Introduction
Translationally-controlled tumor protein (TCTP) is
an evolutionally-conserved protein in yeast and humans (1–4), which
has important roles in cell cycle (5,6),
apoptosis (7), cytoskeleton
(6), protein synthesis (8), immune response (3), development (9) and cancer (7). In addition, a previous study reported
that TCTP expression was elevated in cancer tissues as well as
during liver regeneration (10); the
protein also has an important anti-tumor role (7,11).
Therefore, TCTP is considered a target for cancer therapy. TCTP is
phosphorylated by polo-like kinase (12) at mitosis, and is localized to
microtubules via binding with tubulin, which results in
depolarization and stabilization of microtubules (6). Under stress conditions, TCTP localizes
to the surface of mitochondria and inhibits B cell lymphoma
2-associated X protein dimerization to block apoptosis (6). TCPT binds with Chfr, which is a
G2/M checkpoint protein that controls the cell cycle
under stressful conditions (6). A
study on knockout mice demonstrated that the TCTP heterozygous
(TCTP+/−) mouse was developmentally normal
and the homozygous mutants (TCTP−/−) were
lethal at the embryonic stage (9),
suggesting the importance of TCTP in the developmental process.
Conversely, it was reported that TCTP activates octamer-binding
transcription factor 4 (Oct-4) expression (13), which is a stem cell marker and has a
role in stemness; it was also reported that TCTP downregulates
Oct-4 expression in mouse pluripotent cells (14). The potential use of stem cells for
therapy has a vital role in the field of regenerative medicine
(15). During kidney development, it
has been reported that stem cells are located in the metanephric
mesenchyme, which is the origin for various structures in the
mature kidney, except collecting duct, interstitium and vasculature
(16,17). In addition, it was reported that
kidney stem cells present in the adult kidneys of skates and
freshwater teleosts are able to participate in novel nephron
formation following partial nephrectomy (18–20).
Little is known regarding the molecular mechanisms and activation
of kidney-derived stem cells. In order to understand the
association between Oct-4 and TCTP in kidney-derived stem cells,
immunoblotting and reverse transcription-polymerase chain reaction
(RT-PCR) analysis of a TCTP null mutant was performed. The aim of
the present study was to demonstrate that TCTP activates the
transcription of Oct-4 in kidney-derived stem cells.
Materials and methods
Experimental animals
Animal use in the present study was approved by the
institutional animal care and ethical committee. A total of 12
heterozygous offspring of transgenic (Tg) rats (Rat ID,
RGD_ID1302921) generated from Fisher 344 with the insertion of
homology-directed repair (hDTR) cDNA were used throughout the
experiments. Tg rats were gifted by Dr Wei Zhang (School of
Science, Sun Yan-Sen University, Guangzhou, China). Rats were
maintained under standard conditions with free access to feed and
water and a 12-h light/dark cycle (lights on at 07:00), as
previously outlined (21).
Diphtheria toxin (DT; D0564; Sigma-Aldrich; Merck Millipore,
Darmstadt, Germany) was used to stimulate kidney injury in the
rats. The minimum lethal dose of DT for humans was reported to be
100 ng/kg body weight (21,22). Based upon the standard protocol
(23), 10 ng/kg DT is efficient at
causing kidney injury in Tg rats. Tg rats received 10 ng/kg per day
for three days via intraperitoneal (IP) injection. Both the control
Tg rats and DT-injected Tg rats were carefully monitored at regular
time intervals (every 6 h). Urine and feces samples were collected
from the animals to identify the presence of dead cells and to
determine the protein levels (TCTP, Pax, β-actin and Oct-4).
Following analysis for kidney damage via Tryphan blue in the urine,
the podocytes began to deplete slowly and samples were collected at
the 10th day after DT injection. Day 10 is the optimal time to
collect the kidney samples (based upon the preliminary
standardization protocol) to identify the presence and role of
kidney stem cells in the process to restore the injured cells
caused by the DT
Cell culture experiments
Pentobarbital sodium injection method was used to
sacrifice Tg rats. Briefly, rats were anesthetized with
pentobarbital sodium (60 mg/kg body weight; IP) prior to surgery.
