Introduction
Islet transplantation has progressed to be a
possible cure for type 1 diabetes mellitus (1), but is clinically limited due to the
shortage of donor islets and immune rejection (2). Mesenchymal stem cells (MSCs) are
multipotent stromal cells that can differentiate into a variety of
cell types, and can be easily derived from many different tissues
with no ethic and legal issues. Most importantly is that they have
been reported to exhibit unique immunomodulation properties, which
make them optimal cell types in treating autoimmune diseases and
graft versus host disease (GvHD). MSCs have been successfully
induced to differentiate into insulin-producing cells (IPCs) in
vitro, and these IPCs may be a new cell source for islet
transplantation, thus they have great potential in cell replacement
therapy for diabetes (3–5).
MSCs express low levels of class I major
histocompatibility complex (MHC) molecules, but not class II and
co-stimulatory molecules. MSCs suppress lymphocyte proliferation
in vitro when co-cultured with PBMCs, and exert resistance
against the cytotoxicity of cytotoxic lymphocytes. Due to their
hypo-immunogenicity and immunosuppressive properties (6–8),
clinical trials on allogenic MSCs injection in many different acute
and chronic diseases have been registered and progressed
(https://clinicaltrials.gov). However,
MSCs may become immunogenic after differentiation and
transplantation to host, due to induction process and the
microenvironment of transplanted sites (9–14). In
vitro differentiation of rat bone marrow-derived MSCs into
muscle cells caused elevated expression of MHC-Ia and MHC-II, and
became immunogenic. After transplantation to the infracted
myocardium of allogenic rat, their survival and repair effects were
much weaker than those of autologous transplantation (12). The induction process of muscle cells
in vitro could reduce the secretion of immunomodulatory
molecule PEG2, thus influence the survival of the differentiated
cells in the host (15). The
situation was similar when bone marrow-derived MSCs were induced
into chrondocytes in vitro. The differentiated chondrocytes
with increased immunogenicity could promote DC maturation,
stimulate PBMC proliferation and activate cytotoxic T lymphocytes
(14). We were interested in the
immunogenic changes after MSC differentiation into IPCs in
vitro and, after transplantation into the diabetic model.
Therefore, we induced human umbilical cord MSCs (hUCMSCs) to
differentiate into IPCs in vitro and transplanted these
differentiated cells into diabetic mice to determine whether they
could combat against hyperglycemia. We investigated the
immunological properties of the differentiated IPCs in
vitro, including the expression of HLA-ABC, HLA-DR, the
costimulatory molecules CD40 and CD80, and stimulation of allogenic
PBMCs proliferation. On immunogenic changes of the IPCs after
transplantation into diabetic mice, we observed the immune cell
infiltration of the transplanted sites.
Materials and methods
Isolation and characterization of
hUCMSCs
Institutional Review Board approval was obtained for
all the procedures. Following the written informed consent of
parents, fresh human umbilical cords were obtained after birth from
the Second Affiliated Hospital of Jinan University. hUCMSCs were
isolated and cultured as previously described (16). The cells were cultured in Dulbecco's
modified Eagle's medium (DMEM)/F12 (Gibco, Grand Island, NY, USA)
with 10% fetal bovine serum (FBS), 100 U/ml penicillin G and 100
mg/ml streptomycin and maintained for 3–5 days in a humidified
incubator at 37°C with 5% CO2. Flow cytometry on an
Altra (Beckman Coulter, Fullerton, CA, USA) was performed to
analyze characteristic MSC markers including CD29, CD34, CD90F,
CD45, CD106 and HLA-DR (eBioscience, Inc., San Diego, CA, USA).
hUCMSCs were also characterized by differentiation toward
adipogenic and osteogenic lineages using previously described
protocols (Alizarin red and Oil Red O assay) by commercial kits
(Cyagen Biosciences Inc., Santa Clara, CA, USA).
Pancreatic β-cell differentiation
For pancreatic β-cell differentiation, hUCMSCs were
induced with a modified four-step protocol with the addition of
islet neogenesis-associated protein pentadecapeptide (INGAP-PP), as
previously described (17,18). Differentiation was induced by
treating the cells with CMRL medium supplemented with 10% FBS
(Gibco), 10 mM nicotinamide (Sigma-Aldrich, St. Louis, MO, USA),
and 4 nM activin A (R&D Systems, Inc., Minneapolis, MN, USA),
25 ng/ml recombinate EGF (Invitrogen-Biosource, Carslbad, CA, USA),
and 0.5 mM β-mercaptoethanol (Gibco) for 3 days. Then the cells
were treated with second stage induction medium for 5 days. The
second stage medium was CMRL medium supplemented with 10% FBS, 10
mM nicotinamide, 4 nM activin A, and 25 ng/ml recombinate EGF. At
the third induction stage, the cells were cultured in DMEM/F12
medium supplemented with 2% FBS, 10 mM nicotinamide 10 nM Exendin-4
(AnaSpec, Fremont, CA, USA), 10 µg/ml INGAP-pp (Sangon Biotech Co.,
Ltd., Shanghai, China) and 1X ITS (Gibco) for 7 days. An last
induction stage, the cells were treated with high-glucose DMEM
supplemented with 10 mM nicotinamide, 10 nM Exendin-4, 10 µg/ml
INGAP-pp and 1X ITS, and recombinant bFGF (Prospec, East Brunswick,
NJ, USA) for 7 days.
RT-qPCR analysis of pancreatic
developmental-related genes
hUCMSCs, day 14 IPCs, day 21 IPCs and human islets
were harvested for gene expression analysis. The total RNA was
isolated using TRIzol reagent (Takara Bio, Inc., Otsu, Japan), and
was reverse-transcribed into cDNA using a PrimeScript RT Reagent
kit with gDNA eraser (Takara) according to the manufacturer's
instructions. Quantitative gene expression of Pdx1, Ngn3, MafA,
NeuroD1, and insulin was determined using SYBR-Green Premix Ex Taq
(Takara) and performed using standard methods as previously
described (19). The primer sets
used were: Pdx-1 forward, 5′-ttgaacttgaccgagagacaca-3′ and reverse,
5′-cgcttcttgtcctcctcc ttt-3′; Ngn3 forward,
5′-cttcgcccacaactacatctg-3′ and reverse,
5′-ctgggagactggggagtagag-3′; NeuroD1 forward,
5′-tgaaagccctctgactgattg-3′ and reverse,
5′-cggtgcctgagaagattgat-3′; MafA forward,
5′-cttcagcaaggaggaggtcat-3′ and reverse,
5′-agttggcacttctcgctctc-3′; insulin forward,
5′-cagcctttgtgaaccaacac-3′ and reverse, 5′-cgggtcttgggtgtgtagaa-3′;
β-actin forward, 5′-ctgggac gacatggagaaaa-3′ and reverse,
5′-aaggaaggctggaagagtgc-3′.
Immunofluorescence staining for
pancreatic developmetal-related markers
hUCMSCs and day 21 IPCs were fixed in 4%
paraformaldehyde for 20 min and blocked with 0.2% Triton X-100
supplemented with 2% BSA for 1 h. Cells were then incubated with a
rabbit anti-human Pdx1 polyclonal antibody (1:100; cat. no.
sc-25403) and MafA polyclonal antibody (1:100; cat. no. sc-66958)
(both from Santa Cruz Biotechnology, Inc., Santa Cruz, CA, USA),
rabbit anti-human C-peptide polyclonal antibody (1:100; cat. no.
ab14181; Abcam, Cambridge, UK), goat anti-human Nkx6-1 polyclonal
antibody (1:200; cat. no. MBS622560; Invitrogen-Biosource) at 4°C
overnight. Then, cells were incubated with Rhodamine Red-conjugated
goat anti-rabbit IgG Rhodamine Red antibody (1:200; cat. no.
