Introduction
Curcuma longa (turmeric) is a rhizomatous
herbaceous perennial plant of the ginger family, Zingiberaceae. It
is also known as kasturi manjal, and has been used in Siddha
medicine for more than a thousand years in Asia (1) as a medicine for various ailments,
including skin diseases, pulmonary issues, aches, pains, wounds,
sprains and gastrointestinal system issues (2). Furthermore, it has been used to treat
stomach and liver diseases, and it contains a high antimicrobial
potential (3). The active component
of C. longa extract is a curcuminoid (4). C. longa extract is known to have
anti-inflammatory (5,6), antioxidant (7) and anti-depressant potential (8–11).
Previous studies have reported that C. longa extract is very
effective against inflammation and Alzheimer's disease (12,13).
Parkinson's disease (PD) is a neurodegenerative
disorder characterized by damage to the dopamine-producing cells in
the substantia nigra of the brain. Environment factors and genetic
predisposition have a crucial role in the progression of PD. Tieu
(14) has previously reported that
exposure to several endogenous and exogenous toxins has been
associated with PD. Therefore, these endogenous and exogenous
toxins are often used to study PD. The endogenous toxin, salsolinol
is a neuromodulator of dopamine-producing cells in the substantia
nigra (15). Dysregulation of
salsolinol metabolism is known to contribute to the progressive
development of PD (16). Increased
levels of salsolinol and its derivatives in the cerebrospinal fluid
and urine may serve as an index for the PD (17,18).
Previous studies have reported that the extract of
C. longa may be useful in the treatment of PD (19,20).
Therefore, the present study investigated this effect. The toxicity
of salsolinol was investigated in SH-SY5Y cells derived from human
neuroblastoma cells (21,22), in order to investigate the
therapeutic potential of C. longa extract.
Materials and methods
Materials
SH-SY5Y human neuroblastoma cells were purchased
from American Type Culture Collection (Manassas, VA, USA). Dimethyl
sulphoxide and sulforhodamine B (SRB), Dulbecco's modified Eagle's
medium (DMEM), fetal bovine serum (FBS), penicillin-streptomycin
and trypsin-EDTA were obtained from were obtained from
Sigma-Aldrich (Merck Millipore, Darmstadt, Germany). Acridine
orange (AO), ethidium bromide (EB) Fluorescein diacetate, propidium
iodide (PI) and 2,7-dichlorodihydrofluorescein diacetate (DCFH-DA)
were purchased from Santa Cruz Biotechnology, Inc., (Dallas, TX,
USA).
Cell culture
SH-SY5Y human neuroblastoma cells (sixth passage)
were cultured in DMEM supplemented with 10% FBS and 1%
penicillin-streptomycin. Cells were grown in a CO2
incubator at 37°C and 5% CO2. The protocol for the
present study was approved by the Ethics Committee of the
Affiliated Hospital of Qingdao University.
Preparation of C. longa extracts
Water extraction of C. longa extract was
performed by boiling 100 g in 1,000 ml distilled water for 15 min
over a low flame. The flask was subsequently plugged, removed from
the heat and allowed to cool. The extract was filtered and dried to
prepare the required concentrations after cooling the content of
the flask.
C. longa extract pre-treatment
Various concentrations of C. longa extract
(0.001, 0.01, 0.05, 0.1, 0.2 and 0.4 mg/ml) were added 1 h prior to
the addition of salsolinol to investigate its therapeutic potential
against PD. SRB assay was used to assess cell viability after 24
and 48 h of treatment.
Salsolinol treatment
Once the SH-SY5Y cells reached 90% confluence, the
cells were plated in 96-well plate and allowed to adhere for 24 h.
Various concentrations of salsolinol (0.001, 0.01, 0.05, 0.1, 0.5
and 1 mg/ml) were added to the cells (2×104 cells/well).
SRB assay was used to assess cell viability after 24 and 48 h of
treatment.
SRB assay for cell viability
SH-SY5Y cells were cultured and grown at a density
of 2.5×104 cells/well in 96-well plates. Cells were
treated with C. longa extract and salsolinol at various
concentrations for 24 and 48 h. The cytotoxic effect of salsolinol
and therapeutic potential of C. longa extract on SH-SY5Y
cells were measured by the SRB assay, which was performed as
previously described (23).
Fluorescence microscopy
Cells were cultured and grown at a density of
2.2×104 cells/well into 6-well plates. SH-SY5Y cells
were treated with 0.05 mg/ml salsolinol, 1 h prior to the addition
of C. longa extract (0.05 and 0.1 mg/ml). After 24 and 48 h,
cells were removed from the wells and centrifuged at 500 × g for 3
min. The supernatant was withdrawn from the tubes and
phosphate-buffered saline (PBS) was added to the cells. Cell volume
was adjusted to 105-106 cells/ml. A total of
95 µl cell suspension was added to the microtube, and 5 µl AO and
EB was added to the same tube. Cells were incubated at 37°C for
15–30 min in the dark. Subsequently, 10 µl cell staining solution
was added to a glass slide and covered with a cover glass. Cells
were examined with a fluorescence microscope (Axiovert 2000; Carl
Zeiss AG, Oberkochen, Germany) (24).
Confocal laser scanning microscope
analysis (CLSM)
CLSM is widely used to investigate morphological
features of apoptosis and apoptotic DNA fragmentation (24). AO is an organic compound that is
extensively used as a nucleic acid-selective fluorescent cationic
dye for the determination of the cell cycle (24). AO interacts with DNA and RNA by
intercalation or electrostatic attractions respectively, and
spectrally fluorescein when bound to DNA with an excitation maximum
at 502 nm, and an emission maximum at 525 nm (24). SH-SY5Y cells (2×104
cells/well) were seeded in a confocal dish and subsequently treated
with 0.05 mg/ml salsolinol 1 h prior to the addition of C.
longa extracts (0.05 and 0.1 mg/ml). After 24 and 48 h, cells
were washed thrice with PBS and stained with AO (20 µg/ml) for 5
min. Cells were immediately viewed under a CLSM (1X81R motorized
inverted microscope; Olympus Corp., Tokyo, Japan) (24).
Determination of reactive oxygen
species (ROS) production
ROS was determined using a fluorescent probe,
DCFH-DA (25). SH-SY5Y cells were
seeded in 96-well plates in growth medium at a density of 4,000
cells/well under standard conditions. Cells were treated with 0.05
mg/ml salsolinol, 1 h prior to the addition of C. longa
extract (0.05 and 0.1 mg/ml). After 24 and 48 h, the medium was
removed, and the cells were incubated with 5 µM DCFH-DA in the
growth medium for 30 min at 37°C and 5% CO2.
