Introduction
Natural killer (NK) cells are innate immune cells
whose function is critical in the first-line of defense against
different types of pathogens, including viruses, bacteria and fungi
(1,2). Furthermore, NK cells are associated
with immune surveillance against tumors, hinder the dissemination
of metastatic tumors, and perform essential functions in regulating
autoimmune diseases (3,4). Unlike T and Blymphocytes, NK cells are
constitutively able to kill target cells without prior
sensitization; additionally, NK cells exhibit a variety of
activating and inhibitory receptors that allow for activation and
inhibition signaling, which may determine the cytotoxicity of NK
cells (5).
NK cells are present in all lymphoid organs and
mature NK cells may be detected in the liver, peripheral blood and
spleen (6). NK1.1 has been used to
distinguish NK cells in specific murine strains, including C57Bl/6
(7). Cluster of differentiation
(CD)-49b may associate with DX5 antibody and is also a well-known
NK cell marker in different mouse strains, uch as BALB/c (8). The isolation of purified and newly
separated NK cells poses a challenge as NK cells only constitute
~2.5% of mouse splenocytes (9).
Previous studies have determined multiple approaches
for purifying splenic NK, such as the methods of NK cell isolation
proposed by Ravnik et al (10) and Patel and Linna (11), which were based on the
differentiation of cells via density gradient centrifugation with
continuous or discontinuous percoll gradients. However, flow
cytometry has indicated that <40% of density-separated cells
were NK1.1+CD3ε−, particularly from spleens
of C57BL/6 mice (10,11). Advancement in technology has allowed
for the development of the novel method, magnetic-activated cell
sorting (MACS). MACS sorting is a popular method applied in areas
concerning immunology, cancer research, neuroscience, and stem cell
research. Through this approach, cells are positively or negatively
separated, depending on specific antigens present (12). For NK cell sorting, positive
selection may be gaged by selecting antibodies against NKp46 or
CD49b (DX5) and negative selection may be achieved for naïve NK
cell purification using commercially available kits.
Different conclusions and several problems have been
identified in the purification of murine NK cells as the result of
using different commercial kits (13). For that reason, an extensive
comparative study of four different NK cells isolation kits based
on MACS separation in C57Bl/6 mice was performed in the present
study. The present study recognized that NK cells are short-lived
and IL-2-dependent in vitro. Thus, the concentration of IL-2
was an important factor to consider when evaluating the effects of
various drugs on NK cells. Consequently, the approach of the
present study required freshly isolated NK cells and evaluated the
purity and viability of NK cells in the absence or presence of
different concentration of IL-2 for 24, 48 or 72 h.
Materials and methods
Animals
A total of 30 female 6-8-week-old C57BL/6 mice
weighing 18–20 g were obtained from Harlan Slac Laboratory Animals
Co., Ltd. (Shanghai, China). Mice were maintained at China Medical
University in a pathogen-free animal house. Mice were housed in a
temperature (21±1°C), humidity (55±10%), and 12-h light/dark cycle
controlled room. Food and water were available ad libitum.
All experiments with animals were performed in accordance with the
Guide for the Care and Use of Laboratory Animals as approved by the
China National Institutes of Health.
Cell culture
Freshly isolated NK cells using the Stemcell Mouse
NK Cell Isolation kit (catalogue no. 19855; STEMCELL Technologies,
Inc., Cambridge, UK) were cultured in Roswell Park Memorial
Institute-1640 medium (Biological Industries, Kibbutz Beit-Haemek,
Israel) supplemented with 10% fetal bovine serum (FBS; Biological
Industries), 2 mM L-glutamine (Lonza, Shanghai, China), 50 µM
β-mercapto ethanol (Sigma-Aldrich; Merck Millipore, Darmstadt,
Germany) and antibiotics (100 U/ml penicillin, 100 µg/ml
streptomycin and 100 µg/ml kanamycin) (HyClone; GE Healthcare Life
Sciences, Logan, UT, USA) at a density of 5×105 cells/ml
in 96-well flat plates. NK cells were cultured for 24, 48 or 72 h
in the absence or presence of IL-2 (PeproTech, Inc., Rocky Hill,
NJ, USA). At the designated time points, NK cells were assessed for
viability and surface phenotype using flow cytometry (FCM). Cells
were maintained in a humidified atmosphere containing 5%
CO2 and 95% air at 37°C.
Sample preparation for magnetic
activated cell sorting (MACS)
Mice were anesthetized by intraperitoneal injection
of Inactin (thiobutabarbital, 100 mg/kg; Sigma-Aldrich; Merck
Millipore) and sacrificed by cervical dislocation, and spleens were
removed by mechanical dissociation and placed on a petri dish
containing pre-cooled phosphate-buffered saline (PBS) supplemented
with 2% FBS as described previously (12). Spleens were mashed using a
moisturized cell strainer (70-µm nylon mesh) using a plunger and a
cell strainer positioned above a 50-ml conical tube was used to
transfer the single cell suspension through the strainer into the
tube. The petri dish and the strainer were rinsed with pre-cooled
PBS supplemented with 2% FBS. Erythrocytes were lysed with ammonium
chloride potassium lysing buffer. Subsequently, murine splenocytes
were transferred through another 70-µm cell strainer into a new
50-ml conical tube. Cell numbers were counted by trypan blue
exclusion. Following the different cell isolation procedures, NK
cell fractions were harvested, visualized under a light microscope
and the number of cells counted on a hemocytometer using trypan
blue (the percentage of viable cells=total cells-blue cells/total
cells). Subsequently, FCM analysis was performed (as described in
Flow cytometry) to determine the percentage of NK cells (the
percentage of NK cells=all cells at lymphocyte locations/total
cells) vs. contaminating lymphocytes (the percentage of
contaminating lymphocytes=all cells at granulocyte locations/total
cells) using BD FACS DIVA version 6.0 software (BD Biosciences, San
Jose, CA, USA).
Miltenyi NK Cell Isolation Kit II
The NK Cell Isolation kit II was used according to
the manufacturer's instructions (130-096-892) by using an LS column
on the Midi MACS separator (both Miltenyi Biotec GmbH, Bergisch
Gladbach, Germany). Splenocyte suspensions (1×108
cells/ml) were centrifuged at 300 × g for 10 min at 4°C and
resuspended in 400 µl isolation buffer (containing PBS, pH 7.2,
0.5% bovine serum albumin and 2 mM ethylenediaminetetraacetic
acid). Subsequently, 100 µl NK cell biotin-antibody mixture was
added, and the mixture was incubated for 5 min in the refrigerator
(2–8°C). Following washing with isolation buffer, splenocytes were
resuspended in 800 µl isolation buffer. Subsequently, 200 µl
anti-biotin microbeads were added and cells were incubated for an
additional 10 min in the refrigerator (2–8°C). During the
incubation period, the LS column was prepared by rinsing with 3 ml
of isolation buffer. Cell suspensions were loaded onto the column
and the flow-through containing unlabeled cells was collected,
representing the enriched NK cells. To obtain more NK cells, the
column was subsequently washed with 3 ml isolation buffer again and
the flow-through cells were collected.
