Introduction
The rate of emergence of drug-resistant strains has
been increasing in recent years owing to the widespread and
inappropriate use of conventional antibiotics. This is significant
threat to human health (1), and the
development of novel antimicrobial agents that are effective
against antibiotic-resistance microbes is urgently required
(2). Antimicrobial peptides (AMPs)
are important components of the innate host defense system and are
present in virtually all species of life. AMPs have broad spectrum
activity against bacteria, fungi, viruses, parasites and even
tumors, and have exhibited a similar efficiency against strains
that are resistant and susceptible to antibiotics (3). Since they exert nonspecific
membrane-lytic activities, it is extremely difficult for bacteria
to develop resistance to AMPs. Therefore, AMPs have been regarded
with increasing interest for development as novel therapeutic
agents for the control and prevention of infectious diseases,
particularly those caused by antibiotic-resistant bacteria
(4).
Among the AMPs, cathelicidins are one of the most
prominent families of host defence peptides and are recognized as
having a role in the defence against microbial invasion (5). The majority of cathelicidins are
differentially expressed in a variety of tissues and exhibit
broad-spectrum antimicrobial activity against gram-negative and
gram-positive bacteria, fungi, protozoa and enveloped viruses
(6,7). Fowlicidin-2 is a novel member of the
cathelicidins family that was identified in chicken (8). The peptide has previously been shown to
exert highly potent antibacterial and lipopolysaccharide
(LPS)-neutralizing activities (8,9), and is
among the most efficacious cathelicidins to have been reported
(8). Due to its broad-spectrum and
salt-insensitive antibacterial activities, as well as its potent
LPS-neutralizing activity (8),
fowlicidin-2 may be an excellent candidate as a novel antimicrobial
and antisepsis agent.
There are three pathways for AMP production:
Extraction from natural resources, chemical synthesis and
recombinant expression. Due to the complexity, low-yield and high
cost of chemical peptide synthesis (10), genetic engineering technology using
microorganisms as host cells to express AMPs may be a more
efficient method for AMP production. The methylotrophic yeast
Pichia pastoris has become widely used as a heterologous
gene expression system (11). Yeasts
have become increasingly important in genetic engineering due to
the ease by which they may be genetically manipulated, their
ability to grow rapidly in inexpensive medium to high cell
densities and their capability for complex post-translational
modifications, including folding, disulfide bridge formation and
glycosylation (12,13). The slow production of fowlicidin-2
has been, to date, a bottleneck for its development in scientific
and industrial research. In our previous study, fowlicidin-2 was
expressed in Escherichia coli (14); however, the yield (~6 mg/l) and the
cost of production requires further improvement to meet the needs
of industrial production. In the present study, fowlicidin-2 was
recombinantly expressed using a P. pastoris expression
system, and the bioactivity of the recombinant peptide was
investigated in vitro.
Materials and methods
Strains, vectors, reagents and
enzymes
The expression host, P. pastoris X-33, and
the pPICZα-A expression vector were purchased from Thermo Fisher
Scientific, Inc. (Waltham, MA, USA). E. coli DH5α, E.
coli 25922, Staphylococcus aureus 29213, Pseudomonas
aeruginosa 27853 and Salmonella typhimurium C77-31 were
preserved in our laboratory. A375 human malignant melanoma cells
and RAW264.7 mice macrophages were purchased from Type Culture
Collection of the Chinese Academy of Sciences (Shanghai, China).
Chick embryo fibroblasts (CEFs) were isolated from 9-day-old
chicken embryos, provided by the Harbin Veterinary Research
Institute, Chinese Academy of Agricultural Sciences (Harbin,
China). The experimental protocol was reviewed and approved by the
ethics committee of the Hospital of Northeast Agricultural
University (NEAU; Harbin, China).
Construction of the expression
vector
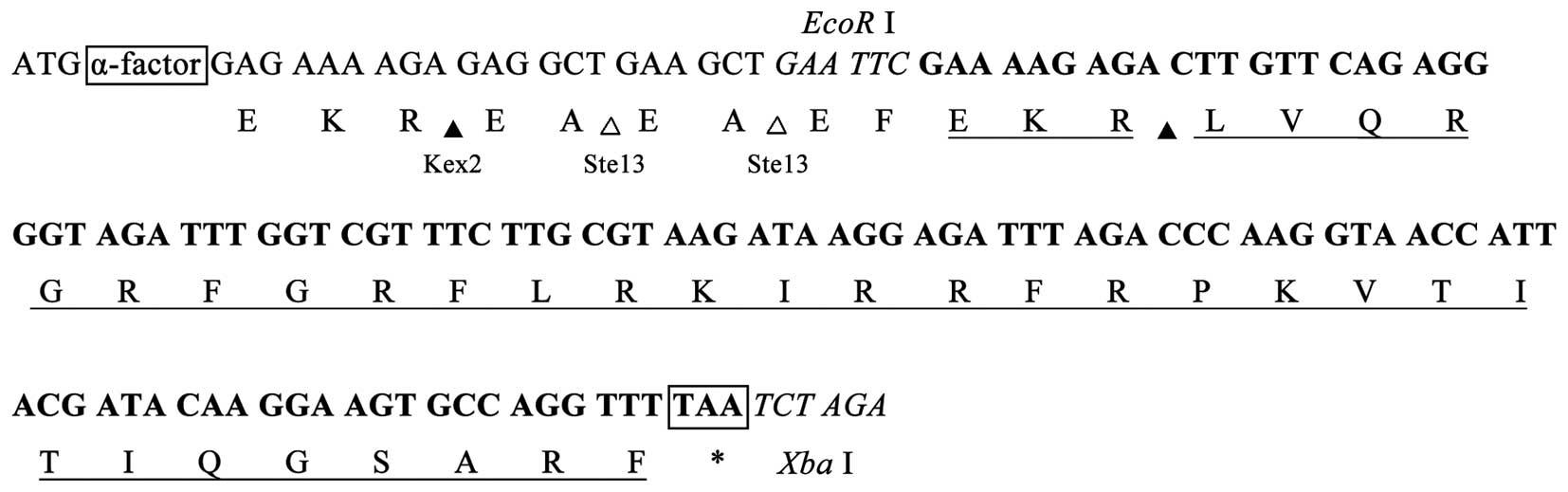
A DNA fragment encoding the mature peptide of
fowlicidin-2 was designed, using DNA software (version 2.0;
http://www.dna20.com) according to the codon bias
of the methylotrophic yeast P. pastoris. The fragment was
constructed using two complementary oligonucleotides (oligos), as
follows: Sense, 5′-GAATTC
GAAAAGAGACTTGTTCAGAGGGGTAGATTTGGTCGTTTCTTGCGTAAGATAAGGAGATTTAGACCCAAGGTAACCATTACGATACAAGGA
