Introduction
Since the use of biodegradable implants to heal
fractures by Rokkanen et al in 1985, the method of using
biodegradable materials has been widely applied in orthopedic
surgeries (1–3). Biodegradable materials exhibit various
advantages in the treatment of fractures compared with the use of
traditional metal implants, including the elimination of implant
removal, reduction of the ‘stress shielding’ effect, improvement of
biocompatibility, reduction of radiological artifacts and
utilization of magnetic resonance imaging assessment following
surgery (4,5). Poly-D-L lactide (PDLLA) is a material
with an intermediate degradation time (PDLLA begins to degrade at
~12 weeks) and may be completely replaced by bone tissue following
surgery, thus, it is considered to be one of the most effective
biodegradable materials for the treatment of fractures (6–8).
However, with the exception of clinical outcomes, the differences
in the biological processes following the use of PDLLA and metal
fixation to treat a fracture remain unknown.
The fracture healing process has been widely
accepted to comprise a series of overlapping phases; these include
inflammation, repair and remodeling events (9). This multistage repair process involves
a variety of complex and well-orchestrated cellular and molecular
processes, and should be considered as a specialized post-natal
recapitulation of embryological skeletal development (9,10).
Runt-related transcription factor 2 (Runx2) is one member of the
Runx family that belongs to a small transcription factor family
sharing a common runt domain (11).
Previous studies indicated that Runx2 was a crucial transcriptional
regulator for skeletal development and served essential roles in
biological events including osteoblast differentiation,
intramembranous, endochondral ossification and in the process of
fracture healing, vascular invasion (12–14).
However, whether the expression of Runx2 during fracture healing
may be altered by the use of PDLLA compared to that of a
traditional metal material for fracture treatment remains
unknown.
A previous study demonstrated that the canonical Wnt
signaling served essentials roles in regulating bone formation
during the fracture repair process (15). In the canonical Wnt signaling
pathway, the Wnts proteins, a family of secreted glycoproteins,
bind to the membrane receptor Frizzled (Fz) leading to downstream
sub-cellular events. The disheveled protein, along with other
co-receptors, is activated to inhibit the activity of glycogen
synthase kinase-3β (GSK-3β). β-catenin is able to avoid the
degradation of the protein ubiquitinylation system and translocate
into the nucleus in order to associate with transcription factors,
including T cell factor-1 (TCF-1), in order to regulate the
transcriptional activities of relevant genes (16). A number of studies have indicated
that the expression of certain Wnt proteins, including Wnt4, 5b,
10b, 11, and 13, and Fz receptors such as Fz1, 2, 4, and 5, are
increased during the fracture healing process (17,18). The
expression of Runx2 may also be regulated by the activity of
canonical Wnt signaling through its binding with β-catenin/TCF-1 in
certain promoter regions of the runx2 gene (19). However, whether and to what extent
the activity of the canonical Wnt signaling in the bone repair
process may be changed by PDLLA, compared to that of traditional
metal material for fracture treatment, has not yet been
determined.
Overall, the present study established an open
osteotomy model of Sprague-Dawley rats in order to determine
whether and to what extent the expression of Runx2 and the activity
of canonical Wnt signaling during bone repair process may be
differentially regulated. The use of traditional metal material
(Kirschner wire) and PDLLA intramedullary rod under internal
fixation for fracture treatment was assessed. The results of the
present study indicate that compared to the traditional metal
material, the curative efficacy of PDLLA internal fixation for
fracture treatment is improved, mediated by the increased Runx2
expression associated with the enhanced activity of canonical Wnt
signaling.
Materials and methods
Experimental animals
A total of 36 male Sprague-Dawley rats (six months
old) weighing 280–300 g were supplied by the Laboratory Animal
Research Center of Soochow University (Suzhou, China). They were
maintained under standard conditions for at least one week, and
individually housed in plastic cages in an animal room in a
temperature-controlled environment of 22–25°C and relative humidity
of 40–70% with a 12-h light/dark cycle and ad libitum access
to commercial pellets and water. Animals were randomly assigned
into control and PDLLA groups (n=18 per group). Following the
transverse osteotomy at the mid-shaft of right femur, the traumatic
fracture was stabilized intramedullary by the Kirschner wire and
PDLLA rod in control and PDLLA groups, respectively. All procedures
were approved by the Institutional Animal Care Service (Second
Affiliated Hospital of Soochow University).
Surgical procedure of femoral open
osteotomy model
The osteotomy was performed using a circular saw
with a diameter of 1.6 cm and a thickness of 0.1 mm at mid-shaft of
the right femur, under the intraperitoneal injection of 50 mg/kg
sodium pentobarbital (Sinopharm Chemical Reagent, Co., Ltd.,
Shanghai, China). The fracture was stabilized by intramedullary
insertion of a sterilized 1 mm diameter Kirschner wire (Nantong
Healthcare Medical Instrument, Co., Ltd., Nantong, China) in the
control group and a sterilized 1 mm diameter PDLLA rod (Nantong
Healthcare Medical Instrument, Co., Ltd.) in the PDLLA group. At
weeks 2, 4 and 6 post-surgery, 50 mg/kg sodium pentobarbital was
used to anaesthetize 6 rats from each group prior to sacrifice via
cervical dislocation.
Alkaline phosphatase and
osteoprotegerin serum levels
Blood samples (0.5 ml each) were obtained using an
intravenous tube implanted in the femoral vein following 2, 4 and 6
weeks and stored at −20°C until analysis. The concentrations of
alkaline phosphatase (ALP) and osteoprotegrin (OPG) were detected
in the serum using commercially available ELISA kits (C506082-0096,
and C506509-0048; Sangon Biotech, Co., Ltd., Shanghai, China).