Miltenyi CD49b (DX5) positive
selection kit
CD49b (DX5) microbeads were used according to the
manufacturer's instructions (130-052-501) using an MS Column on the
MiniMACS separator (Miltenyi Biotec, GmbH). Splenocyte suspensions
at a density of 1×108 cell/ml were centrifuged at 300 ×
g for 10 min at 4°C and resuspended in 900 µl isolation buffer. A
total of 100 µl CD49b (DX5) microbeads were added and cells were
incubated for 15 min at 4–8°C. Following washing with isolation
buffer, splenocytes were resuspended in 500 µl isolation buffer and
the MS column was prepared by rinsing with 500 µl isolation buffer.
The cell suspension was applied onto the column and cells were
washed with isolation buffer three times. Subsequently, the column
was removed from the separator and placed on a 15 ml conical tube.
A total of 1 ml buffer was pipetted onto the column and the
fraction was immediately flushed out using magnetically labeled
cells by firmly applying the plunger supplied with the column,
providing the purified CD49b+ cells.
Stemcell CD49b Positive Selection
kit
The Mouse CD49b Positive Selection kit was performed
according to the manufacturer's instructions (18755; Stemcell
Technologies UK Ltd., Cambridge, UK) using Falcon 5-ml polystyrene
round-bottom tubes (352058; BD Biosciences) on a purple EasySep
magnet (18000; Stemcell Technologies UK Ltd.). Splenocyte
suspensions (1×108 cells/ml) were centrifuged at 300 × g
for 10 min at 4°C and resuspended in 1 ml sorting buffer (PBS and
2% FBS with 1 mM ethylenediaminetetraacetic acid) in a 5 ml (12×75
mm) polystyrene tube. Subsequently, 50 µl EasySep mouse CD49b
phycoerythrin (PE) labeling reagent was added, and the suspension
was incubated at room temperature (15–25°C) for 15 min. A total of
100 µl EasySep PE selection cocktail was added and the cells were
incubated at room temperature for 15 min. Subsequently, 50 µl
nanoparticles were added and the cells were incubated at room
temperature for 10 min. Finally, the cell suspensions were made to
a total volume of 2.5 ml by adding the recommended medium, and were
placed into the magnet for 5 min. The supernatant fraction was
poured off three times following 5 min separations in the magnet.
Magnetically-labeled cells (positively selected cells) remained
inside the tube; the tube was removed from the magnet and cells
were resuspended in 1 ml desired medium.
Stemcell Mouse NK Cell Isolation
kit
The Mouse NK Cell Isolation kit was used according
to the manufacturer's instructions (19855; Stemcell Technologies UK
Ltd.) using Falcon 5-ml polystyrene round-bottom tubes on the
EasySep magnet. Splenocyte suspensions at a density of
1×108 cells/ml were centrifuged at 300 × g for 10 min at
4°C and resuspended in 1 ml of recommended medium in a 5-ml
polystyrene tube. A total of 50 µl EasySep mouse NK cell isolation
cocktail was added, and cells were incubated at room temperature
(15–25°C) for 10 min. Subsequently, 100 µl EasySep Streptavidin
RapidSpheres 50002 was added, and cells were incubated at room
temperature (15–25°C) for 5 min. Cell suspensions were brought up
to a total volume of 2.5 ml by adding the recommended medium, and
were placed into the magnet for 5 min at room temperature
(15–25°C). The EasySep magnet was removed and the desired fraction
was poured off into a 15-ml tube three times following 5 min
separations in the magnet. The isolated cells in the 15-ml tube
were naïve NK cells.
Modified NK cell isolation
To obtain high purity NK cells, the Miltenyi CD3ε
MicroBead kit (130-094-973) was used in combination with a Miltenyi
CD49b (DX5) positive selection kit. Splenocytes were labeled with
CD3ε-biotin and the cells were magnetically labeled with
anti-biotin micro beads. Cell suspensions were loaded onto the MACS
column, which was placed in the magnetic field of a MACS separator.
Magnetically labeled CD3ε+ cells were retained within
the column, while the unlabeled cells ran through. This cell
fraction was depleted of CD3ε+ cells. Subsequently, the
CD49b (DX5) micro beads were used according to the manufacturer's
instructions to isolate NK cells from the cell fraction depleted of
CD3ε+ cells.
Flow cytometry
Anti-mouse antibodies used for flow cytometry were
as follows: PE-conjugated anti-NK1.1 (PK136; catalogue no. 108707;
1:100), allophycocyanin-conjugated anti-NK1.1 (PK136; catalogue no.
108709; 1:100), Fluorescein isothiocyanate-conjugated anti-CD3ε
(145-2C11; catalogue no. 100305; 1:100), PE-conjugated anti-F4/80
(BM8; catalogue no. 123109; 1:100), PE-conjugated anti-CD8a
(53-6.7; catalogue no. 100707; 1:100), peridininchlorophyll protein
complex-CY5.5-conjugated anti-CD4 (GK1.5; catalogue no. 100434;
1:100) (BioLegend, Inc., San Diego, CA, USA). Cell viability was
confirmed by staining the cells with 7-aminoactinomycin D (420404;
1:100; BioLegend, Inc.). NK cell purity was evaluated using flow
cytometry as described previously (12). Briefly, freshly isolated NK cells or
cultured NK cells were collected and washed in PBS twice. Then
cells were resuspended in 100 µl staining buffer (421002;
BioLegend, Inc.) and incubated on ice for 15 min with combination
of PE-conjugated anti-NK1.1 or APC-conjugated anti-NK1.1 and
FITC-conjugated anti-CD3ε antibodies. After extensive washing with
PBS, cells were resuspended in 500 µl PBS and 5 µl 7-AAD was added
into the suspension for 10 min. The stained cells were then
analyzed by FCM. A minimum of 10,000 events was acquired using a
FACS Canto II flow cytometer and analyzed with BD FACS DIVA version
6.0 software (BD Biosciences).
Statistical analysis
Statistical analysis was performed using SPSS 16.0
statistical software package (SPSS, Inc., Chicago, IL, USA). Values
were expressed as the mean ± standard deviation from at least three
independent experiments. Statistical significance of differences
was determined by a one-way analysis of variance analysis using the
Bonferroni t-test. P<0.05 was considered to indicate a
statistically significant difference.