MBS235303; Invitrogen-Biosource) FITC-conjugated goat anti-rabbit
IgG (1:1,000; cat. no. sc-2012) and FITC-conjugated donkey
anti-goat IgG antibody (1:200; sc-2024) (both from Santa Cruz
Biotechnology, Inc.). The nuclei were counterstained with DAPI
(1:800), and cells were visualized under a fluorescence microscope
(Eclipse TE2000-U; Nikon GmbH, Düsseldorf, Germany).
Immune antigen expression
analysis
The immuno-phenotype (HLA-ABC, HLA-DR, CD40 and
CD80) of hUCMSCs and day 21 IPCs was examined by flow cytometry on
a flow cytometer (Epics Altra; Beckman Coulter) with specific
phycoerythrin or FITC-conjugated monoclonal antibodies
(eBioscience, Inc.) and performed as previously described (20). CellQuest software (BD Biosciences,
Franklin Lakes, NJ, USA) was used for data analysis. Results are
expressed as a percentage of positive cells or as mean relative
fluorescence intensity, obtained as a ratio between the mean
fluorescence intensity of cells stained with specific mAb and the
mean fluorescence intensity obtained with isotype control.
Purification of IPCs
Newport Green (NG) is a fluorescent dye specifically
binding to zinc. Insulin is stored in β cells as crystals
containing zinc, so NG can be used to label functional β cells and
NG diacetate (NG-AC) is suitable for live cell staining (21). Differentiated IPCs were washed twice
with phosphate-buffered saline (PBS), then incubated with 1–10 µM
NG-AC and 1 µl/ml Pluronic F127 at 37°C for 30 min. After washing
in PBS with 5% FBS, the cells were re-suspended and was subjected
to FACS analysis.
One-way mixed lymphocyte reaction
hPBMCs as effector cells were isolated from healthy
donors following informed consent. hUCMSCs at passage 3 or day 21
IPCs pretreated with 25 mg/ml mitomycin C (Roche Diagnostics,
Indianapolis, IN, USA) were used as stimulator cells. A total of
1×105 effector cells were co-cultured with
1×104 stimulator cells in 96-well plates. Effector cells
treated with PHA (5 mg/ml; Sigma-Aldrich) were used as a positive
control, and effector cells alone were used as a negative control.
After co-culture for 72 h, the proliferation of effector cells was
assayed with a cell counting kit (Dojindo Laboratories, Kumamoto,
Japan) and the OD at 450 nm was measured with a Bio-Rad 550
microplate reader (Bio-Rad, Tokyo, Japan). The SI was calculated
using the formula: SI = (co-cultured sample OD - stimulator
OD)/(negative control OD - blank OD).
Cell-mediated lympholysis (CML)
test
C57BL/6 mice were randomly divided into two groups,
hUCMSCs and IPCs group, 5 mice in each group. On day 1 and 6,
1×107 hUCMSCs or purified IPCs were peritoneally
injected to immune animals. On day 10, the mice were sacrificed.
Spleen was removed and made to single cell suspension at a density
of 1.0×107/ml, as effector cells. A total of
1×105 cells of splenocytes was incubated with a serial
titration of the same stimulating cells for 72 h at 37°C. The
cytotoxicity was evaluated by staining with Annexin V and PI, and
the apoptosis rate was analyzed by FACS.
Interleukin (IL)-2, IL-4 and
interferon (IFN)-γ secretion in CML
IL-2, IL-4 and IFN-γ secretion was tested with the
supernatants collected from the wells of CML plates. The assay
procedure followed instructions of the mouse ELISA kit (4A Biotech
Co., Ltd., Beijing, China). Absorbance was measured using a Bio-Rad
680 microplate reader (Bio-Rad) at 450 nm.
For peritoneal inflammatory cell infiltration test
male C57BL/6 mice with an initial body weight of 20–25 g were
randomly assigned into three groups: hUCMSCs (n=4),
1.0×107 hUCMSCs at passage 3 were injected into
peritoneal cavity; IPCs (n=4), 1.0×107 purified IPCs
were injected into peritoneal cavity; control group (n=4), 1 ml PBS
were intraperitoneally injected. Four-hour post-injection,
peritoneal cavity lavage was performed by injecting 8 ml PBS into
the cavity. Then the lavage was centrifuged at 1,000 × g for 5 min
and treated with red blood cell lysis buffer at room temperature
for 10 min. The cells were then stained with FITC anti-mouse CD3e
and PE anti-mouse CD45 and analyzed by FACS.
Cell transplantation and histological
analysis
C57B/L mice were purchased from the Experimental
Animal Center of Southern Medical University and housed under
specific pathogen-free conditions. All animal procedures were
approved by the Institutional Animal Care and Use Committee at
Shenzhen PKU-HKUST Medical Center. Male mice with an initial body
weight of 20–25 g were fasted for 12 h and then intraperitoneally
injected with streptozotocin (Sigma-Aldrich) at a single dose of
170 mg/kg. After 48 h, tail vein blood samples were obtained for
blood glucose (BG) measurements using a BG device (Roche Accu-Chek
III; Roche Diagnostics, Basel, Switzerland). Mice with a
non-fasting BG of 300 mg/dl for 3 consecutive days were considered
diabetic. The diabetic animals were randomly assigned based on the
transplanted cell types; 1.0×107 hUCMSCs or purified
IPCs were transplanted under the left kidney capsule, marked as
hUCMSCs (n=10) and IPCs group (n=10). Diabetic group (n=10) and
non-diabetic mice (n=6) were injected with the same amount of PBS,
as positive and negative control. Non-fasting BG levels were
measured at a certain time every 3 days post-transplantation until
day 30, and the body weight was monitored at the same time.
The left kidney was removed from mice in hUCMSCs
(n=3) and IPCs group (n=3) on day 15 and 30 post-transplantation,
and then fixed in 10% buffered formalin. Paraffin-embedded 5 µm
sections were routinely stained with hematoxylin and eosin. The
hematoxylin and eosin-stained kidney sections were analyzed by
counting the number of inflammatory cells infiltrating into the
left kidney in 10 high-power fields (×400) per sample (22).
Statistical analysis
Data were expressed as the mean ± standard deviation
(SD). Statistical analysis was performed using one-way analysis of
variance (ANOVA) to compare the difference among different groups.
Independent sample t-tests were used to statistically compare the
difference between two groups. Sequential data were compared using
repeated measures ANOVA. P<0.05 was considered to indicate a
statistically significant difference.
