Introduction
Allergic rhinitis (AR) is a chronic, reversible
allergic airway disease that has become a significant global public
health concern and >500 million people around the world are
estimated to currently be suffering with AR (1). Symptoms of AR include rhinorrhoea,
nasal obstruction, nasal itching and sneezing, and it is often
associated with ocular symptoms. Impairment of quality-of-life is
observed in the majority of patients. They may suffer from sleep
disorders and emotional problems, and experience impairments in
completing activities and proper social functioning.
AR is a Type I allergic disease caused by an
immunoglobulin E (IgE)-mediated adaptive immune response. IgE
production results from complex interactions between B cells, T
cells, mast cells and basophils, and involves the presence of
interleukin (IL)-4, IL-13 and IL-18 cytokines, as well as a
physical interaction between T and B-cells by a number of surface
and adhesion molecules (2). T-helper
2 (Th2) cells (3) and a
downregulation of T-regulatory cell responses (4–6) drive
the synthesis of IgE and the recruitment, maturation, survival and
effector function of accessory cells, such as eosinophils,
basophils and mast cells. The influx of eosinophils and Th2 cells,
producing IL-4, IL-5 and IL-13, is the main feature of AR (7).
Chinese traditional herbal medicines have long been
used to maintain the immune balance and to treat various allergic
diseases, such as allergic rhinitis (8–11),
asthma (12) and atopic dermatitis
(13). Biminne is the first herbal
preparation to be clinically tested for AR internationally. It has
anti-allergic and anti-inflammatory effects not only through its
ability to restrain inflammatory cells degranulation and
antagonistic inflammatory medium, but it can also reduce and remove
serum IgE levels (8). Herbal
formulas in traditional Chinese medicine referred to as
Yu-ping-feng-san (9),
Bu-zhong-yi-qi-tang (10) and
Xin-yi-san (11) are also considered
to be effective medicines for allergic rhinitis treatment. There is
little research focus on the isolation of herbal in allergic
rhinitis. Sinomenine (SN) was first isolated from Sinomenium
acutum in the 1920′s (14), and
since then a vast number of pharmacological and clinical studies
have been performed in China and Japan, demonstrating that the pure
alkaloid extract of SN possesses anti-inflammatory and
immune-regulatory properties (14,15).
Therefore, it is hypothesized that SN may be capable of
immune-modulatory effects on the allergic inflammation of the
airways. The aim of the present study was to evaluate whether SN
had an effect on inflammation of the nasal mucosa, as well as on
the immune response in an AR mouse model.
Materials and methods
Animals
A total of 40 male BALB/c mice (5-week-old; 16–18 g)
raised and maintained under specific pathogen-free conditions were
obtained from the Hubei Center for Disease Control and Prevention
(Wuhan, China). All mice were maintained under standard
conventional conditions: 12-h light/dark cycle, temperature
(18–22°C) and humidity (50–60%), with food and water ad
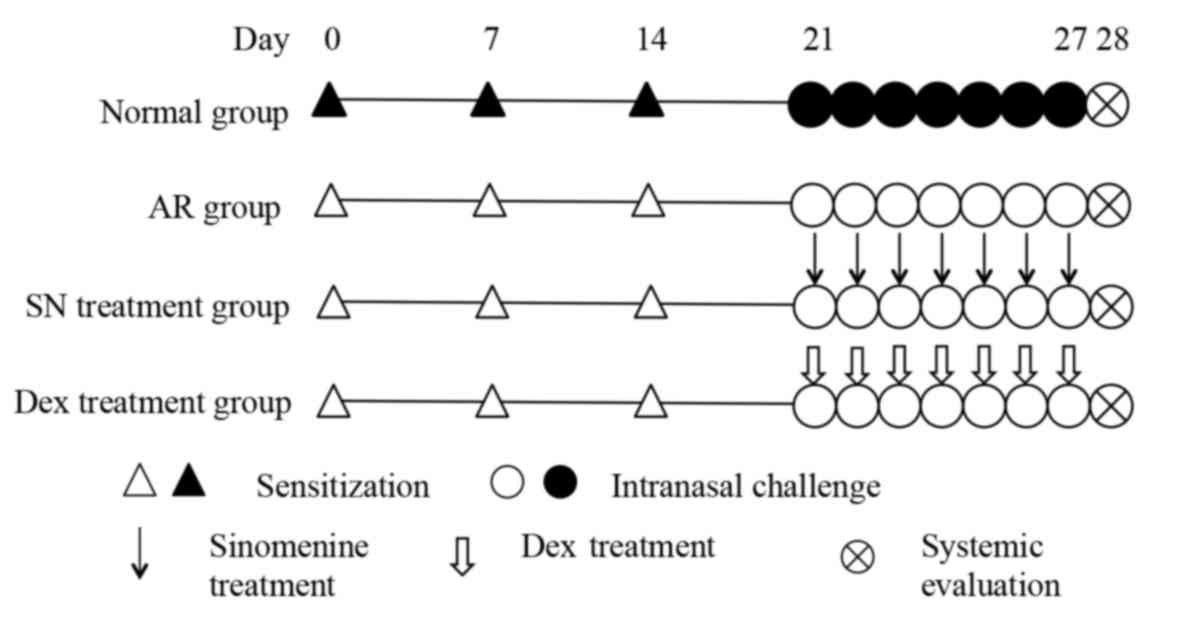
libitum. The animals were randomly divided into four groups:
Normal group, AR group, SN group and dexamethasone (Dex) group and