A G T G C C A G G T T T TAA T C T AGA-3′ and
antisense, 5′-TCTAGA
TTAAAACCTGGCACTTCCTTGTATCGTAATGGTTACCTTGGGTCTAAATCTCCTTATCTTACGCAAGAAACGACCAAATCTACCCCTCTGAACAAGTCTCTTTTC
GAATTC-3′. Restriction sites for EcoRI and
XbaI are indicated in the oligos in italics. A terminal TAA
codon is followed by the sequence encoding fowlicidin-2 in the
sense strand. GAAAAGAGA, as the codon for a glutamic acid, lysine
and arginine (EKR) sequence, provide the cleavage site of Kexin
protease 2 (Kex2) in the translated peptide (Fig. 1). The two complementary oligos were
synthesized chemically by Sangon Biotech Co., Ltd. (Shanghai,
China). A total of 20 µl of each sense and antisense oligos
solution (100 µm) were mixed, heated at 95°C for 2 min and cooled
down to room temperature. The annealed product was purified using
the gel extraction kit (cat. no. 28704; Qiagen, Beijing, China).
Following digestion by EcoRI and XbaI, the annealed
product was ligated using a DNA Ligation kit (cat. no. 6022;
version 2.1; Takara Biotechnology Co., Ltd., Dalian, China)
according to the manufacturer's instruction between the
EcoRI and XbaI sites of the pPICZα-A vector, which
contains the Saccharomyces cerevisiae α-factor secretion
signal and the alcohol oxidase 1 promoter. A total of 10 µl of the
resultant plasmid (30 ng) was added to 100 µl of competent E.
coli DH5α cells (Tiangen Biotech Co., Ltd., Beijing, China) and
held on ice for 30 min prior to heat shocking at 42°C for 90 sec.
After incubation on ice for 2 min, the cells were diluted into 0.8
ml LB culture (0.5% yeast extract, 1% tryptone, 1% NaCl) (Hopebio,
Qindao, China) and cultured at 37°C for 30 min. Then, appropriate
dilutions were plated on LB plates (containing 20 µg/ml Zeocin) and
incubated for 12 h at 37°C (15).
Positive transformants were screened by colony polymerase chain
reaction (PCR) using a Taq PCR Mastermix kit (cat. no.
KT201; Qiagen) and the following primers: P1,
5′-GACTGGTTCCAATTGACAAGC-3′ and P2, 5′-GCAAATGGCATTCTGACATCC-3′.
The following thermal cycling conditions were used: 94°C for 5 min
followed by 32 cycles (95°C for 30 sec, 58°C for 30 sec and 72°C
for 1 min) and finally 72°C for a 5 min extension. Finally, they
were verified by DNA sequencing.
Transformation of P. pastoris and
selection of transformants
The pPICZα-A-fowlicidin-2 construct was linearized
using the SacI restriction enzyme and transformed into P.
pastoris X-33 cells by electroporation using a Gene Pulser
apparatus (Bio-Rad Laboratories, Inc., Hercules, CA, USA) in 0.2-cm
electroporation cuvettes (200 Ω, 25 µF, 1,500 V). The pPICZα-A
vector was similarly linearized and transformed into P.
pastoris X-33 as a negative control. The two types of
transformed P. pastoris were incubated at 30°C in YPD plates
(Hopebio) containing 1% yeast extract, 2% tryptone, 2% dextrose, 2%
agar and 100 µg/ml Zeocin (Invitrogen; Thermo Fisher Scientific,
Inc.). In order to determine the methanol-utilizing phenotype of
the transformants, all Zeocin-resistant transformants cultured in
YPD medium were selected on MD plates (1.34% YNB,
4×10−5% biotin, 2% dextrose and 2% agar) and MM plates
(1.34% YNB, 4×10−5% biotin, 0.5% methanol and 1.5% agar)
(both purchased from Invitrogen; Thermo Fisher Scientific, Inc.).
Zeocin+ Mut+ phenotype colonies were further
confirmed by colony-PCR using the P1 and P2 primers.
Heterologous expression of recombinant
fowlicidin-2 (rfowlicidin-2)
A total of 10 pPICZα-A-fowlicidin-2 transformants
were cultured at 30°C in a shaking flask containing 20 ml buffered
glycerol complex medium (1% yeast extract, 2% peptone, 0.3%
K2HPO4, 1.18% KH2PO4,
1.34% YNB, 4×10−5% biotin, 1% glycerol, pH 6.0) to
optical density (OD)600=2.0–6.0. Cells were harvested by
centrifugation at 1,500 × g for 5 min at room temperature and
resuspended to an OD600 of 1.0 in buffered
methanol-complex medium (BMMY; 1% yeast extract, 2% peptone, 0.3%
K2HPO4, 1.18% KH2PO4,
1.34% YNB, 4×10−5% biotin, 0.5% methanol, pH 6.0). To
induce expression of rfowlicidin-2, methanol was added every 24 h
to a final concentration of 0.5% (v/v), after which the resuspended
culture was grown for 72 h and the supernatant was obtained by
centrifugation at 12,000 × g for 5 min at 4°C. The expression of
fowlicidin-2 in the supernatant was determined using Tricine-sodium
dodecyl sulfate-polyacrylamide gel electrophoresis
(Tricine-SDS-PAGE). Total protein yield in the culture medium was
measured using a Bradford protein assay kit. The highest expressing
P. pastoris clone identified as P. pastoris X33-Fow2
was singled out and used for large-scale production of
rfowlicidin-2.