Results
Purity of NK cells purified with
Miltenyi NK Cell Isolation kit II
The proportion of NK cells was first detected in
female C57Bl/6 mice of identical age. For this purpose, FCM
analysis of NK cells, defined as NK1.1+CD3ε−,
was performed in viable splenocytes. Results indicated that in
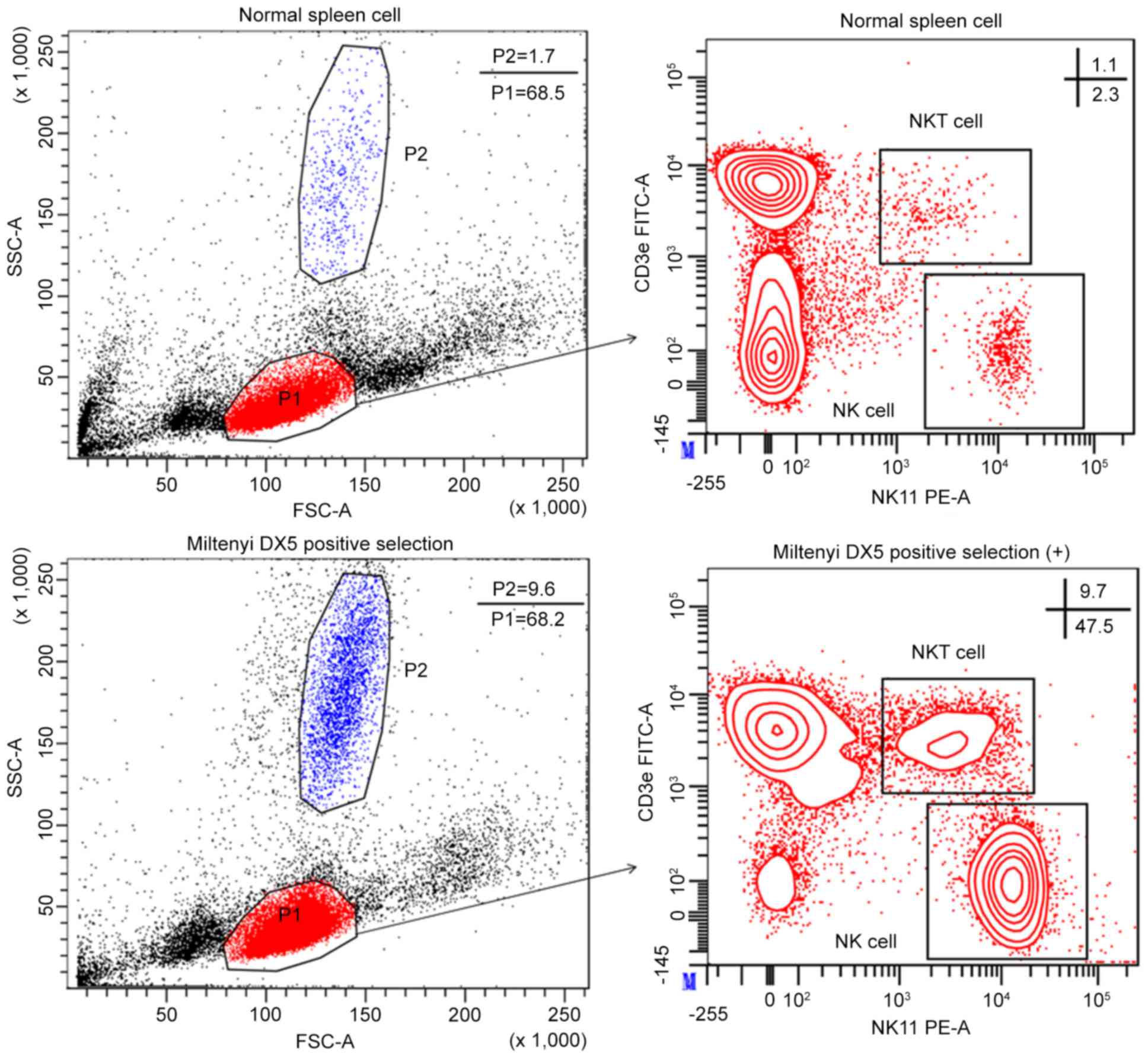
C57Bl/6 mice, the spleen contained 2.63±0.31% NK cells (Fig. 1). To confirm cell purity and the
success of the isolation procedure of the Miltenyi NK Cell
Isolation kit II, isolated cells were phenotyped by FCM for the
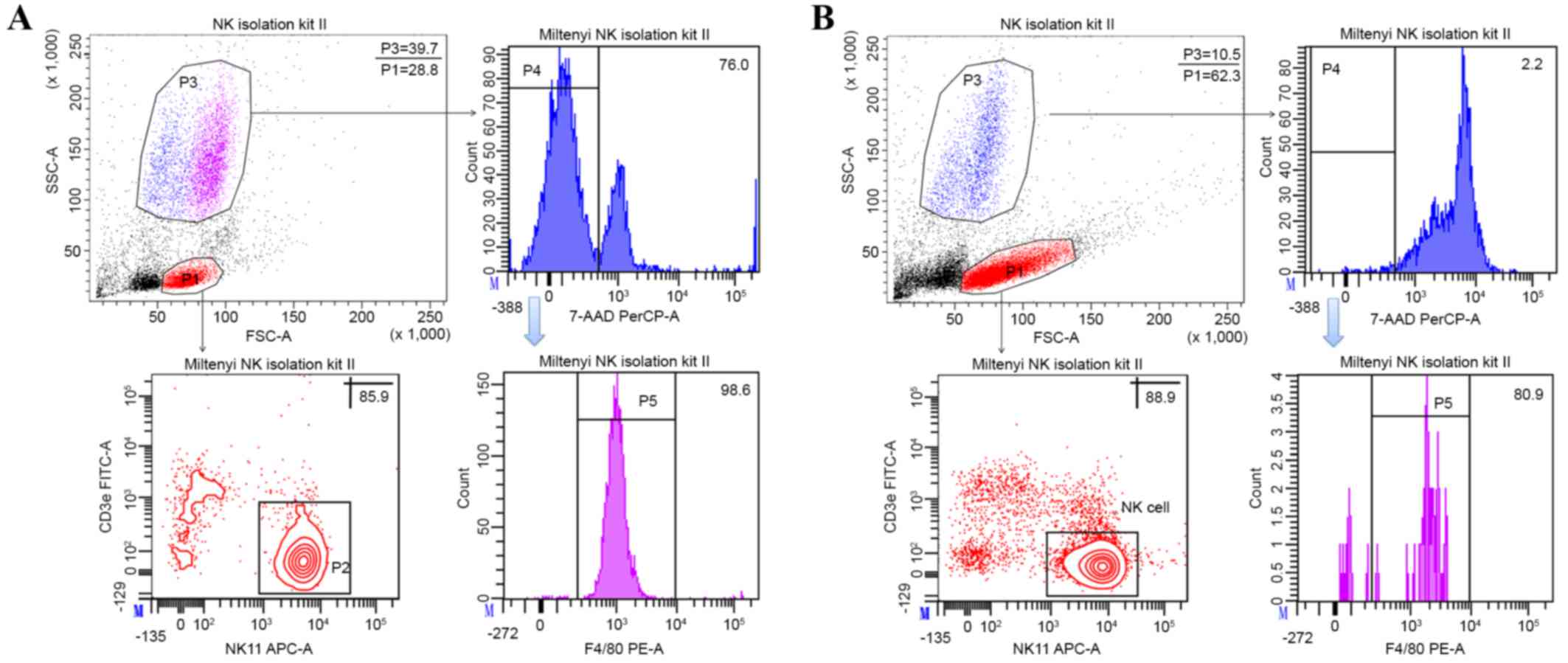
expression of NK1.1 and CD3ε. NK cell purity in the gated
population was 81.47±3.98% (Fig.
2A); however, the percentage of the gated NK cells population
based on forward and side scatter characteristics was 34.37±5.41%
(Fig. 2A). There was a large
population (33.83±5.58%; Fig. 2A) at
the position of granulocytes; however, the percentage was only
1.83±0.15% in the splenocytes (Fig.
1). To identify the prosperity of this population, cells were
stained with F4/80 antibody. The findings revealed that almost all
of the population was F4/80 positive (Fig. 2). Isolated cells were cultured with
500 U/ml IL-2 for 24 h to deplete this population. Notably,
following the subsequent depletion step, almost all cells underwent
cell death and the percentages of gated NK cells and NK cell purity
were increased (Fig. 2B).
Purity obtained with Miltenyi CD49b
(DX5) positive selection kit
Isolated cells from the Miltenyi CD49b (DX5)
positive selection kit were phenotyped by FCM for the expression of
NK1.1 and CD3ε. Results indicated that the NK cell purity from this
kit was ~51.50±4.52% (Fig. 1).