each group consisted of 10 mice. Corticosteroids such as
dexamethasone, the most potent therapeutic agents used for allergic
rhinitis, profoundly inhibit the activity of T cells largely
through the inhibition of expression of various cytokines (1,16), thus
we make it as a positive control. Mice in the AR, SN and Dex groups
were sensitized by intraperitoneal injection with 500 µl
phosphate-buffered saline (PBS) containing 10 µg ovalbumin (OVA;
grade V; Sigma-Aldrich; Merck Millipore, Darmstadt, Germany) and 1
mg aluminum hydroxide on days 0, 7 and 14. Mice in the AR, SN and
Dex groups were subjected to intranasal challenge with 20 µl PBS
containing 500 µg OVA for 7 days, between days 21 and 27 (17). Mice in the normal group were injected
with PBS alone, and PBS was administered intranasally following the
same schedule. Along with sensitization and challenge, selected
groups of mice were administered 100 mg/kg SN (purity, ≥98%) or 2
mg/kg Dex (purity, ≥98%; both Sigma-Aldrich; Merck Millipore),
which were dissolved in 200 µl PBS and orally administered daily 2
h before intranasal OVA challenge between days 21 and 27 (18). The experimental protocol is shown in
more detail in Fig. 1. The protocols
of the current study were approved by the Animal Ethics Committee
of Renmin Hospital of Wuhan University (Wuhan, China).
Measurement of nasal symptoms and
tissue preparation
At 20 min after the final OVA/PBS challenge on day
27, four observers blinded to the study groups recorded the
frequencies of nasal rubbing and sneezing in each group. After 24 h
from the last challenge with OVA/PBS, mice were anesthetized with
1–2 ml diethyl ether (60–29-7; Sinopharm Chemical Reagent Co.,
Ltd., Shanghai, China) volatilized in a 2 liter seal pot. Blood was
drawn blood from the eyeballs and mice were sacrificed quickly by
cervical dislocation. The nasal mucosa of mice was rapidly
collected once the mice were sacrificed using a small curette under
a microscope meticulously and was immediately immersed in liquid
nitrogen and stored until further use in reverse
transcription-quantitative polymerase chain reaction (RT-qPCR) and
western blot analyses.
Histopathological evaluation of nasal
cavity
Nasal tissues were removed 24 h after the last
challenge with OVA/PBS, fixed in 10% neutral buffered formalin and
decalcified. The coronal nasal section (5 µm) was stained with
hematoxylin and eosin, and the number of eosinophils was counted
under a microscope at four random high-power fields (HPFs) of the
submucosal region of the nasal cavity at ×400 magnification
(19), then the mean was taken as
the number of eosinophils in each group.
Enzyme-linked immunosorbent assay
(ELISA)
Mice serum was obtained immediately after sacrifice,
and stored at 4°C. Serum levels of anti-OVA specific IgE (N509;
R&D Systems, Inc., Minneapolis, MN, USA), interferon-γ (IFN-γ;
BMS6027), IL-4 (BMS613) and transforming growth factor-β (TGF-β;
BMS608/4; eBioscience, Inc., San Diego, CA, USA) in the animals
were measured using commercially available ELISA kits, according to
the manufacturer's recommendations. The concentrations of anti-OVA
specific IgE, IFN-γ, IL-4 and TGF-β were calculated from the
equations obtained from standard curve plots for the standard
solutions in the kits.