Purification of rfowlicidin-2
The fermentation medium obtained from the P.
pastoris X33-fowlicidin-2 fermentation was harvested by
centrifugation at 12,000 × g for 10 min at 4°C. A total of 30 ml of
the resultant supernatant was loaded onto a Q-Sepharose Fast Flow
column (10 ml broth/5 ml resin; GE Healthcare Life Sciences,
Chalfont, UK) pre-equilibrated with 20 mM sodium phosphate buffer
(pH 7.2; buffer A) for purification by ion exchange chromatography.
The column was washed again with buffer A, and then eluted with a
linear gradient of 0.0–1.0 M NaCl in buffer A. Fractions containing
fowlicidin-2 were collected and further purified by reversed phase
high-performance liquid chromatography (RP-HPLC) on a C18 column
(250 × 4.6 mm, 5 µm; silica pore size, 300 Å; Supelco, Inc.,
Bellefonte, PA, USA), which had been pre-equilibrated using 0.1%
trifluoroacetic acid (TFA). Bound proteins were eluted with a
linear gradient of acetonitrile (20–45%, v/v; 1.5% per min) in 0.1%
TFA. The flow rate was 1.0 ml/min and the absorbance of the eluent
was monitored at 214 nm. Purification was monitored by 15%
Tricine-SDS-PAGE. The purified peptide was lyophilized and stored
at −20°C until further analysis. The peptide yield was determined
using a Bradford protein assay kit.
Mass spectrometry and N-terminal
sequencing analysis
The purified peptide (30 ng) was dissolved in a
saturated solution of α-cyano-4-hydroxycinnamic acid containing 50%
acetonitrile and 0.1% TFA, and then analyzed using a
Matrix-Assisted Laser Desorption/Ionization Time-of-Flight mass
spectrometer (Applied Biosystems; Thermo Fisher Scientific, Inc.).
For N-terminal sequencing analysis, the purified peptide was run on
a 15% tricine-SDS-polyacrylamide gel, and then transferred to a
polyvinylidene difluoride (PVDF) membrane (EMD Millipore,
Billerica, MA, USA) by semi-dry electrophoretic transfer.
N-terminal sequence analysis was performed by automated Edman
degradation using a PPSQ-33A Sequencing System (Shimadzu
Corporation, Kyoto, Japan), according to the manufacturer's
protocol.
Antibacterial activity assay
Antibacterial activity of fowlicidin-2 against
various microorganisms was tested using the microbroth dilution
method (16). Briefly, bacteria
subcultured in the log phase were diluted to
2×105-7×105 colony forming units/ml with
fresh Mueller Hinton Broth medium (Hopebio). Aliquots (50 µl) from
each strain suspension were distributed into the wells of a 96-well
polypropylene microtiter plate, after which 50 µl peptide was added
to each well. Cultures were grown for 18 h at 37°C with agitation.
The absorbance was measured at 600 nm using a microplate reader.
Experiments were performed in triplicate. The minimal inhibitory
concentration (MIC) was defined as the lowest concentration of
peptide able to fully inhibit bacterial growth, as determined by OD
measurements.
Hemolytic activity assay
The hemolytic activity of rfowlicidin-2 was
determined using human red blood cells (hRBCs), as described
previously (17). Briefly, fresh
anticoagulated human blood from a healthy donor was washed thrice
with phosphate-buffered saline (PBS) and diluted to a concentration
of 4% (v/v) in PBS. The experimental protocol was reviewed and
approved by the ethics committee of the Hospital of NEAU. The
erythrocytes (90 µl) were dispensed into a 96-well plate, followed
by addition in triplicate of 10 µl serially diluted peptides. The
resulting suspension was incubated for 1 h at 37°C. Following
centrifugation of the plate at 1,000 × g for 5 min at room
temperature, the supernatants were transferred to a new 96-well
plate and the absorbance was measured at 570 nm using an ELISA
plate reader for analysis of released hemoglobin. Negative controls
consisted of cells suspended in PBS only, and positive controls
consisted of cells suspended in 1% Triton X-100. The percentage of
hemolysis was calculated as follows: Percentage of hemolysis (%) =
[(A570 nm, peptide - A570 nm, PBS)/(A570
nm, 1% Triton X-100 - A 570 nm, PBS)] × 100.
Determination of anticancer activity
in vitro
The anticancer activity of rfowlicidin-2 was
determined using the
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT)
method, as described previously (18). Briefly, A375 human malignant melanoma
cells, RAW264.7 mice macrophages and CEFs were seeded into a
96-well plate at a density of 2.5×104 cells/well in
Dulbecco's modified Eagle's medium (DMEM; Gibco; Thermo Fisher
Scientific, Inc.) containing 10% fetal bovine serum (FBS; Gibco;
Thermo Fisher Scientific, Inc.) and cultured at 37°C in a
humidified 5% CO2 incubator. When the cell fusion
reached more than 80%, the cells were washed once with DMEM,
followed by the addition of 90 µl fresh DMEM containing 5% FBS and
various concentrations (0 to 128 µM) of 10 µl peptide in triplicate
into each well. Cells were incubated at 37°C for 24 h, after which
10 µl alamarBlue dye was added to each well and incubated for a
further 6 h. The fluorescence was measured at 490 nm. The
anticancer activity of rfowlicidin-2 was defined as the peptide
concentration that induced 50% cell death (EC50).
Statistical analysis
Data were analyzed by one-way analysis of variance
using SPSS version 17.0 software (SPSS, Inc., Chicago, IL, USA).
Differences among groups were compared with Student's t-test.
