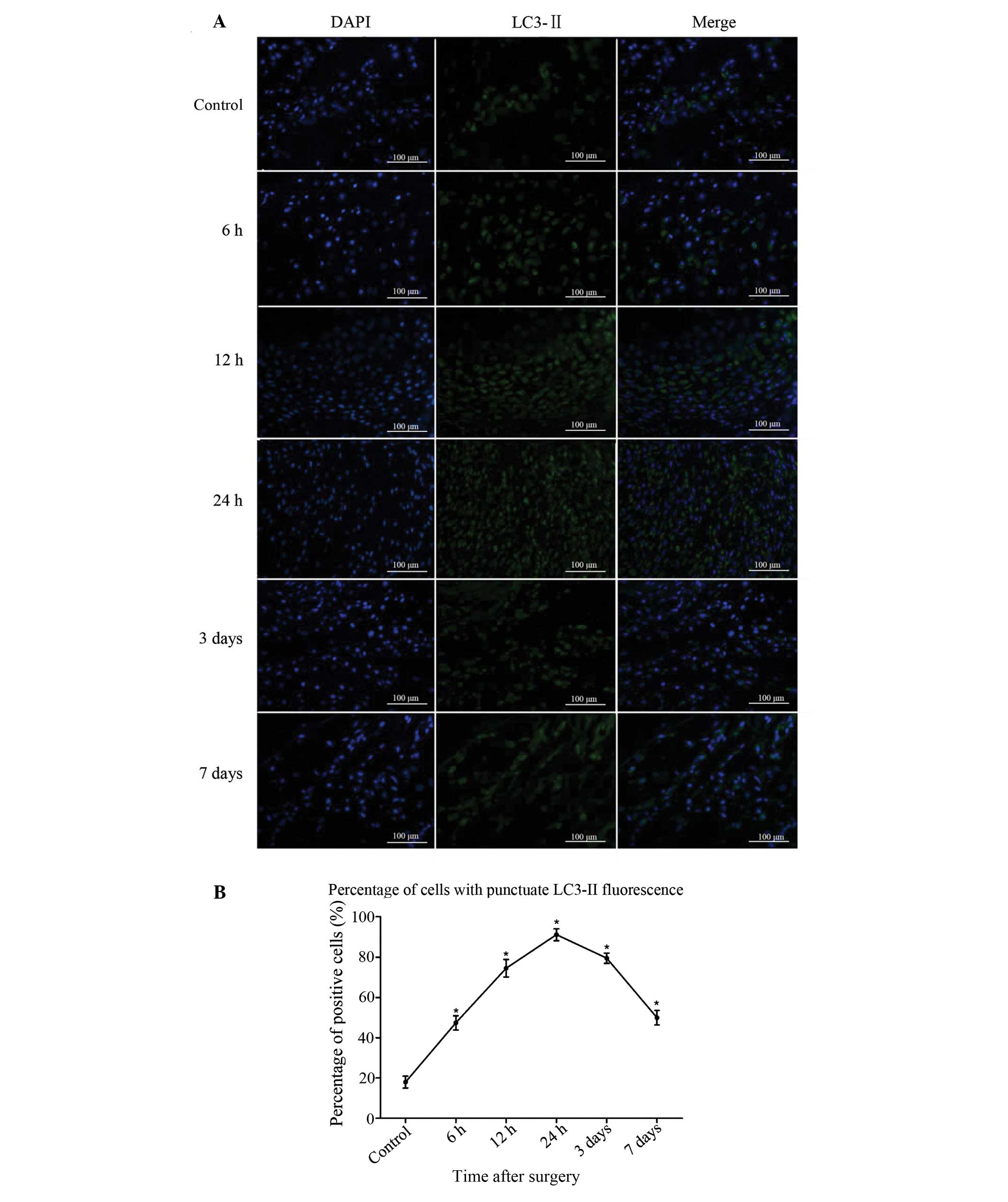
LC3-II-positive cells were sparse in the bone of the
sham-surgery group. At 6 h after injury, the number of
LC3-II-positive cells began to increase, peaked at 24 h and then
reduced gradually from 3 days after bone fracture (Fig. 2A). The difference between the
experimental and sham-surgery groups was significant at all
time-points (P<0.05). Significant differences were also found
among the data from the 5 experimental groups (P<0.05). The
results demonstrate that the process of autophagy was induced in
the bone tissue following fracture injury (Fig. 2B).