RT-qPCR
Total RNA was prepared from the nasal mucosa of the
mice using TRIzol reagent (N15596-026, Invitrogen; Thermo Fisher
Scientific, Inc., Waltham, MA, USA). Complementary DNA (cDNA) was
synthesized from 2 µg total RNA using Superscript Reverse
Transcriptase and oligo (dT) primers (K1629; Fermentas; Thermo
Fisher Scientific, Inc.). For the analysis of TGF-β and GAPDH
levels, specifically-designed primers and probes were purchased
from Invitrogen (Thermo Fisher Scientific, Inc.). The mRNA
expression of TGF-β and GAPDH was determined by qPCR, through
amplifying 25 ng cDNA in 50 µl 1X SYBR-Green PCR Master Mix
(DRR041A, Takara Biotechnology Co., Ltd., Dalian, China) containing
200 nM primers. qPCR primers were as follows: TGF-β forward,
5′-AGGGCTACCATGCCAACTTC-3′ and reverse, 5′-CCACGTAGTAGACGATGGGC-3′;
and GAPDH forward, 5′-ACCCAGAAGACTGTGGATGG-3′ and reverse,
5′-TGCTGTAGCCAAATTCGTTG-3′. Experiments were performed in
triplicate, using an ABI Prism 7500 Sequence Detection system
(Applied Biosystems; Thermo Fisher Scientific, Inc.). The average
transcript levels of genes were then normalized to GAPDH. Negative
controls (Master Mix containing untranscribed total RNA, or sample
without any cDNA or RNA) were used in each experiment. Relative
quantitation of TGF-β mRNA expression was calculated as the fold
increase in expression using the 2−ΔΔCq method (20) and the housekeeping gene was
GAPDH.
Western blot analysis
Proteins were obtained from the nasal mucosa of each
group using lysis buffer (containing 0.5% Triton X-100, 150 mM
NaCl, 15 mM Tris (pH 7.4) 1 mM CaCl2 and 1 mM
MgCl2). The protein concentrations were determined using
a BCA protein assay reagent (Thermo Fisher Scientific, Inc.).
Samples (20 µl; 2.7 µg/ml) were separated by 12% SDS-PAGE and
transferred onto polyvinylidene difluoride membranes (EMD
Millipore, Billerica, MA, USA). Next, the samples were
immunoblotted with primary antibodies against TGF-β (3711) and
GAPDH (5174; Cell Signaling Technology, Inc., Danvers, MA, USA) at
4°C overnight. Following washing with 1X TBST solution, membranes
were immunoblotted with secondary antibodies (7074; Cell Signaling
Technology, Inc.) at room temperature for 1 h. Proteins were
subsequently detected with an enhanced chemiluminescence reagent
(P0018; Beyotime Institute of Biotechnology, Shanghai, China) and
transferred to X-ray films. Finally, Quantity One software (version
4.6.2; Bio-Rad Laboratories, Inc., Hercules, CA, USA) was used to
quantify the results.
Statistical analysis
The results are presented as the mean ± standard
error of the mean. A Mann-Whitney U-test was used to compare
results between different groups. Statistical analyses were
performed by the SPSS statistical software (version 16.0; SPSS,
Inc., Chicago, IL, USA). P-values of <0.05 were considered as
indicating statistically significant associations.