Results
Screening of positive
transformants
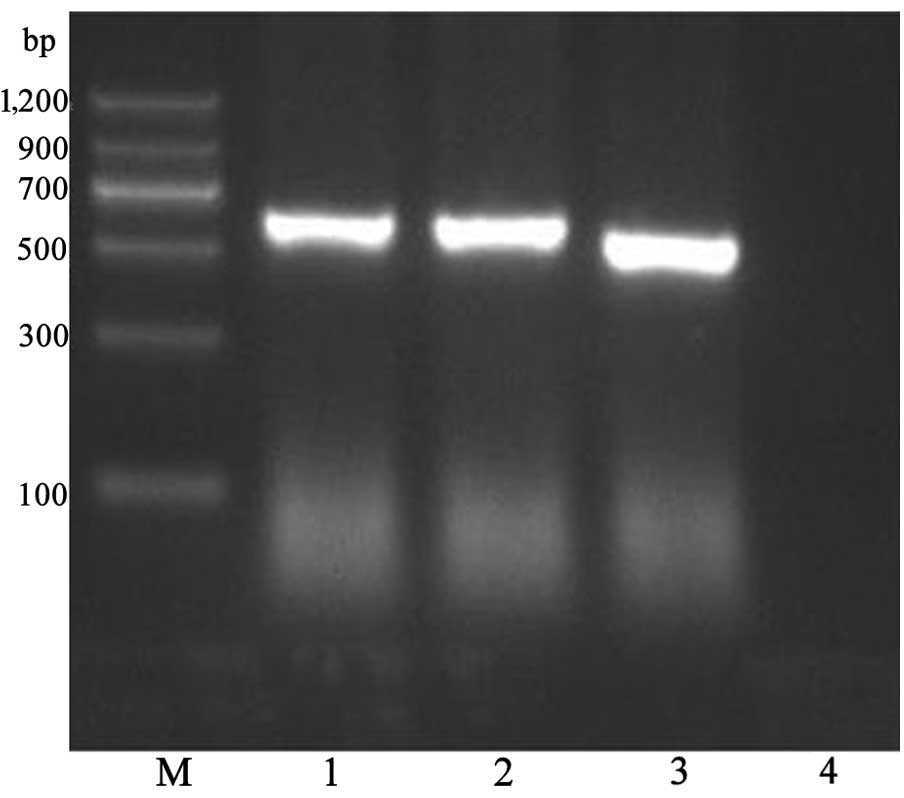
A fowlicidin-2 DNA fragment was inserted into the
EcoRI and XbaI sites of pPICZα-A to construct the
recombinant pPICZα-A-fowlicidin-2 vector. The pPICZα-A-fowlicidin-2
vector was transformed into P. pastoris X-33 cells by
electroporation, and the transformants that grew on MD plates were
identified by PCR (Fig. 2). A total
of 31 multicopy transformants were screened out on MD plates under
selection by Zeocin (100 µg/ml) and were confirmed by PCR
amplification. A total of 10 colonies presenting the
Zeocin+ Mut+ phenotype and with
strong PCR amplification products of fowlicidin-2 DNA were used for
the induced expression of the peptide.
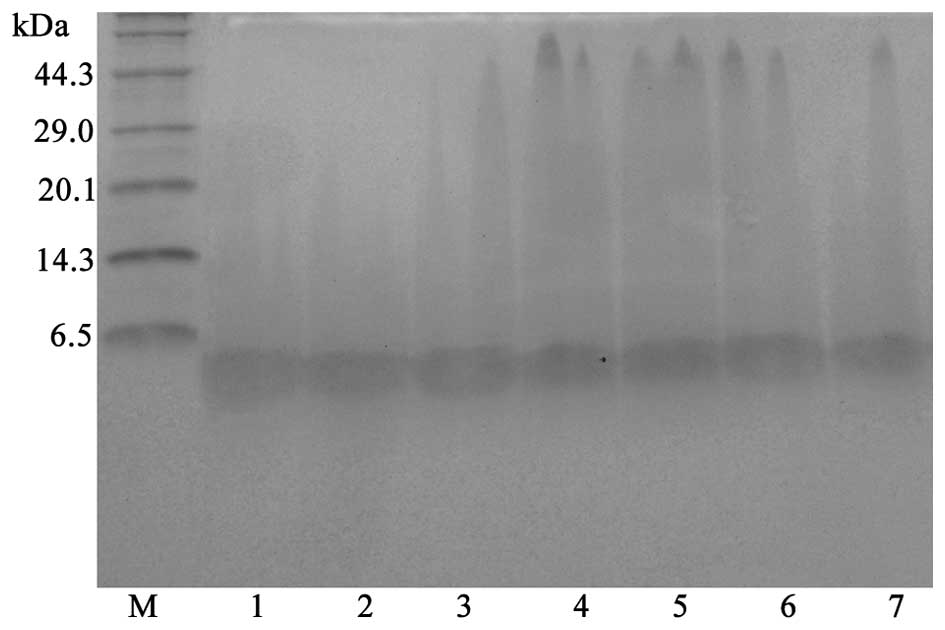
Expression of rfowlicidin-2
pPICZα-A-fowlicidin-2 transformants were analyzed
for the extracellular expression of fowlicidin-2 in BMMY broth at
28°C for 72 h. The highest-expressing transformant, P.
pastoris X33-Fow2, was selected for further high-density
cultivation. Aliquots of the culture supernatant were withdrawn at
12-h intervals and analyzed by Tricine-SDS-PAGE. As shown in
Fig. 3, a major protein band of ~3.7
kDa, which corresponded to fowlicidin-2, appeared following
induction with methanol. The highest expression level of
fowlicidin-2 was ~121 mg/l at 60 h of culture in the optimized
fermentation conditions (28°C, pH 6.0, 0.5% methanol in BMMY
medium).
