Introduction
Galanin is a 29-amino acid peptide naturally present
in the tissues and fluids of humans and animals (1). To date three galanin receptors (GalR1,
GalR2 and GalR3) have been discovered to be widely distributed in
the mammalian central and peripheral nervous system (2). It is acknowledged that galanin has a
critical role in the regulation of energy homeostasis (3). Leptin, another adipocyte-derived
hormone, is also a key factor in the regulation of body weight and
energy expenditure and acts in rodents via hypothalamus receptors
to inhibit feeding and increase thermogenesis (4). Notably, a previous study confirmed that
galanin inhibits leptin expression and secretion in rat adipose
tissue and 3T3-L1 adipocytes (5).
Leptin has been shown to possess direct profibrogenic activity in
the liver, and the absence of leptin is associated with a marked
attenuation of the hepatic response to a diverse range of fibrotic
stimuli (6). It has been
hypothesized that leptin acts directly on hepatic stellate cells
(HSCs) to trigger downstream response pathways that ultimately lead
to the deposition of extracellular matrix (ECM) (7). However, little is known about the
effects of galanin on HSCs. Since the activation and proliferation
of HSCs are the key factors in the progress of liver fibrosis, in
view of the counteracting effect of galanin on leptin in food
intake and energy metabolism, the effect of galanin on the
proliferation of HSCs becomes an interesting research topic.
It may be hypothesized that galanin is able to
inhibit the activation and proliferation of HSCs. In the present
study, to test this hypothesis, the galanin and GalR mRNA
expression in quiescent and activated HSCs was first examined. The
effects of galanin and GalRs on the proliferation and secretion of
HSCs were also explored.
Materials and methods
Animals
A total of 3 male Sprague-Dawley rats (230–300 g;
7–8 weeks-old) were obtained from the Shanghai Experimental Animal
Center (Shanghai, China). All rats were maintained in cages under
pathogen-free condition on a 12-hour light/12-hour dark schedule,
and had ad libitum access to water and food. All animal
handling and experimental procedures were approved by the Animal
Care and Use Committee of the Shanghai Jiaotong University School
of Medicine (Shanghai, China).
Primary HSC isolation
HSCs were isolated from the male Sprague-Dawley rats
(375–400 g) as described previously (8). Briefly, the rat liver was perfused in
situ with D-Hanks buffer for 5 min, 0.02% pronase for 5 min and
0.04% collagenase (type IV) for 10 min at a flow rate of 5 ml/min.
Following in situ digestion, the liver was removed, minced
and further digested with 0.08% collagenase at 37°C for 30 min. The
cell suspension was carefully layered on top of two layers (6% and
10%) of OptiPrep Density Gradient Medium (Sigma-Aldrich; Merck
Millipore, Darmstadt, Germany). After centrifugation at 1400 × g
for 20 min, the cells from the interface were collected and were
used as primary HSCs. Isolated HSCs were suspended in Dulbecco's
modified Eagle's medium (DMEM; Sigma-Aldrich; Merck Millipore)
supplemented with 10% fetal bovine serum (FBS; Gibco; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) (10% FBS/DMEM), 100 IU/ml
penicillin and 100 mg/ml streptomycin (all Gibco), and were plated
at 2×105 cells/cm2 on non-coated 6-well
plastic plates (Costar; Corning Incorporated, Corning, NY, USA).
HSCs were cultured for 7 days in vitro and were used as
fully activated HSCs. Cell purity was 95% as determined by
examination of morphology and vitamin A autofluorescence.
Reverse transcription-polymerase chain
reaction (RT-PCR)
Total RNA was extracted from HSCs using TRIzol
reagent (Thermo Fisher Scientific, Inc.) according to the
manufacturer's protocol. Total RNA from rat hypothalamus was used
as a positive control. Total RNA was subjected to DNase
(Invitrogen; Thermo Fisher Scientific, Inc.) treatment prior to
reverse transcription. For RT-PCR, 1 µg total RNA was reverse
transcribed with Moloney murine leukemia virus reverse
transcriptase (M-MLV; Santa Cruz Biotechnology, Inc., Dallas, TX,
USA) according to the manufacturer's instructions. cDNAs were then
amplified using specific sets of primers for each gene. Taq DNA
polymerase purchased from Promega Corporation was used for PCR. The
amplification cycle numbers for Galanin, GalR1, GalR2 and GalR3
were 40, 37, 37 and 40, respectively. The sequences of each pair of
primers, product sizes and amplification conditions are listed in
Table I. In parallel, PCR was
performed with primers coding for GAPDH to control for equal
amounts of template cDNAs. Analysis was conducted using 5–20 µl
total PCR product in a 2.5% agarose gel. The gels were scanned
using a densitometer (Furi Science & Technology Ltd., Shanghai,
China). Primer specificity was confirmed with sequencing of PCR
products.