However, this kit presented with certain issues. The percentage of
the granulocyte population (9.07±1.19%) was significantly increased
when compared with that in the normal splenocytes (1.83±0.15%;
P=0.005). Furthermore, the percentage of natural killer T (NKT)
cells, defined as the NK1.1+CD3ε+
(10.8±1.49%) population, was increased in the isolated viable
lymphocyte population when compared with the splenocytes
(1.13±0.25%), which were also contaminated with numerous
CD3ε+ NK1.1− cells (32.97±4.17%; Fig. 1).
Purity of Stemcell CD49b Positive
Selection kit
Since the positively selected cells of the Stemcell
CD49b Positive Selection kit were already PE-labeled, the purity
was assessed directly by FCM. Anticipated results were achieved
using the recommended marker in this kit and the CD49b+
(DX5) cell content of selected cells was 92.90±2.36% (Fig. 3). To confirm NK cell purity, cells
were phenotyped by FCM for the expression of NK1.1 and CD3ε,
following purification. Surprisingly, the NK cell purity was only
64.93±2.62% and the gated cell population was also slightly
contaminated with NKT cells (6.60±1.21%). Furthermore, purified
cells were analyzed using another NK cell marker,
CD49b+CD3ε−. The percentage of
CD49b+CD3ε− cell population was 79.40±2.95%
and the CD49b+ cells included a large population of
CD3ε+ cells (13.87±2.56%; Fig. 3).
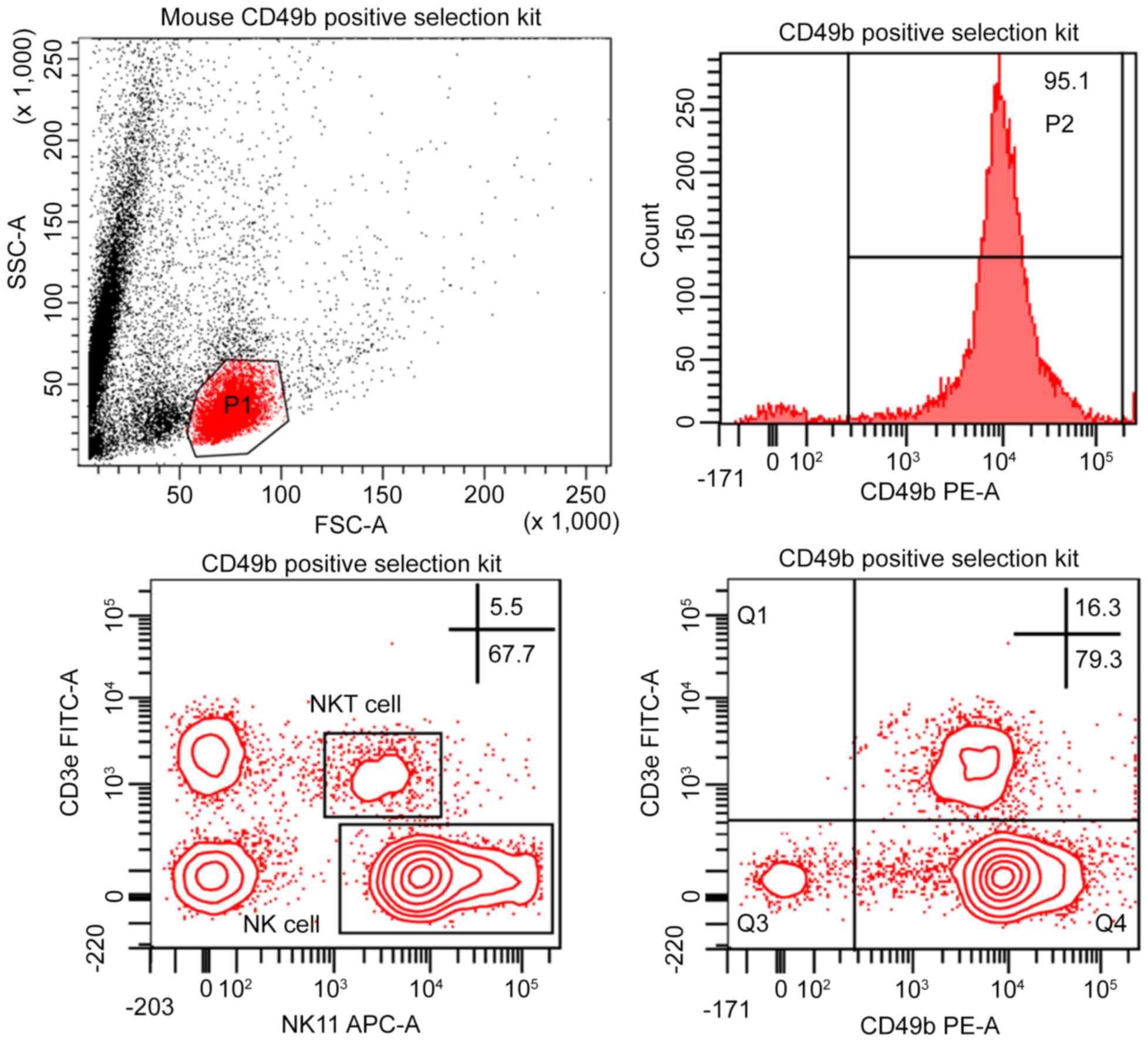 | Figure 3.FCM analysis of purified splenic NK
cells using a Stemcell CD49b positive selection kit. NK cell purity
is shown as a percentage of the total gated viable lymphocyte
population (P1). The histogram represents the percentage of
CD49b+ cells of P1. NK cells were defined as
NK1.1+CD3ε− or
CD49b+CD3ε−. The indicated percentages
represent the purity efficacy represented as the NK cell
population/total viable lymphocyte population (P1) ×100. The
percentage of NKT cell, defined as
NK1.1+CD3ε+, was also indicated. This plot is
one representative experiment obtained from at least three separate
assays performed in parallel. FCM, flow cytometry; NK, natural
killer; CD, cluster of differentiation; FSC, forward scatter; SSC,
side scatter; FITC, fluorescein isothiocyanate; PE, phycoerythrin;
NKT cell, natural killer T cell. |
Purity of Stemcell Mouse NK Cell
Isolation kit
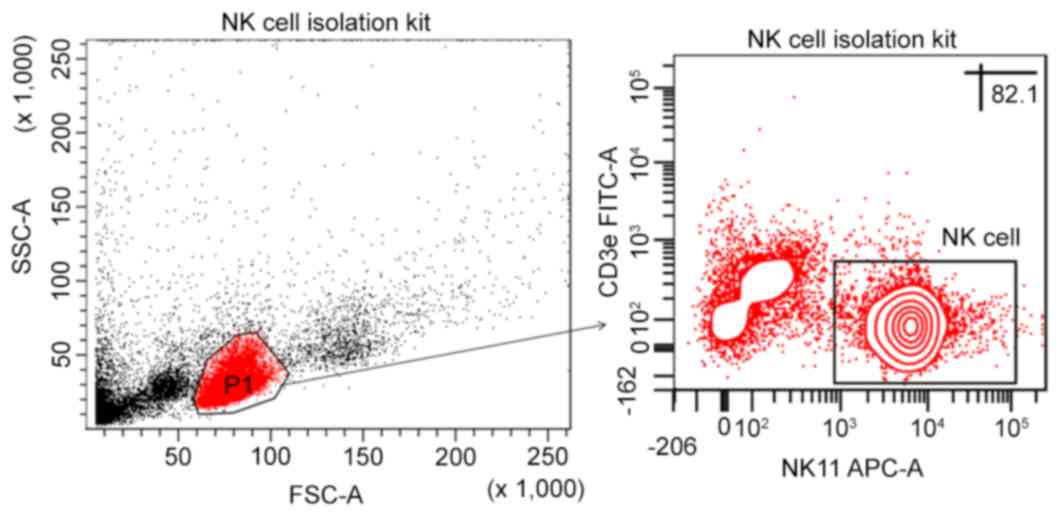
NK cell purity as determined by the Stemcell Mouse
NK Cell Isolation kit was 81.97±3.40% in the viable lymphocyte
population, following purification. No contaminated cells were
identified at the position of granulocytes and no NKT cell
contamination was observed (Fig.