Results
Isolation and characterization of
hUCMSCs
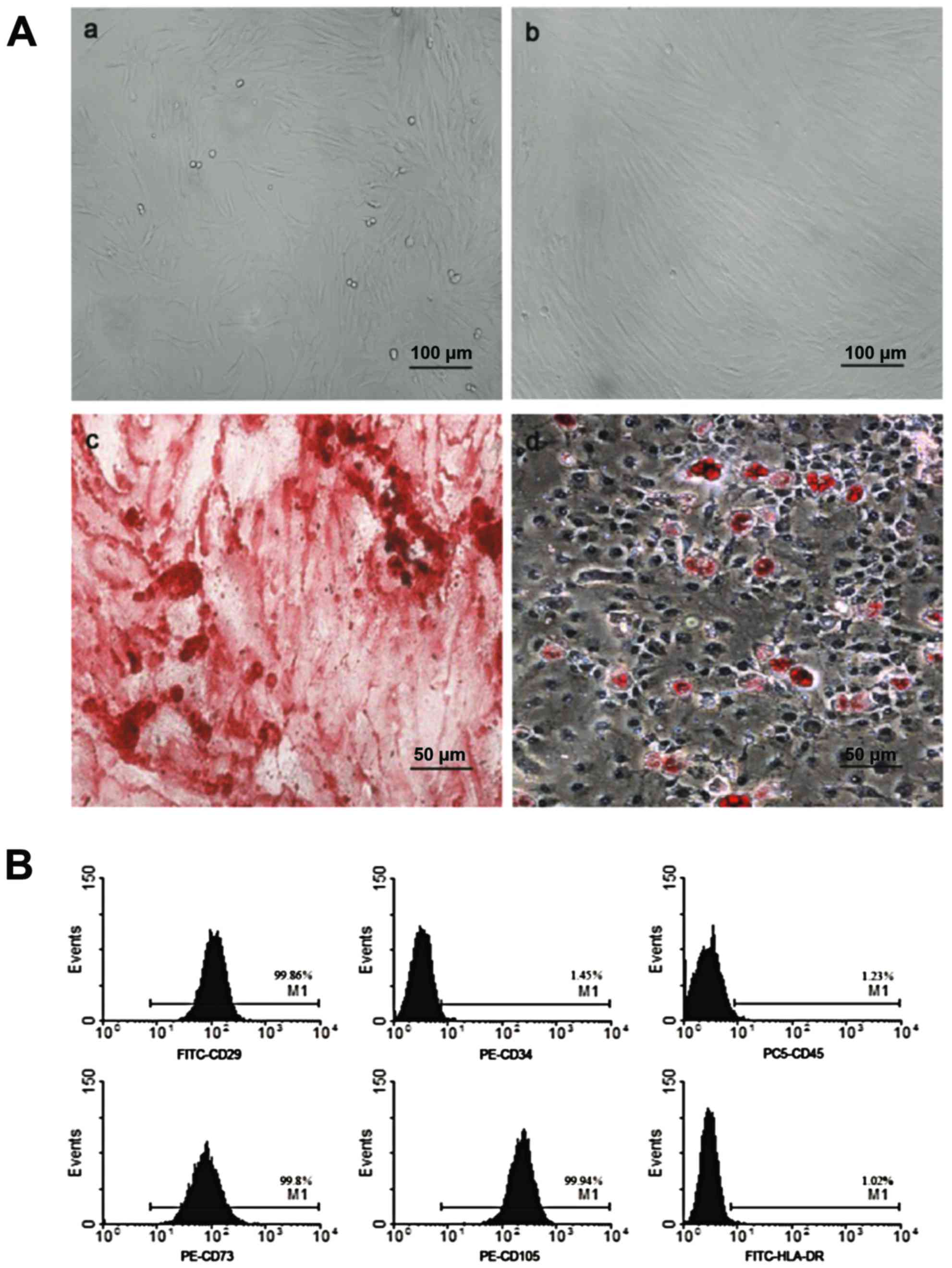
After purification, hUCMSCs were spindle-shaped and
attached to the plate during cell culture (Fig. 1A-a and b). Flow cytometric analysis
demonstrated that these cells were positive for CD29 (99.86%), CD73
(99.8%), CD105 (99.94%), but negative for CD45 (1.23%), CD34
(1.45%) and HLA-DR (1.02%), which was in accordance with MSC
characteristics as reported previously (Fig. 1B). After adipogenic induction, the
cells were stained positive for Oil Red O, showing lipid-filled
vesicles. After osteogenic induction, these cells displayed
osteogenesis, as shown by Alizarin red staining of calcium deposits
(Fig. 1A-c and d).
IPCs induction from hUCMSCs
Following the addition of INGAP-pp and certain small
molecules, modified four-step induction protocol was used to induce
IPCs from hUCMSCs. During induction, the cells started to become
polygonal and rhombic on day 14, then cells aggregated to grow,
became oval and formed a regular pavement-like morphology on day 21
(Fig. 2A). The maturity of
differentiated IPCs was investigated by immunofluorescent staining
and glucose stimulation test. In immunofluorescent staining,
differentiated IPCs expressed C-peptide, indicating the production
of insulin, not absorption from the medium (Fig. 2C). Furthermore, these cells also
expressed the mature β cell markers PDX1, NKX6.1, but no MAFA
(Fig. 2C), indicating moderate but
not full maturity of the differentiated IPCs. Flow cytometric
analysis showed that the rate of C-peptide expression of fully
differentiated IPCs was 12.53±0.92%, while almost no C-peptide
positive cells were found in hUCMSCs (Fig. 2D). This indicated that about
12.03±0.92% cells were successfully differentiated into IPCs. The
test of glucose stimulation showed that these induced IPCs secreted
a low level of C-peptide (48.32±0.39) with stimulation of low
glucose (5.5 mM) (Fig. 2E). After
incubation with high glucose (25 mM), C-peptide level in the
supernatant were 3-fold higher than that of low glucose (Fig. 2E). These results indicated that the
induced IPCs by our modified protocol could secrete insulin in a
glucose-regulated manner.