Kidneys were exposed with the help of holding clamps and were
surgically removed after the 25 min of IP injection. Kidney-derived
stem cells were isolated from the Tg rat kidneys as follows: The
rat kidneys were surgically removed, harvested, minced, and
partially digested using collagenase (Sigma-Aldrich; Merck
Millipore) in the presence of trypsin inhibitor (T9253;
Sigma-Aldrich; Merck Millipore). The cell suspension was washed
with and plated in a medium composed of 58% Dulbecco's modified
Eagle's medium-low glucose and 42% MCDB-201, and supplemented with
1X insulin-transferrin-selenium, 1 mg/ml bovine serum albumin
(BSA), 0.05 M dexamethasone, 0.1 mM ascorbic acid 2-phosphate, 100
U penicillin, 1,000 U streptomycin, 2% fetal bovine serum, 10 ng/ml
epidermal growth factor, 10 ng/ml platelet-derived growth factor-BB
and 10 ng/ml leukemia inhibitory factor (all Sigma-Aldrich; Merck
Millipore). The medium composition used for the present study was
partially modified from the protocol previously described by Gupta
et al (24). The cells were
seeded on fibronectin-coated culture flasks (BD Biosciences, San
Jose, CA, USA) at low density (300 cells/cm2), to avoid
cell-cell contact, and cultured at 37°C in the presence of 5%
CO2. Single clones of cells were obtained by reseeding
the cells at non touching density and following experiments were
performed to characterize the isolated cells as a kidney derived
stem cells.
Generation of TCTP null mutant
TCTP−/− and normal embryo
control samples [both from embryos at day 9.5 (E9.5)] were used in
the present study. A TCTP null mutant
(TCTP−/−) was generated a previously
described (9). The 129/Svj mouse
strain was used. Genotyping of Tg rat tissues was performed by PCR
using Platinum Taq DNA polymerase (11146–057; Invitrogen; Thermo
Fisher Scientific, Inc., Waltham, MA, USA) and the following
primers from Sigma-Aldrich (Merck Millipore): K1,
5′-TCTAGAAAAGTGGAGGCGGAGC-3′ and K5, 5′-GGTGACTACTGTGCTTTCGGTA-3′
for the wild-type and floxed alleles, and K1 and K4,
5′-AAAGCAGATCCAGAATAACCCC-3′ for the deleted allele. A T100 Thermal
Cycler (Bio-Rad Laboratories, Inc., Hercules, CA, USA) was used for
PCR analysis. PCR products were separated by 2% agarose gel
electrophoresis with ethidium bromide. A Gel Doc EZ system and its
in-built software program (Bio-Rad Laboratories, Inc.) was used to
analyze the PCR results. All animal experiments were performed in
accordance with the guidelines set by the Institutional Animal Care
and Utilization Committee.
Immunostaining and RT-PCR
analysis
Kidney-derived stem cells were fixed with 4%
paraformaldehyde, permeabilized with Triton X-100 and blocked with
1% BSA in phosphate-buffered saline (PBS) for 1 h. The cells were
then incubated with anti-Oct-4 (1:100; SAB2500713) and anti-TCTP
(1:50; WH0007178M1; both Sigma-Aldrich; Merck Millipore) primary
antibodies overnight at 4°C. The plates were washed in 1X PBS and
incubated with the following horseradish peroxidase
(HRP)-conjugated secondary antibodies for 45 min at room
temperature in the dark: Rabbit anti-goat IgG HRP (ab97023), goat
anti-mouse IgG H&L (FITC) (ab6785), donkey anti-mouse IgG
H&L (Texas Red) (ab6818) and donkey anti-goat IgG H&L
(Texas Red) (all 1:10,000; ab6883; all Abcam). Plates were
developed with 3,3′-diaminobenzidine (DAB) substrate
(Sigma-Aldrich; Merck Millipore). Cells were then observed under a
Nikon Ti-S fluorescent microscope (Nikon Corporation, Tokyo,
Japan). For RT-PCR analysis, 2.4 µg total RNA was isolated from the
kidney-derived cultured stem cells using an Rneasy Mini kit
according to the manufacturer's protocol (Qiagen GmbH, Hilden,
Germany) and treated with DNase 1 (Invitrogen; Thermo Fisher
Scientific, Inc.), prior to being stored at −70°C. The quality and
quantity of the RNA were validated by standard procedures, which
included cross-checking of the quality of RNA by agarose gel
electrophoresis and of the quantity via a Nanodrop
spectrophotometer (Nanodrop Technologies, Pittsburgh, PA, USA). RT
to cDNA was performed using using Platinum Taq DNA polymerase with
0.5 µg total RNA as a template and PCR was performed on 1/20 of the
RT product using the following primers from Sigma-Aldrich (Merck
Millipore): Oct-4 forward, 5′-CTGTAACCGGCGCCAGAA-3′, and reverse,
5′-TGCATGGGAGAGCCCAGA-3′; paired box-2 (Pax-2) forward,
5′-TGGAGAGGCCTGCCAAGTA-3′, and reverse, 5′-AAGAGTGGGAGTTGCTGTTG-3′;
and TCTP forward, 5′-AAACCAGAAAGGGTAAAGCC-3′, and reverse,
5′-TCCACTCCAAATAAATCACGG-3′. Thermal cycling was performed under
standard conditions as follows: 40 cycles of two-step PCR (95°C for
15 sec and 60°C for 60 sec) after initial denaturation (95°C for 10
min) with 1 µl cDNA. PCR products were separated by 2% agarose gel
electrophoresis with ethidium bromide. The bands were observed and
documented using a Gel Doc EZ gel documentation unit (Bio-Rad
Laboratories, Inc.).