4).
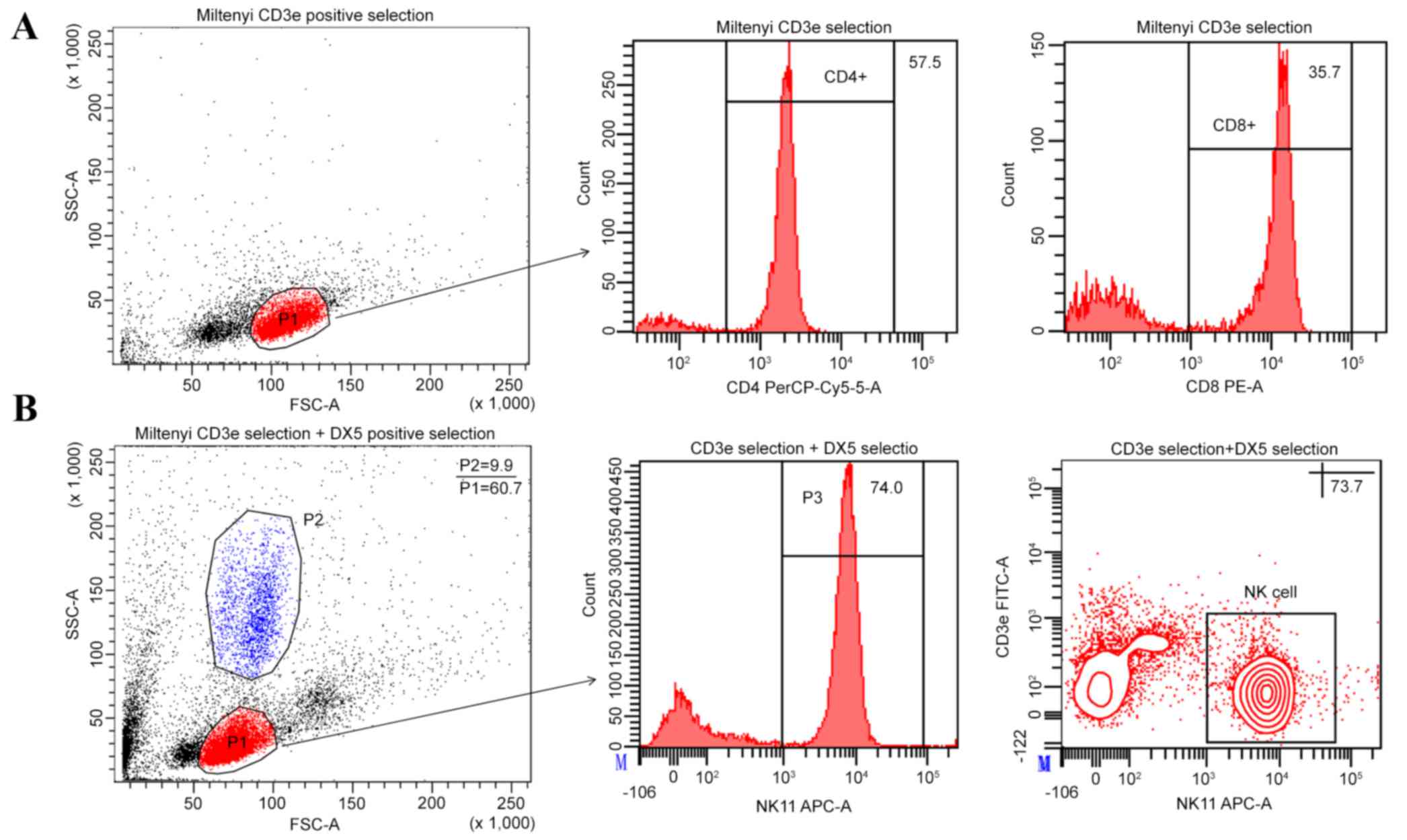
Purity of modified NK cell
isolation
In the present study, the Miltenyi NK cell positive
selection kit (DX5 positive selection kit) was observed to be
contaminated with CD3ε+ cells. In order to deplete this
population of cells, the Miltenyi CD3ε MicroBead kit was used in
combination with the Miltenyi CD49b (DX5) positive selection kit.
The efficiency of the Miltenyi CD3ε MicroBead kit was identified,
and the results indicated a high purity of CD3+ cells
were enriched in the positive portion without granulocyte
contamination. Subsequently, the results of the Miltenyi CD3ε
MicroBead kit were used combination with the Miltenyi CD49b (DX5)
positive selection kit and showed that the NK cell purity was
73.23±2.83% and the NKT cell contamination was depleted. However,
the contamination of granulocytes was not depleted (8.5±1.35%;
Fig. 5).
Purity and viability of NK cells in
presence of different concentrations of IL-2
Freshly isolated NK cells from murine stem cells
were cultured using an NK cell isolation kit at a density of
5×105 cells/ml in 96-well flat plates for 24, 48 or 72 h
in the absence or presence of IL-2 (0, 100, 300 or 500 U/ml). The
purity and viability of NK cells was assessed using FCM following
culturing for 24, 48 or 72 h (Fig.
6A-C) and the statistical analysis of the purity and viability
were performed as presented in Fig. 6E
and F. Detailed results were presented in Table I and Fig.
6. The results revealed that at higher IL-2 concentration, the
NK cell purity and viability are significantly higher (Fig. 6E and F; P<0.05). The majority of
NK cells under went cell death following 24 h in culture in absence
of IL-2.