An increase in the number of
PCNA-positive cells is observed in bones from rats with surgically
induced fracture
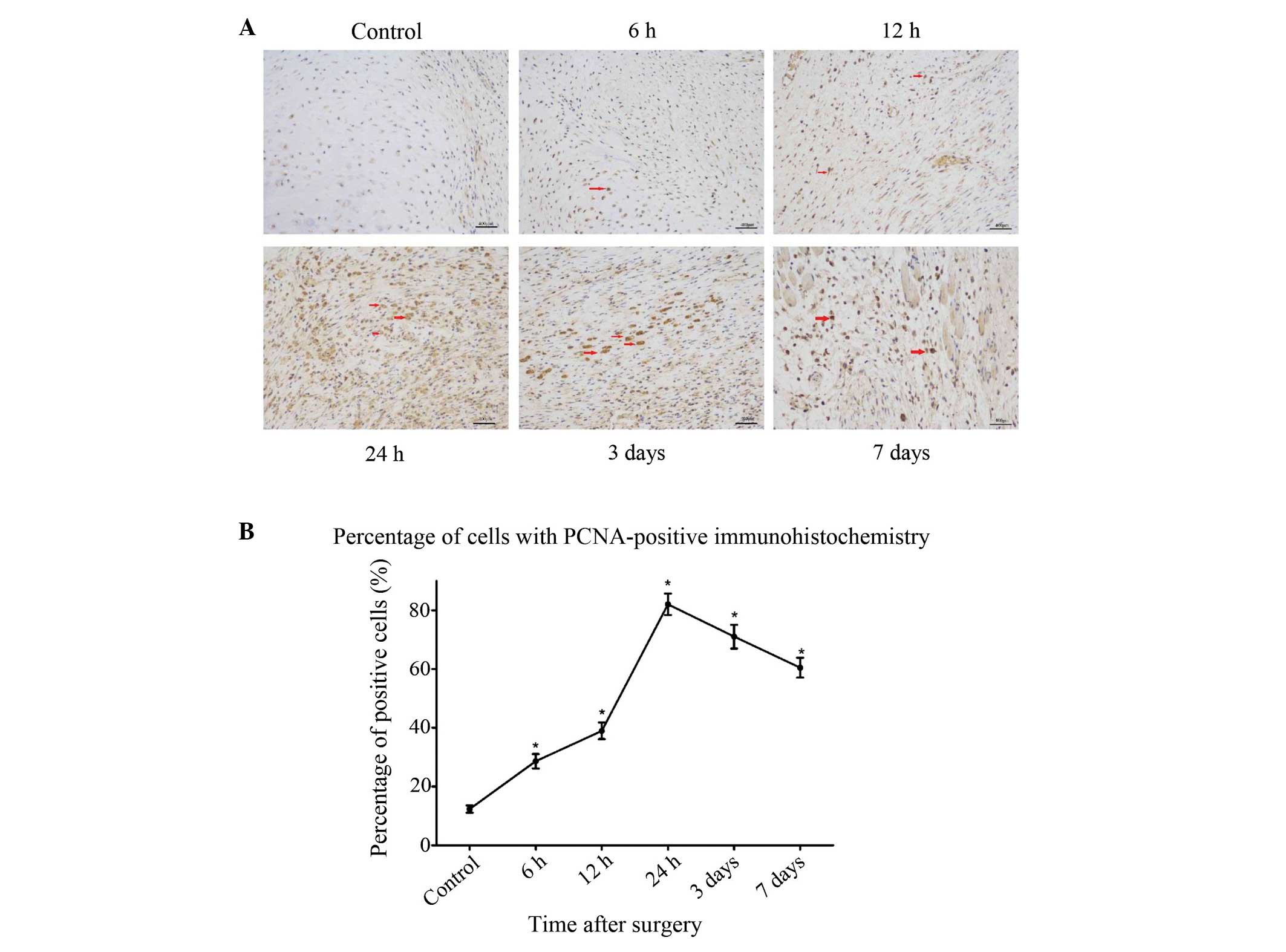
To further investigate the association between
autophagy and cell proliferation following fracture injury,
immunohistochemical staining for the cell proliferation marker PCNA
was performed on the same samples that were analyzed for the
autophagy marker in the preceding experiments. At 6 h after
fracture, an increased number of PCNA-positive cells was observed
compared with the sham-surgery group, and this increase lasted for
24 h after the surgery (P<0.05) (Fig.
3). After 3 days, the pathological changes began to decrease,
which coincided with the recovery of blood flow and nutrition.