Immunoblotting analysis
Protein samples were isolated from the
TCTP−/− and normal embryo control samples
(both E9.5) using 2X protein sample buffer and incubating the
homogenate in a boiling water bath for 5 min. Protein quantity was
estimated according to Lowrys method of protein estimation using
BSA as standard. Protein samples (80 µg) were subsequently
separated by 8% SDS-PAGE. The resolved protein samples from the
SDS-PAGE gel were transferred to a polyvinylidene fluoride membrane
(Sigma-Aldrich; Merck Millipore), blocked with 4% BSA for 1 h and
incubated with the following primary antibodies: Anti-Oct-4
(1:100), anti-TCTP (1:50) and anti-Pax-2 (1:100; SAB1404166)
antibodies (all Sigma-Aldrich; Merck Millipore) overnight at 4°C.
Anti-β-actin antibody (1:100; A5316; Abcam, Cambridge, UK) was used
as a loading control. The non-specific binding of the primary
antibody was eliminated through washing with 1X Tris-buffered
saline with Tween-20. The following secondary antibody conjugated
with HRP were incubated for 1 h at room temperature: Rabbit
anti-goat IgG HRP (ab97023), goat anti-mouse IgG H&L (FITC)
(ab6785), donkey anti-mouse IgG H&L (Texas Red) (ab6818) and
donkey anti-goat IgG H&L (Texas Red) (all 1:10,000; ab6883; all
Abcam). The washed membrane was developed according to the
manufacturer's protocol using the DAB/H2O2
substrate (Amresco LLC, Solon, OH, USA) to produce a brown-colored
product, which appeared on the membranes and was clear to the naked
eye.
Results
Injection of DT
Tg rats express hDTR specifically in rat podocytes.
Injection of DT into Tg rats induces podocyte loss in a
dose-dependent manner. Damages in the kidney were noted on the 7th
day after DT injection, using Tryphan blue in the urine (data not
shown). The podocytes began to deplete slowly and samples were
collected at the 10th day after DT injection.
Isolation and culture of
kidney-derived stem cells
In order to isolate the kidney-derived stem cells,
cell culture technique was performed. Control Tg rats and
DT-injected Tg rats were used for the experiments to isolate the
kidney-derived stem cells. Kidney samples from both the control and
DT-injected Tg rats were surgically removed and processed for cell
culture. After 5 weeks, the majority of the cell types had died
(data not shown). In addition, it was observed that the cultures
became monomorphic and the cells appeared spindle-shaped when
observed under the phase contrast microscope (data not shown).
Immunostaining and RT-PCR
analysis
To confirm and validate that the isolated cells were
kidney-derived stem cells, immunostaining was performed. The
isolated cultured cells were immunostained with anti-Oct-4 and
anti-TCTP antibodies and analyzed under a fluorescent microscope
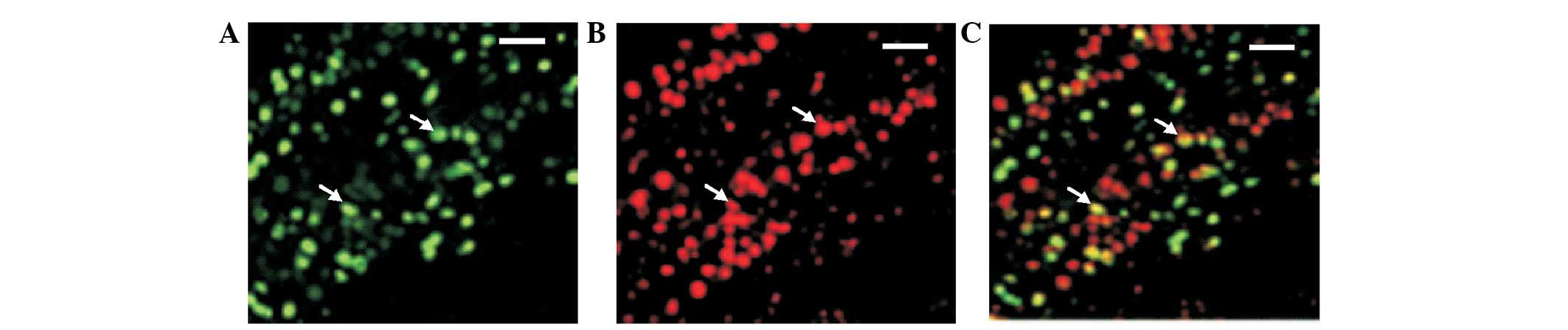
(Fig. 1). Oct-4-positive cells were
observed in the plates as shown in the Fig. 1A. TCTP expression was also noted in
the same cells (Fig. 1B).
Co-expression is shown in Fig. 1C.