 | Table I.PCR primers, annealing temperature and
amplicon size of galanin and galanin receptors. |
Table I.
PCR primers, annealing temperature and
amplicon size of galanin and galanin receptors.
| Gene | Sequence | Annealing T (°C) | Size (bp) |
|---|
| Galanin | Foward:
5′-TACCCCTGCCTGAGAGCAAT-3′ | 61 | 118 |
|
| Reverse:
5′-TCTTCTGAGGAGGTGGCCAA-3′ |
|
|
| GalR1 | Foward:
5′-TGCTTTTGCTATGCCAAGGTTC-3′ | 61 | 359 |
|
| Reverse:
5′-TCGGTTCTTTTCTTTAGCATCGC-3′ |
|
|
| GalR2 | Foward:
5′-TTCTGCCTCTGTTGGATGCC-3′ | 58 | 362 |
|
| Reverse:
5′-TCTACTCGAGGCTGTGCAGTTG-3′ |
|
|
| GalR3 | Foward:
5′-AGCCAAGCAGTACCACAGAT-3′ | 62 | 161 |
|
| Reverse:
5′-GGTGAGGTAGATGAGCAGAT-3′ |
|
|
| GAPDH | Foward:
5′-CATGGTCTACACGTACCAGT-3′ | 58 | 349 |
|
| Reverse: 5′-
GGCAAGCAGTI'GGTGGTGC-3′ |
|
|
Immunofluorescence
The HSC-T6 cell line, which is an immortalized rat
liver stellate cell line that has a stable phenotype and
biochemical characteristics compared with primary stellate cells
(9), was obtained from the Chinese
Academy of Sciences (Shanghai, China). HSC-T6 cells were seeded in
a 4-well cell culture chamber in 10% FBS/DMEM at 2×103
cells/cm2. The culture medium was changed daily and
cells were fixed in ice-cold 4% paraformaldehyde after 3 days.
After further washes, cells were incubated at room temperature for
30 min with TBS containing 10% goat serum and 1% bovine serum
albumin (both purchased from Vector Laboratories, Burlingame, CA,
USA) to prevent non-specific binding of primary antibody. For GalR2
staining, fixed HSC-T6 cells were incubated with anti-GalR2 goat
polyclonal antibody (cat. no. ab59029; Abcam, Cambridge, MA, USA;
1:100 dilution) at 4°C overnight followed by incubation with
fluorescein isothiocyanate-conjugated secondary antibodies (cat.
no. ab150141; Abcam, Cambridge, MA; 1:100 dilution) at 37°C for 2
h. A negative control without primary antibody staining was
included.
Cell proliferation assay
HSC-T6 cells were seeded on 94-well plastic plates
in 10% FBS/DMEM at 2×103 cells/cm2. At 3 days
after seeding, HSCs were serum-starved for 24 h in 2% FBS/DMEM and
then subjected to treatment with galanin (Sigma-Aldrich) at
concentrations of 1–10,000 nmol/l for 24 h. Cell proliferation was
measured using an MTT assay. A working concentration of galanin
(100 nmol/l) was identified and used for the following
experiments.
Small interfering RNA (siRNA)
transfection and cell treatment
The siRNAs against mouse GalR2 and GalR3 mRNA were
designed and synthesized by Life Technologies (Thermo Fisher
Scientific, Inc.). The most effective one was selected for the
construction of the siRNA expression vector. The siRNAs had the
following sequences: GalR2-siRNA, forward:
5′CUCCAAGCAUUUCCGUAAAdTdT3′ and reverse:
5′UUUACGGAAAUGCUUGGAGdTdT3′; GalR3-siRNA, forward:
5′CCUCGCACUGCCUCGCCUAdTdT3′ and reverse
5′UUAGUCUAGUCUCUCCACCdTdT3′. Scrambled siRNA (forward
5′GCTCATATTACCAGTCACATT3′ and reverse 5′GAGGTGGATCGATTTATTCTA3′)
was also chemically synthesized and served as a negative control to
assess non-specific gene silencing effects (Life Technologies).