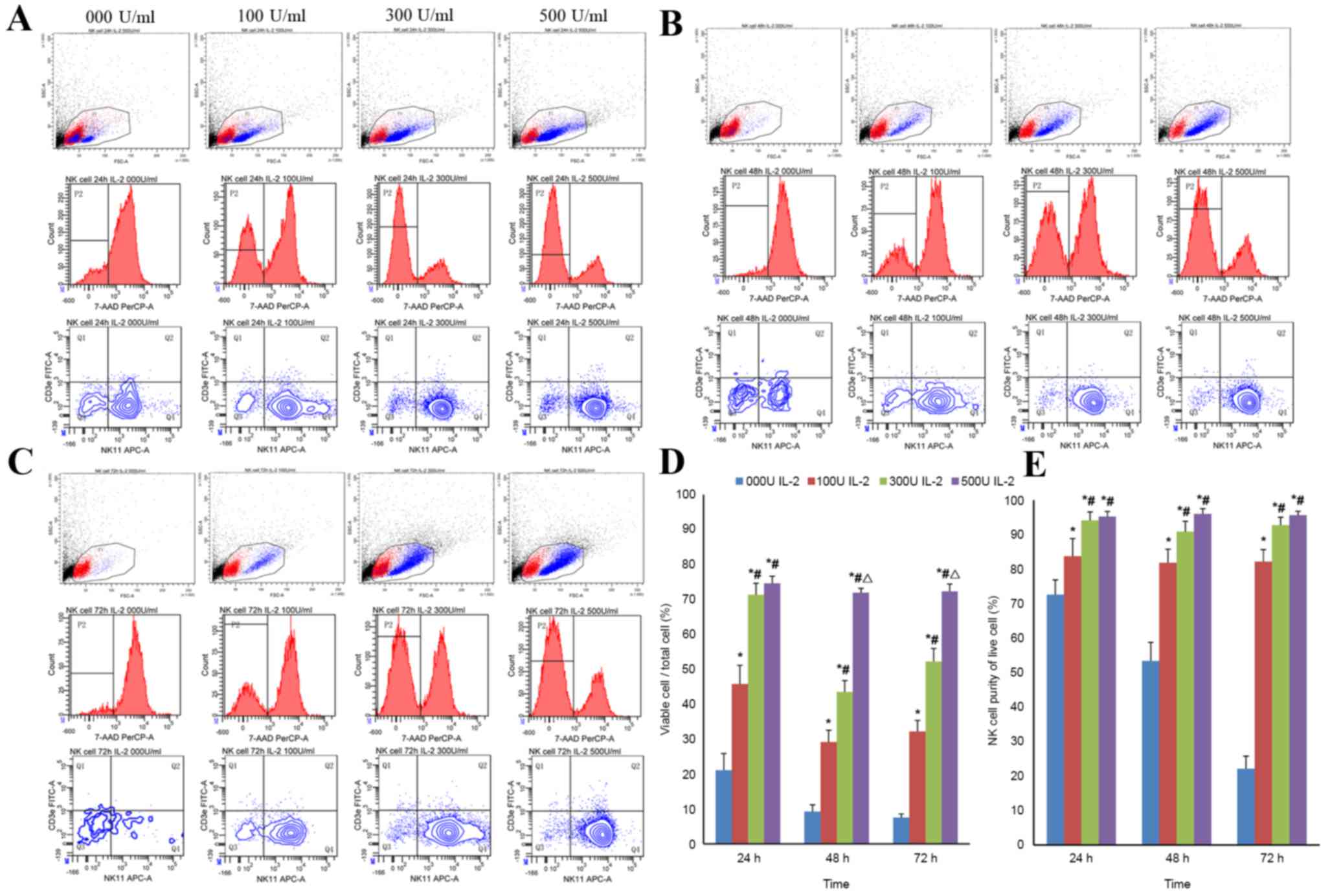 | Figure 6.Analysis of the purity and viability
of NK cells in the presence of different concentrations of IL-2 by
FCM. Purity and viability of NK cell were evaluated using FCM
following culturing for (A) 24, (B) 48 or (C) 72 h. All cells,
except debris, were gated based on FSC and SSC. Subsequently, 7-AAD
was used to identify the percentage of viable cells. NK cell purity
was assessed in viable cells using
NK1.1+CD3ε−. (D) Viability and (E) purity of
NK cells following culturing for 24, 48, and 72 h. Data are
expressed as mean ± standard deviation. *P<0.05 vs. the group
treated with 0 U/ml of IL-2; #P<0.05 vs. the group
treated with 100 U/ml of IL-2; ΔP<0.05 vs. the group
treated with 300 U/ml of IL-2. FCM, flow cytometry; NK, natural
killer; 7-AAD, 7-aminoactinomycin D; IL-2, interleukin-2; FSC,
forward scatter; SSC, side scatter; FITC, fluorescein
isothiocyanate; PE, phycoerythrin. |
 | Table I.Purity and viability of NK cells with
different concentration of IL-2. |
Table I.
Purity and viability of NK cells with
different concentration of IL-2.
|
| NK cells purity of
living cell, % | Viable cell/total
cell, % |
|---|
|
|
|
|
|---|
| IL-2 concentration,
U/ml | 24 h | 48 h | 72 h | 24 h | 48 h | 72 h |
|---|
| 0 | 72.57±4.30 | 53.37±5.44 | 22.13±3.67 | 21.10±4.78 | 9.23±1.93 | 7.50±1.05 |
| 100 |
83.70±5.16a |
81.83±3.95a |
82.17±3.50a |
45.77±5.34a |
29.13±3.39a |
32.17±3.15a |
| 300 |
94.07±2.51a,b |
90.83±3.03a,b |
92.73±2.30a,b |
71.30±3.29a,b |
43.50±3.24a,b |
52.23±3.70a,b |
| 500 |
95.20±1.49a,b |
95.97±1.55a,b |
95.63±1.14a,b |
74.60±2.02a,b |
71.90±1.25a–c |
72.30±2.10a–c |
Comparison of the purity and time
required of different NK cell isolation kits
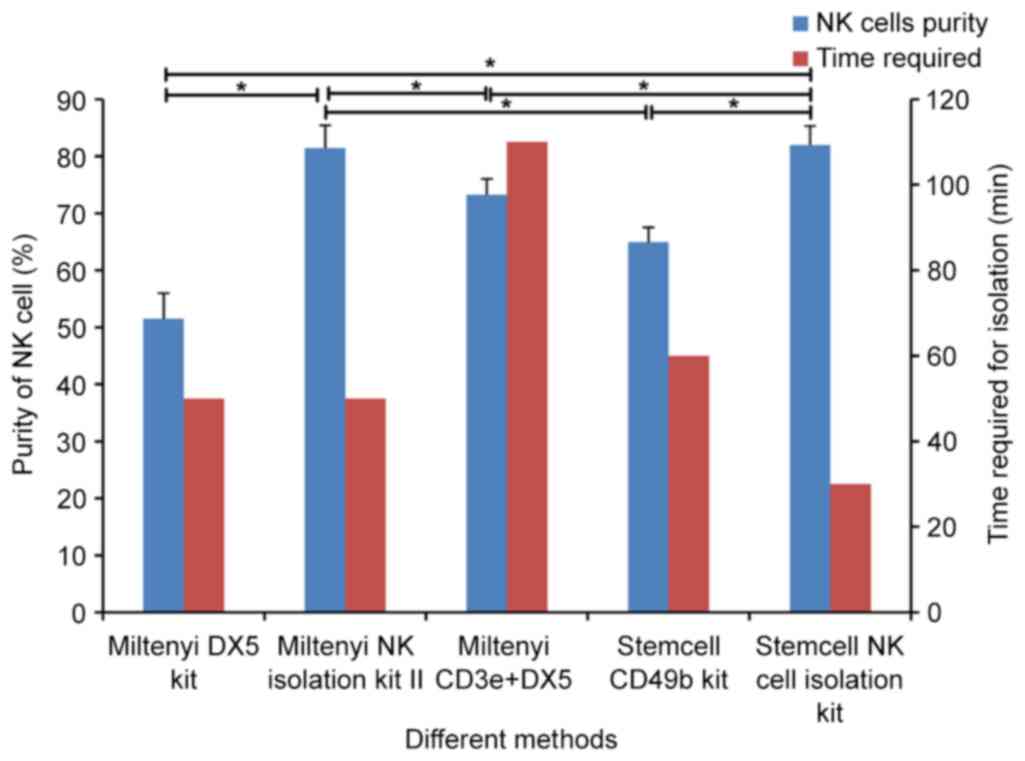
The purity and time required for NK cell isolation
using different kits were compared as presented in Fig. 7. Without consideration of the yield
of purified NK cells, the NK cells purity and the time required of
different kits are as follows: Miltenyi DX5 kit (51.50±4.52%, 50
min), Miltenyi NK Cell Isolation kit II (81.47±3.98%, 50 min),
modified NK cell isolation (73.23±2.83%, 110 min), Stemcell CD49b
Positive Selection kit (64.93±2.62%, 60 min), Stemcell Mouse NK
Cell Isolation kit (81.97±3.40%, 30 min).