These results suggest that the cells isolated and cultured from Tg
rat kidneys were kidney-derived stem cells. In addition,
co-expression of TCTP in Oct-4-expressing cells revealed that TCTP
may be involved in the activation of Oct-4, or conversely Oct-4 may
induce the activation of TCTP. Furthermore, in order to determine
whether Oct-4-positive cells co-express TCTP in the kidney-derived
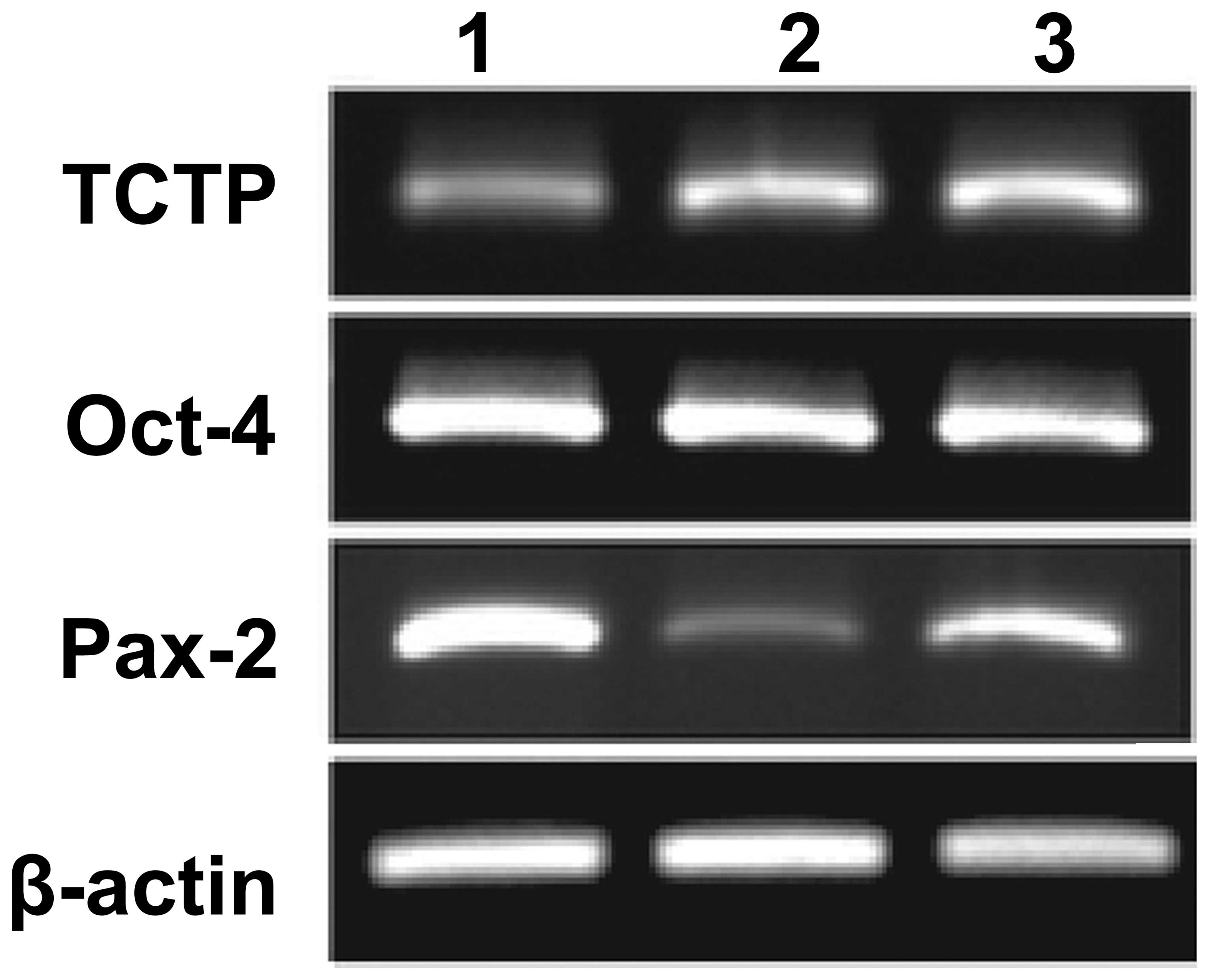
stem cells, RT-PCR was performed. The following expression profile
markers were analyzed using RT-PCR: Oct-4, TCTP, Pax-2 and β-actin.
Total RNA was isolated from the cultured kidney-derived stem cells
and RT-PCR was performed following a standard protocol. The results
demonstrated that co-expression of Oct-4 and TCTP occurred in the
kidney-derived stem cells (Fig. 2).
These results suggested TCTP may be involved in the activation of
Oct-4, or conversely Oct-4 may induce the activation of TCTP in
kidney-derived stem cells. Pax-2 was used as a kidney-specific
marker. Pax-2 expression was noted in the E9.5
TCTP−/− null mutant embryos and also in the
controls.