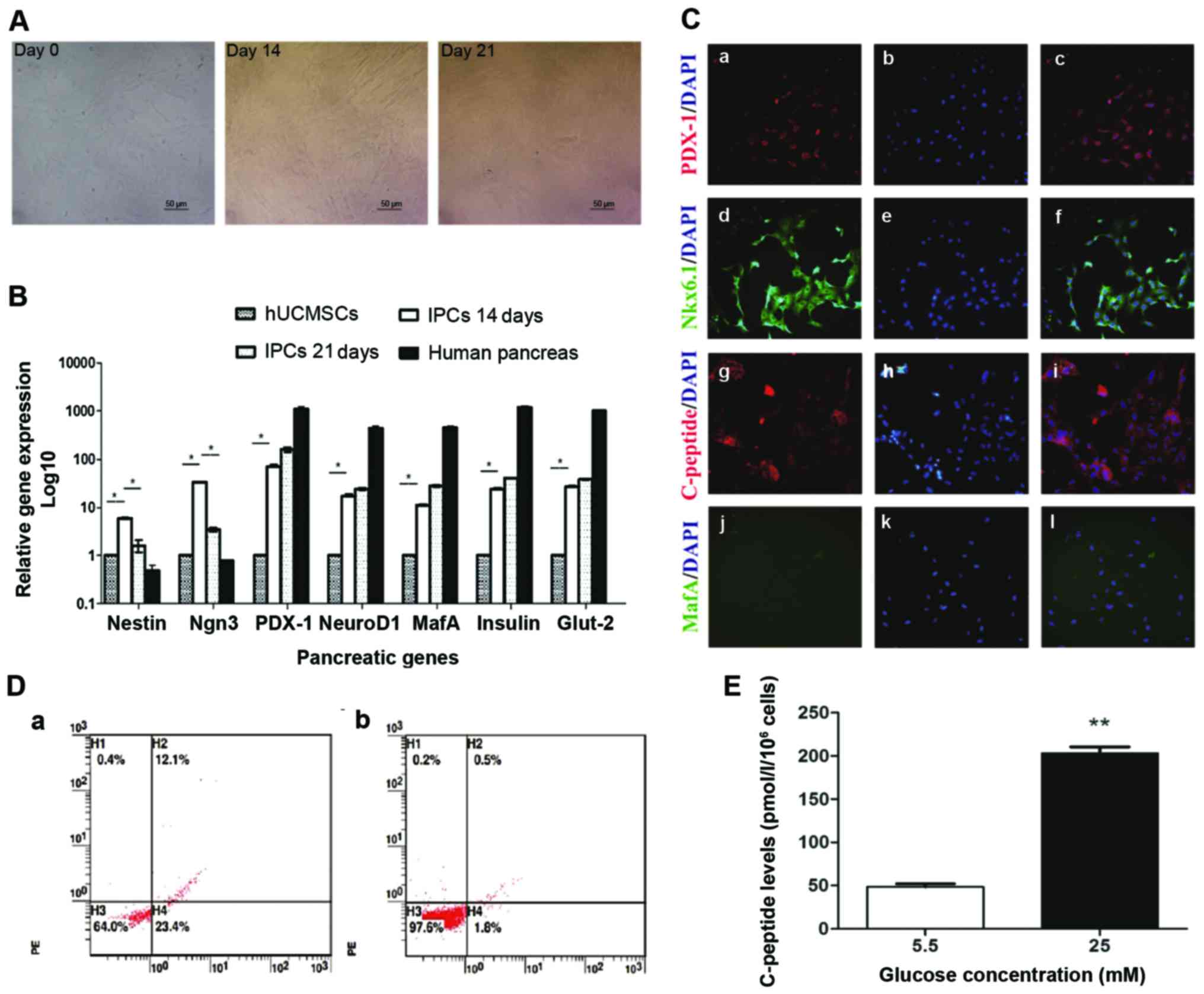 | Figure 2.IPCs induction and function analysis.
(A) Morphology changes during IPC induction from hUCMSCs on days 14
and 20; ×200 magnification. (B) mRNA levels of pancreatic
developmental-related genes. Total mRNA was extracted to evaluate
the expression of pancreatic genes (Nestin, Ngn3, Pdx1, NeuroD1,
MafA, insulin and Glut-2) by RT-qPCR. *P<0.05. (C) Induced IPCs
were stained positive for Pdx1, Nkx6.1 and C-peptide by
immunofluorescence staining; ×200 magnification. (D) Flow
cytometric detection of c-peptide expression on induced IPCs.
Induced IPCs on (a) day 21 and (b) isotype control. (E) Glucose
challenge test, day 21 IPCs were incubated with low glucose (5.5
mM) and high glucose (25 mM) for 3 h, then C-peptide release was
measured in the supernatants. *P<0.01. IPCs, insulin-producing
cells; hUCMSCs, human umbilical cord mesenchymal stem cells. |
RT-qPCR analysis of the levels of pancreatic
developmental gene mRNA. Induced IPCs were stained with NG and
sorted by FACS. Upon FACS analysis, 6–8% of the cells were selected
as positive for NG, no green fluorescence was observed in the
control group (data not shown). RT-qPCR analysis was performed to
investigate the changes of pancreatic developmental gene expression
during induction and differentiation. In induced IPCs, Nestin, a
marker for pancreatic development, was upregulated on day 14 and
then decreased on day 21; the same trend was also observed on
Ngn3-a pancreatic endocrine progenitor marker. Compared to hUCMSCs,
Ngn3 mRNA was significantly upregulated (P<0.05) on day 14, and
its level was reduced on day 21. There was an increased expression
of β-cell transcripts such as insulin, Pdx1, MafA, NeuroD1 and
Glut2 on days 14 and 21, but albeit much lower than the level of
adult human islets (Fig. 2B).
Immune antigen expression on IPCs and
lymphocyte proliferation assay in vitro
FACS showed that the IPCs expressed high levels of
HLA-ABC (97.93±1.71), and no HLA-DR, CD40 and CD80 in vitro,
which is similar to hUCMSCs without induction (Fig. 3A). In one-way mixed lymphocyte
reaction assay, the stimulation index of hPBMSCs after co-culturing
with hUCMSCs was 0.24±0.04, which was significantly different
compared with the positive control group (PHA) and negative control
group (auto-hPBMCs) (P<0.05). The index of stimulation of IPC
co-culture group was 0.79±0.06, which was lower compared with the
positive control group, while no difference was found with the
negative control group (P>0.05) (Fig.
3B), indicating that induced IPCs had low immunogenicity in
vitro, but did not have the immunosuppressive property as
hUCMSCs did.
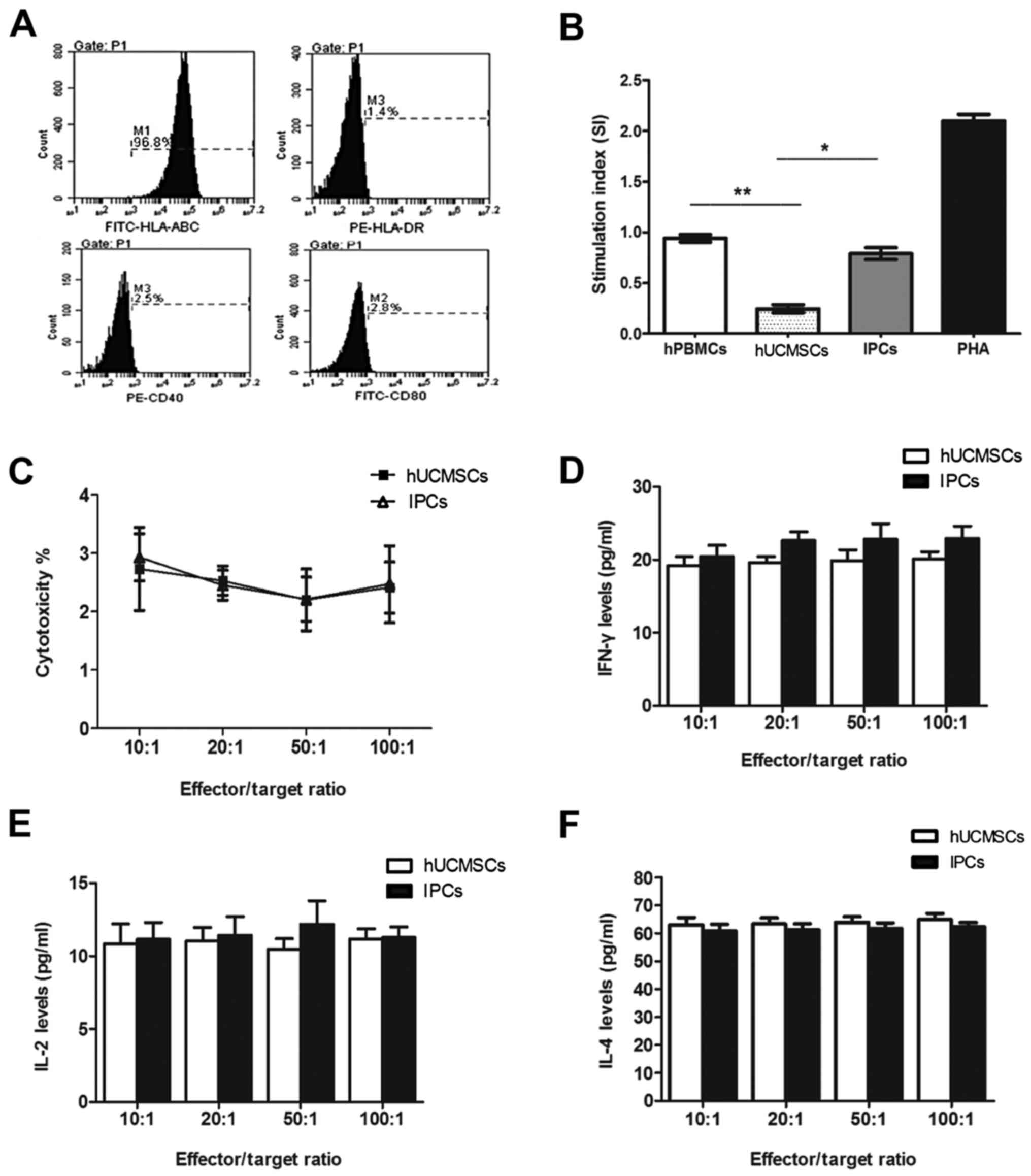 | Figure 3.In vitro immunological
characteristics of induced IPCs. (A) FACS shows that induced IPCs
expressed MHC-I and did not express HLA-DR, CD40 and CD80. (B)
Allogenic PBMCs were co-cultured with hUCMSCs or IPCs for 72 h. No