Discussion
NK cells are innate lymphocytes that provide host
protection against infectious diseases and cancer. In vitro
studies of NK cells are necessary to obtain fundamental information
on their function and the mechanisms of their interaction with
other cells. Mouse models are considered useful tools in developing
pre-clinical adoptive NK cell transfer immunotherapy against human
tumors (14). A prerequisite for
further detailed functional characterization of NK cells is how to
optimize the purification method. In the present study, the purity
of NK cells was identified to be varied among the different
purification kits used, despite the same method being applied. More
granulocytes were detected in the purified NK cells using the
Miltenyi sorting kit, particularly while using the negative
selection kit. The main drawback of DX5-positive selection using
Stemcell and Miltenyi kits was that a high percentage of
CD3ε+ cells were mixed into the isolated NK cells.
Furthermore, a significant difference in NK cell purity was
observed while the purification was performed using different
surface markers. Therefore, the positive selection kit procedure
was modified and a higher purity and yield of NK cells was
obtained. Moreover, the purity of NK cells was compared with the
viability with or without a range of concentrations of IL-2. These
findings revealed that the higher IL-2 concentrations resulted in a
higher purity of NK cells.
The purity and time required for NK cells isolation
to occur in different kits was compared. Without consideration of
the time required and the yield of purified NK cells, the NK cells
purity in the gated viable mononuclear cell population of negative
selection was higher than that of positive selection. As for the
specific kits, NK cell purity of the Stemcell kit was higher when
compared with the Miltenyi kit, particularly the positive selection
kit. Compared with Stemcell kits, there was a severe issue with
granulocyte contamination of the purified NK cells based on forward
scatter and side scatter properties when using Miltenyi sorting
kits, particularly with the negative selection kit, which affected
the yield of NK cells. Following FCM identification, this
contaminated population was identified as almost entirely F4/80
positive. Thus, it was speculated that this population was likely
macrophages. In order to deplete this population from purified NK
cells, the total isolated cells were cultured for 24 h. However,
this population consisted of non-adherent cells and almost all
cells underwent cell death following 24 h in culture. Thus, there
are no macrophages in the contaminated population, thus they may be
neutrophils, eosinophilic or basophile granulocytes because these
cells are non-adherent and short-lived (15). The exact type of these contaminating
cells requires further explanation. Additionally, the CD3ε-positive
selection kit and DX5-positive selection kit were combined to
achieve a high purity of NK cells. When the CD3ε-positive selection
kit was initially used, granulocyte contamination was not
identified in the positive fraction; however, after using the
DX5-positive kit, the granulocytes contamination was detected
again. These results confirmed the theory that contaminated cells
were reserved due to CD49b expression, although the exact type of
these contaminating cells (CD49b+F4/80+)
requires further explanation. When NK cells were isolated using
Miltenyi kits, the normal prepared spleen cells were not
advantageous due to the enriched granulocytes observed following
purification. We recommend that the spleen cells are pretreated
first to deplete the granulocytes, which may be achieved by
gradient centrifugation. Another disadvantage of NK cell isolation
using DX5-positive selection according to both Stemcell and
Miltenyi kits was that the isolates contained a high percentage of
contaminating CD3ε+ cells, including NKT cells
(NK1.1+CD3ε+) and some T cells
(NK1.1−CD3ε+). The predominant reason for
this problem is in the expression of CD49b. NKT cells and certain T
cells may express CD49b (16,17).
Thus, using CD49b as sorting target may cause these cells to be
reserved in the mixture. This problem requires a solution by
alternative means, such as combining this kit with the CD3-positive
selection kit, which can remove all CD3+ cells,
including NKT cells and therefore improve the purity of NK cells.
Furthermore, it was identified with DX5-positive selection that the
NK cell purity of isolates was variable based on the different
markers used. The Stemcell CD49b Positive Selection kit is a
typical representation. Following isolation, the purity of
CD49b-positive cells was >95% in the isolated cells, whereas the
purity of CD49b+CD3ε− cells was ~80% and the
purity of NK1.1+CD3ε− cells was ~65%. The
different purity of NK cells based on different markers was also
associated with the expression of CD49b (16,17).
Therefore, when the purity of isolated NK cells was identified
using the DX5-positive selection kit, the classic surface markers
were revealed to be the preferential choice over other markers.
Consequently, isolating NK cells using only the CD49b-positive
strategy is not recommended; however, this kit provides improved
results when combined with the CD3-positive selection kit.
Although combining the use of CD3ε positive
selection with DX5 positive selection may improve the purity of NK
cell, the time required for isolation was extended. Additionally,
the NK cell is a functionally and phenotypically heterogeneous
population (18) and perhaps only a
specific subpopulation is isolated by positive selection. It has
also been described that in vitro cultured NK cells are no
longer recognized by DX5 antibodies (19). Moreover, DX5 is not expressed on
immature NK cells and tissue-resident NK cells in the liver
(20,21). Therefore, the isolation of NK cells
via CD49b as a positive selection marker should be undertaken
depending on the experiment requirements. The negative selection of
NK cells can capture naïve cells. It is critical that functional
studies are performed in the future. There are several advantages
of negative selection with one of these being that the NK cells are
not coated with antibody and, therefore, are not at risk of
functional perturbation by antibody cross-linking. Additionally,
negative selection provides a way to isolate diverse subpopulations
of NK cells without selectively purifying a specific subpopulation
(13). In our comparative
experiments, high purity and yield of NK cells were obtained in a
shorter time frame using the negative selection method,
particularly with the Stemcell Mouse NK Cell Isolation kit. The
Miltenyi NK Cell Isolation kit II was also indicated to be a
suitable choice of kit if the granulocytes of the spleen are
depleted prior to sorting.
NK cells are short-lived and IL-2 dependent in
vitro (22). IL-2 exposure;