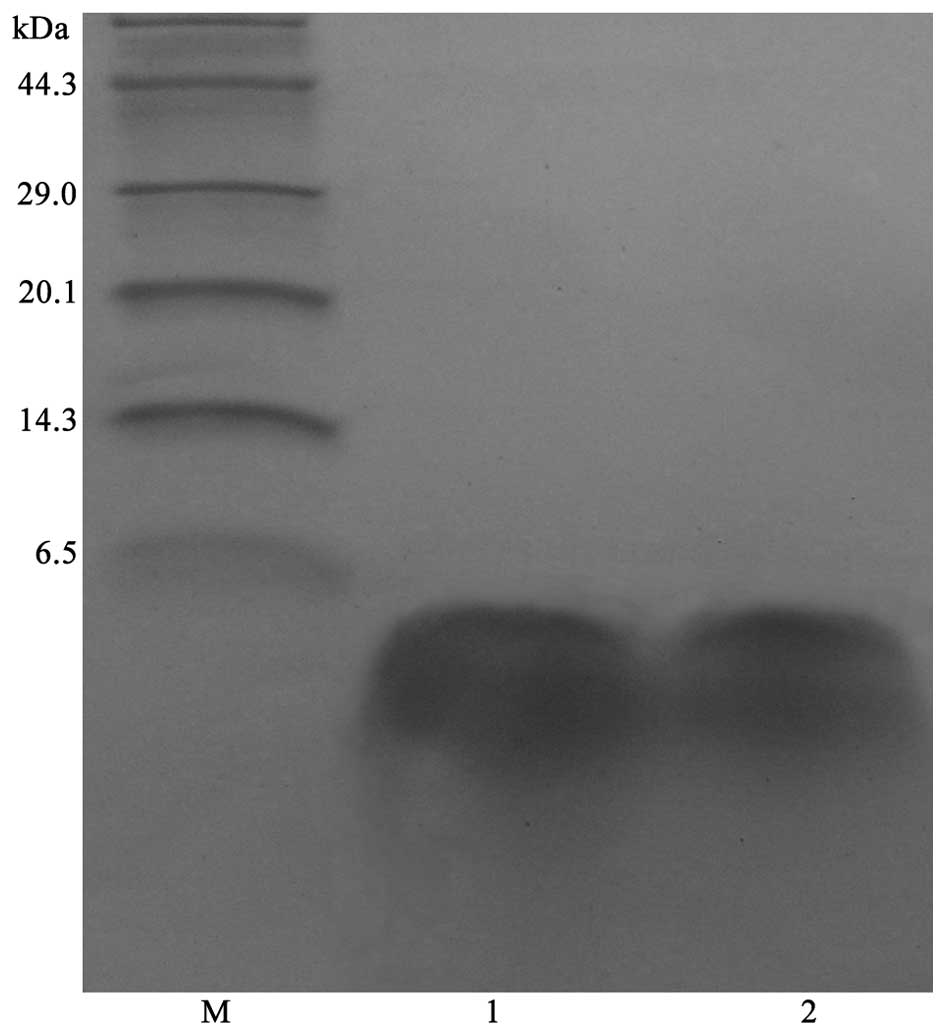
Purification and identification of
rfowlicidin-2
After 60 h of methanol induction, the cultures were
harvested and the supernatants were collected for the purification
of rfowlicidin-2 using a Q-Sepharose Fast Flow column and RP-HPLC.
The highest peak absorbance fractions, which were pooled and
detected as a single band by Tricine-SDS-PAGE (Fig. 4), contained the majority of
rfowlicidin-2. The average volume of rfowlicidin-2 recovered from 1
liter culture medium was ~85.6 mg, with a purity of >95%. These
results indicate that rfowlicidin-2 may be successfully purified
from methylotrophic yeast, with a relatively high recovery rate and
purity, using a two-step purification strategy. The purified
rfowlicidin-2 was transferred to a PVDF membrane, and sequenced for
up to 4 cycles. The N-terminus began with a
leucine-valine-glutamine-arginine sequence, which was consistent
with the predicted sequence of rfowlicidin-2 (8). This indicates that the rfowlicidin-2
peptide that was produced did not have any additional amino acids
at its N-terminal end.
Antibacterial activity of
rfowlicidin-2
The MICs of the peptide for several microorganisms
are shown in Table I. Fowlicidin-2
showed strong and broad-spectrum antimicrobial activities against
both gram-positive bacteria (S. aureus ATCC 29213) and
gram-negative bacteria (E. coli 25922, P. aeruginosa
ATCC 27853, S. typhimurium C77-31). For these strains, the
MIC values were in the range of 1–4 µM.
 | Table I.MICs of rfowlicidin-2 for four tested
bacteria. |
Table I.
MICs of rfowlicidin-2 for four tested
bacteria.