proliferation was observed in the IPCs group compared with the
PBMCs auto-proliferation and PHA (positive) group. *P<0.05,
**P<0.01. (C) Splenocytes collected from recipients were
considered as effector cells, and then co-cultured with IPCs
(target cell to effector cell ratio: 1:10, 1:20, 1:50 and 1:100)
for 72 h. Percentage of apoptotic MSCs were evaluated by Annexin
V-APC/PI staining and flow cytometry. (D-F) IL-2, IL-4 and IFN-γ
secretion in CML supernatants at different effector/target ratio.
IPCs, insulin-producing cells; hUCMSCs, human umbilical cord
mesenchymal stem cells; MSCs, mesenchymal stem cells; CML,
cell-mediated lympholysis. |
Cell-mediated lysis test
In order to observe sensitization of the host
lymphocytes by the induced IPCs, we pre-sensitized the mice with
hUCMSCs or IPCs twice, on days 1 and 6. Then splenocytes were
isolated from the pre-sensitized mice and co-cultured with the same
cells for sensitization with different ratios. No cytolysis
difference was observed among groups with different effector/target
ratio in either hUCMSC or IPC co-culturing groups (P>0.05)
(Fig. 3C). When cells co-cultured in
the highest effector/target ratio (100:1), the apoptotic rates of
hUCMSCs and IPCs were 2.4±0.44 and 2.47±0.66% respectively, with no
difference (P>0.05) (Fig. 3C).
This indicated that hUCMSCs were low immunogenic and
immunosuppressive, thus could not activate memory T cells and
cytolysis T cells in vivo. After the in vitro
induction, the purified IPCs did not activate immune cells or
elicit cytolysis in vivo due to its hypo-immunogenicity.
Cytokine secretion in CML
IFN-γ, IL-2 and IL-4 are Th1/Th2 cytokines which are
very important in mediating and regulating immunity. We tested
these cytokines in the supernatants of the co-cultured cells in
CML. The results showed that there was no significant difference of
cytokine secretion between IPCs and hUCMSC co-culture groups at
different ratios (Fig. 3D-F). These
results suggested that hUCMSCs and induced IPCs could not activate
immune cells and no Th1/Th2 cytokine secretion changes occurred
when transplanted the second time.
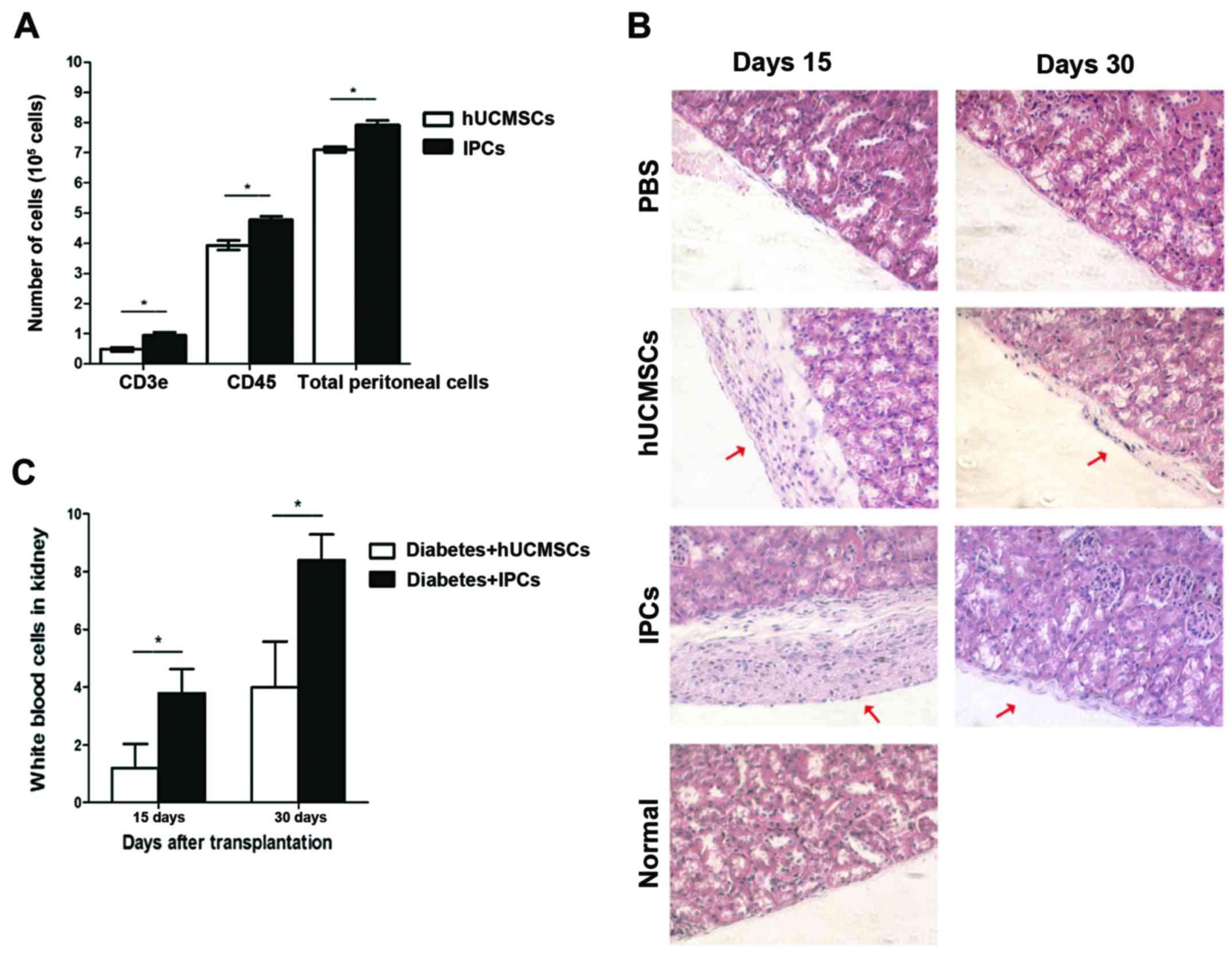
Immune cells in peritoneal lavage
To determine the acute rejection of hUCMSCs and
IPCs, cells were injected into the peritoneal cavity. The
peritoneal lavage was extracted and cells positive for leukocyte
(CD45+) and T lymphocytes (CD3e+) were
analyzed by FACS. Total cells in peritoneal lavage extracted from
the hUCMSCs group were 7.10±0.55×105, 55% of the cells
was CD45+, and 6.8% was CD3e+. Compared to
the hUCMSCs group, an increased number of cells were found in
peritoneal lavage from the IPCs injection group (P<0.05), total
cells were 7.92±0.09×105, in which 60% of the cells
expressed CD45+ and 12% were CD3e+, which was
much higher than that in the hUCMSCs group (Fig. 4A) (P<0.05). This indicates that
induced IPCs attract immune cells to infiltrate into the injection
sites, showing some immunogenicity in vivo.