however, will result in differences owing to activation, such as NK
cell killing of a broader panel of targets, which may be
inappropriate for certain experimental settings. Therefore, a
suitable concentration of IL-2 is critical for specific studies. In
the present report, the NK cell purity and viability was compared
with or without different concentrations of IL-2 (0, 100, 300 or
500 U/ml), following culturing for 24, 48 or 72 h using the
Stemcell negative selection kit. Without IL-2, the purity of NK
cells significantly decreased over time; however, with the addition
of 100 U/ml IL-2, the NK cell purity may be maintained at >80%
after 72 h. The purity increased with increased concentrations of
IL-2 at the same time point, and with the same concentration of
IL-2 the purity was approximately the same at different time
points. Consequently, the purity of NK cells was able to meet the
requirement of the experiment no matter what concentration of IL-2
was used. However, the IL-2 concentration was critical for the
viability of NK cells at different time points. Only 20% of NK
cells were viable following 24 h without IL-2, and after 72 h ~10%
were viable. Therefore, if studies wish to use NK cells in
vitro for >24 h, IL-2 is required. The viability of NK cells
was increased with high concentrations of IL-2 at the same time
point, particularly at 48 and 72 h. At a concentration of 100 or
300 U/ml of IL-2, the viability of NK cell was increased at 72 h
when compared with 48 h exposure. Notably, the viability of NK
cells was similar at different time points with 500 U/ml IL-2,
which may be explained by NK cell proliferation. High
concentrations (500 U/ml) of IL-2 are able to activate more NK
cells to proliferate, which results in a balance between death and
proliferation of NK cells. Thus, at different time points, the
viability of NK cells is similar. However, low concentrations (100
or 300 U/ml) of IL-2 are not able to activate more NK cells to
proliferate; therefore, if a study requires the use of NK cells
in vitro for 24 h, the concentration of 300 U/ml IL-2 is
recommended, based on the present results. If a study requires the
use of NK cells in vitro for 48 or 72 h, a concentration of
500 U/ml IL-2 is recommended.
In conclusion, it is important to consider that
different NK cell isolation kits affect the purity and yield of
isolated NK cells. When isolating NK cells using Miltenyi kits, the
spleen cells should be pretreated to deplete the granulocytes prior
to sorting. Combined use of the CD3ε selection kit with the DX5
selection kit is able to produce higher purity and yield of NK
cells. We favor the use of the negative selection kit from
Stemcell, which can produce naïve NK cells that may be further
analyzed. The present study revealed that IL-2 is essential for the
viability of NK cells in vitro. The present study concludes
that a concentration of 300 U/ml of IL-2 is recommended for the
culture NK cells in vitro for 24 h to obtain optimal
resolution and 500 U/ml of IL-2 is suitable for NK cells in
vitro for 48 or 72 h.
Acknowledgments
The present study was supported by Chinese National
Natural Science Foundation (grant no. 81373162) and Nature Science
of Foundation of Liaoning Province, China (grant no.
2012225016).
Glossary
Abbreviations
Abbreviations:
|
FCM
|
flow cytometry
|
|
MACS
|
magnetic activated cell sorting
|
|
NK cells
|
natural killer cells
|
|
NKT cells
|
natural killer T cells
|
|
IL-2
|
interleukin 2
|
References
|
1
|
Jost S and Altfeld M: Control of human
viral infections by natural killer cells. Annu Rev Immunol.
31:163–194. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Bär E, Whitney PG, Moor K, Sousa Reise C
and LeibundGut-Landmann S: IL-17 regulates systemic fungal immunity
by controlling the functional competence of NK cells. Immunity.
40:117–127. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Zitvogel L, Galluzzi L, Smyth MJ and
Kroemer G: Mechanism of action of conventional and targeted
anticancer therapies: Reinstating immunosurveillance. Immunity.
39:74–88. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Tian Z, Gershwin ME and Zhang C:
Regulatory NK cells in autoimmune disease. J Autoimmun. 39:206–215.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kim HS, Das A, Gross CC, Bryceson YT and
Long EO: Synergistic signals for natural cytotoxicity are required
to overcome inhibition by c-Cbl ubiquitin ligase. Immunity.
32:175–186. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Shi FD, Ljunggren HG, La Cava A and Van
Kaer L: Organ-specific features of natural killer cells. Nat Rev
Immunol. 11:658–671. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
7
|
Carlyle JR, Mesci A, Ljutic B, Belanger S,
Tai LH, Rousselle E, Troke AD, Proteau MF and Makrigiannis AP:
Molecular and genetic basis for strain-dependent NK1.1
alloreactivity of mouse NK cells. J Immunol. 176:7511–7524. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Charbonnier LM, van Duivenvoorde LM,
Apparailly F, Cantos C, Han WG, Noël D, Duperray C, Huizinga TW,
Toes RE, Jorgensen C and Louis-Plence P: Immature dendritic cells
suppress collagen-induced arthritis by in vivo expansion of CD49b+
regulatory T cells. J Immunol. 177:3806–3813. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Hackett J Jr, Tutt M, Lipscomb M, Bennett
M, Koo G and Kumar V: Origin and differentiation of natural killer
cells. II. Functional and morphologic studies of purified NK-1.1+
cells. J Immunol. 136:3124–3131. 1986.PubMed/NCBI
|
|
10
|
Ravnik SE, Gage S and Pollack SB:
Self-generating density gradients of Percoll provide a simple and
rapid method that consistently enriches natural killer cells. J
Immunol Methods. 110:161–168. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Patel MR and Linna TJ: Enrichment of mouse
splenic natural killer cells using discontinuous
polyvinylpyrrolidone silica (Percoll) gradients. Immunology.
53:721–729. 1984.PubMed/NCBI
|
|
12
|
Meinhardt K, Kroeger I, Abendroth A,
Müller S, Mackensen A and Ullrich E: Influence of NK cell magnetic
bead isolation methods on phenotype and function of murine NK
cells. J Immunol Methods. 378:1–10. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Pak-Wittel MA, Piersma SJ, Plougastel BF,
Poursine-Laurent J and Yokoyama WM: Isolation of murine natural
killer cells. Curr Protoc Immunol. 105:3.22.1–9. 2014. View Article : Google Scholar
|
|
14
|
Davis ZB, Felices M, Verneris MR and
Miller JS: Natural killer cell adoptive transfer therapy:
Exploiting the first line of defense against cancer. Cancer J.
21:486–491. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Nowarski R, Gagliani N, Huber S and
Flavell RA: Innate immune cells in inflammation and cancer. Cancer
Immunol Res. 1:77–84. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wang C, Liu X, Li Z, Chai Y, Jiang Y, Wang
Q, Ji Y, Zhu Z, Wan Y, Yuan Z, et al: CD8 (+)NKT-like cells
regulate the immune response by killing antigen-bearing DCs. Sci
Rep. 5:141242015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kaufman G, d'Ovidio R, Kaldawy A, Assy B,
Ullmann Y, Etzioni A, Paus R and Gilhar A: An unexpected twist in
alopecia areata pathogenesis: Are NK cells protective and CD49b+ T
cells pathogenic? Exp Dermatol. 19:e347–e349. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Meinhardt K, Kroeger I, Bauer R, Ganss F,
Ovsiy I, Rothamer J, Büttner M, Atreya I, Waldner M, Bittrich M, et
al: Identification and characterization of the specific murine NK
cell subset supporting graft-versus-leukemia- and reducing
graft-versus-host-effects. Oncoimmunology. 4:e9814832015.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Arase H, Saito T, Phillips JH and Lanier
LL: Cutting edge: The mouse NK cell-associated antigen recognized
by DX5 monoclonal antibody is CD49b (alpha 2 integrin, very late
antigen-2). J Immunol. 167:1141–1144. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kim S, Iizuka K, Kang HS, Dokun A, French
AR, Greco S and Yokoyama WM: In vivo developmental stages in murine
natural killer cell maturation. Nature immunology. 3:523–528. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Peng H, Jiang X, Chen Y, Sojka DK, Wei H,
Gao X, Sun R, Yokoyama WM and Tian Z: Liver-resident NK cells
confer adaptive immunity in skin-contact inflammation. J Clin
Invest. 123:1444–1456. 2013. View
Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kuribayashi K, Gillis S, Kern DE and
Henney CS: Murine NK cell cultures: Effects of interleukin-2 and
interferon on cell growth and cytotoxic reactivity. J Immunol.
126:2321–2327. 1981.PubMed/NCBI
|