TCTP null mutant
In order to prove the hypothesis that TCTP activates
Oct-4 expression in kidney-derived stem cells, the following
experiments were designed: TCTP null mutant
(TCTP−/−) was generated. Although the TCTP
null mutant (TCTP−/−) is embryonically
lethal, E9.5 was used throughout the studies. E9.5 days embryos
were collected from the control and TCTP null mutant
(TCTP−/−) and subjected to immunoblotting.
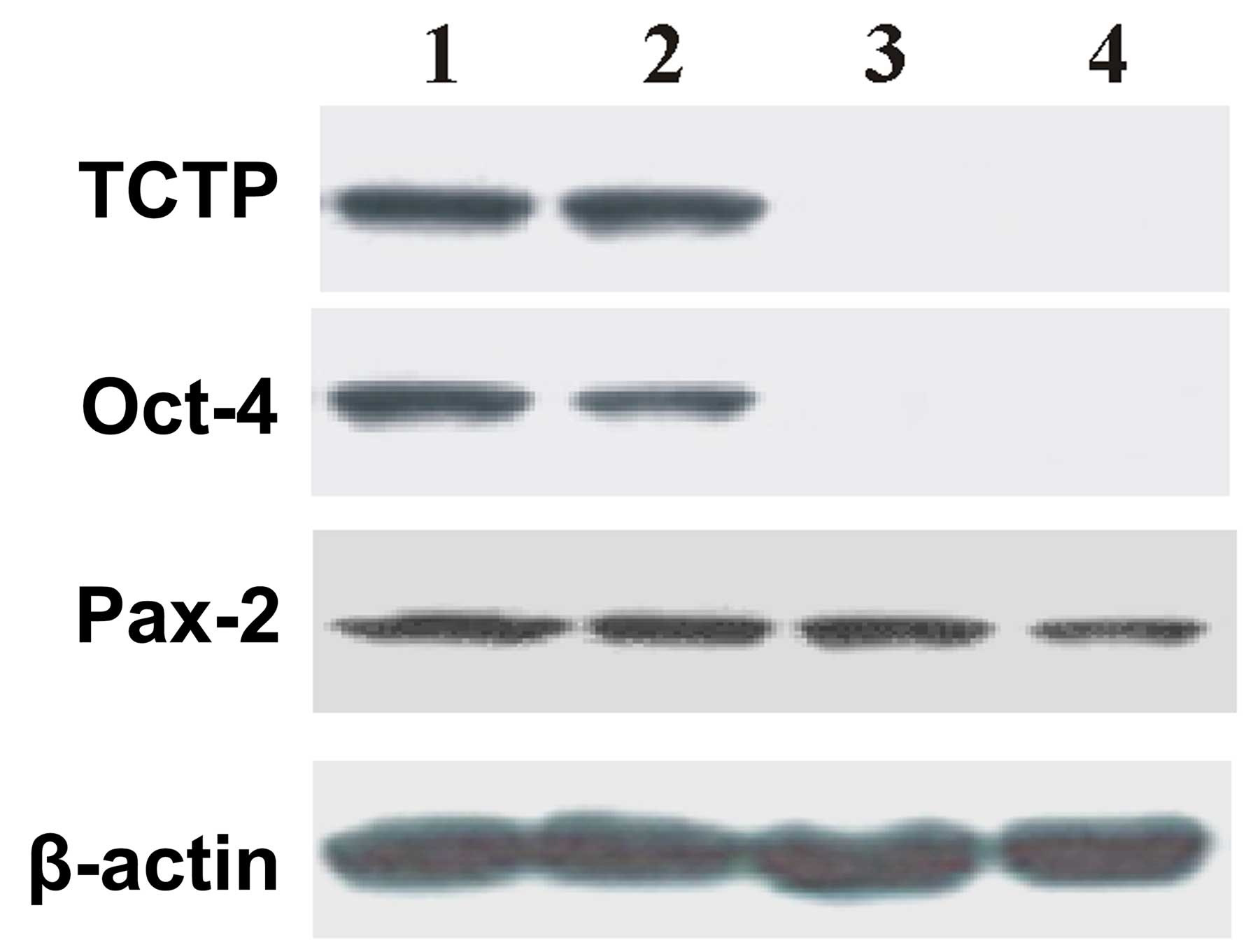
The results of the immunoblotting are shown in the Fig. 3, which demonstrates that there was no
expression of Oct-4 or TCTP in the TCTP null mutant
(TCTP−/−) embryos. The data suggests that
TCTP is associated with the activation of Oct-4. In addition, Pax-2
expression was noted in the E9.5 TCTP null mutant (TCTP-/-)
embryos. To further validate the data, total RNA was isolated from