| Tested
bacteria | MIC (µM) |
|---|
| Escherichia
coli ATCC25922 | 1 |
| Staphylococcus
aureus ATCC29213 | 4 |
| Pseudomonas
aeruginosa ATCC27853 | 4 |
| Salmonella
typhimurium C77-31 | 2 |
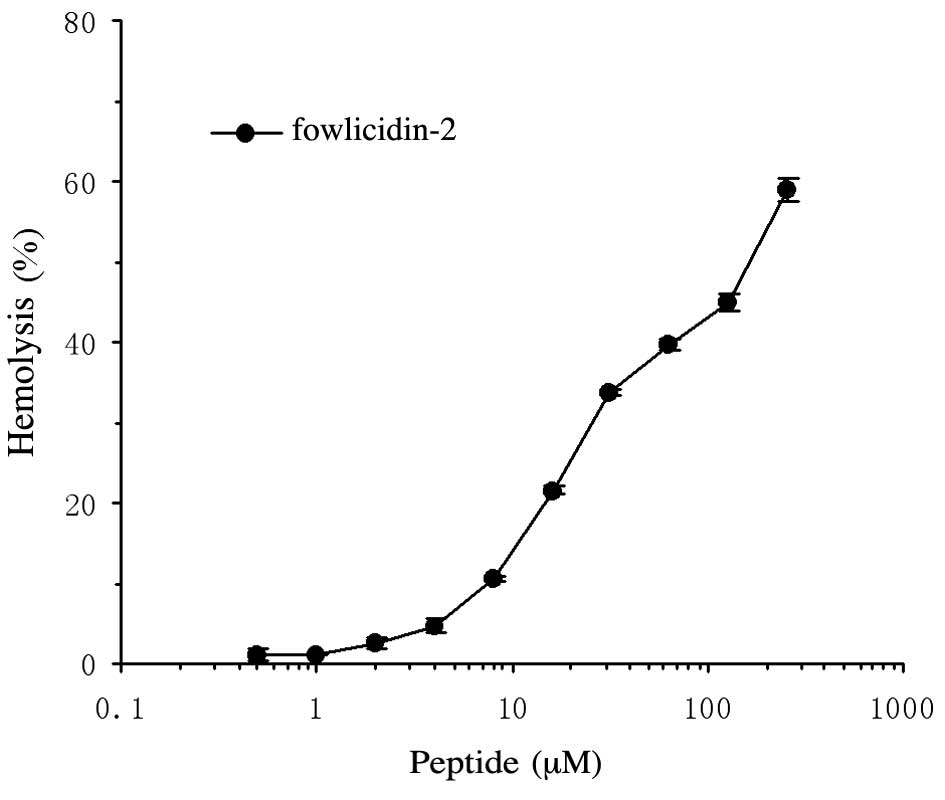
Hemolytic activities of
rfowlicidin-2
To evaluate the hemolytic activity of fowlicidin-2
against red blood cells, fresh hRBCs were used. The peptide
exhibited a dose-dependent hemolytic activity against hRBCs, with
the EC50 occurring within the range of 128–256 µM
(Fig. 5).
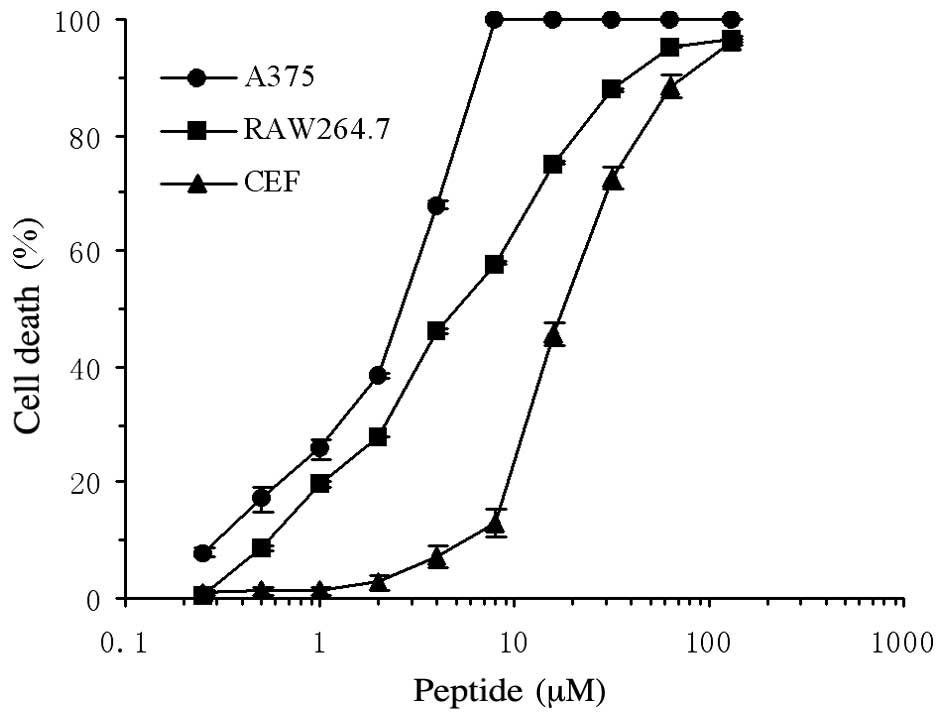
Anticancer activity of
rfowlicidin-2
The cytotoxic activity of rfowlicidin-2 was assessed
against A375 human malignant melanoma cells, RAW264.7 mouse
macrophages and CEFs. Concentration-response studies demonstrated
that the peptide exerted a significantly higher cytotoxic activity
against the human malignant A375 cells (P<0.01), as compared
with the two other types of cells (Fig.
6). Fowlicidin-2 showed a 2-fold higher activity against the
human A375 cells (EC50=2–4 µM), as compared with the
RAW264.7 cells (EC50=4–8 µM), and an 8-fold higher
activity, as compared with the CEFs (EC50=16–32 µM;
Fig. 6).
Discussion
The emergence of pathogenic microorganisms with
resistance to conventional antibiotics has driven the search for
novel types of antimicrobial agents. Although numerous novel
strategies have been investigated, there are a number of avenues
that have yet to be explored (19).
AMPs are promising candidates for the development of novel
antimicrobial agents, since they have been shown to be an important
component of immunity and have a broad antimicrobial spectrum
(3). However, the production of
active AMPs in a cost-effective and scalable method is challenging.
It has been recognized that recombinant DNA techniques are a
potential means for producing AMPs in large quantities. Numerous
AMPs have been successfully produced using genetic engineering
methods (10). Various host cells
have been used for the expression of AMPs; however, P.
pastoris has emerged as one of the most efficient recombinant
bioreactors (20). Similar to
prokaryotes, P. pastoris is able to grow rapidly in
inexpensive medium, and is amenable to high-cell-density
fermentations (20,21). Furthermore, it is non-pathogenic and
is capable of secreting functional extracellular proteins directly
into the culture medium (21,22).