Fluorescence was measured at 24 and 48 h using a fluorescent plate
reader at excitation⁄emission (490 and 525 nm respectively). Images
were captured using a fluorescence microscope.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
SH-SY5Y cells (2×104 cells/well) were
seeded into a T25 flask and subsequently treated with 0.05 mg/ml
salsolinol, 1 h prior to the addition of C. longa extract
(0.05 and 0.1 mg/ml). After 24 and 48 h, 100 ng total RNA was
isolated from the control and treated samples (26). RNA was reverse transcribed using Taq
DNA polymerase (D1806; Sigma-Aldrich; Merck Millipore) with a total
reaction volume of 25 µl. Following treatment with DNase (AMPD1;
Sigma-Aldrich; Merck Millipore), qPCR was performed using cDNA
equivalent to 10 ng total RNA from each sample in a total reaction
volume of 10 µl, containing SYBR Green (A25741; Thermo Fisher
Scientific, Inc.), 12.5 µl PCR Master Mix (2X), 0.25–2.5 µl
upstream primer (10 µM), 0.25–2.5 µl downstream primer (10 µM), 1–5
µl DNA template and nuclease-free water. Primers were specific for
p53 (forward, 5′-TAACAGTTCCTGCATGGGCGGC-3′ and reverse,
5′-AGGACAGGCACAAACACGCACC-3′), Bcl-2-associated X protein (Bax)
(forward, 5′-TGGAGCTGCAGAGGATGATTG-3′ and reverse,
5′-GAAGTTGCCGTCAGAAAACATG-3′), caspase 3 (forward,
5′-TTAATAAAGGTATCCATGGAGAACACT-3′ and reverse,
5′-TTAGTGATAAAAATAGAGTTCTTTTGTGAG-3′) and a housekeeping gene GAPDH
(forward, 5′-GGTCACCAGGGCTGCTTTT-3′ and reverse,
5′-ATCTCGCTCCTGGAAGATGGT-3′). Thermal cycling conditions were as
follows and were performed on an Applied Biosystems PCR thermal
cycler (Applied Biosystems; Thermo Fisher Scientific, Inc.):
Annealing at 50–68°C for 1 min, followed by 40 cycles at 68°C for 1
min. Relative ratios were calculated based on the 2−ΔΔCq
method (27). PCR was monitored
using the CFX96TM Real-Time System (Bio-Rad Laboratories, Inc.,
Hercules, CA, USA). A total of 25 repeats were performed and GAPDH
was used as the reference gene.
Caspase 3 activity assay
SH-SY5Y cells (2×104 cells/well) were
seeded in the culture dish and subsequently treated with 0.05 mg/ml
of salsolinol, 1 h before the addition of C. longa extract
(0.05 and 0.1 mg/ml). After 24 and 48 h, caspase 3 enzyme activity
was measured based on the method outlined by Muthuraman (28).
Statistical analysis
Data were expressed as the mean ± standard error of
the mean. Differences between control and treated cells were
evaluated using Student's t-test. P<0.05 was considered to
indicate a statistically significant difference.
Results
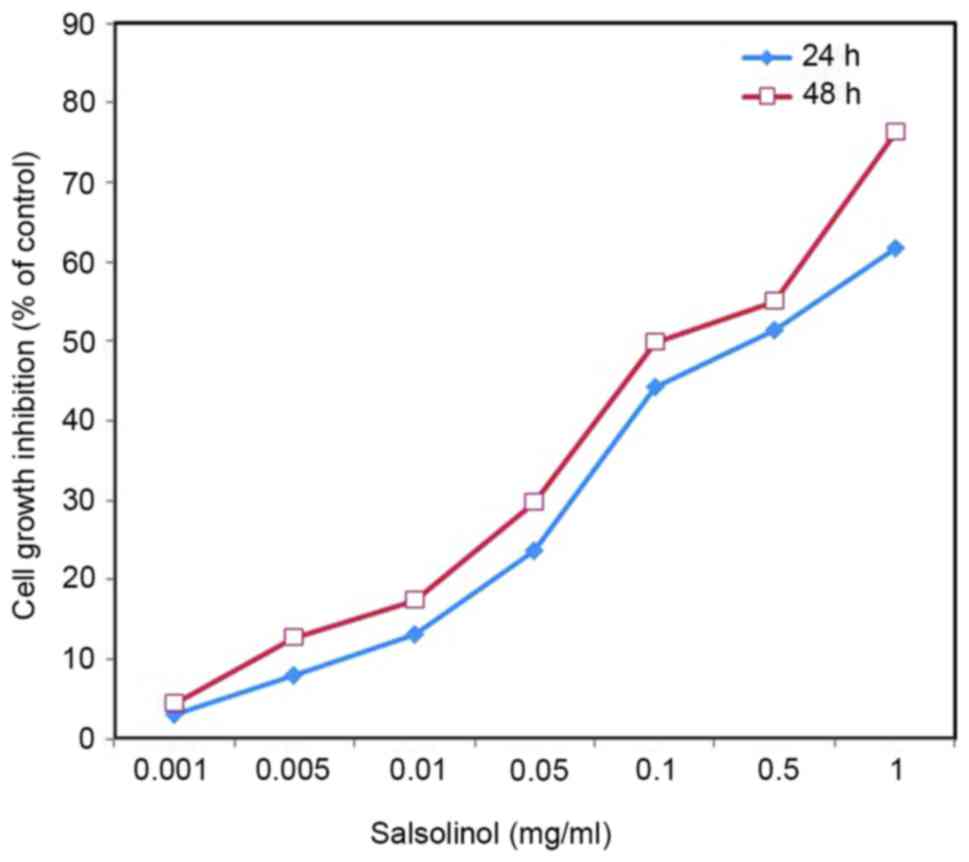
Effect of salsolinol on cell
viability
Salsolinol induced a neurotoxic effect on SH-SY5Y
cells in a dose-dependent manner (Fig.
1). As 1 mg/l salsolinol was able to significantly inhibit the
growth of SH-SY5Y cells, 0.05 mg/l salsolinol was used for
subsequent experiments. An inverted microscope was used to observe
cell shape and morphological changes. Control SH-SY5Y cells
exhibited normal distinct morphology, whereas salsolinol-treated
cells exhibited sporadic distribution, loss of adhesion and a
rounding/oval profile (data not shown).