Results
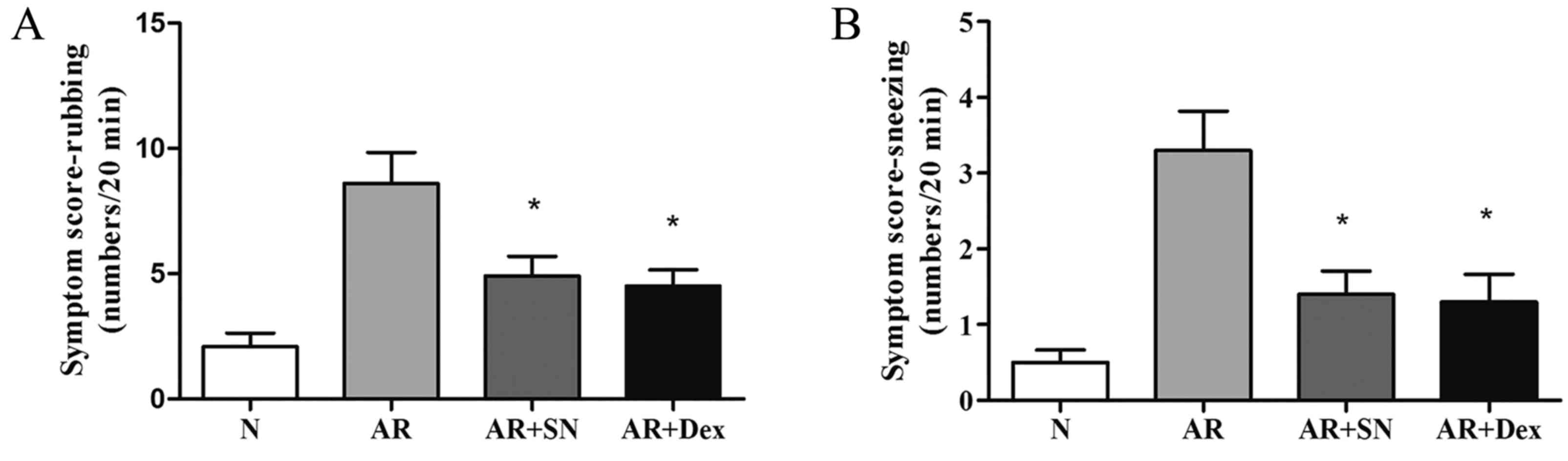
Symptom scoring
The nasal rubbing and sneezing symptom scores in the
AR group were significantly elevated when compared with those in
the normal group. The frequencies were 8.6±1.2 and 5.8±1.7 times/20
min in the AR group, respectively. By contrast, the scores were
significantly lower in the two treatment groups when compared with
the untreated AR group (Nasal rubbing: AR vs. AR + SN, P=0.0218; AR
vs. AR + Dex, P=0.0091; Sneezing: AR vs. AR + SN, P=0.0054; AR vs.
AR + Dex, P=0.0055; P<0.05), and the SN treatment group showed
no significant decrease in symptom incidence compared with the Dex
treatment group (Fig. 2).
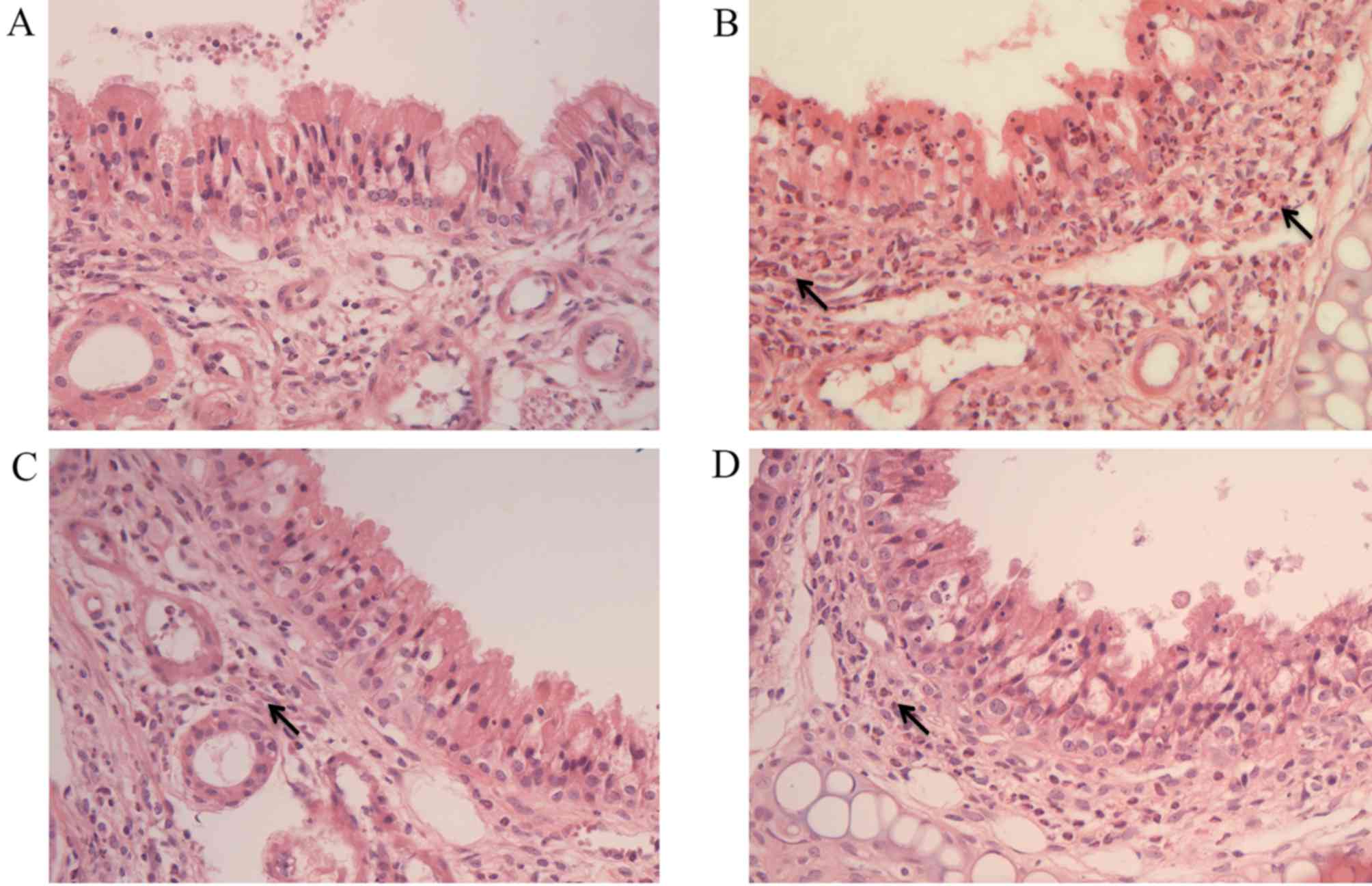
Histology of nasal mucosa and
eosinophil infiltration
Histological images of the nasal tissue of each