HSC-T6 cells were transfected with a mixture of plasmid DNA and
Lipofectamine 2000 (Life Technologies) in Opti-MEM I medium without
serum as recommended by the manufacturer. The medium was later
replaced with standard RPMI 1640 medium (containing 10% FBS;
Sigma-Aldrich; Merck Millipore). Cultured HSC-T6 cells were divided
to four groups: Control, galanin treatment group (galanin 100
nmol/l), GalR2 siRNA treatment group (GalR2 siRNA + galanin) and
GalR3 siRNA treatment group (GalR3 siRNA + galanin). The
proliferation of HSC-T6 cells was measured using an MTT assay.
Western blot analysis
Cultured HSC-T6 cells were homogenized in ice-cold
radioimmunoprecipitation assay buffer containing protease and
phosphatase inhibitors (Beyotime Biotechnology, Shanghai, China).
Western blotting was performed as previously described (10). The mouse antibodies anti-α-smooth
muscle actin (α-SMA; cat. no. ab18147; 1:2,000 dilution),
anti-transforming growth factor (TGF)-β1 (cat. no. ab64715; 1:2,000
dilution), anti-peroxisome proliferator-activated receptor (PPAR)-γ
(cat. no. ab41928; 1:1,000 dilution) and anti-β-actin (cat. no.
ab8226; 1:3,000 dilution) were obtained from Abcam (Cambridge, MA,
USA), and secondary horseradish peroxidase-conjugated goat
anti-mouse IgG antibody (cat. no. A9917; 1:60,000 dilution) was
purchased from Sigma-Aldrich (Merck Millipore). Protein bands were
quantified with Bio-Rad Image Lab 2.0 software (Bio-Rad
Laboratories, Inc., Hercules, CA, USA). β-actin was used as the
loading control.
Statistical analysis
Results were expressed as mean ± standard error of
the mean. At least three independent experiments were performed for