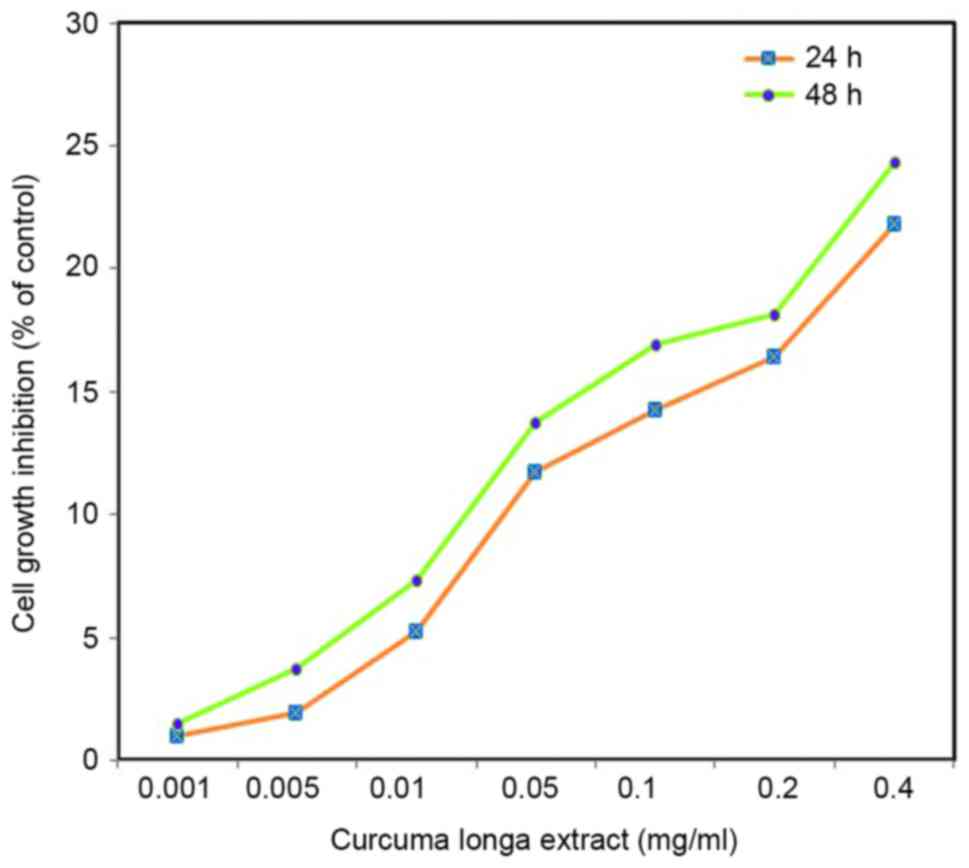
Effect of C. longa extract on cell viability.
C. longa extract had a dose-dependent therapeutic effect on
the viability of SH-SY5Y cells (Fig.
2). As 1 mg/l C. longa extract induced an adverse effect
on normal cells, 0.05 and 0.1 mg/l C. longa extract was used
for subsequent experiments. An inverted microscope was used to
observe cell shape and morphological changes. Control cells
exhibited irregular morphology, such as rounding and a reduction in
nuclear volume. C. longa extract-treated cells demonstrated
a significant reduction in cell growth inhibition (P<0.05;
Fig. 2).
Effect of C. longa extract on apoptosis.
Fluorescence microscopy was performed to determine whether the
neuroprotective impact of C. longa extract was associated
with the induction of apoptosis, by assessing the morphological
features of cell death. This method combines the dual uptake of
fluorescent DNA binding dyes (AO and EB), and chromatin
condensation in the stained nucleus can be used to differentiate
between viable, apoptotic, and necrotic cells (24). Control cells exhibited irregular
morphological features, such as rounding and a reduction in nuclear