The present study used P. pastoris to express
fowlicidin-2 at a high level. The Kex2 recognition site was
introduced at the N-terminal of fowlicidin-2. The Kex2 endoprotease
is a member of the membrane-bound endoprotease family and is
responsible for site-specific endoproteolysis at pairs of basic
amino acid residues, in particular EKR, in the α-factor precursor
peptide (23,24). The translated peptide fowlicidin-2
was successfully released by Kex2, and was shown to have the
predicted amino acid sequence and molecular weight. Furthermore,
sequencing revealed that there were no additional residues at the
N-terminus of the peptide following Kex2 digestion, which was
important since the natural N-terminus is critical for the activity
of AMPs (25,26).
The antibacterial assay in the present study
indicates that rfowlicidin-2 is prominent among the most potent
cathelicidins discovered to date, showing activity against a
variety of bacteria at concentrations of ≤4 µM. In the present
study, the lowest detected MIC of rfowlicidin-2 was towards E.
coli 25922 (1 µM). The peptide exhibited a similar activity
against S. aureus ATCC29213 and P. aeruginosa
ATCC27853. These results demonstrate that the rfowlicidin-2 peptide
was functional and effective against gram-negative and
gram-positive bacteria. In addition, rfowlicidin-2 was shown to
exert a hemolytic effect on hRBCs, with EC50 occurring
at 128–256 µM, which is markedly higher than that reported by Xiao
et al (8). The underlying
mechanism is unclear and may be due to the different spatial
structures of the rfowlicidin-2 obtained in the present study and
the chemically synthesized fowlicidin-2 used in the study of Xiao
et al (8).
Several cationic amphipathic AMPs or their
derivatives have previously been reported to exert anticancer
activity, display selective cytotoxicity against cancer cells in
vitro and show efficacy in various in vivo xenograft
models (27,28). However, only a small range of AMPs
have been shown to have anticancer activity. Various
cathelicidin-related or cathelicidin-modified AMPs, including LL-37
(29), Ceragenin CSA-13 (30), FK-16 (31) and BF-30 (32), have been reported to exhibit
anticancer activity. However, to the best of our knowledge, there
have been no previous reports on the anticancer activity of
fowlicidin-2. Cationicity, α-helicity and amphipathicity are among
the most important physicochemical parameters that determine the
functional properties of α-helical AMPs (9,33).
Increasing the degree of α-helicity and mean hydrophobicity of AMPs
generally result in an increase in cytotoxicity against cancer
cells (34). The secondary structure
of fowlicidin-2 is dominated by α-helices. The N-terminal α-helix
of the peptide adopts a typical amphipathic structure, whereas the
C-terminal helix is more hydrophobic (9). In the present study, the anticancer
activity of fowlicidin-2 was determined using the MTT method in
vitro. The survival of cells was reduced by treatment with
fowlicidin-2, and the percentage of cell death increased in a
concentration-dependent manner. However, the EC50 of
fowlicidin-2 for A375 human cancer cells was 2–4 µM, and few A375
cells survived when treated with 8 µM fowlicidin-2. Conversely,
fowlicidin-2 showed relatively weak cytotoxic activity against
RAW264.7 cells (EC50=4–8 µM) and CEFs
(EC50=16–32 µM). These results suggested that
fowlicidin-2 may be considered a promising candidate for anticancer
applications. However, the mechanism of action of the peptide on
cancer cells requires further investigation.
In conclusion, in the present study bioactive
fowlicidin-2 was successfully expressed at a high level (85.6 mg/l)
and high purity (>95%) using the yeast P. pastoris as a
recombinant expression system. In addition, rfowlicidin-2 showed
strong and broad-spectrum antimicrobial activity and anticancer
activity. The present study was the first to demonstrate an
inhibitory effect of fowlicidin-2 on cancer cells in
vitro.
Acknowledgements
The present study was supported by the Program for
Young Aged Academic Staff in the Heilongjiang Province Ordinary
College (grant no. 1252G010)U, the Research Fund for Innovation
Talents of Science and Technology in Harbin City (grant no.
2012RFQXN022) and the ‘Academic Backbone’ Project of Northeast
Agricultural University (grant no. 15XG15).
References
|
1
|
Rai J, Randhawa GK and Kaur M: Recent
advances in antibacterial drugs. Int J Appl Basic Med Res. 3:3–10.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Spížek J, Novotná J, Rezanka T and Demain
AL: Do we need new antibiotics? The search for new targets and new
compounds. J Ind Microbiol Biotechnol. 37:1241–1248. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Zasloff M: Antimicrobial peptides of
multicellular organisms. Nature. 415:389–395. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hancock RE and Sahl HG: Antimicrobial and
host-defense peptides as new anti-infective therapeutic strategies.
Nat Biotechnol. 24:1551–1557. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
Tomasinsig L and Zanetti M: The
cathelicidins - structure, function and evolution. Curr Protein
Pept Sci. 6:23–34. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zaiou M and Gallo RL: Cathelicidins,
essential gene-encoded mammalian antibiotics. J Mol Med (Berl).
80:549–561. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zaiou M, Nizet V and Gallo RL:
Antimicrobial and protease inhibitory functions of the human
cathelicidin (hCAP18/LL-37) prosequence. J Invest Dermatol.
120:810–816. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Xiao Y, Cai Y, Bommineni YR, Fernando SC,
Prakash O, Gilliland SE and Zhang G: Identification and functional
characterization of three chicken cathelicidins with potent
antimicrobial activity. J Biol Chem. 281:2858–2867. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Xiao Y, Herrera AI, Bommineni YR, Soulages
JL, Prakash O and Zhang G: The central kink region of fowlicidin-2,
an alpha-helical host defense peptide, is critically involved in
bacterial killing and endotoxin neutralization. J Innate Immun.