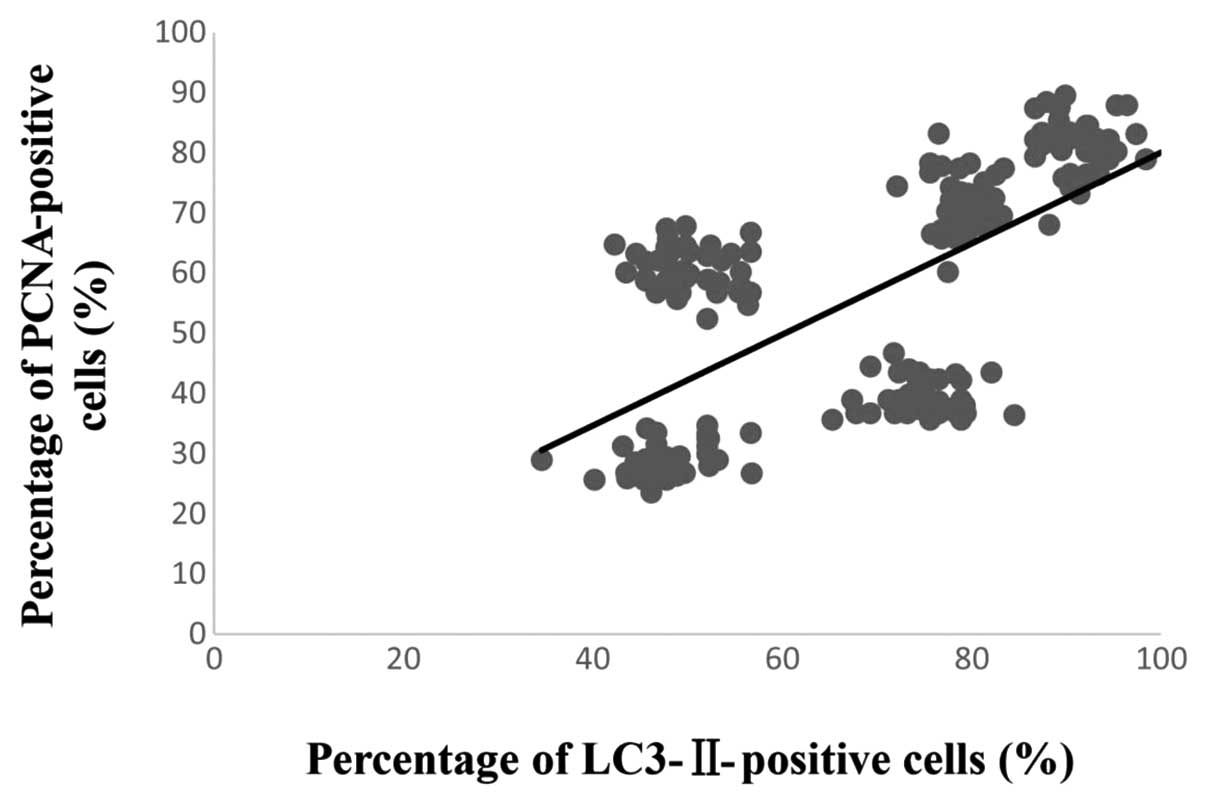
Correlation between the number of
LC3-II- and PCNA-positive cells
Simple linear regression was used to analyze the
correlation between the number of LC3-II- and PCNA-positive cells.
The result showed that a significant correlation existed between
the degree of LC3-II and PCNA expression following fracture injury
(R2=0.43, P<0.01) (Fig.
4). These findings demonstrate that surgically induced femur
fracture is associated with the activation of autophagy (LC3-II
expression) along with an increase in cell proliferation (PCNA
expression).
Discussion
In the present study, the occurrence of certain
cellular events, i.e. autophagy and proliferation, was analyzed in
the various phases of bone fracture healing following internal
fixation in a rat femoral fracture model. Autophagy, type II
programmed cell death, is a cellular mechanism of self-cycling for
survival in eukaryotes. The study of autophagy is an emerging field
within bone research. To investigate the degree of autophagy in
bone tissue following fracture injury, immunofluorescence was used
with an LC3-II polyclonal antibody, a method that has already been
described and used previously (23).
In the present study, the protein expression of LC3-II increased in
the bone tissue following fracture, peaking at 24 h, and then
exhibited a gradual decline from 3 days. Significant differences
were observed at each time-point (P<0.05). A few LC3-II-positive
cells were additionally observed in the sham-surgery group,
indicating that autophagy exists within a narrow homeostatic range
and in delicate balance with other cellular systems that regulate
protein homeostasis and cell survival. This theory has been
described in a previous study (24).