the control and TCTP null mutant (TCTP-/-) embryos (E9.5) and
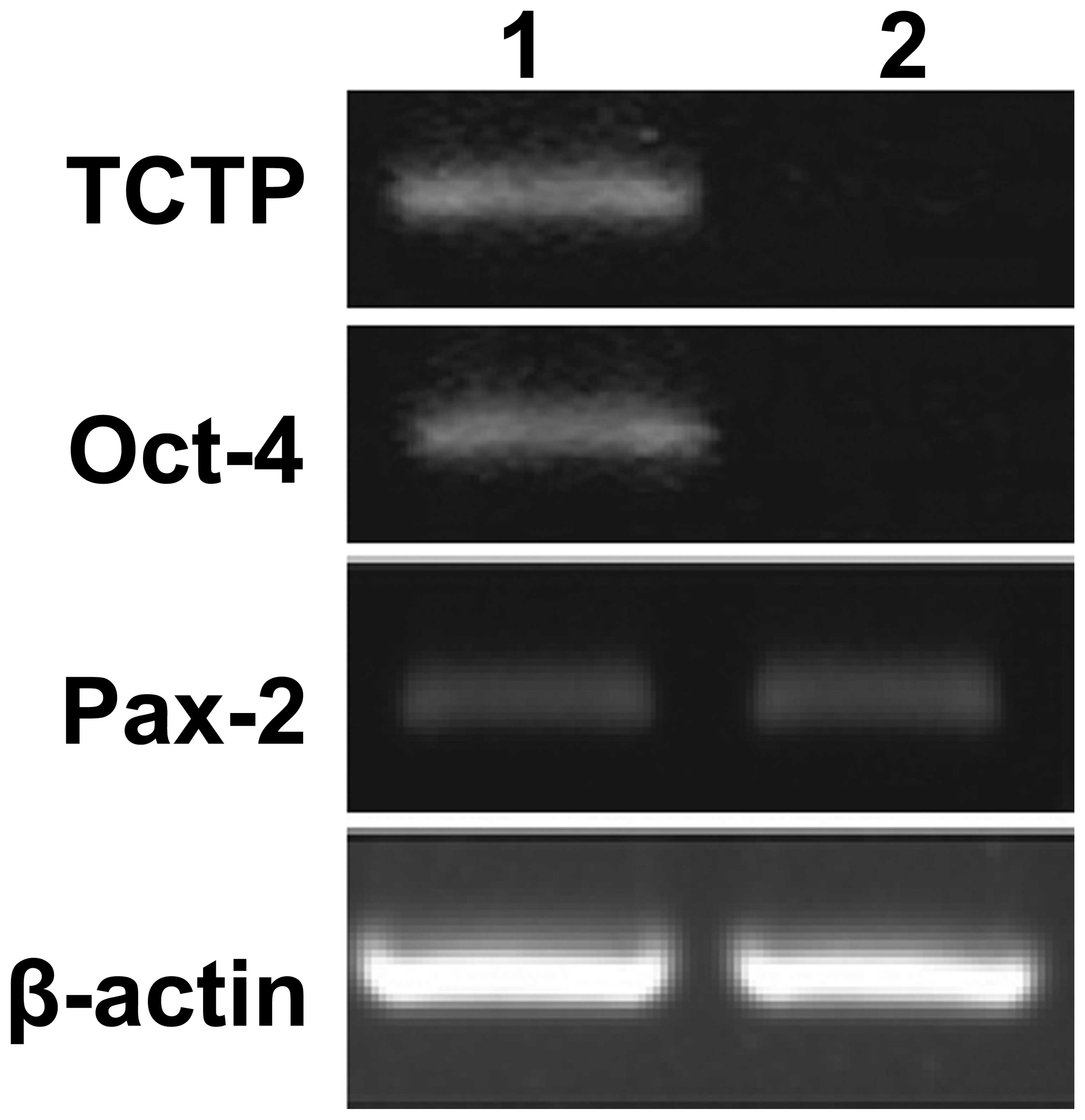
subjected to RT-PCR. As shown in Fig.
4, no expression of Oct-4 was observed in the TCTP null mutant
(TCTP-/-) embryos. In addition, Pax-2 transcripts were identified
both in the control and TCTP null mutant (TCTP-/-) embryos. These
results suggest that TCTP activates the transcription of Oct-4 in
kidney-derived stem cells.
Discussion
Little is known regarding the basic molecular
mechanisms underlying TCTP and its role in the activation of Oct-4
in kidney-derived stem cells. Adult stem cells and their niches has
been well characterized in numerous organs such as bone marrow,
intestine, skin, gastrointestinal mucosa, liver, prostate and brain
(25–29). Understanding the molecular mechanism
underlying kidney-derived stem cells processes will be useful for
regenerative medicine.