MicroCT scanning and assessment of
bone quality parameters
At weeks 2, 4 and 6 post-surgery, six rats randomly
selected from each group were anesthetized with intraperitoneal
injections of sodium pentobarbital (Sinopharm Chemical Reagent,
Co., Ltd., 50 mg/kg) prior to micro-CT scanning. The femora were
scanned by a desktop micro-CT system (IVIS Quantum FX; Cold Spring
Biotech Corporation, Shanghai, China) with the resolution of 47 µm
and scanning time of 15 min each. Values of bone volume fraction
(BV/TV), bone surface density (BS/BV), trabecular thickness
(TB.TH), trabecular number (TB.N), and trabecular separation
(TB.SP) were determined in each group under the certain regions of
interest. The scan area was centered through the fracture line.
Western blot analysis
Protein samples were routinely extracted from callus
tissue using a Total Protein Extraction kit according to the
manufacturer's protocol (310003; BestBio Biotechnology Co., Ltd.,
Shanghai, China) 2, 4 and 6 weeks following surgery, as described
previously (20). Samples of protein
(80 µg) were loaded in each lane for electrophoresis in a 4%
SDS-PAGE gel (0.1% SDS and 10% polyacrylamide) and transferred to
immobilon P polyvinyldifluoride membranes (EMD Millipore,
Billerica, MA, USA). Each membrane was blocked with Tris-buffered
PBS containing 5% bovine serum albumin (Chinese Academy of
Sciences, Shanghai, China) plus 0.1% (v/v) Tween (TBS-T) on a
gentle shaker for 1 h at room temperature, followed by incubation
at 4°C overnight with rabbit polyclonal antibodies (Santa Cruz
Biotechnology, Inc. Santa Cruz, CA, USA) against Runx2 (sc-10758,
1:1,000), hypoxia inducible factor 2A (HIF2A; sc-28706, 1:1,000),
Wnt4 (sc-13962, 1:1,000), Wnt10b (sc-25524, 1:1,000), GSK-3β
(sc-9166, 1:1,000), β-catenin (sc-7199, 1:1,000) and TCF-1
(sc-13025, 1:1,000). Membranes were then washed three times for 10
min in TBS-T solution and incubated at room temperature with the
secondary horseradish peroxidase-conjugated goat anti-rabbit
antibody (1:5,000; A0208; Beyotime Institute of Biotechnology,
Haimen, China) for 2 h. Proteins were then visualized using
enhanced chemiluminescence detection reagents (ECL reagent RPN2232,
GE Healthcare Life Sciences, Chalfront, UK) and exposed to X-ray
film according to the manufacture's protocol. The β-actin protein
(primary antibody from EMD Millipore, working concentration:
1:1,000) was blotted in the same membrane as an internal control to
normalize the relative density. Results were quantified and
analyzed using a Kodak electrophoresis documentation and analysis
system, and Kodak ID image analysis software (Kodak, Rochester, NY,
USA). Three replicates were performed per tissue sample.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
At weeks 2, 4 and 6 post-surgery, total tissue RNA
was routinely isolated from the callus tissue using Eastep Super
Total RNA Extraction kit (Promega Corporation, Madison, WI, USA)
according to the manufacturer's protocol. Reverse transcription and
first strand cDNA synthesis was performed using M-MLV Reverse
Transcriptase according to the manufacture's protocol (Invitrogen;
Thermo Fisher Scientific Inc., Waltham, MA, USA). RT-qPCR analysis
was completed for Runx2, HIF2A, Wnt4, Wnt10b, GSK-3β, β-catenin and
TCF-1 gene expression in callus tissue. The GAPDH gene was used as
an internal control and serial dilutions of the positive control
(the synthesized tissue cDNA from rat bone; RD-107, Zyagen, San
Diego, CA, USA) were performed on each plate to create a standard
curve. The primer sequences (Sangon Biotech, Co., Ltd.) are as
follows: For rat Runx2 are forward, 5′-CACAAGTGCGGTGCAAACTT-3′ and
reverse, 5′-AATGACTCGGTTGGTCTCGG-3′; for rat HIF2A are forward
5′-AACCTTAAGTCGGCCACCTG-3′ and reverse, 5′-TTGCTGTCCAAGGGGATGTC-3′;
for rat Wnt4 are forward, 5′-TACGGATGAGGACCTGGTGT-3′ and reverse,
5′-GACGTCTTGTTGCAAGTGCG-3′; for rat Wnt10b are forward,
5′-CCGTGAGTTAGGTCGAGCAG-3′ and reverse, 5′-GTGGGGAAACTGTGTGGAGT-3′;
for rat GSK-3β are forward, 5′-CGAACTCCACCAGAGGCAAT-3′ and reverse,
5′-GAGTTGGAGGCTGATGCAGA-3′; for rat β-catenin are forward,
5′-CTGCTGATCTCGGACTGGAC-3′ and reverse, 5′-GGCAGCCCATCAACTGGATA-3′;
for rat TCF-1 are forward, 5′-CAGAATCCACAGATACAGCA-3′ and reverse,
5′-CAGCCTTTGAAATCTTCATC-3′; for rat GAPDH are forward,
5′-GGTGGTCTCCACGGACTTTA-3′ and reverse, 5′CAAGGAGGGGCCTTTATTTC3′.
RT-qPCR (iCycler iQ™ Real-time PCR detection system; Bio-Rad
Laboratories, Inc., Hercules, CA, USA) was performed using 25 µl of
each sample, in 96-well plates (Takara Bio, Inc., Otsu, Japan)
under the following protocol: 42°C for 30 min, 95°C for 15 min
followed by 40 cycles of 95°C for 20 sec, 56°C for 1 min and 72°C
for 20 sec. The amount of target gene was normalized to the GAPDH
reference to obtain the relative threshold cycle (ΔΔCq), then
2−ΔΔCq was calculated to determine the relative
abundance for target gene expression between control and PDLLA
groups (21).
Statistical analysis
All data were expressed as mean ± standard
deviation. With regard to the different mRNA level of PDLLA and
control groups, statistical significance was accepted (P<0.05)
when the ratio of 2−ΔΔCq exceeded 1.7. The two-way
analysis of variance and independent t-test, where appropriate, was
used to evaluate the differences between PDLLA and control groups.