Once the balance is disrupted, autophagy can be induced as a
survival mechanism in normal cells in response to hypoxic
conditions or stress (25). There is
growing evidence that energy status and nutrient levels are key
modulators of autophagy, and autophagy is defined as a catabolic,
energy-generating mechanism for the cell (26,27). In
the present study it was hypothesized that autophagy would be
activated in bone tissue following fracture injury due to the
sudden reduction or interruption of the nutrient supply.
Cell proliferation appears to be a major cellular
response at the beginning of a cellular event, such as
inflammation, chondrogenesis and endochondral or intramembranous
ossification. PCNA is a key marker for bone cell proliferation and
plays an important role in both pathological and physiological
activity (28). In the present
study, PCNA-positive cells were detected at 6 h near the fracture
site, concurring with the findings of Iwaki et al (29), which suggested that fracture healing
began with cell proliferation. In addition, the present results
showed that the percentage of PCNA- and LC3-II-positive cells
exhibited similar trends in variation following fracture injury at
each experimental time-point. Linear regression analysis
demonstrated that a significant correlation was present between
PCNA and LC3-II protein expression following fracture injury
(R2=0.43, P<0.01). In a previous study, Morrow et
al (30) demonstrated that
rapamycin caused the activation of the nuclear translocation of
PCNA in CD4+ T cells. It has been reported that one of
the key suppressors of autophagy is mammalian target of rapamycin
(mTOR); rapamycin exerts significant inhibitory effects on mTOR,
with no off-target effects on other enzymes (31,32).
Based on the findings of Morrow et al (30), we propose that the activation of the
nuclear translocation of PCNA may be associated with the induction
of autophagy; however, the potential mechanism underlying the
association between the process of autophagy and PCNA expression
has not yet been elucidated in the context of fracture injury and
healing, although it may involve the self-regulation pathway of the
cell, among others. An inhibitor or enhancer of autophagy, such as
3-methyladenine or rapamycin, respectively, should be used in
further studies to attempt to illustrate the specific role of
autophagy in bone following fracture injury.
In conclusion, this is the first study, to the best
of our knowledge, to report the activation of autophagy in bone
tissue following fracture. This phenomenon may indicate that
autophagy is associated with the occurrence of trauma and could be
used as a target to improve bone fracture healing. Future studies
should investigate the role of autophagy in bone fracture, in order
to facilitate the development of treatments aimed at improving
fracture healing.
References
|
1
|
Megas P: Classification of non-union.
Injury. 36(Suppl 4): S30–S37. 2005.PubMed/NCBI
|
|
2
|
Rueff-Barroso CR, Milagres D, Valle J,
Casimiro-Lopes G, Nogueira-Neto JF, Zanier JF and Porto LC: Bone
healing in rats submitted to weight-bearing and non-weight-bearing
exercises. Med Sci Monit. 14:BR231–BR236. 2008.PubMed/NCBI
|
|
3
|
Mizushima N, Levine B, Cuervo AM and
Klionsky DJ: Autophagy fights disease through cellular
self-digestion. Nature. 451:1069–1075. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Levine B and Kroemer G: Autophagy in the
pathogenesis of disease. Cell. 132:27–42. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Lin Y, Tang C, He H and Duan R: Activation
of mTOR ameliorates fragile X premutation rCGG repeat-mediated
neurodegeneration. PloS One. 8:e625722013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Su H, Li J, Osinska H, Li F, Robbins J,
Liu J, Wei N and Wang X: The COP9 signalosome is required for
autophagy, proteasome-mediated proteolysis, and cardiomyocyte
survival in adult mice. Circ Heart Fail. 6:1049–1057. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Caramés B, Hasegaa A, Taniguchi N, Miyaki
S, Blanco FJ and Lotz M: Autophagy activation by rapamycin reduces
severity of experimental osteoarthritis. Ann Rheum Dis. 71:575–581.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Vellai T: Autophagy genes and ageing. Cell
Death Differ. 16:94–102. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Klionsky DJ and Emr SD: Autophagy as a
regulated pathway of cellular degradation. Science. 290:1717–1721.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Thorburn A: Apoptosis and autophagy:
Regulatory connections between two supposedly different processes.
Apoptosis. 13:1–9. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Bao XH, Naomoto Y, Hao HF, Watanabe N,
Sakurama K, Noma K, Motoki T, Tomono Y, Fukazawa T, Shirakawa Y, et
al: Autophagy: Can it become a potential therapeutic target? Int J
Mol Med. 25:493–503. 2010.PubMed/NCBI
|
|
12
|
Beau I, Mehrpour M and Codogno P:
Autophagosomes and human diseases. Int J Biochem Cell Biol.