The present study used DT in order to determine
whether the kidney has stem cells. It is well-accepted that stem
cells are involved in renewable and compensation processes during
tissue injury or disease. In order to create an injury to the
kidney, DT-mediated injury was created in Tg rats as a model for
kidney injury. Potential kidney-derived stem cells were isolated
from the Tg rat kidneys, and were observed to express Oct-4 and
Pax-2. The results demonstrated that the cultured cells were
kidney-derived stem cells. In addition, the cultured kidney-derived
stem cells were immunostained with anti-Oct-4 and anti-TCTP
antibodies. The results demonstrated the following: i) The
expression of Oct-4 (stem cell marker) and Pax-2 (kidney-specific
marker) in the cells isolated and cultured from the Tg rats kidneys
indicated that the cells were kidney-derived stem cells; ii)
co-expression of TCTP in Oct-4-expressing cells revealed that TCTP
was associated with the activation of Oct-4 or vice
versa.
To validate the data and to confirm that the
identified cells were kidney-derived stem cells, RT-PCR was
performed. The expression profiles of Oct-4, Pax-2 and TCTP in the
kidney-derived stem cells were examined. The results demonstrated
that the co-expression of Oct-4, TCTP and Pax-2 was observed in the
kidney-derived stem cells. The results suggested that TCTP induced
the activation of Oct-4 or Oct-4 induced the activation of TCTP in
kidney-derived stem cells.
Immunoblotting experiments with TCTP null mutant
(TCTP−/−) embryos (E9.5) demonstrate the
absence of Oct-4 and TCTP co-expression. In addition, the
expression of Pax-2 and β-actin in the
TCTP−/− embryos indicated the following: (i)
The TCTP null mutant (TCTP−/−) embryos (E9.5)
had kidney-derived stem cells; (ii) TCTP has no role in the
activation of Pax-2; and (iii) the expression of β-actin was not
changed in the TCTP null mutant (TCTP−/−).
The results suggested that the TCTP is important for the activation
of Oct-4 expression.
RNA samples were prepared from the TCTP null mutant
(TCTP−/−) embryos (E9.5) and subjected to
RT-PCR with Oct-4, Pax-2 and TCTP primers. The results of the
experiment demonstrated that there was no expression of Oct-4 and
TCTP in the TCTP null mutant (TCTP−/−)
embryos (E9.5). However, Pax-2 expression was noted in the control
as well as the TCTP null mutant (TCTP−/−)
embryos (E9.5). These results suggested that TCTP activates the
transcription of Oct-4 in kidney-derived stem cells.
In conclusion, the results of the present study
demonstrated that TCTP activates the transcription of Oct-4 in
kidney-derived stem cells. The characteristics and functional
nature of TCTP in connection with Oct-4 in kidney-derived stem
cells was identified. These results may serve in regenerative
medicine and kidney diseases in future.
Acknowledgements
The authors are grateful to the institutional review
ethical board approval committee for the successful completion of
this project.
References
|
1
|
Yenofsky R, Cereghini S, Krowczynska A and
Brawerman G: Regulation of mRNA utilization in mouse
erythroleukemia cells induced to differentiate by exposure to
dimethyl sulfoxide. Mol Cell Biol. 3:1197–1203. 1983. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Böhm H, Benndorf R, Gaestel M, Gross B,
Nürnberg P, Kraft R, Otto A and Bielka H: The growth-related
protein P23 of the Ehrlich ascites tumor: Translational control,
cloning and primary structure. Biochem Int. 19:277–286.
1989.PubMed/NCBI
|
|
3
|
MacDonald SM, Rafnar T, Langdon J and
Lichtenstein LM: Molecular identification of an IgE-dependent
histamine-releasing factor. Science. 269:688–690. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Thaw P, Baxter NJ, Hounslow AM, Price C,
Waltho JP and Craven CJ: Structure of TCTP reveals unexpected
relationship with guanine nucleotide-free chaperones. Nat Struct
Biol. 8:701–704. 2001. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
Brioudes F, Thierry AM, Chambrier P,
Mollereau B and Bendahmane M: Translationally controlled tumor
protein is a conserved mitotic growth integrator in animals and
plants. Proc Natl Acad Sci USA. 107:16384–16389. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Gachet Y, Tournier S, Lee M,
Lazaris-Karatzas A, Poulton T and Bommer UA: The growth-related,
translationally controlled protein P23 has properties of a tubulin
binding protein and associates transiently with microtubules during
the cell cycle. J Cell Sci. 112:1257–1271. 1999.PubMed/NCBI
|
|
7
|
Tuynder M, Susini L, Prieur S, Besse S,
Fiucci G, Amson R and Telerman A: Biological models and genes of
tumor reversion: Cellular reprogramming through tpt1/TCTP and
SIAH-1. Proc Natl Acad Sci USA. 99:14976–14981. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Cans C, Passer BJ, Shalak V,
Nancy-Portebois V, Crible V, Amzallag N, Allanic D, Tufino R,
Argentini M, Moras D, et al: Translationally controlled tumor
protein acts as a guanine nucleotide dissociation inhibitor on the
translation elongation factor eEF1A. Proc Natl Acad Sci USA.