volume, whereas C. longa extract-treated cells demonstrated
normal morphological features (Figs.
3 and 4).
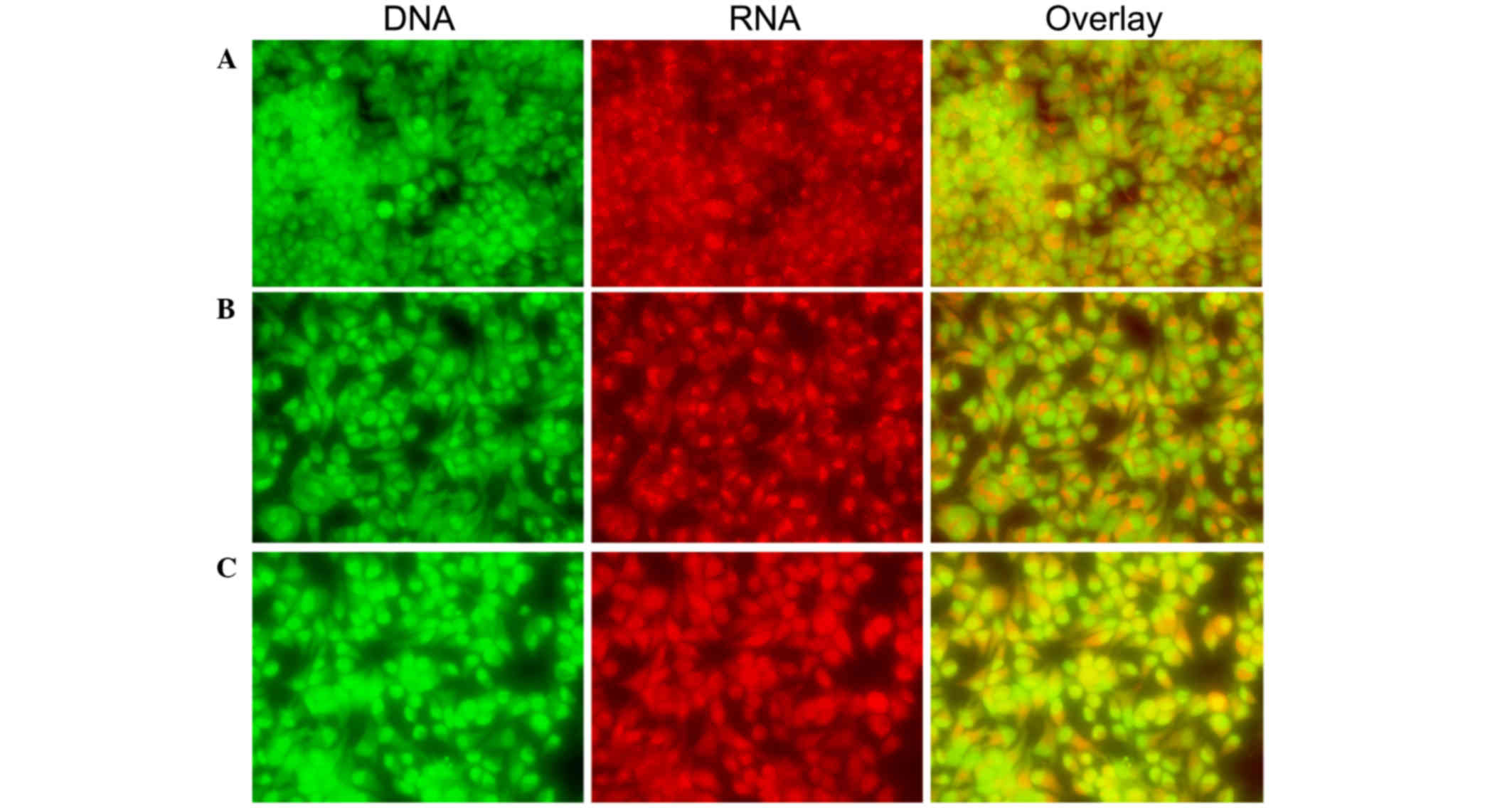
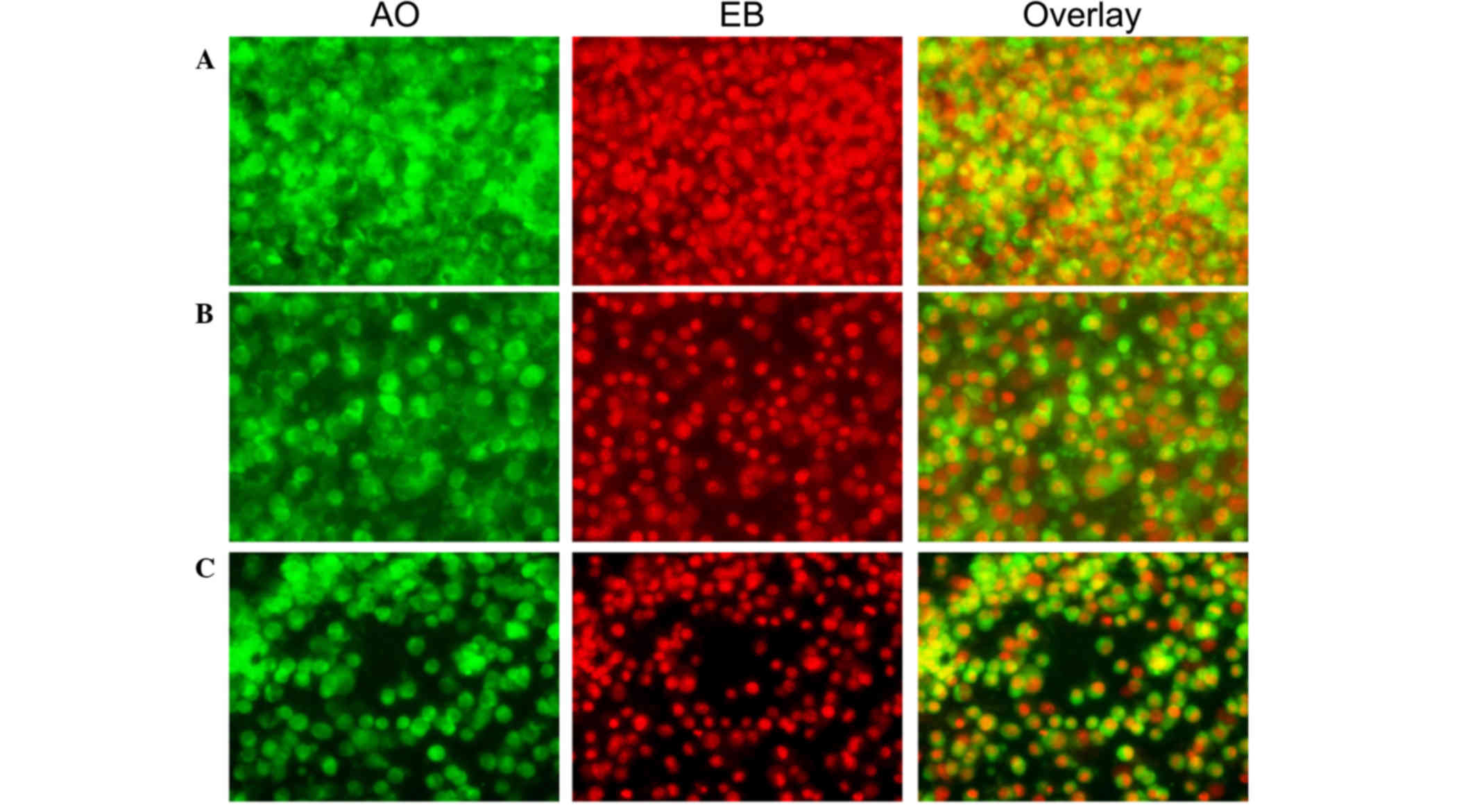 | Figure 3.Morphological observation with AO/EB
double staining by fluorescence microscopy (magnification, ×40).
SH-SY5Y cells were seeded in 6-well plates and following 24-h
adherence, were treated with (A) 0, (B) 0.5 and (C) 0.1 mg/ml
Curcuma longa extract for 24 h. Following treatment, 95 µl
cell suspension was mixed with 5 µl dye mixture, containing 100
mg/l of AO and 100 mg/l of EB in phsophate-buffered saline.
Subsequently, stained cells were visualized immediately under a
fluorescence microscope. Viable cells were detected in the
controls. Fragmented nuclei, condensed chromatin, and apoptotic
cells were detected in the C. longa extract-treated cells.
Representative images from three independent experiments. AO,
acridine orange; EB, ethidium bromide. |
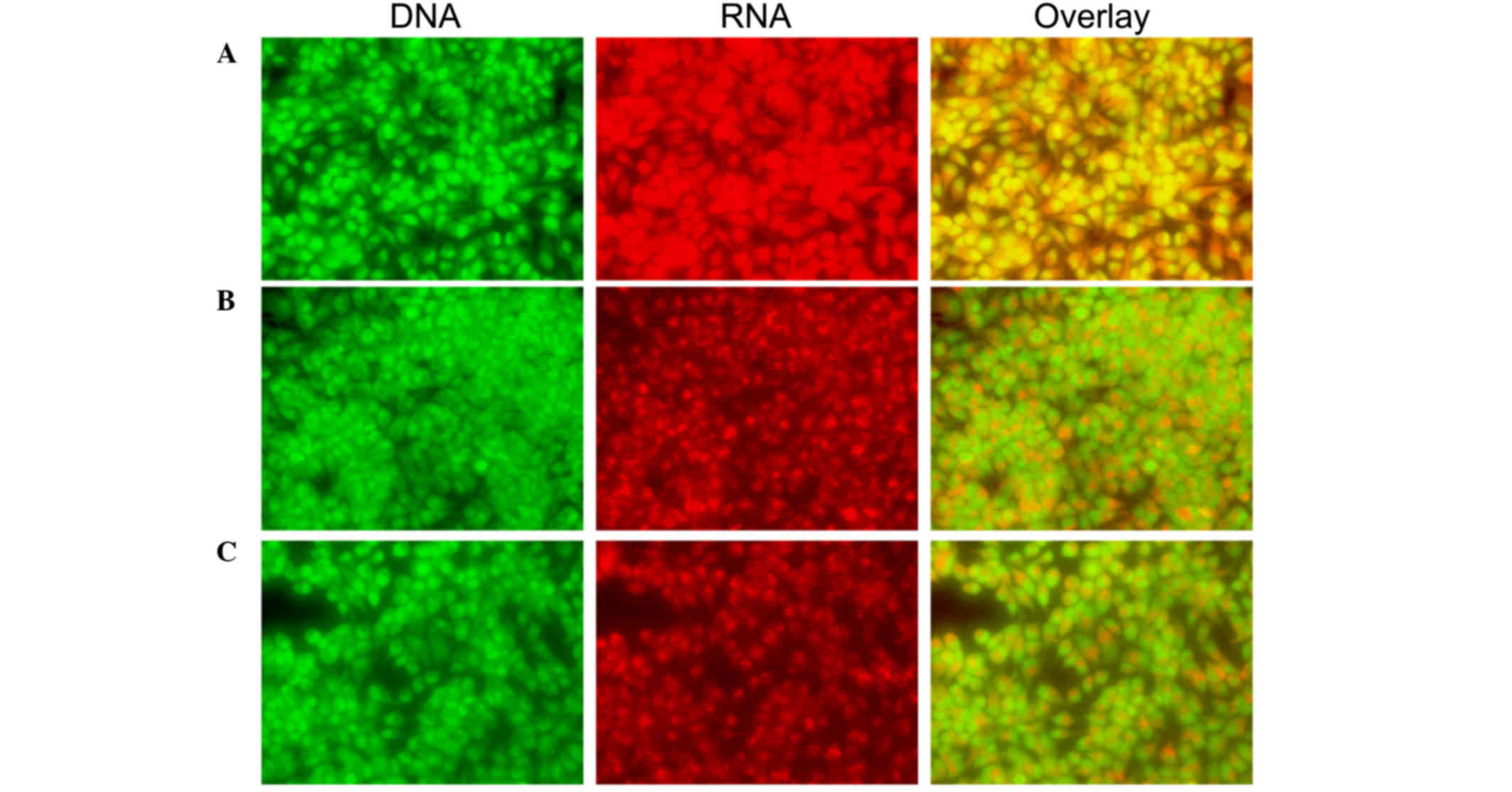
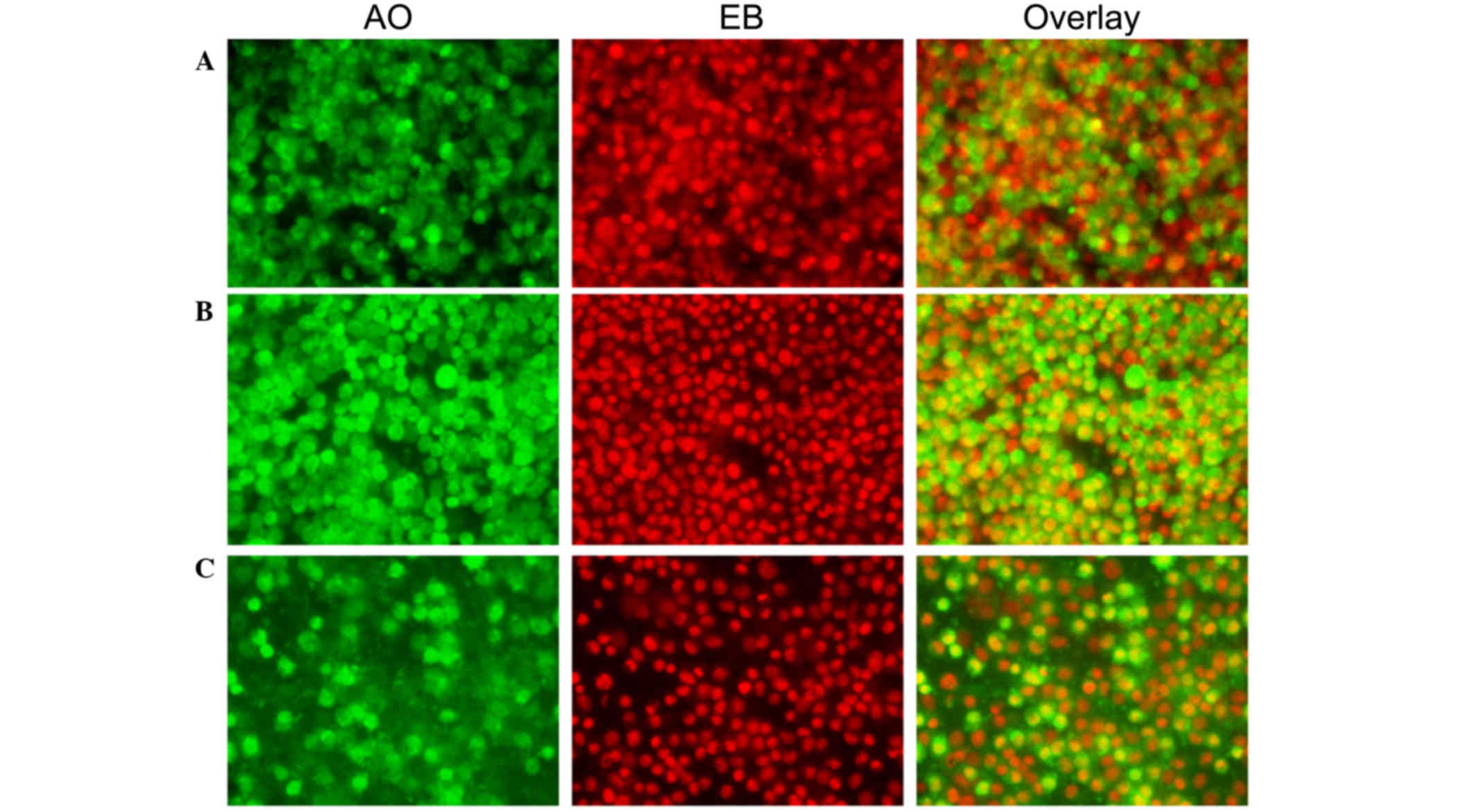 | Figure 4.Morphological observation with AO/EB
double staining by fluorescence microscopy (magnification, ×40).
SH-SY5Y cells were seeded in 6-well plates and, following 48-h
adherence, were treated with (A) 0, (B) 0.5 and (C) 0.1 mg/ml
Curcuma longa extract for 48 h. Following treatment, 95 µl
cell suspension was mixed with 5 µl dye mixture, containing 100
mg/l of AO and 100 mg/l of EB in phsophate-buffered saline.
Subsequently, stained cells were visualized immediately under a
fluorescence microscope. Viable cells were detected in the
controls. Fragmented nuclei, condensed chromatin, and apoptotic
cells were detected in the C. longa extract-treated cells.
Representative images from three independent experiments. AO,
acridine orange; EB, ethidium bromide. |
CLSM was used to investigate the morphological
features of apoptosis and apoptotic DNA fragmentation in the
SH-SY5Y cells. Control cells exhibited irregular morphological
features, such as rounding and a reduction in nuclear volume, as
compared with treated cells. Morphological changes included
rounding, compact granular masses in the nucleus and reduced
nuclear volume in the treated cells. A bright green nucleus
indicated the induction of apoptosis in the treated cells (Figs. 5 and 6).