A statistically significant difference was accepted if P<0.05.
SPSS statistical software (version 18; SPSS, Inc., Chicago, IL,
USA) was used for all statistical analyses.
Results
MicroCT analysis of fracture
calluses
MicroCT analysis indicated that, compared with the
control, the values of trabeculae quality of the traumatic femur
including BV/TV, BS/TV, TB.TH, TB.N and TB.SP were significantly
improved in the PDLLA group, 4 and 6 weeks following the
intramedullary fixation for femoral osteotomy (P<0.05). However,
at week 2 there was no significant difference in the values of
trabeculae growth between the PDLLA group compared with the control
(P>0.05; Table I).
 | Table I.Comparison of microCT analysis of
fracture calluses between PDLLA group and control group across
time-points. |
Table I.
Comparison of microCT analysis of
fracture calluses between PDLLA group and control group across
time-points.
| microCT
parameters | Weeks after
fixation | Con | PDLLA |
|---|
| BV/TV, % | 2 | 2.469±0.14 | 2.435±0.11 |
|
| 4 | 3.188±0.16 |
3.765±0.13a |
|
| 6 | 5.132±0.18 |
6.123±0.13a |
| BS/BV, 1/m | 2 | 1.887±0.14 | 1.869±0.12 |
|
| 4 | 2.025±0.11 |
2.183±0.12a |
|
| 6 | 3.102±0.12 |
3.244±0.16a |
| BV/TV, % | 2 | 0.0740±0.0011 | 0.0738±0.0014 |
|
| 4 | 0.0751±0.0013 |
0.0768±0.0011a |
|
| 6 | 0.0831±0.0012 |
0.0857±0.0015a |
| BS/BV, l/m | 2 | 0.334±0.031 | 0.333±0.043 |
|
| 4 | 0.491±0.034 |
0.522±0.062a |
|
| 6 | 0.654±0.051 |
0.709±0.061a |
| BV/TV, % | 2 | 1.106±0.0051 | 1.108±0.0044 |
|
| 4 | 1.083±0.0072 |
1.073±0.0035a |
|
| 6 | 1.035±0.010 |
1.018±0.012a |
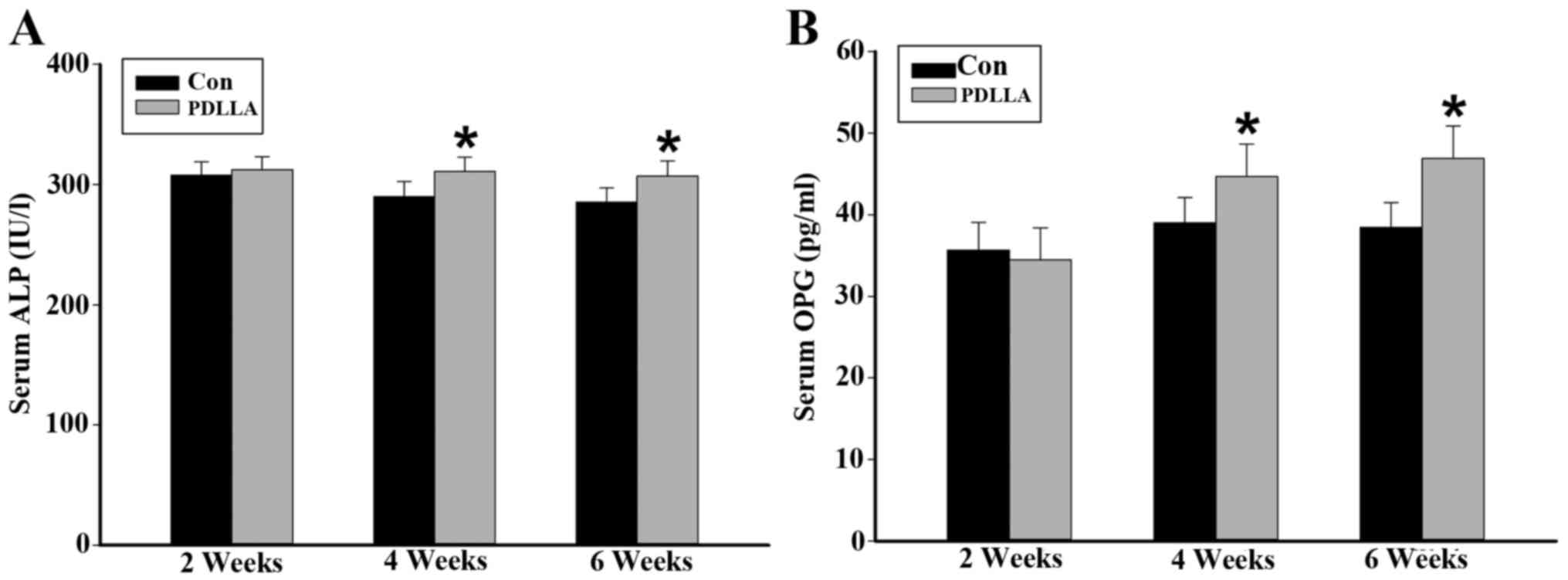
Serum values of ALP and OPG
In order to further evaluate the differences in
osteogenesis between the PDLLA and control group, the circulatory
serum levels of ALP and OPG were detected as they are widely
considered to be markers of bone formation. The ELISA assay
indicated that weeks 4 and 6 following the intramedullary fixation
for femoral osteotomy, the serum levels of ALP and OPG were
significantly increased in the PDLLA group compared with that of
the control (P<0.05). However, results from week 2 demonstrate
that the serum levels of ALP and OPG are not significantly
different in the PDLLA group compared to that of control
(P>0.05; Fig. 1).
Protein expression of Runx2, HIF2A,
Wnts, GSK-3β, β-catenin and TCF-1 in callus tissue
To compare the differences of cellular and molecular
events involved in the process of fracture healing between the
PDLLA and control groups, runx2, canonical Wnt signaling and
associated regulators were dynamically examined. Western blot
analysis demonstrated that 4 and 6 weeks following the
intramedullary fixation for femoral osteotomy, the protein
expression levels of Runx2, HIF2A, Wnts and β-catenin in callus
tissue were significantly increased in the PDLLA group compared
with the control (P<0.05; Fig.