Histopathological analysis
On day 15, no leukocyte infiltration was observed in
left kidney of hUCMSC-transplanted group, while in the IPCs groups,
a small number of immune cells was found in the transplanted region
(P<0.05); on day 30, in the IPCs group, more immune cells
infiltrated to the left kidney compared to day 15 (P<0.05), but
in the hUCMSCs group, only few immune cells were observed (Fig. 4B and C). This result suggests that
the immunogenicity of induced IPCs was enhanced after
transplantation in vivo, which could attract leukocyte
infiltrate to the transplanted sites.
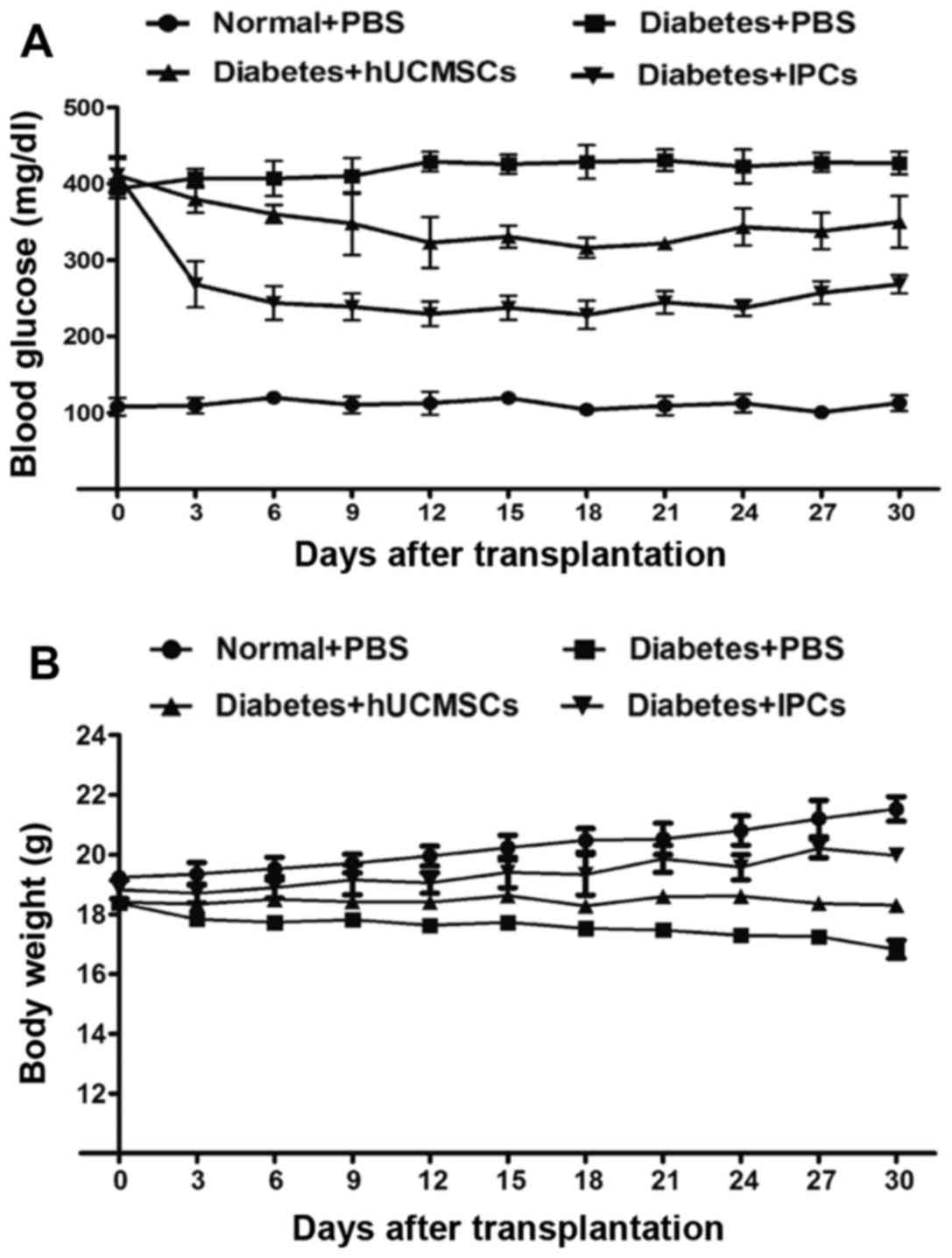
Reversal of hyperglycemia after IPCs
transplantation
After IPCs administration, the BG levels decreased
rapidly to 229±16 mg/dl by day 12, and reached 268±12 mg/dl by day
30 (Fig. 5A). In the
hUCMSC-transplanted group, the BG decreased slightly to 323±33
mg/dl by day 12, and eventually returned to 350±34 mg/dl, and this
ability to reduce BG may be due to tissue repair and paracrine
effects. The weight of diabetic mice was improved to some extent in
both hUCMSC and IPC treated groups by the end of the study
(Fig. 5B). These data showed that
the induced IPCs improved the conditions of diabetes.
Discussion
MSCs can be derived from different tissues,
including adipose, bone marrow or umbilical cord (23). MSCs were considered low immunogenic
due to their low expression of immune antigens on the surface
(24,25), which makes them hard to be recognized
by T lymphocytes (6,10). In both in vitro and in
vivo settings, MSCs can inhibit T-lymphocyte proliferation,
suppress proliferation and activation of NK cells and DCs, and
reduce the cytokine secretion of T cells and NK cells. They also
lower the immune rejections, induce immune tolerance, and thus
improve the success rate of organ transplantation (26). However, immunogenicity of MSCs may be
enhanced due to the differentiation process and transplantation
microenvironments (9–14). After differentiation or
transplantation, MSCs may elicit immune responses, thus reducing
the survival and further differentiation of the cells, and finally
hamper the clinical application of MSCs. In the present study, we
successfully induced hUCMSCs to differentiate into functional IPCs
by modified induction protocol with the addition of INGAP-pp. Then
we investigated the immunological properties of induced IPCs in
vitro and immune responses after transplantation into diabetic
mice, in order to explore the effects on MSC immunogenicity of
induction process and further provide basis for future clinical
treatments of diabetes.