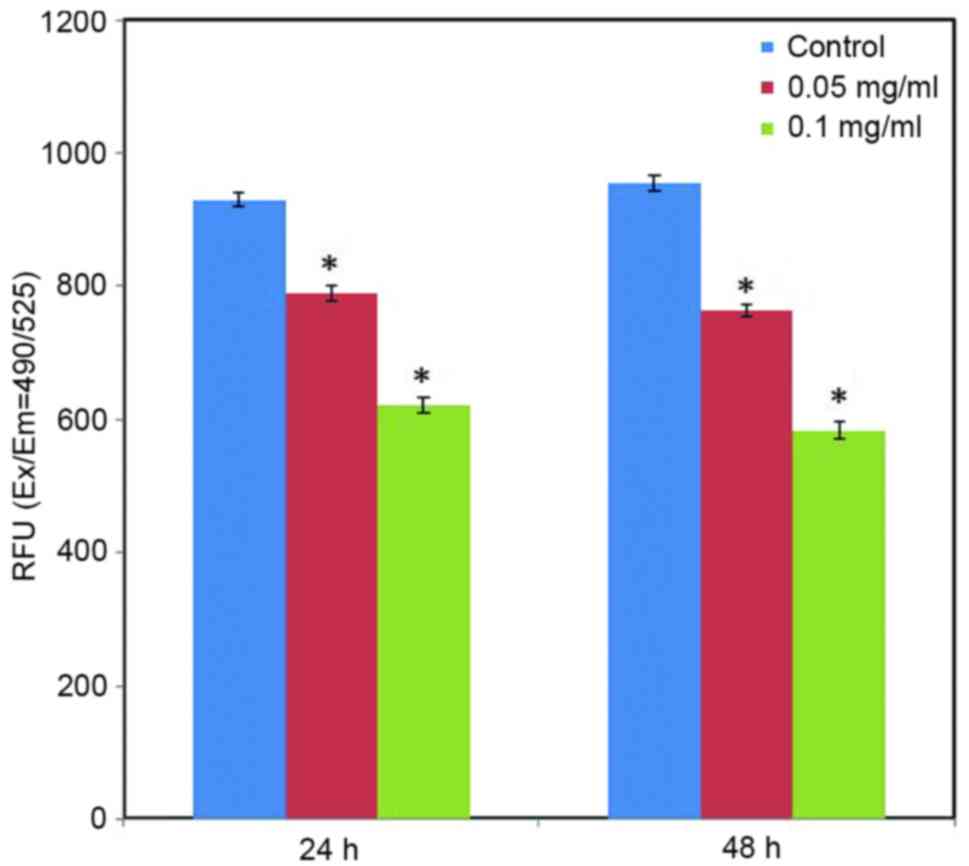
Effect of C. longa extract on ROS level. ROS
have an essential role in facilitating signal transduction
processes within the intracellular region. Intracellular ROS
generation was determined using a DCFH-DA fluorescent probe.
Fluorescence analysis indicated that the green fluorescence
intensity of DCF was significantly reduced in the treated cells, as
compared with the control cells (Fig.
7; P<0.05). Furthermore, fluorescence analysis indicated the
apparent action of C. longa extract on intracellular ROS
generation in a dose-dependent manner (Fig. 7).
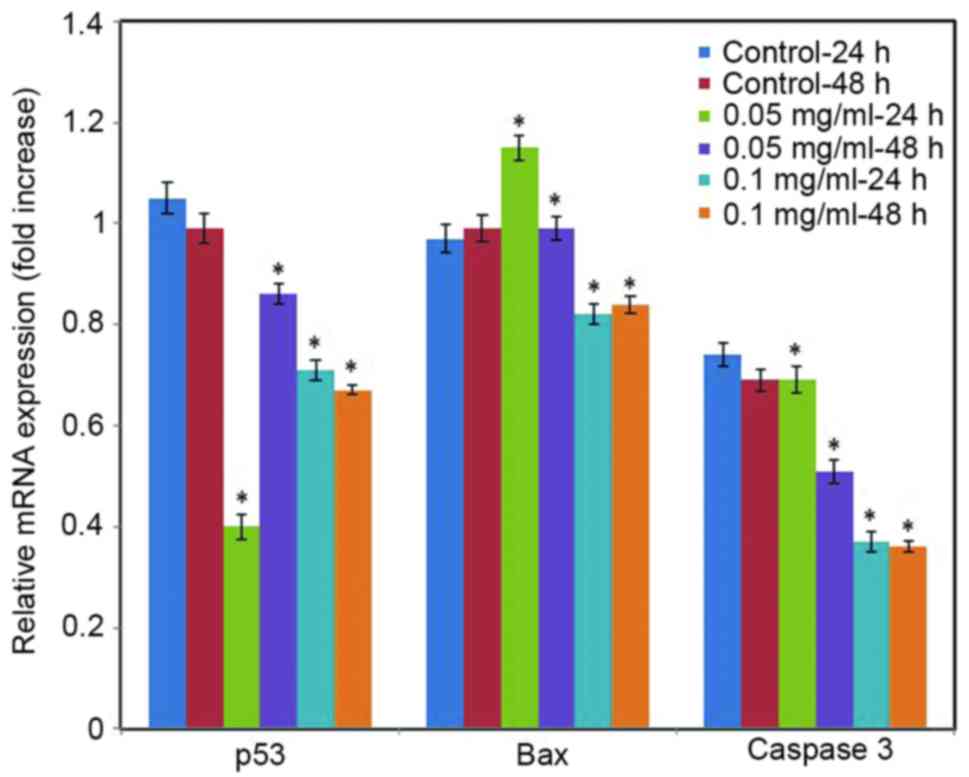
Effect of C. longa extract on gene expression
levels. To further assess the impact of C. longa extract on
apoptosis, RT-qPCR analysis was performed in the present study to
quantify the expression levels of apoptotic markers. Cells exposed
to various concentrations of the C. longa extract exhibited
significantly downregulated mRNA expression levels of p53, Bax and
caspase 3 in a time- and concentration-dependent manner (Fig. 8; P<0.05).