2A-E). Furthermore, the level of GSK-3β protein expression in
callus tissue, at week 4 and 6, was significantly decreased in the
PDLLA group compared to that of control (P<0.05; Fig. 2F). However, at week 2 following the
intramedullary fixation, the level of protein expression for Runx2,
HIF2A, Wnts, β-catenin and GSK-3β exhibited no significant
differences in the PDLLA group compared with that of control
(P>0.05; Fig. 2A-F). With respect
to TCF-1 at weeks 2, 4 and 6 following the intramedullary fixation
for femoral osteotomy, the protein expression levels exhibited no
significant differences in the PDLLA group compared with that of
control (P>0.05; Fig. 2G).
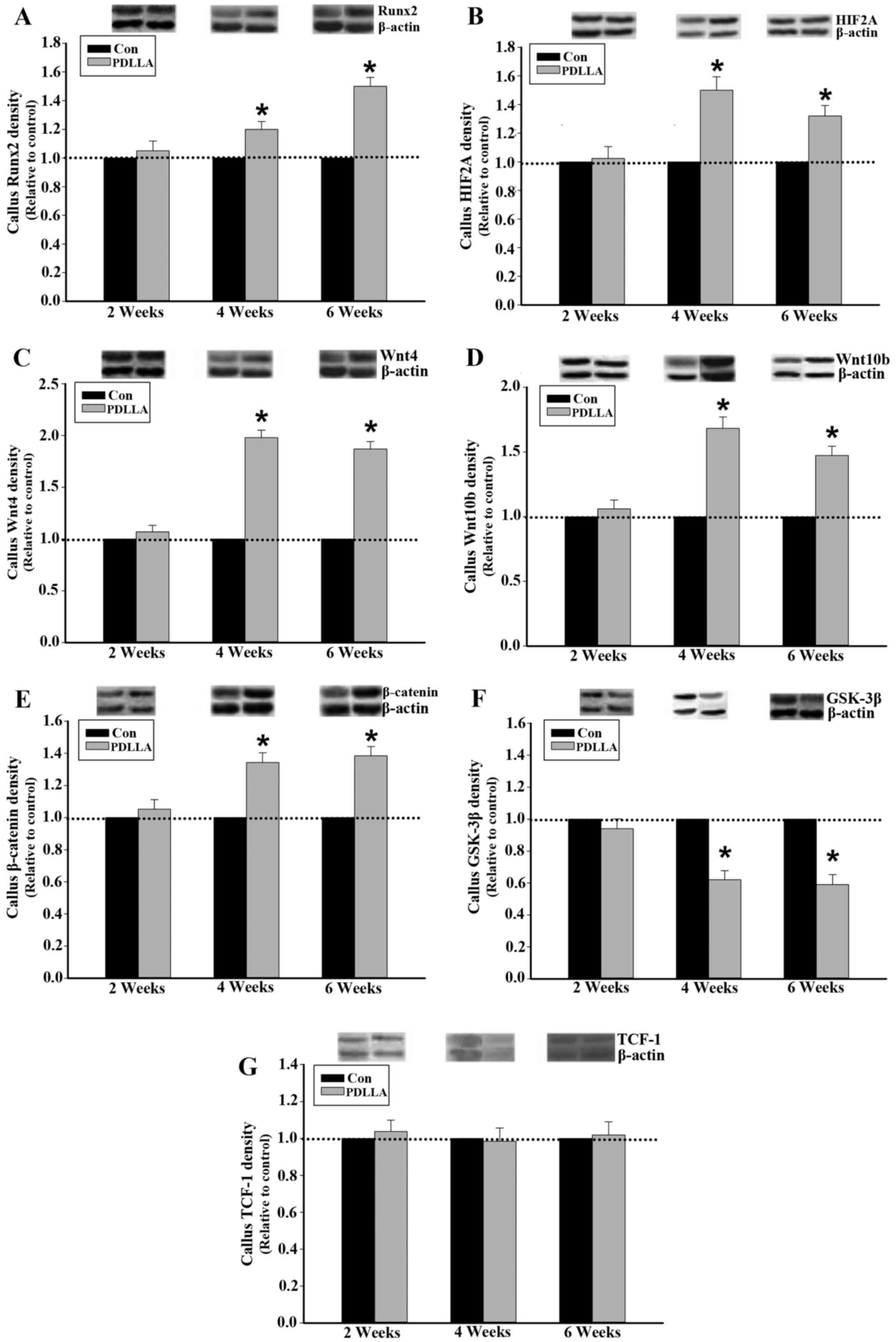 | Figure 2.Protein expression in callus tissue of
control and PDLLA treated rats 2, 4 and 6 weeks post-surgery.
Expression level of (A) Runx2, (B) HIF2A, (C) Wnt4, (D) Wnt10b, (E)
GSK-3β, (F) β-catenin and (G) TCF-1 under PDLLA intramedullary
fixation (PDLLA group) compared to that of Kirschner wire (control
group). Β-actin is a control. Values are presented as mean ±
standard deviation, *P<0.05. Con, control group; PDLLA, Poly-D-L
lactide treated group; Runx2, Runt-related transcription factor 2;
HIF2A, hypoxia inducible factor 2A; GSK-3β, glycogen synthase
kinase-3β; TCF-1, T cell factor-1. |
Runx2, HIF2A, Wnts, GSK-3β, β-catenin
and TCF-1 mRNA in callus tissue
It was demonstrated following RT-qPCR that, similar
to the of protein expression, 4 and 6 weeks following the
intramedullary fixation for femoral osteotomy, the expression of
Runx2, HIF2A, Wnts and β-catenin mRNA in callus tissue were
significantly increased in the PDLLA group compared with that of
the control (P<0.05; Fig. 3A-E).