Induced IPCs expressed high levels of HLA-ABC, but
no HLA-DR and co-stimulatory molecules CD40 and CD80. MHC molecules
selectively recognize and bind with antigen peptide, and help
presenting antigens. They bind with naïve T cells and form first
signal of T-cell activation. Co-stimulatory molecules CD80 and CD40
produce second signal of T-cell activation. IPCs failed to present
antigen to CD4+ cells due to absence of MHC-II on the
surface. In one-way, mixed lymphocyte reaction assay
antigen-presenting cells (APCs) in allogenic hPBMCs can indirectly
present alloantigen to the host lymphocytes, but we did not find
allogenic hPBMC proliferation when co-cultured with induced IPCs.
This indicated that induced IPCs could not activate naïve T cells
in vitro even with the assistance of exogenous APCs.
Therefore, induction of IPCs in vitro did not change the
immune phenotype and maintained hypo-immunogenic, which makes them
escape the presentation of antigen and fail to activate naïve T
cells. Similarly, hUCMSCs were induced to differentiate into
hepatocyte-like cells, similar to its undifferentiated progenitors,
it did not express MHC-II molecules and significantly inhibited
lymphocyte proliferation (27). MSCs
from rabbit bone marrow did not express MHC-II molecules after
osteogenesis in vitro, and failed to stimulate allogenic
PBMC proliferation and cytolytic T-cell activation, but increased
secretion of anti-immunological cytokine IL-10 (28).
In the cell-mediated lysis assay, when hUCMSCs or
IPCs were co-cultured with present-sized splenocytes, there was no
significant difference of the apoptotic rate between the IPCs and
hUCMSCs group. This indicates that IPCs could not activate CTLs and
memory T cells though the expression of MHC-I molecules in the
surface of IPCs was higher than that of hUCMSCs. Liu et al
(28) transplanted induced
osteoblasts from MSCs into rabbits, and observed expression of
MHC-II molecule on day 7 post-transplantation, but the transplanted
cells survived for 28 days. When fibroblast was transplanted to the
same host for the second time, no rejection occurred to the
transplanted fibroblasts, indicating no activation of CTLs and
memory T cells (28). Memory T cell
production and activation needs the assistance of CD4+ T
cells. Studies demonstrated that CD4+ T cells did not
activate and there was no upregulation of IFN-γ and IL-2 secretion
after intravenous injection of allogenic MSCs. Our data were in
accordance with those results. In our study, compared to
undifferentiated hUCMSCs, Th1 cytokines (IL-2 and IFN-γ) and Th2
cytokines (IL-4) did not change in differentiated IPCs co-culture
group, indicating that the balance between Th1 and Th2 was not
broken by induced IPCs and the activation of T cells (29).
Differentiated IPCs exhibit low immunogenicity in
vitro, but how they behave after transplantation to the host
remains to be determined. Additionally, whether they elicit
inflammatory responses and cause acute or chronic rejection should
be investigated. In the present study, we found leukocyte
infiltration to the peritoneal lavage and a slight upregulation of
T lymphocytes 4 h post-IPC injection to the peritoneal cavity. This
may be due to the upregulation of MHC-I in differentiated IPCs,
leading to the activation of the monocyte-macrophage system (innate
immunity) and the infiltration of immune cells. Similarly,
histopathological analysis revealed that a few IPCs were found to
be alive on day 14 after transplantation under the capsule of the
left kidney. Leukocyte infiltration to the transplanted sites was
observed in IPCs but hUCMSCs transplanted mice. On day 30, very few
IPCs survived but elevated inflammatory cells infiltration was seen
in the transplanted region. On the contrary, almost no immune cell
infiltration was found in hUCMSCs group. The above demonstrated
that IPCs became immunogenic after transplantation, which might be
the consequences of the disease microenvironment. Accordingly, BG
reduced rapidly after IPCs transplantation into diabetic mice, but
eventually returned to hyperglycemia again due to the loss of
transplanted IPCs. The possible explanation may be the enhancement
of IPC immunogenicity by the disease microenvironment.
Our previous data demonstrated that allogenic bone
marrow-derived MSCs continued to differentiate into IPCs after
transplantation under pancreatic capsule of diabetic rats. However,
with the occurrence of differentiation, MHC-II expression was
positive on MSCs and alloantibodies were detected in the serum,
indicating that differentiation initiates an immune ‘switch’ that
alters the immune characteristics of MSCs in vivo (30). Huang et al found that MSCs
started to express myocardiocyte-specific markers and also the
MHC-I and MHC-II after transplanted to infracted myocardium
(12). Microenvironment in the
disease recipients could promote differentiation of MSCs, and
change its immunopreviliged state in vivo (12).
It has been demonstrated that the body was in an
inflammatory state in the STZ-induced diabetes model. The secretion
of inflammatory cytokines possibly induced immune antigens
expression in the surface of IPCs, thus eliciting immune rejection.
IFN-γ has been demonstrated to induce MHC-I and MHC-II expression
on the surface of MSCs in vitro and in vivo (31–33). To
clarify the underlying mechanism of immunological changes after IPC
differentiation, the relationship between disease immune
microenvironment and immune characteristic alteration should be
examined.
In conclusion, although the immunogenicity of IPCs
derived from hUCMSCs was not altered in vitro, they can
become immunogenic by the interaction with disease microenvironment
and induce inflammatory responses and recruit immune cell
infiltration into the transplanted sites.
Acknowledgements
This study was supported by the National 973 Special
Plan of China (no. 2007CB516811), the National Natural Science
Foundation of China (no. 30772042), the Natural Science Foundation
of Guangdong (no. 6027540) and the Science and Technology Profect
of Shenzhen (no. 201001005).
References
|
1
|
Shapiro AMJ, Lakey JRT, Ryan EA, Korbutt
GS, Toth E, Warnock GL, Kneteman NM and Rajotte RV: Islet
transplantation in seven patients with type 1 diabetes mellitus
using a glucocorticoid-free immunosuppressive regimen. N Engl J
Med. 343:230–238. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Saidi RF: Current status of pancreas and
islet cell transplantation. Int J Organ Transplant Med. 3:54–60.
2012.PubMed/NCBI
|
|
3
|
Xie H, Wang Y, Zhang H, Qi H, Zhou H and
Li FR: Role of injured pancreatic extract promotes bone
marrow-derived mesenchymal stem cells efficiently differentiate
into insulin-producing cells. PLoS One. 8:e760562013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Prabakar KR, Domínguez-Bendala J, Molano
RD, Pileggi A, Villate S, Ricordi C and Inverardi L: Generation of
glucose- responsive, insulin-producing cells from human umbilical
cord blood-derived mesenchymal stem cells. Cell Transplant.
21:1321–1339. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Gabr MM, Zakaria MM, Refaie AF, Ismail AM,
Abou-El-Mahasen MA, Ashamallah SA, Khater SM, El-Halawani SM,
Ibrahim RY, Uin GS, et al: Insulin-producing cells from adult human
bone marrow mesenchymal stem cells control streptozotocin-induced
diabetes in nude mice. Cell Transplant. 22:133–145. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Uccelli A, Moretta L and Pistoia V:
Mesenchymal stem cells in health and disease. Nat Rev Immunol.