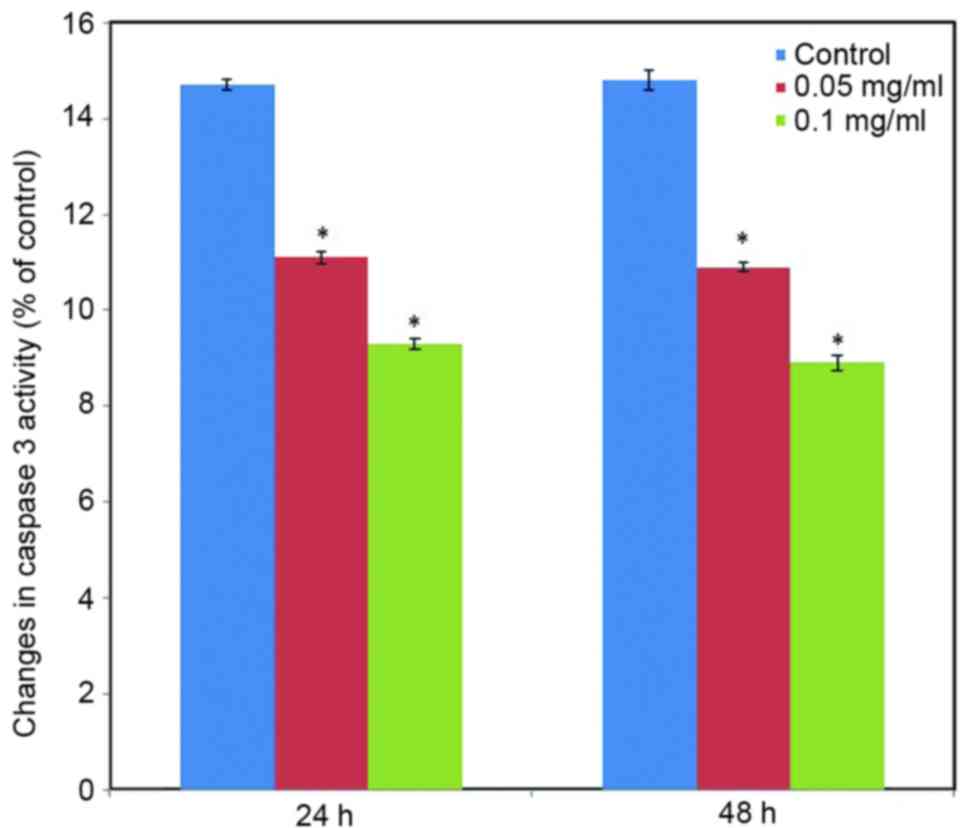
Effect of C. longa extract on caspase 3
activity. To further examine the participation of caspase 3 in the
effect induced by C. longa extract on apoptosis, caspase 3
activity was measured at the translational level. Caspase 3
activity significantly was reduced in the C. longa
extract-treated SH-SY5Y cells (Fig.
9; P<0.05).
Discussion
The results of the present study demonstrated that
C. longa extract has a neuroprotective effect in SH-SY5Y
cells and seemed to exhibit a clear dose-dependence when used in
higher concentrations. Induction of tumor cell apoptosis is an
essential mechanism of anti-cancer compounds (29). Apoptosis is characterized by
morphological and biochemical changes and, the apoptosis of
different cells in the same tissue does not occur at the same time.
Morphological observation is critical for the assessment of
apoptosis in the early stage as DNA multiples may be observed after
the initiation of apoptosis. In the present study, C. longa
extract exerted a therapeutic effect on SH-SY5Y cells at various
concentrations. Morphological observation was performed to
investigate whether the cytotoxic impact of the C. longa
extract was correlated with the apoptotic process.
The findings showed that salsolinol is able to
induce neurotoxicity in SH-SY5Y cells (30,31), and
the effect of treatment with C. longa extract suggested a
therapeutic potential in the PD. Treatment with a low concentration
of C. longa extract was able to significantly protect
against the neurotoxicity induced by salsolinol in the SH-SY5Y
cells. Salsolinol has been demonstrated to induce neurotoxicity via
the inhibition of mitochondrial complex II, and the initiation of
apoptosis through the increased production of free radicals
(32). Salsolinol may contribute the
pathogenesis of PD through the formation of inflammation as the
neurotoxicity induced by salsolinol is mediated through apoptosis,
and is caspase 3 dependent (33).
C. longa extract is known to induce
neuroprotective effects through the inhibition of apoptosis with a
consequent reduction in the gene expression levels of apoptotic
markers, including p53, Bax and caspase 3. Even when administered
at high concentrations, the C. longa extract was able to
partially recover/inhibit the toxicity induced by salsolinol in
SH-SY5Y cells. However, the effect of C. longa extract was
demonstrated to be significant. Subsequent investigation of C.
longa extract in combination with other agents is required to
elucidate their potentially synergistic actions (34), and it would be helpful to determine
the therapeutic effect of C. longa extract as a monotherapy
and in combination with other agents.
Antioxidant and anti-inflammatory properties of the
C. longa extract would be helpful in the treatment of PD
(5). There are several studies
suggesting a significant co-morbidity between depression and PD
(35). The mood regulating potential
of the C. longa extract would be additionally helpful it
treating PD (11).
In conclusion, salsolinol was demonstrated to be
cytotoxic to SH-SY5Y cells when observed by SRB assay. Notably,
this cytotoxicity was reversed by treatment with the C.
longa, as demonstrated by SRB assay. Morphological and
apoptotic changes were evaluated by fluorescence microscopy and
CLSM. RT-qPCR demonstrated that the gene expression levels of
apoptotic markers were downregulated, indicating that treatment
with C. longa extract reduced apoptosis. Furthermore,
caspase 3 activity decreased, indicating that the autocatalysis of
apoptosis was reduced. ROS levels were reduced in the present study
due to a reduction in oxidative stress in response to treatment
with C. longa extract. These findings indicated that
salsolinol successfully exerted cytotoxicity in SH-SY5Y cells
through the apoptotic pathway, and treatment with C. longa
extract was able to significantly reverse this toxicity.
References
|
1
|
Chattopadhyay I, Biswas K, Bandyopadhyay U
and Banerjee RK: Turmeric and curcumin: Biological actions and
medicinal applications. Curr Sci. 87:44–53. 2004.
|
|
2
|
Khalsa SVK: Turmeric. The Golden Healer.
http://www.healthy.net/recipe/health/turmeric_the_golden_healer/47Accessed.
2013.
|
|
3
|
Chaturvedi TP: Uses of turmeric in
dentistry: An update. Indian J Dent Res. 20:107–109. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Tayyem RF, Heath DD, Al-Delaimy WK and
Rock CL: Curcumin content of turmeric and curry powders. Nutr
Cancer. 55:126–131. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Aggarwal BB and Harikumar KB: Potential
therapeutic effects of curcumin, the anti-inflammatory agent,
against neurodegenerative, cardiovascular, pulmonary, metabolic,
autoimmune and neoplastic diseases. Int J Biochem Cell Biol.
41:40–59. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Jurenka JS: Anti-inflammatory properties
of curcumin, a major constituent of Curcuma longa: A review
of preclinical and clinical research. Altern Med Rev. 14:141–153.