The mRNA expression levels of GSK-3β in callus tissue, at weeks 4
and 6 were significantly decreased in the PDLLA group compared with
the control (P<0.05). However, 2 weeks after the intramedullary
fixation, the level of mRNA expression observed for Runx2, HIF2A,
Wnts, β-catenin and GSK-3β exhibited no significant difference in
the PDLLA group compared with the control (Fig. 3A-F; P>0.05). With respect to
TCF-1, at weeks 2, 4 and 6 following the intramedullary fixation
for femoral osteotomy, the mRNA expression levels exhibited no
significant differences in PDLLA group compared to that of control
(Fig. 3G; P>0.05).
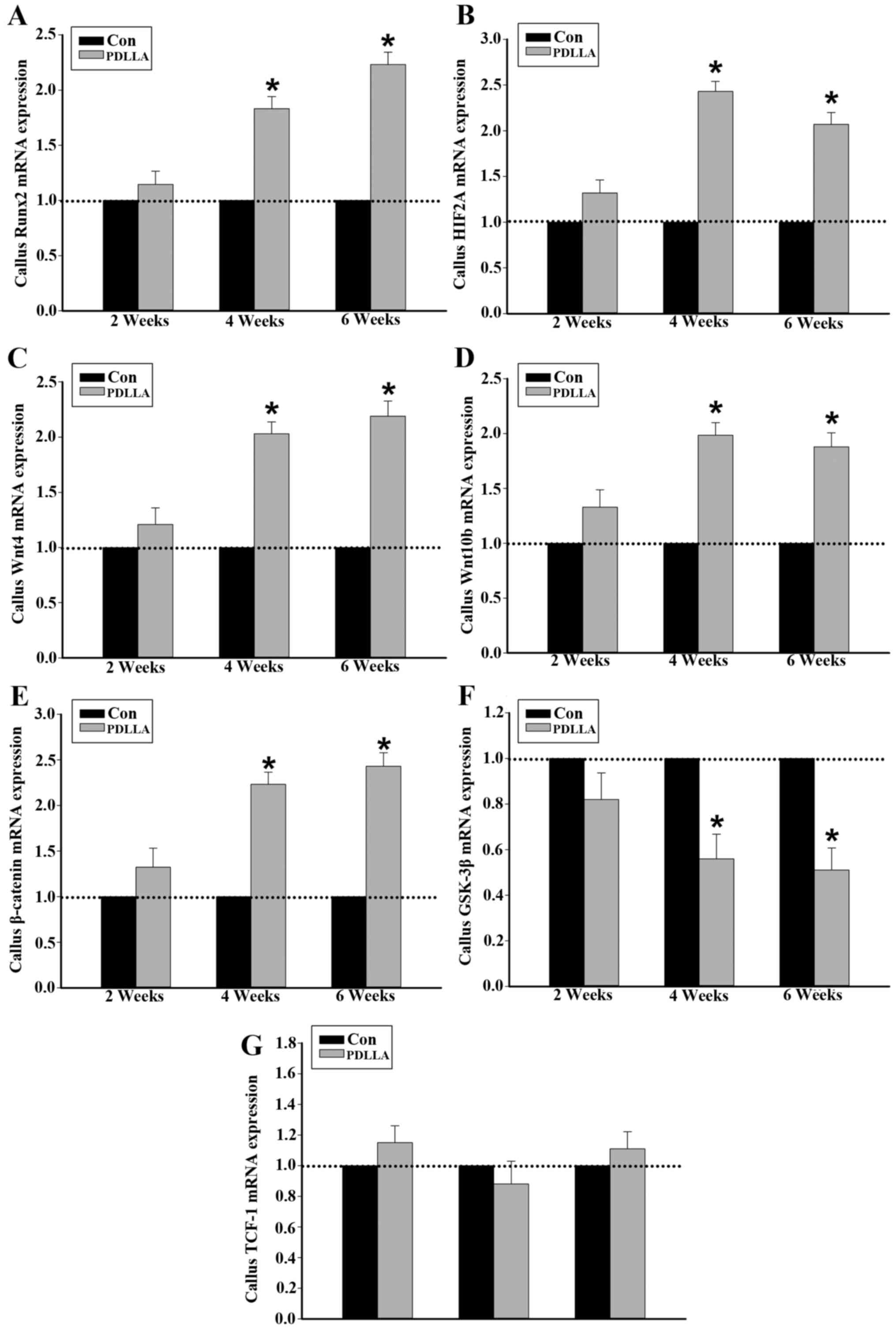 | Figure 3.mRNA expression in callus tissue of
control and PDLLA treated rats 2, 4 and 6 weeks post-surgery.
Expression level of (A) Runx2, (B) HIF2A, (C) Wnt4, (D) Wnt10b, (E)
GSK-3β, (F) β-catenin and (G) TCF-1. Β-actin is a control. Values
are presented as mean ± standard deviation, *P<0.05. Con,
control group; PDLLA, Poly-D-L lactide treated group; Runx2,
Runt-related transcription factor 2; HIF2A, hypoxia inducible
factor 2A; GSK-3β, glycogen synthase kinase-3β; TCF-1, T cell
factor-1. |
Discussion
In the present study, the fracture bone repair under
intramedullary fixation was improved following the use of PDLLA
instead of traditional metal material. Furthermore, compared with
the traditional metal material, the expression of Runx2 in callus
tissue, which is closely associated with the further activation of
canonical Wnt signaling, was significantly increased under PDLLA
intramedullary fixation.