each experiment. Data were analyzed by Student's t-test or one way
analysis of variance followed by Mann Whitney U test using the SAS
system (release 8.02, TS level 02M0; SAS Institute, Cary, NC, USA).
P<0.05 was considered to indicate a statistically significant
result.
Results
Differential expression patterns of
galanin and GalRs in quiescent and activated HSCs
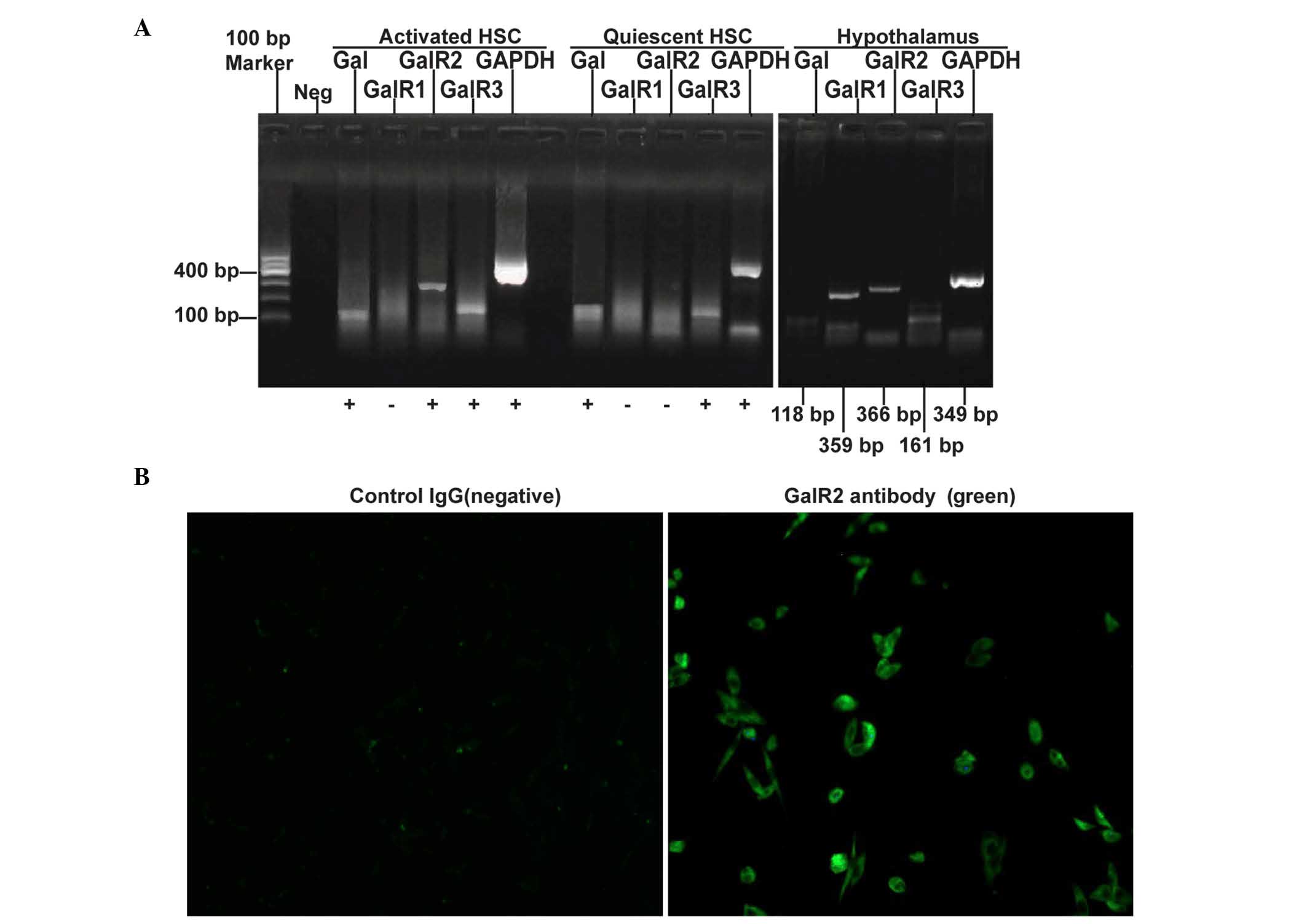
The expression levels of galanin and GalRs were
evaluated using RT-PCR, and it was found that galanin and GalR3
mRNA are expressed in both quiescent and activated HSCs while GalR2
mRNA is undetectable in quiescent HSCs but is highly expressed in
activated HSCs. GalR1 mRNA was undetectable in both quiescent and
activated HSCs (Fig. 1A). The
expression of GalR2 protein in cultured HSC-T6 cells was confirmed
using immunofluorescence staining (Fig.
1B).
Galanin and GalRs regulate HSC
proliferation and profibrogenic protein expression
Whether galanin and its receptors have roles in HSC
proliferation and the profibrogenic process was next investigated.
HSC-T6 cells were treated with different concentration of galanin
ranging from 1–10,000 nmol/l and it was found that 100 and 1,000
nmol/l galanin significantly inhibited the proliferation of HSCs.
The 100 nmol/l concentration of galanin showed the most marked
inhibitory effect (Fig. 2A). To
study the roles of GalRs on HSC growth, GalRs were depleted in
HSC-T6 cells using siRNA and it was found that GalR2 knockdown did
not have a notable effect on cell proliferation while GalR3
depletion significantly inhibited cell proliferation (P<0.05;
Fig. 2B).