100:13892–13897. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Chen SH, Wu PS, Chou CH, Yan YT, Liu H,
Weng SY and Yang-Yen HF: A knockout mouse approach reveals that
TCTP functions as an essential factor for cell proliferation and
survival in a tissue-or cell type-specific manner. Mol Biol Cell.
18:2525–2532. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhu WL, Cheng HX, Han N, Liu DL, Zhu WX,
Fan BL and Duan FL: Messenger RNA expression of translationally
controlled tumor protein (TCTP) in liver regeneration and cancer.
Anticancer Res. 28:1575–1580. 2008.PubMed/NCBI
|
|
11
|
Arcuri F, Papa S, Carducci A, Romagnoli R,
Liberatori S, Riparbelli MG, Sanchez JC, Tosi P and del Vecchio MT:
Translationally controlled tumor protein (TCTP) in the human
prostate and prostate cancer cells: Expression, distribution, and
calcium binding activity. Prostate. 60:130–140. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yarm FR: Plk phosphorylation regulates the
microtubule-stabilizing protein TCTP. Mol Cell Biol. 22:6209–6221.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Koziol MJ, Garrett N and Gurdon J: Tpt1
Activates transcription of oct4 and nanog in transplanted somatic
nuclei. Curr Biol. 17:801–807. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Cheng X, Li J, Deng J, Li Z, Meng S and
Wang H: Translationally controlled tumor protein (TCTP)
downregulates Oct4 expression in mouse pluripotent cells. BMB Rep.
45:20–25. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Weissman IL: Stem cells: Units of
development, units of regeneration, and units in evolution. Cell.
100:157–168. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Herzlinger D, Koseki C, Mikawa T and
al-Awqati Q: Metanephric mesenchyme contains multipotent stem cells
whose fate is restricted after induction. Development. 114:565–572.
1992.PubMed/NCBI
|
|
17
|
Oliver JA, Maarouf O, Cheema FH, Martens
TP and Al-Awqati Q: The renal papilla is a niche for adult kidney
stem cells. J Clin Invest. 114:795–804. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Drummond IA, Mukhopadhyay D and Sukhatme
VP: Expression of fetal kidney growth factors in a kidney tumor
line: Role of FGF2 in kidney development. Exp Nephrol. 6:522–533.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Elger M, Hentschel H, Litteral J, Wellner
M, Kirsch T, Luft FC and Haller H: Nephrogenesis is induced by
partial nephrectomy in the elasmobranch Leucoraja erinacea. J Am
Soc Nephrol. 14:1506–1518. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Salice CJ, Rokous JS, Kane AS and
Reimschuessel R: New nephron development in goldfish (Carassius
auratus) kidneys following repeated gentamicin-induced
nephrotoxicosis. Comp Med. 51:56–59. 2001.PubMed/NCBI
|
|
21
|
Pappenheimer AM Jr: The story of a toxic
protein, 1888–1992. Protein Sci. 2:292–298. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wharram BL, Goyal M, Wiggins JE, Sanden
SK, Hussain S, Filipiak WE, Saunders TL, Dysko RC, Kohno K, Holzman
LB and Wiggins RC: Podocyte depletion causes glomerulosclerosis:
Diphtheria toxin-induced podocyte depletion in rats expressing
human diphtheria toxin receptor transgene. J Am Soc Nephrol.
16:2941–2952. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Humphreys BD, Valerius MT, Kobayashi A,
Mugford JW, Soeung S, Duffield JS, McMahon AP and Bonventre JV:
Intrinsic epithelial cells repair the kidney after injury. Cell
Stem Cell. 2:284–291. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Gupta S, Verfaillie C, Chmielewski D, Kren
S, Eidman K, Connaire J, Heremans Y, Lund T, Blackstad M, Jiang Y,
et al: Isolation and characterization of kidney-derived stem cells.
J Am Soc Nephrol. 17:3028–3040. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Alison MR, Poulsom R and Forbes SJ: Update
on hepatic stem cells. Liver. 21:367–373. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Bernard-Kargar C and Ktorza A: Endocrine
pancreas plasticity under physiological and pathological
conditions. Diabetes. 50:(Suppl 1). S30–S35. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Forbes SJ, Poulsom R and Wright NA:
Hepatic and renal differentiation from blood-borne stem cells. Gene
Ther. 9:625–630. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Morrison SJ, White PM, Zock C and Anderson
DJ: Prospective identification, isolation by flow cytometry, and in
vivo self-renewal of multipotent mammalian neural crest stem cells.
Cell. 96:737–749. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Wright NA: Epithelial stem cell repertoire
in the gut: Clues to the origin of cell lineages, proliferative
units and cancer. Int J Exp Pathol. 81:117–143. 2000. View Article : Google Scholar : PubMed/NCBI
|