With the development and improvement of pixel
resolution and the mathematical combination with finite element
analysis, microCT has been widely applied in assessing the
biomechanical features and qualities, including bone mineral
density and Young's modulus, of cortical and trabecular bone
(22). In the present study,
microCT-scanning results indicated that following 4 and 6 weeks
post-surgery, the fracture repair indices, including BV/TV, BS/BV,
TB.TH, TB.N and TB.SP, were significantly improved in the PDLLA
group compared with the control group. ALK and OPG were expressed
in the bone tissue, specifically in osteoblastic lineages and their
serum concentrations were considered as the markers for the
evaluation of osteogenesis during development and bone formation
throughout fracture healing (23,24). The
results of the current study indicate that at weeks 4 and 6
following intramedullary fixations for osteotomy, the serum levels
of ALK and OPG were significantly increased in the PDLLA group,
compared with the control. Thus, the current study indicates that
compared to the traditional metal material, the osteogenesis in the
process of fracture healing was increasingly activated and
therefore further improved the curative efficacy of PDLLA
intramedullary fixation for fracture treatment.
It has been indicated that the Runx2 is extensively
expressed in the callus tissue during the process of fracture
healing (25). In the present study,
the level of mRNA and protein expression of Runx2 in the callus
tissue of the PDLLA group were significantly increased, compared to
that of control, at 4 and 6 weeks following intramedullary fixation
for osteotomy. Genetic engineering studies demonstrated that the
altered expression of Runx2 may lead to abnormal and lethal bone
development and ossification (26).
Studies have indicated that Runx2 may serve as an essential
transcriptional factor to extensively regulate the osteogenesis
mediated by a number of pathways, including the differentiation and
maturation of osteoblasts and osteoclasts, expression of bone
matrix protein genes and angiogenesis controlled by vascular
endothelial growth factor (27,28).
Thus, the present study proposed that the improved curative
efficacy of PDLLA in the intramedullary fixation for traumatic
fracture was closely associated with the increased expression of
Runx2 in callus tissue; an important regulator for osteogenesis in
the process of traumatic bone repair. However, the pathways
involved and the aforementioned underlying cellular and molecular
mechanisms may be differentially regulated by the increased
expression of Runx2, thus more studies are required.
The Runx2 gene spans ~210 kb with its two
predominant transcripts, governed by P1 and P2 promoters (29,30).
Runx2 RNA, driven by the P1 promoter, is extensively stimulated and
highly expressed in osteoblastic lineages during skeletal
development (31). Previous results
have demonstrated that the specific loss of expression of the
Runx2-P1 promoter in mice leads to severe developmental defects
with cleidocranial dysplasia-like symptoms (32). Thus, the activity of the P1 promoter
serves a crucial role in the regulation of Runx2 transcription in
the process of osteogenesis. Previous experiments indicated that
HIF2A may bind to the −155 and −75 bp region of the Runx2-P1
promoter and serve as upstream regulators for the basal
transcription of the Runx2 gene (33). In the present study, results
indicated that 4 and 6 weeks following intramedullary fixation for
osteotomy, the mRNA and protein expression of HIF2A in callus
tissue were significantly increased in the PDLLA group compared to
the control. It was therefore hypothesized that, compared to
traditional metal material, the use of PDLLA for internal fixation
of fractures increases the expression of HIF2A. This further
promotes the transcriptional activity of Runx2 through an
interaction between HIF2A and the Runx2-P1 promoter during the
process of traumatic bone repair (34). Notably, various studies indicated
that, from the second week following PDLLA implantation, the
material began to be degraded (35,36). The
biodegraded monomer would then enter into the citric acid cycle to
yield carbon dioxide and water (35). As a consequence, when compared to the
traditional metal material, the oxygen level in the local repair
tissue decreases at weeks 4 and 6 following intramedullary
fixations by PDLLA. Thus, under PDLLA internal fixation for
fracture treatment, the expression of HIF2A may have increased due
to the relative hypoxia condition in the callus tissue. However,
the concerned data supporting this hypothesis in this study is
lacking and the detailed mechanism requires further study to be
elaborated.
It is acknowledged that the canonical Wnt signaling
may govern the lineage commitment and differentiation of progenitor
cells, specifically the mesenchymal stem cells, into the
chondrocytes and osteoblasts, so as to serve the critical roles in
the process of osteogenesis (37).
Gain- and loss-of-function mutations in canonical Wnt signaling
components including Wnt ligands have elaborated the critical
functions of canonical Wnt signaling during endochondral bone
formation (15). The canonical Wnt
signaling is initiated by the binding of Wnt proteins and its Fz
receptors. Following the activation of the Fz receptor complex, the
protein phosphorylation cascade, triggered by GSK-3β, may be
inhibited to stabilize the intracellular β-catenin levels (38). Subsequently, β-catenin is
translocated to the nucleus to form a heterodimeric DNA-binding
complex with TCF-1 to regulate the transcription activity of
downstream genes such as connexin 43 and Runx2 (39). The current study demonstrated that at
weeks 4 and 6 following intramedullary fixation for osteotomy, the
mRNA and protein expression levels of Wnt4, Wnt10b and β-catenin
were significantly increased, while mRNA and protein expression
levels of GSK-3β were significantly decreased in the PDLLA group
compared with the control. Consequently, it is suggested that,
compared to the traditional metal material, the canonical Wnt
signaling pathway was further activated under PDLLA internal
fixation for fracture treatment. Notably, previous findings
indicated that, the heterodimeric DNA-binding complex of β-catenin
and TCF-1 may target the −97 and −93 bp region of the Runx2-P1
promoter, enhancing the transcription activity of Runx2 gene
(19). Although the expression of
TCF-1 was not changed, the increased expression of β-catenin may,
at least in part, contribute to further activation and
transcriptional activity of Runx2 gene under PDLLA intramedullary
fixation compared to that of metal material. The binding sites for
β-catenin, TCF-1 and HIF2A in the P1 promoter region of the Runx2
gene overlapped, the current study hypothesized that the function
of β-catenin, TCF-1 and HIF2A may synergistically involve the
transcriptional regulation of Runx2 gene.
It should be noted that at week 2 following the
intramedullary fixation for osteotomy, compared with traditional
metal materials, all parameters detected in the current study
presented little difference following the use of PDLLA
biodegradable material. A number of potential reasons for this are
as follows: In one aspect, previous results have indicated that
PDLLA degrades relatively slowly in the first two weeks in
vivo (40). This indicated that
the mechanical strength PDLLA is greater than the callus tissue and
the released monomer and changes to microenvironment in local
tissue are limited. As such, compared to the traditional metal
material, the differences concerning stress shielding and local
inflammatory effects induced by PDLLA would be negligible.