8:726–736. 2008. View
Article : Google Scholar : PubMed/NCBI
|
|
7
|
Poncelet AJ, Vercruysse J, Saliez A and
Gianello P: Although pig allogeneic mesenchymal stem cells are not
immunogenic in vitro, intracardiac injection elicits an immune
response in vivo. Transplantation. 83:783–790. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
English K, French A and Wood KJ:
Mesenchymal stromal cells: facilitators of successful
transplantation? Cell Stem Cell. 7:431–442. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lohan P, Coleman CM, Murphy JM, Griffin
MD, Ritter T and Ryan AE: Changes in immunological profile of
allogeneic mesenchymal stem cells after differentiation: should we
be concerned? Stem Cell Res Ther. 5:992014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Le Blanc K, Tammik C, Rosendahl K,
Zetterberg E and Ringdén O: HLA expression and immunologic
properties of differentiated and undifferentiated mesenchymal stem
cells. Exp Hematol. 31:890–896. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Liu CT, Yang YJ, Yin F, Wang X, Yu XH,
Wang QH, Wang XL and Xie M: The immunobiological development of
human bone marrow mesenchymal stem cells in the course of neuronal
differentiation. Cell Immunol. 244:19–32. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Huang XP, Sun Z, Miyagi Y, Kinkaid H
McDonald, Zhang L, Weisel RD and Li RK: Differentiation of
allogeneic mesenchymal stem cells induces immunogenicity and limits
their long-term benefits for myocardial repair. Circulation.
122:2419–2429. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Technau A, Froelich K, Hagen R and
Kleinsasser N: Adipose tissue-derived stem cells show both
immunogenic and immunosuppressive properties after chondrogenic
differentiation. Cytotherapy. 13:310–317. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Chen X, McClurg A, Zhou GQ, McCaigue M,
Armstrong MA and Li G: Chondrogenic differentiation alters the
immunosuppressive property of bone marrow-derived mesenchymal stem
cells, and the effect is partially due to the upregulated
expression of B7 molecules. Stem Cells. 25:364–370. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Dhingra S, Li P, Huang XP, Guo J, Wu J,
Mihic A, Li SH, Zang WF, Shen D, Weisel RD, et al: Preserving
prostaglandin E2 level prevents rejection of implanted allogeneic
mesenchymal stem cells and restores postinfarction ventricular
function. Circulation. 128:(Suppl 1). S69–S78. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Seshareddy K, Troyer D and Weiss ML:
Method to isolate mesenchymal-like cells from Wharton's Jelly of
umbilical cord. Methods Cell Biol. 86:101–119. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wang HS, Shyu JF, Shen WS, Hsu HC, Chi TC,
Chen CP, Huang SW, Shyr YM, Tang KT and Chen TH: Transplantation of
insulin-producing cells derived from umbilical cord stromal
mesenchymal stem cells to treat NOD mice. Cell Transplant.
20:455–466. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Li J, Wang Y, Yu X, Chen H, Wu Y, Han X,
Guo X, Zhang C, Chen Q, Chen J, et al: Islet neogenesis-associated
protein-related pentadecapeptide enhances the differentiation of
islet-like clusters from human pancreatic duct cells. Peptides.
30:2242–2249. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Pfaffl MW: A new mathematical model for
relative quantification in real-time RT-PCR. Nucleic Acids Res.
29:e452001. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Morandi F, Levreri I, Bocca P, Galleni B,
Raffaghello L, Ferrone S, Prigione I and Pistoia V: Human
neuroblastoma cells trigger an immunosuppressive program in
monocytes by stimulating soluble HLA-G release. Cancer Res.
67:6433–6441. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Lukowiak B, Vandewalle B, Riachy R,
Kerr-Conte J, Gmyr V, Belaich S, Lefebvre J and Pattou F:
Identification and purification of functional human beta-cells by a
new specific zinc-fluorescent probe. J Histochem Cytochem.
49:519–528. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ye J, Liao YT, Jian YQ, Zhang XD, Wei P,
Qi H, Deng CY and Li FR: Alpha-1-antitrypsin for the improvement of
autoimmunity and allograft rejection in beta cell transplantation.
Immunol Lett. 150:61–68. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wang HW, Lin LM, He HY, You F, Li WZ,
Huang TH, Ma GX and Ma L: Human umbilical cord mesenchymal stem
cells derived from Wharton's jelly differentiate into
insulin-producing cells in vitro. Chin Med J (Engl). 124:1534–1539.
2011.PubMed/NCBI
|
|
24
|
Casado JG, Gomez-Mauricio G, Alvarez V,
Mijares J, Tarazona R, Bernad A and Sanchez-Margallo FM:
Comparative phenotypic and molecular characterization of porcine
mesenchymal stem cells from different sources for translational
studies in a large animal model. Vet Immunol Immunopathol.
147:104–112. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lee JM, Jung J, Lee HJ, Jeong SJ, Cho KJ,
Hwang SG and Kim GJ: Comparison of immunomodulatory effects of
placenta mesenchymal stem cells with bone marrow and adipose
mesenchymal stem cells. Int Immunopharmacol. 13:219–224. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Shi Y, Hu G, Su J, Li W, Chen Q, Shou P,
Xu C, Chen X, Huang Y, Zhu Z, et al: Mesenchymal stem cells: A new
strategy for immunosuppression and tissue repair. Cell Res.
20:510–518. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhao Q, Ren H, Li X, Chen Z, Zhang X, Gong
W, Liu Y, Pang T and Han ZC: Differentiation of human umbilical
cord mesenchymal stromal cells into low immunogenic hepatocyte-like
cells. Cytotherapy. 11:414–426. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Liu H, Kemeny DM, Heng BC, Ouyang HW,
Melendez AJ and Cao T: The immunogenicity and immunomodulatory
function of osteogenic cells differentiated from mesenchymal stem
cells. J Immunol. 176:2864–2871. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Amirzargar A, Lessanpezeshki M, Fathi A,
Amirzargar M, Khosravi F, Ansaripour B and Nikbin B: TH1/TH2
cytokine analysis in Iranian renal transplant recipients.
Transplant Proc. 37:2985–2987. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Sadeghi M, Daniel V, Naujokat C, Schmidt
J, Mehrabi A, Zeier M and Opelz G: Evidence for IFN-γ up- and IL-4
downregulation late post-transplant in patients with good kidney
graft outcome. Clin Transplant. 21:449–459. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Stubbendorff M, Deuse T, Hua X, Phan TT,
Bieback K, Atkinson K, Eiermann TH, Velden J, Schröder C,
Reichenspurner H, et al: Immunological properties of extraembryonic
human mesenchymal stromal cells derived from gestational tissue.
Stem Cells Dev. 22:2619–2629. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhang X, Tang T, Shi Q, Fernandes JC and
Dai K: The immunologic properties of undifferentiated and
osteogenic differentiated mouse mesenchymal stem cells and its
potential application in bone regeneration. Immunobiology.
214:179–186. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Elias D, Prigozin H, Polak N, Rapoport M,
Lohse AW and Cohen IR: Autoimmune diabetes induced by the beta-cell
toxin STZ. Immunity to the 60-kDa heat shock protein and to
insulin. Diabetes. 43:992–998. 1994. View Article : Google Scholar : PubMed/NCBI
|