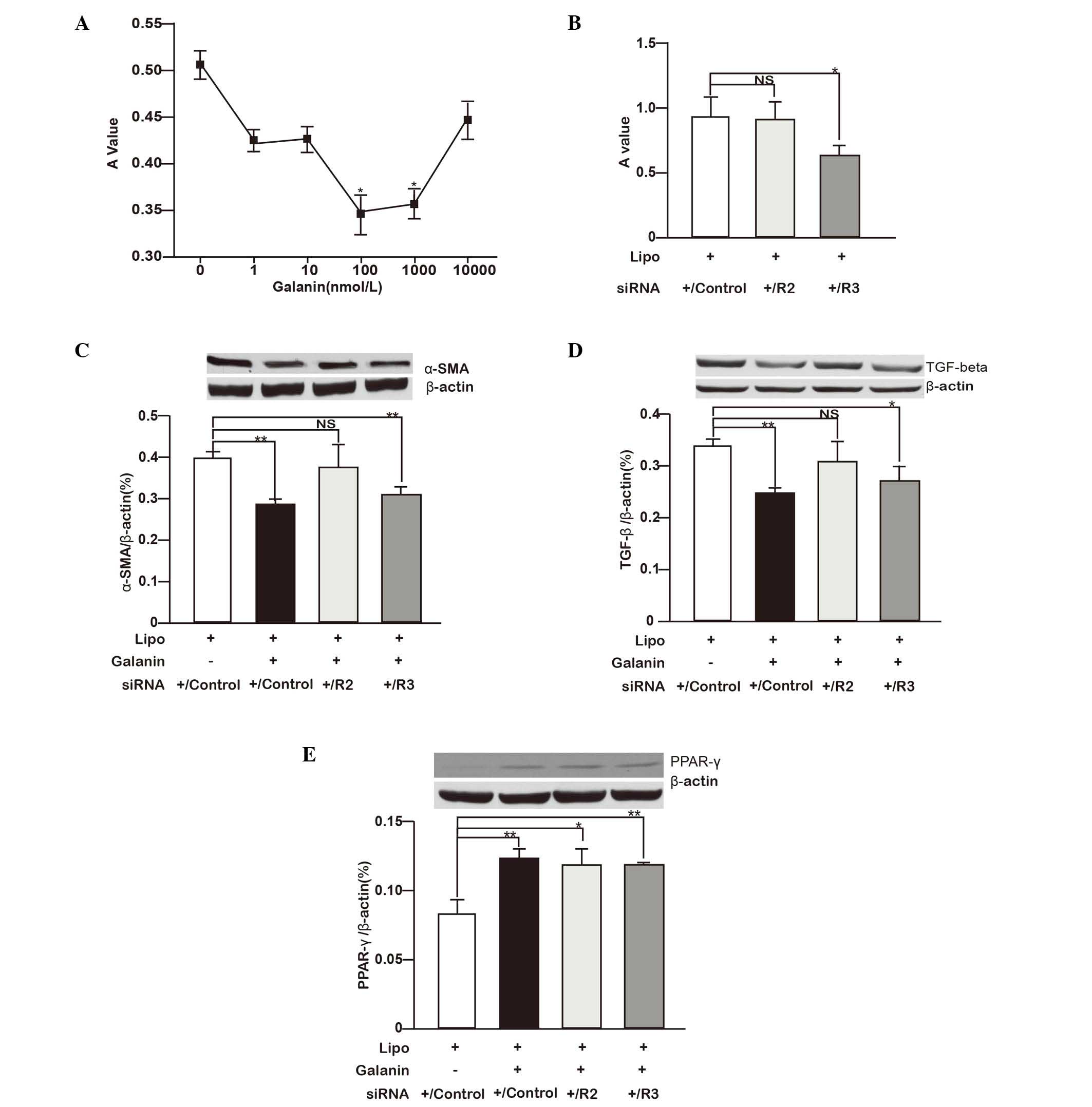 | Figure 2.Galanin and GalRs regulate HSC
proliferation and profibrogenic gene expression. (A) HSC-T6 cells
were treated with different concentrations of galanin (1–10,000
nmol/l) and the cell proliferation was measured with an MTT assay.
The A value measured after MTT administration represents the viable
cell number. *P<0.05 vs. control (0 nmol/l). (B) HSC-T6 cells
were treated with siRNA against GalR2 and GalR3, or scrambled siRNA
(control siRNA). Cell proliferation is shown as the rate relative
to the controls. *P<0.05. (C-E) Effects of galanin (100 nmol/l)
treatment and siRNA knockdown of GalR2, GalR3 or control siRNA on
(C) α-SMA, (D) TGF-β1 and (E) PPAR-γ protein expression in HSC-T6
cells are shown. The proteins detected by western blotting were
quantified with image analysis software and the relative expression
levels of α-SMA, TGF-β1 and PPAR-γ to β-actin are shown as bar
graphs. Western blot images are shown on top of the bar graphs.