Secondly, during the first two weeks following the fracture in
rats, the bone repair process is still in the inflammatory and
initial repair phases (9,10). This indicates that the biological
processes of fracture healing were in the most active period and
may not be clearly altered by the relatively subtle differences
between the traditional metal and PDLLA materials. Over time, the
bone repair process moves into the late repair and remodeling
phases and new blood vessels also become mature. Furthermore, the
degradation process of PDLLA material in vivo would become
more active and the differences between traditional metal and PDLLA
materials, including stress shielding and local inflammatory
effects, would become progressively apparent. However, further
investigation is required to assess which of the aforementioned
factors serve as the trigger and are therefore important for
changing the pathways examined in this study.
In conclusion, the findings of the present study
provide a novel insight into the differences of cellular and
molecular pathways in the process of bone repair under the internal
fixation for traumatic fracture treatment using PDLLA, a
biodegradable material compared to that of traditional metal
materials. Although these results were obtained from the rat model,
the present study may facilitate further studies of the relevant
biological mechanisms underlying PDLLA in clinical use.
Acknowledgements
The present study was supported by research grants
from Suzhou Science and Education Project (no. KJXW2014010) and
Suzhou Science and Technology Project (no. SYS201405).
References
|
1
|
Rokkanen P, Böstman O, Vainionpää S,
Vihtonen K, Törmälä P, Laiho J, Kilpikari J and Tamminmäki M:
Biodegradable implants in fracture fixation: Early results of
treatment of fractures of the ankle. Lancet. 1:1422–1424. 1985.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Zantop T, Weimann A, Schmidtko R, Herbort
M, Raschke MJ and Petersen W: Graft laceration and pullout strength
of soft-tissue anterior cruciate ligament reconstruction: In vitro
study comparing titanium, poly-d,l-lactide, and
poly-d,l-lactide-tricalcium phosphate screws. Arthroscopy.
22:1204–1210. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Mukherjee DP and Pietrzak WS:
Bioabsorbable fixation: Scientific, technical, and clinical
concepts. J Craniofac Surg. 22:679–689. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Schrumpf MA, Lee AT and Weiland AJ:
Foreign-body reaction and osteolysis induced by an intraosseous
poly-L-lactic Acid suture anchor in the wrist: Case report. J Hand
Surg Am. 36:1769–1773. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kim SH, Oh JH, Lee OS, Lee HR and Hargens
AR: Postoperative imaging of bioabsorbable anchors in rotator cuff
repair. Am J Sports Med. 42:552–557. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Fu D, Xiao B, Yang S and Li J: Open
reduction and bioabsorbable pin fixation for late presenting
irreducible supracondylar humeral fracture in children. Int Orthop.
35:725–730. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Waris E, Ashammakhi N, Raatikainen T,
Törmälä P, Santavirta S and Konttinen YT: Self-reinforced
bioabsorbable versus metallic fixation systems for metacarpal and
phalangeal fractures: A biomechanical study. J Hand Surg Am.
27:902–909. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Waris E, Ashammakhi N, Kaarela O,
Raatikainen T and Vasenius J: Use of bioabsorbable osteofixation
devices in the hand. J Hand Surg Br. 29:590–598. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Claes L, Recknagel S and Ignatius A:
Fracture healing under healthy and inflammatory conditions. Nat Rev
Rheumatol. 8:133–143. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Gerstenfeld LC, Cullinane DM, Barnes GL,
Graves DT and Einhorn TA: Fracture healing as a post-natal
developmental process: Molecular, spatial, and temporal aspects of
its regulation. J Cell Biochem. 88:873–884. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Rennert J, Coffman JA, Mushegian AR and
Robertson AJ: The evolution of Runx genes I. A comparative study of
sequences from phylogenetically diverse model organisms. BMC Evol
Biol. 3:42003. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Bustamante M, Nogués X, Agueda L, Jurado
S, Wesselius A, Cáceres E, Carreras R, Ciria M, Mellibovsky L,
Balcells S, et al: Promoter 2 −1025 T/C polymorphism in the RUNX2
gene is associated with femoral neck bmd in Spanish postmenopausal
women. Calcif Tissue Int. 81:327–332. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Qin X, Jiang Q, Matsuo Y, Kawane T, Komori
H, Moriishi T, Taniuchi I, Ito K, Kawai Y, Rokutanda S, et al: Cbfb
regulates bone development by stabilizing Runx family proteins. J
Bone Miner Res. 30:706–714. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wu M, Li C, Zhu G, Wang Y, Jules J, Lu Y,
McConnell M, Wang YJ, Shao JZ, Li YP and Chen W: Deletion of
core-binding factor β (Cbfβ) in mesenchymal progenitor cells
provides new insights into Cbfβ/Runxs complex function in cartilage
and bone development. Bone. 65:49–59. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Xu H, Duan J, Ning D, Li J, Liu R, Yang R,
Jiang JX and Shang P: Role of Wnt signaling in fracture healing.
BMB Rep. 47:666–672. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Eastman Q and Grosschedl R: Regulation of
LEF-1/TCF transcription factors by Wnt and other signals. Curr Opin
Cell Biol. 11:233–240. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhong N, Gersch RP and Hadjiargyrou M: Wnt
signaling activation during bone regeneration and the role of
Dishevelled in chondrocyte proliferation and differentiation. Bone.
39:5–16. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Chen Y, Whetstone HC, Lin AC, Nadesan P,
Wei Q, Poon R and Alman BA: Beta-catenin signaling plays a
disparate role in different phases of fracture repair: Implications
for therapy to improve bone healing. PLoS Med. 4:e2492007.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Gaur T, Lengner CJ, Hovhannisyan H, Bhat
RA, Bodine PV, Komm BS, Javed A, van Wijnen AJ, Stein JL, Stein GS
and Lian JB: Canonical WNT signaling promotes osteogenesis by
directly stimulating Runx2 gene expression. J Biol Chem.