2009.PubMed/NCBI
|
|
7
|
Sandur SK, Ichikawa H, Pandey MK,
Kunnumakkara AB, Sung B, Sethi G and Aggarwal BB: Role of
pro-oxidants and antioxidants in the anti-inflammatory and
apoptotic effects of curcumin (diferuloylmethane). Free Radic Biol
Med. 43:568–580. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kulkarni SK, Bhutani MK and Bishnoi M:
Antidepressant activity of curcumin: Involvement of serotonin and
dopamine system. Psychopharmacology (Berl). 201:435–442. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Bhutani MK, Bishnoi M and Kulkarni SK:
Anti-depressant like effect of curcumin and its combination with
piperine in unpredictable chronic stress-induced behavioral,
biochemical and neurochemical changes. Pharmacol Biochem Behav.
92:39–43. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Li YC, Wang FM, Pan Y, Qiang LQ, Cheng G,
Zhang WY and Kong LD: Antidepressant-like effects of curcumin on
serotonergic receptor-coupled AC-cAMP pathway in chronic
unpredictable mild stress of rats. Prog Neuropsychopharmacol Biol
Psychiatry. 33:435–449. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Hurley LL, Akinfiresoye L, Nwulia E,
Kamiya A, Kulkarni AA and Tizabi Y: Antidepressant-like effects of
curcumin in WKY rat model of depression is associated with an
increase in hippocampal BDNF. Behav Brain Res. 239:27–30. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ng TP, Chiam PC, Lee T, Chua HC, Lim L and
Kua EH: Curry consumption and cognitive function in the elderly. Am
J Epidemiol. 164:898–906. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Aggarwal BB, Sundaram C, Malani N and
Ichikawa H: Curcumin: The Indian solid gold. Adv Exp Med Biol.
595:1–75. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Tieu K: A guide to neurotoxic animal
models of Parkinson's disease. Cold Spring Harb Perspect Medicine.
1:a0093162011. View Article : Google Scholar
|
|
15
|
Mravec B: Salsolinol, a derivate of
dopamine, is a possible modulator of catecholaminergic
transmission: A review of recent developments. Physiol Res.
55:353–364. 2006.PubMed/NCBI
|
|
16
|
Antkiewicz-Michaluk L: Endogenous risk
factors in Parkinson's disease: Dopamine and
tetrahydroisoquinolines. Pol J Pharmacol. 54:567–572.
2002.PubMed/NCBI
|
|
17
|
Maruyama W, Dostert P, Matsubara K and
Naoi M: N-methyl(R)salsolinol produces hydroxyl radicals:
Involvement to neurotoxicity. Free Radic Biol Med. 19:67–75. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Moser A, Siebecker F, Vieregge P,
Jaskowski P and Kömpf D: Salsolinol, catecholamine metabolites, and
visual hallucinations in L-dopa treated patients with Parkinson's
disease. J Neural Transm Vienna. 103:421–432. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Alladi PA, Mahadevan A, Yasha TC, Raju TR,
Shankar SK and Muthane U: Absence of age-related changes in nigral
dopaminergic neurons of Asian Indians: Relevance to lower incidence
of Parkinson's disease. Neuroscience. 159:236–245. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Darvesh AS, Carroll RT, Bishayee A,
Novotny NA, Geldenhuys WJ and Van der Schyf CJ: Curcumin and
neurodegenerative diseases: A perspective. Expert Opin Investig
Drugs. 21:1123–1140. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ramlochansingh C, Taylor RE and Tizabi Y:
Toxic effects of low alcohol and nicotine combinations in SH-SY5Y
cells are apoptotically mediated. Neurotox Res. 20:263–269. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Brown D, Tamas A, Reglödi D and Tizabi Y:
PACAP protects against salsolinol-induced toxicity in dopaminergic
SH-SY5Y cells: Implication for Parkinson's disease. J Mol Neurosci.
50:600–607. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Muthuraman P, Gansukh E, Bhupendra M,
Murugesan C, Rafi N and Kim DH: Investigation of the role of
aspartame on apoptosis process in Hela cells. Saudi J Biol Sci.
June 15–2015.(Epub ahead of print).
doi:10.1016/j.sjbs.2015.06.001.
|
|
24
|
Muthuraman P, Gansukh E, Baskar V,
Bhupendra M, Rafi N, Bong YJ and Kim DH: Time and
concentration-dependent therapeutic potential of silver
nanoparticles in cervical carcinoma cells. Biol Trace Elem Res.
170:309–319. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kirkland RA, Windelborn JA, Kasprzak JM
and Franklin JL: A Bax-induced pro-oxidant state is critical for
cytochrome c release during programmed neuronal death. J Neurosci.
22:6480–6490. 2002.PubMed/NCBI
|
|
26
|
Pandurangan M, Veerappan M and Kim DH:
Cytotoxicity of zinc oxide nanoparticles on antioxidant enzyme
activities and mRNA expression in the cocultured C2C12 and 3T3-L1
cells. Appl Biochem Biotechnol. 175:1270–1280. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Pfaffl MW: A new mathematical model for
relative quantification in real-time RT-PCR. Nucleic Acids Res.
29:e452001. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Muthuraman P: Effect of cortisol on
caspases in the co-cultured C2C12 and 3 T3-L1 cells. Appl Biochem
Biotechnol. 173:980–988. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Frankfurt OS and Krishan A:
Apoptosis-based drug screening and detection of selective toxicity
to cancer cells. Anticancer Drugs. 14:555–561. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Das JR and Tizabi Y: Additive protective
effects of donepezil and nicotine against salsolinol-induced
cytotoxicity in SH-SY5Y cells. Neurotox Res. 16:194–204. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Song JX, Shaw PC, Wong NS, Sze CW, Yao XS,
Tang CW, Tong Y and Zhang YB: Chrysotoxine, a novel bibenzyl
compound selectively antagonizes MPP+, but not rotenone,
neurotoxicity in dopaminergic SH-SY5Y cells. Neurosci Lett.
521:76–81. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Storch A, Kaftan A, Burkhardt K and
Schwarz J: 1-Methyl-6,7-dihydroxy-1,2,3,4-tetrahydroisoquinoline
(salsolinol) is toxic to dopaminergic neuroblastoma SH-SY5Y cells
via impairment of cellular energy metabolism. Brain Res. 855:67–75.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhang Y, Wang L, Mu X, Duan J, Zhu Y, Qing
H, Li Y, Xiao S and Deng Y: Assessment of salsolinol N
methyltransferase activity in rat peripheral lymphocytes by liquid
chromatography-electrospray time-of-flight mass spectrometry. Anal
Bioanal Chem. 399:3541–3545. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Copeland RL Jr, Das JR, Kanaan YM, Taylor
RE and Tizabi Y: Antiapoptotic effects of nicotine in its
protection against salsolinol-induced cytotoxicity. Neurotox Res.
12:61–69. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Worth PF: How to treat Parkinson's disease
in 2013. Clin Med. 13:93–96. 2013. View Article : Google Scholar
|