*P<0.05, **P<0.01. GalR, galanin receptor; R2, GalR2; R3,
GalR3; HSC, hepatic stellate cell; A, absorbance; siRNA, small
interfering RNA; Lipo, lipofectamine; NS, not significant; α-SMA,
α-smooth muscle actin; TGF-β, transforming growth factor-β; PPAR-γ,
peroxisome proliferator-activated receptor-γ. |
The regulatory roles of galanin and GalRs on the
profibrogenic features of HSCs were further studied by measuring
the expression of key proteins. As shown in Fig. 2C-E, compared with the corresponding
controls, galanin (100 nmol/l) significantly inhibited TGF-β1 and
α-SMA protein expression while stimulating PPAR-γ expression
(P<0.01). GalR2 knockdown in HSC-T6 cells did not markedly
affect TGF-β1 and α-SMA protein expression but significantly
increased PPAR-γ expression (P<0.05) compared with that in the
control group. By contrast, GalR3 knockdown attenuated TGF-β1
(P<0.05) and α-SMA (P<0.01) protein expression while
increasing PPAR-γ expression (P<0.01) compared with the
respective levels in the control group. When considered together,
the data indicate that galanin inhibits HSC proliferation and
suppresses the profibrogenic features of these cells. Its
regulatory roles are likely mediated by GalR2.
Discussion
In the current study, it was demonstrated that GalR2
expression is undetectable in quiescent HSCs but induced in
activated HSCs, whereas galanin and GalR3 are expressed in both
quiescent and activated HSCs. Galanin (100 nmol/l) significantly
inhibited cell proliferation, α-SMA and TGF-β1 expression and
stimulated PPAR-γ expression in activated HSCs. The results suggest
the effects of galanin might be mediated by GalR2.
Activated HSCs transdifferentiate into
myofibroblasts, which then produce massive amounts of ECM proteins.
The activated HSCs adopt a smooth muscle myofibroblast-like
phenotype that is characterized by the expression of α-SMA
(11). Based on the fact that
galanin can inhibit the proliferation of PC12 cells (12), counteracts leptin, is involved in
fibrogenesis, and undergoes expression and secretion in rat adipose
tissue and 3T3-L1 adipocytes (5,6), it was
proposed that galanin might inhibit the activation and
proliferation of HSCs. The expression of galanin and GalRs in HSCs
was therefore investigated and confirmed in this study (Fig. 1). Cell proliferation assay results
indicated that galanin also inhibits the proliferation of HSCs
(Fig. 2A). Moreover, as a
characteristic of activated HSCs, α-SMA protein was also
downregulated when HSC-T6 cells were treated with galanin (Fig. 2C).
A biphasic inhibitory effect of galanin, occurring
at low concentrations (100–1,000 nmol/l) and disappearing at a
higher concentration (10,000 nmol/l), was observed. The mechanism
of this biphasic inhibitory effect is unclear. It is speculated
that such a high concentration of galanin will result in the
saturation of GalR2 and GalR3. The saturation of these two
receptors may lead to a biological effect totally different from
the variable activation of GalR2 and GalR3 observed at a lower
concentration of galanin (13).
Other biological processes such as receptor endocytosis may also be
involved. Indeed, a similar pharmacological profile has been
reported for other peptides in several types of cells, including
cardiac fibroblasts (13). Although
the underlying mechanism is unknown, others also speculate that it
might be associated with the activation of multiple receptors or
enzymes (13).
Inhibition of HSC activation and proliferation by
galanin may be associated with the downregulation of TGF-β1
protein. TGF-β1 is derived from paracrine and autocrine sources,
and is the most potent fibrogenic cytokine in liver (14). Quiescent HSCs induced by TGF-β1
transdifferentiate into myofibroblasts that secrete ECM (15). HSCs generate fibrosis by increasing
cell number and matrix production per cell (11). Inhibition of HSC proliferation and
downregulation of TGF-β1 expression may contribute to the control
of liver fibrosis (8,14).
Activated reversion of HSCs is another interesting
process that was investigated in the present study. PPAR-γ is
highly expressed in quiescent HSCs where it reinforces the
adipogenic property of the cells (16). The expression of PPAR-γ is markedly
reduced during HSC activation. Forced re-expression of PPAR-γ leads
to a partial reversal of the cells' pro-fibrogenic phenotypes,
including the loss of collagen 1 expression, prevention of TGF-β1
signaling, reduction of proliferation capacity, re-expression of
adipogenic transcription factors and re-uptake of lipid into
intracellular vesicles (16–18). In the present study, it was
demonstrated that galanin upregulates the expression of PPAR-γ
protein in activated HSCs. Indeed, HSCs are a kind of lipocyte and
galanin is also present in adipocytes (18,19).