group are shown in Fig. 3. These
images demonstrated decreased eosinophil infiltration and decreased
epithelial layer disruption in the SN and Dex treatment groups when
compared with the AR group (Fig. 3).
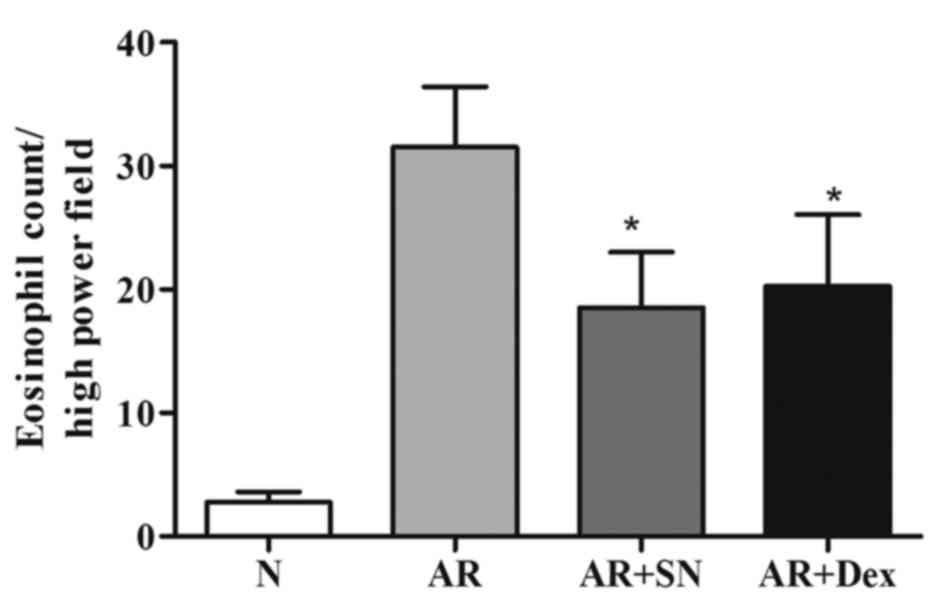
The eosinophil count per HPF in the AR group was 31.5±4.9, which
was significantly higher from that of the normal group (2.8±0.9
eosinophils/HPF; P<0.05). The eosinophil counts per HPF in the
SN treatment group and Dex treatment group were 18.5±4.5 and
20.3±5.7, respectively (Fig. 4);
thus, the number of eosinophils was significantly reduced in the
two treatment groups (P<0.05) when compared with the AR group.
These results indicate that the SN and Dex treatments decreased the
eosinophil migration in the nasal mucosa, but there was no
significant difference between them (P>0.05).