1:268–280. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Parachin NS, Mulder KC, Viana AA, Dias SC
and Franco OL: Expression systems for heterologous production of
antimicrobial peptides. Peptides. 38:446–456. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ahmad M, Hirz M, Pichler H and Schwab H:
Protein expression in Pichia pastoris: Recent achievements and
perspectives for heterologous protein production. Appl Microbiol
Biotechnol. 98:5301–5317. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Gellissen G, Kunze G, Gaillardin C, Cregg
JM, Berardi E, Veenhuis M and van der Klei I: New yeast expression
platforms based on methylotrophic Hansenula polymorpha and Pichia
pastoris and on dimorphic Arxula adeninivorans and Yarrowia
lipolytica -A comparison. FEMS Yeast Res. 5:1079–1096. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Hélène B, Céline L, Patrick C, Fabien R,
Christine V, Yves C and Guy M: High-level secretory production of
recombinant porcine follicle-stimulating hormone by Pichia
pastoris. Process Biochem. 36:907–913. 2001. View Article : Google Scholar
|
|
14
|
Feng X, Xu W, Qu P, Li X, Xing L, Liu D,
Jiao J, Wang J, Li Z and Liu C: High-yield recombinant expression
of the chicken antimicrobial peptide fowlicidin-2 in Escherichia
coli. Biotechnol Prog. 31:369–374. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Sambrook J, Fristsch EF and Maniatis T:
Molecular cloning: A laboratory manualSambrook J: Science Press;
Beijing: pp. 87–102. 2002, (In Chinese).
|
|
16
|
Chen Z, Wang D, Cong Y, Wang J, Zhu J,
Yang J, Hu Z, Hu X, Tan Y, Hu F and Rao X: Recombinant
antimicrobial peptide hPAB-β expressed in Pichia pastoris, a
potential agent active against methicillin-resistant Staphylococcus
aureus. Appl Microbiol Biotechnol. 89:281–291. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Dean SN, Bishop BM and van Hoek ML:
Natural and synthetic cathelicidin peptides with anti-microbial and
anti-biofilm activity against Staphylococcus aureus. BMC Microbiol.
11:1142011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Lin MC, Hui CF, Chen JY and Wu JL: The
antimicrobial peptide, shrimp anti-lipopolysaccharide factor
(SALF), inhibits proinflammatory cytokine expressions through the
MAPK and NF-κB pathways in Trichomonas vaginalis adherent to HeLa
cells. Peptides. 38:197–207. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Moellering RC Jr: New approaches to
developing antimicrobials for resistant bacteria. J Infect
Chemother. 9:8–11. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Jin F, Xu X, Zhang W and Gu D: Expression
and characterization of a housefly cecropin gene in the
methylotrophic yeast, Pichia pastoris. Protein Expr Purif.
49:39–46. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Cereghino JL and Cregg JM: Heterologous
protein expression in the methylotrophic yeast Pichia pastoris.
FEMS Microbiol Rev. 24:45–66. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wang A, Wang S, Shen M, Chen F, Zou Z, Ran
X, Cheng T, Su Y and Wang J: High level expression and purification
of bioactive human alpha-defensin 5 mature peptide in Pichia
pastoris. Appl Microbiol Biotechnol. 84:877–884. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Fuller RS, Brake A and Thorner J: Yeast
prohormone processing enzyme (KEX2 gene product) is a
Ca2+-dependent serine protease. Proc Natl Acad Sci USA.
86:1434–1438. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Seeboth PG and Heim J: In-vitro processing
of yeast alpha-factor leader fusion proteins using a soluble yscF
(Kex2) variant. Appl Microbiol Biotechnol. 35:771–776. 1991.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Li L, Wang JX, Zhao XF, Kang CJ, Liu N,
Xiang JH, Li FH, Sueda S and Kondo H: High level expression,
purification, and characterization of the shrimp antimicrobial
peptide, Ch-penaeidin, in Pichia pastoris. Protein Expr Purif.
392:144–151. 2005. View Article : Google Scholar
|
|
26
|
Wu M and Hancock RE: Interaction of the
cyclic antimicrobial cationic peptide bactenecin with the outer and
cytoplasmic membrane. J Biol Chem. 274:29–35. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Huang W, Seo J, Willingham SB, Czyzewski
AM, Gonzalgo ML, Weissman IL and Barron AE: Learning from
host-defense peptides: Cationic, amphipathic peptoids with potent
anticancer activity. PLoS One. 9:e903972014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Yang QZ, Wang C, Lang L, Zhou Y, Wang H
and Shang DJ: Design of potent, non-toxic anticancer peptides based
on the structure of the antimicrobial peptide, temporin-1CEa. Arch
Pharm Res. 36:1302–1310. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Büchau AS, Morizane S, Trowbridge J,
Schauber J, Kotol P, Bui JD and Gallo RL: The host defense peptide
cathelicidin is required for NK cell-mediated suppression of tumor
growth. J Immunol. 184:369–378. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Kuroda K, Fukuda T, Okumura K, Yoneyama H,
Isogai H, Savage PB and Isogai E: Ceragenin CSA-13 induces cell
cycle arrest and antiproliferative effects in wild-type and p53
null mutant HCT116 colon cancer cells. Anticancer Drugs.
24:826–834. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Ren SX, Shen J, Cheng AS, Lu L, Chan RL,
Li ZJ, Wang XJ, Wong CC, Zhang L, Ng SS, et al: FK-16 derived from
the anticancer peptide LL-37 induces caspase-independent apoptosis
and autophagic cell death in colon cancer cells. PLoS One.
8:e636412013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Wang H, Ke M, Tian Y, Wang J, Li B, Wang
Y, Dou J and Zhou C: BF-30 selectively inhibits melanoma cell
proliferation via cytoplasmic membrane permeabilization and
DNA-binding in vitro and in B16F10-bearing mice. Eur J Pharmacol.
707:1–10. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Conlon JM, Mechkarska M, Prajeep M, Arafat
K, Zaric M, Lukic ML and Attoub S: Transformation of the naturally
occurring frog skin peptide, alyteserin-2a into a potent, non-toxic
anti-cancer agent. Amino Acids. 44:715–723. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Gaspar D, Veiga AS and Castanho MA: From
antimicrobial to anticancer peptides. A review. Front Microbiol.
4:2942013. View Article : Google Scholar : PubMed/NCBI
|