Since galanin exerts its effects through GalRs, and GalR2
expression is induced during HSC activation, whether GalR2 has
regulatory roles in HSCs was then considered.
To investigate this, GalR2 and GalR3 were depleted
in HSC-T6 cells using siRNA. The results suggest that GalR2 may
mediate the function of galanin in HSCs. Following the knockdown of
GalR2 by siRNA, the activation of GalR3 by galanin does not
influence these effects of galanin on HSCs. However, activation of
GalR2 alone by galanin following the knockdown of GalR3 inhibits
HSC proliferation and TGF-β1 and α-SMA expression, in addition to
inducing PPARγ expression. This result is similar to the findings
made by Tofighi et al (12).
They found galanin decreases the proliferation of PC12 cells and
induces cell apoptosis via its subtype 2 receptor. GalR2 functions
through several different classes of G proteins that activate
diverse intracellular pathways. The most frequently reported
pathway involves activation of the Gq/11-type G protein, which
leads to the hydrolysis of inositol phosphate and activation of
protein kinase C (PKC) and phosphoinositide 3-kinase (PI3K)
(19–21). The P13K-Akt/protein kinase B pathway
and PKC family members activate the Ras-mitogen-activated protein
kinase pathway, which is involved in platelet-derived growth factor
signaling, one of the most well-characterized pathways of HSC
activation (22,23).
In summary, the results of the present study
indicate that galanin inhibits HSC activation and suppresses their
profibrogenic features. The underlying mechanisms may involve
inhibition of HSC proliferation, reduction of TGF-β1 expression and
induction of PPARγ expression. These biological functions of
galanin might be mediated by GalR2. Therefore, galanin is a
promising endogenous factor involved in the inhibition of liver
fibrosis. Additional studies using animal models are required to
gain further insight into the biological effects of galanin in
fibrosis.
Acknowledgements
This study was supported by grants from the National
Natural Science Foundation of China (grant no. 81170410 for Dr
Yuanwen Chen and grant no. 81260081 for Dr Yongnian Ding) and
sponsored by the New Talented Young Medical Specialist Cultivating
Program of Shanghai (grant no. XYQ2011010 for Dr Yuanwen Chen), the
Key Research Program for Colleges and Universities of Xinjiang
Uygur Autonomous Region (grant no. XJEDU2012I001 for Dr Yongnian
Ding), and by the State Key Development Program for Basic Research
of China (grant no. 2012CB517501 for Dr Jiangao Fan).
References
|
1
|
Tatemoto K, Rökaeus A, Jörnvall H,
McDonald TJ and Mutt V: Galanin - a novel biologically active
peptide from porcine intestine. FEBS Lett. 164:124–128. 1983.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Gundlach AL, Burazin TC and Larm JA:
Distribution, regulation and role of hypothalamic galanin systems:
Renewed interest in a pleiotropic peptide family. Clin Exp
Pharmacol Physiol. 28:100–105. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Poritsanos NJ, Mizuno TM, Lautatzis ME and
Vrontakis M: Chronic increase of circulating galanin levels induces
obesity and marked alterations in lipid metabolism similar to
metabolic syndrome. Int J Obes (Lond). 33:1381–1389. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Barrachina MD, Martínez V, Wei JY and
Taché Y: Leptin-induced decrease in food intake is not associated
with changes in gastric emptying in lean mice. Am J Physiol.
272:R1007–R1011. 1997.PubMed/NCBI
|
|
5
|
Li RY, Song HD, Shi WJ, Hu SM, Yang YS,
Tang JF, Chen MD and Chen JL: Galanin inhibits leptin expression
and secretion in rat adipose tissue and 3T3-L1 adipocytes. J Mol
Endocrinol. 33:11–19. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wang J, Leclercq I, Brymora JM, Xu N,
Ramezani-Moghadam M, London RM, Brigstock D and George J: Kupffer
cells mediate leptin-induced liver fibrosis. Gastroenterology.