43:460–464. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Mijaljica D, Prescott M and Devenish RJ:
Autophagy in disease. Methods Mol Biol. 648:79–92. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhang L, Guo YF, Liu YZ, Liu YJ, Xiong DH,
Liu XG, Wang L, Yang TL, Lei SF, Guo Y, et al: Pathway-based
genome-wide association analysis identified the importance of
regulation-of-autophagy pathway for ultradistal radius BMD. J Bone
Miner Res. 25:1572–1580. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
15
|
Daroszewska A, van't Hof RJ, Rojas JA,
Layfield R, Landao-Basonga E, Rose L, Rose K and Ralston SH: A
point mutation in the ubiquitin-associated domain of SQSMT1 is
sufficient to cause a Paget's disease-like disorder in mice. Hum
Mol Genet. 20:2734–2744. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kabeya Y, Mizushima N, Ueno T, Yamamoto A,
Kirisako T, Noda T, Kominami E, Ohsumi Y and Yoshimori T: LC3, a
mammalian homologue of yeast Apg8p, is localized in autophagosome
membranes after processing. EMBO J. 19:5720–5728. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Su JC, Tseng PH, Hsu CY, Tai WT, Huang JW,
Ko CH, Lin MW, Liu CY, Chen KF and Shiau CW: RFX1-dependent
activation of SHP-1 induces autophagy by a novel obatoclax
derivative in hepatocellular carcinoma cells. Oncotarget.
5:4909–4919. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Manolagas SC: Birth and death of bone
cells: Basic regulatory mechanisms and implications for the
pathogenesis and treatment of osteoporosis. Endocr Rev. 21:115–137.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Lee FY, Choi YN, Beherns FF, Defoun DO and
Einhorn TA: Programmed removal of chondrocytes during endochondral
fracture healing. J Orthop Res. 16:144–150. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Einhorn TA: The cell and molecular biology
of fracture dealing. Clin Orthop Relat Res. (355 Suppl): S7–S21.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Holstein JH, Menger MD, Culemann U, Meier
C and Pohlemann T: Development of a locking femur nail for mice. J
Biomech. 40:215–219. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Müller E, Koch P, Nazarian S and Schatzker
J: The Comprehensive Classification of Fractures of Long Bones
(1st). New York, NY: Springer. 1994.
|
|
23
|
Hou H, Zhang L, Zhang L, Liu D, Xiong Q,
Du H and Tang P: Acute spinal cord injury could cause activation of
autophagy in dorsal root ganglia. Spinal Cord. 51:679–682. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Moscat J and Diaz-Meco MT: p62 at the
crossroads of autophagy, apoptosis and cancer. Cell. 137:1001–1004.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Dosenko VE, Nagibin VS, Tumanovska LV and
Moibenko AA: Protective effect of autophagy in anoxia-reoxygenation
of isolated cardiomyocyte? Autophagy. 2:305–306. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Mortimore GE and Schworer CM: Induction of
autophagy by amino-acid deprivation in perfused rat liver. Nature.
270:174–176. 1977. View
Article : Google Scholar : PubMed/NCBI
|
|
27
|
Quan W, Jung HS and Lee MS: Role of
autophagy in the progression from obesity to diabetes and in the
control of energy balance. Arch Pharm Res. 36:223–229. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Barnouti ZP, Owtad P, Shen G, Petocz P and
Darendeliler MA: The biological mechanisms of PCNA and BMP in TMJ
adaptive remodeling. Angle Orthod. 81:91–99. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Iwaki A, Jingushi S, Oda Y, Izumi T, Shida
JI, Tsuneyoshi M and Sugioka Y: Localization and quantification of
proliferating cells during rat fracture repair: Detection of
proliferating cell nuclear antigen by immunohistochemistry. J Bone
Miner Res. 12:96–102. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Morrow PW, Tung HY and Hemmings HC Jr:
Rapamycin causes activation of protein phosphatase-2A1 and nuclear
translocation of PCNA in CD4+ T cells. Biochem Biophys Res Commun.
323:645–651. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kobayashi S, Kishimoto T, Kamata S, Otsuka
M, Miyazaki M and Ishikura H: Rapamycin, a specific inhibitor of
the mammalian target of rapamycin, suppresses lymphangiogenesis and
lymphatic metastasis. Cancer Sci. 98:726–733. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Kawahara T, Asthana S and Kneteman NM:
m-TOR inhibitors: What role in liver transplantation? J Hepatol.
55:1441–1451. 2011. View Article : Google Scholar : PubMed/NCBI
|