280:33132–33140. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Rosier RN, O'Keefe RJ and Hicks DG: The
potential role of transforming growth factor beta in fracture
healing. Clin Orthop Relat Res. (Suppl 355). S294–S300. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Mabilleau G, Mieczkowska A, Libouban H,
Simon Y, Audran M and Chappard D: Comparison between quantitative
X-ray imaging, dual energy X-ray absorptiometry and microCT in the
assessment of bone mineral density in disuse-induced bone loss. J
Musculoskelet Neuronal Interact. 15:42–52. 2015.PubMed/NCBI
|
|
23
|
Yonden Z, Aydin M, Alcin E, Kelestemur MH,
Kutlu S and Yilmaz B: Effects of letrozole on bone biomarkers and
femur fracture in female rats. J Physiol Biochem. 65:267–275. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Fahrleitner-Pammer A, Dobnig H,
Piswanger-Soelkner C, Bonelli C, Dimai HP, Leb G and
Obermayer-Pietsch B: Osteoprotegerin serum levels in women:
Correlation with age, bone mass, bone turnover and fracture status.
Wien Klin Wochenschr. 115:291–297. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Yee CS, Xie L, Hatsell S, Hum N, Murugesh
D, Economides AN, Loots GG and Collette NM: Sclerostin antibody
treatment improves fracture outcomes in a Type I diabetic mouse
model. Bone. 82:122–134. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Komori T, Yagi H, Nomura S, Yamaguchi A,
Sasaki K, Deguchi K, Shimizu Y, Bronson RT, Gao YH, Inada M, et al:
Targeted disruption of Cbfa1 results in a complete lack of bone
formation owing to maturational arrest of osteoblasts. Cell.
89:755–764. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Inada M, Yasui T, Nomura S, Miyake S,
Deguchi K, Himeno M, Sato M, Yamagiwa H, Kimura T, Yasui N, et al:
Maturational disturbance of chondrocytes in Cbfa1-deficient mice.
Dev Dyn. 214:279–290. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Ueta C, Iwamoto M, Kanatani N, Yoshida C,
Liu Y, Enomoto-Iwamoto M, Ohmori T, Enomoto H, Nakata K, Takada K,
et al: Skeletal malformations caused by overexpression of Cbfa1 or
its dominant negative form in chondrocytes. J Cell Biol.
153:87–100. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zhang Y, Hassan MQ, Montecino M, Hawse JR,
Spelsberg TC, Stein JL, Lian JB, van Wijnen AJ and Stein GS:
Synergistic regulation of the Runx2 P1 promoter in mesenchymal
cells by a conserved HLH box and purine-rich elements (GAY motifs).
FASEB J. 22:(Suppl 1). S782.7172008.
|
|
30
|
Doecke JD, Day CJ, Stephens AS, Carter SL,
van Daal A, Kotowicz MA, Nicholson GC and Morrison NA: Association
of functionally different RUNX2 P2 promoter alleles with BMD. J
Bone Miner Res. 21:265–273. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Harada H, Tagashira S, Fujiwara M, Ogawa
S, Katsumata T, Yamaguchi A, Komori T and Nakatsuka M: Cbfa1
isoforms exert functional differences in osteoblast
differentiation. J Biol Chem. 274:6972–6978. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Liu JC, Lengner CJ, Gaur T, Lou Y, Hussain
S, Jones MD, Borodic B, Colby JL, Steinman HA, van Wijnen AJ, et
al: Runx2 protein expression utilizes the Runx2 P1 promoter to
establish osteoprogenitor cell number for normal bone formation. J
Biol Chem. 286:30057–30070. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Tamiya H, Ikeda T, Jeong JH, Saito T, Yano
F, Jung YK, Ohba S, Kawaguchi H, Chung UI and Choi JY: Analysis of
the Runx2 promoter in osseous and non-osseous cells and
identification of HIF2A as a potent transcription activator. Gene.
416:53–60. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Hirata M, Kugimiya F, Fukai A, Saito T,
Yano F, Ikeda T, Nakamura K, Chung UI and Kawaguchi H: 060
Molecular network on the C/Ebp-Beta axis including Runx2, Mmp13,
and Hif2a controls osteoarthritis development. Osteoarthritis
Cartilage. 18:(Suppl 2). S342010. View Article : Google Scholar
|
|
35
|
Heidemann W, Jeschkeit S, Ruffieux K,
Fischer JH, Wagner M, Krüger G, Wintermantel E and Gerlach KL:
Degradation of poly (D,L)lactide implants with or without addition
of calciumphosphates in vivo. Biomaterials. 22:2371–2381. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Jansen J, Koopmans SA, Los LI, van der
Worp RJ, Podt JG, Hooymans JM, Feijen J and Grijpma DW: Intraocular
degradation behavior of crosslinked and linear poly(trimethylene
carbonate) and poly(D,L-lactic acid). Biomaterials. 32:4994–5002.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Carroll SH and Ravid K: Differentiation of
mesenchymal stem cells to osteoblasts and chondrocytes: A focus on
adenosine receptors. Expert Rev Mol Med. 15:e12013. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Kim JH, Liu X, Wang J, Chen X, Zhang H,
Kim SH, Cui J, Li R, Zhang W, Kong Y, et al: Wnt signaling in bone
formation and its therapeutic potential for bone diseases. Ther Adv
Musculoskelet Dis. 5:13–31. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Monroe DG, McGee-Lawrence ME, Oursler MJ
and Westendorf JJ: Update on Wnt signaling in bone cell biology and
bone disease. Gene. 492:1–18. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Simon JA, Ricci JL and Di Cesare PE:
Bioresorbable fracture fixation in orthopedics: A comprehensive
review. Part I. Basic science and preclinical studies. Am J Orthop
(Belle Mead NJ). 26:665–671. 1997.PubMed/NCBI
|