137:713–723. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Saxena NK, Saliba G, Floyd JJ and Anania
FA: Leptin induces increased alpha2(I) collagen gene expression in
cultured rat hepatic stellate cells. J Cell Biochem. 89:311–320.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Dooley S, Delvoux B, Streckert M, Bonzel
L, Stopa M, ten Dijke P and Gressner AM: Transforming growth factor
beta signal transduction in hepatic stellate cells via Smad2/3
phosphorylation, a pathway that is abrogated during in vitro
progression to myofibroblasts. TGFbeta signal transduction during
transdifferentiation of hepatic stellate cells. FEBS Lett.
502:4–10. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Vogel S, Piantedosi R, Frank J, Lalazar A,
Rockey DC, Friedman SL and Blaner WS: An immortalized rat liver
stellate cell line (HSC-T6): A new cell model for the study of
retinoid metabolism in vitro. J Lipid Res. 41:882–893.
2000.PubMed/NCBI
|
|
10
|
Chen Y, Pat B, Zheng J, Cain L, Powell P,
Shi K, Sabri A, Husain A and Dell'italia LJ: Tumor necrosis
factor-alpha produced in cardiomyocytes mediates a predominant
myocardial inflammatory response to stretch in early volume
overload. J Mol Cell Cardiol. 49:70–78. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Benyon RC and Arthur MJ: Extracellular
matrix degradation and the role of hepatic stellate cells. Semin
Liver Dis. 21:373–384. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Tofighi R, Joseph B, Xia S, Xu ZQ,
Hamberger B, Hökfelt T and Ceccatelli S: Galanin decreases
proliferation of PC12 cells and induces apoptosis via its subtype 2
receptor (GalR2). Proc Natl Acad Sci USA. 105:2717–2722. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Rhaleb NE, Peng H, Harding P, Tayeh M,
LaPointe MC and Carretero OA: Effect of
N-acetyl-seryl-aspartyl-lysyl-proline on DNA and collagen synthesis
in rat cardiac fibroblasts. Hypertension. 37:827–832. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Inagaki Y and Okazaki I: Emerging insights
into transforming growth factor beta Smad signal in hepatic
fibrogenesis. Gut. 56:284–292. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Breitkopf K, Godoy P, Ciuclan L, Singer MV
and Dooley S: TGF-beta/Smad signaling in the injured liver. Z
Gastroenterol. 44:57–66. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Hazra S, Miyahara T, Rippe RA and
Tsukamoto H: PPAR gamma and hepatic stellate cells. Comp Hepatol.
3(Suppl 1): S72004. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Tsukamoto H, She H, Hazra S, Cheng J and
Miyahara T: Anti-adipogenic regulation underlies hepatic stellate
cell transdifferentiation. J Gastroenterol Hepatol. 21(Suppl 3):
S102–S105. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
She H, Xiong S, Hazra S and Tsukamoto H:
Adipogenic transcriptional regulation of hepatic stellate cells. J
Biol Chem. 280:4959–4967. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Fathi Z, Battaglino PM, Iben LG, Li H,
Baker E, Zhang D, McGovern R, Mahle CD, Sutherland GR, Iismaa TP,
et al: Molecular characterization, pharmacological properties and
chromosomal localization of the human GALR2 galanin receptor. Brain
Res Mol Brain Res. 58:156–169. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lehmann JM, Kliewer SA, Moore LB,
Smith-Oliver TA, Oliver BB, Su JL, Sundseth SS, Winegar DA,
Blanchard DE, Spencer TA and Willson TM: Activation of the nuclear
receptor LXR by oxysterols defines a new hormone response pathway.
J Biol Chem. 272:3137–3140. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wang S, Hashemi T, Fried S, Clemmons AL
and Hawes BE: Differential intracellular signaling of the GalR1 and
GalR2 galanin receptor subtypes. Biochemistry. 37:6711–6717. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wong L, Yamasaki G, Johnson RJ and
Friedman SL: Induction of beta-platelet-derived growth factor
receptor in rat hepatic lipocytes during cellular activation in
vivo and in culture. J Clin Invest. 94:1563–1569. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kelly JD, Haldeman BA, Grant FJ, Murray
MJ, Seifert RA, Bowen-Pope DF, Cooper JA and Kazlauskas A:
Platelet-derived growth factor (PDGF) stimulates PDGF receptor
subunit dimerization and intersubunit trans-phosphorylation. J Biol
Chem. 266:8987–8992. 1991.PubMed/NCBI
|