Serum levels of OVA-specific IgE and
cytokines
In order to evaluate the effects of the SN treatment
on AR, the serum levels of OVA-specific IgE and various cytokines
were detected by ELISA. The results indicated that OVA-specific IgE
levels significantly decreased in the two treatment groups when
compared with those in the AR group (P<0.05; Fig. 5). The SN treatment group demonstrated
a significant decrease (P<0.05) in the Th2 cytokine IL-4 level
in comparison with the level in the AR group, which was consistent
with the observed symptom scores and histological observations.
Similarly, the Th1 cytokine IFN-γ was also significantly reduced
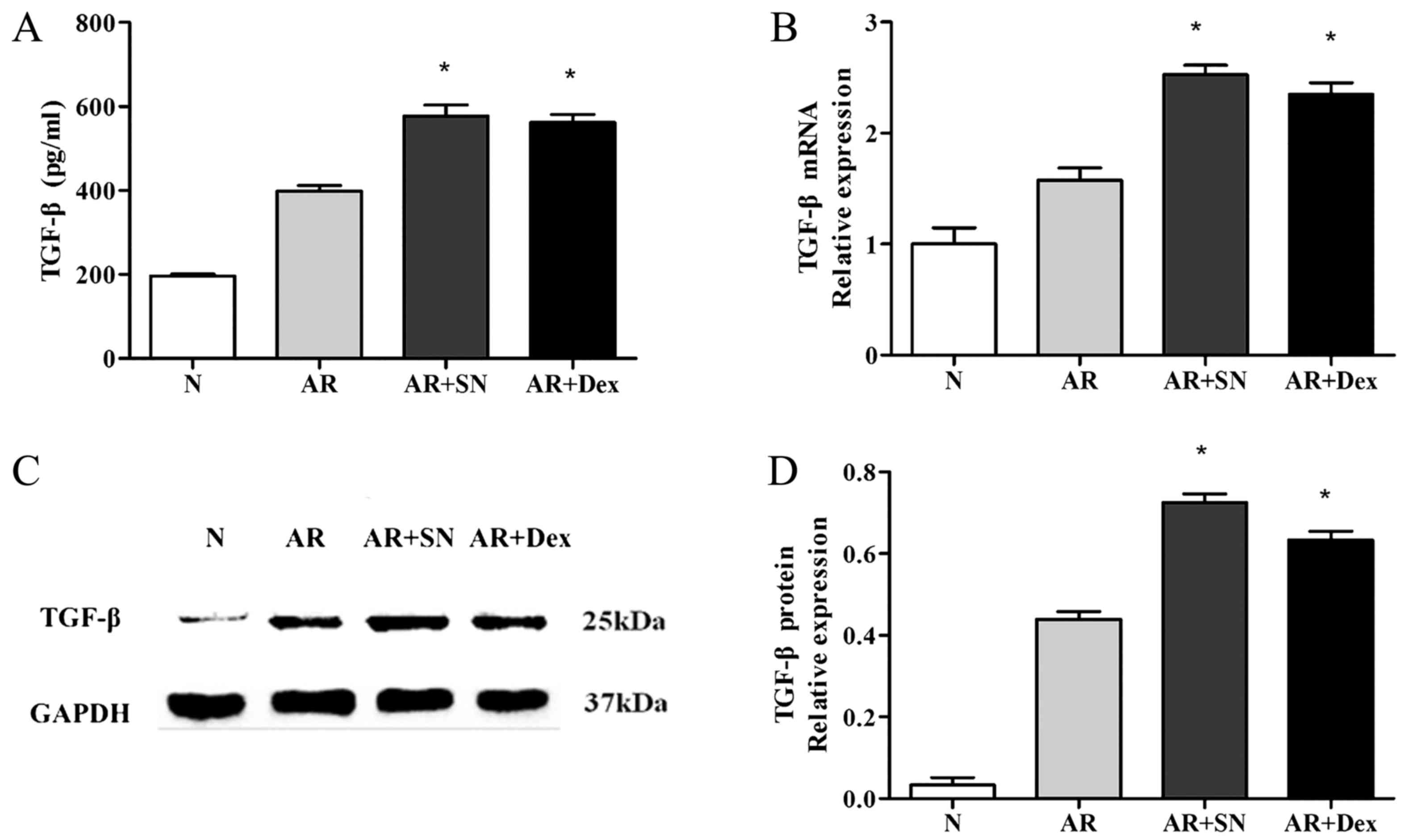
(P<0.05) in the two treatment groups (Fig. 5). Furthermore, treatment with SN
resulted in enhancement of the production of TGF-β in the mouse
serum (Fig. 6A).
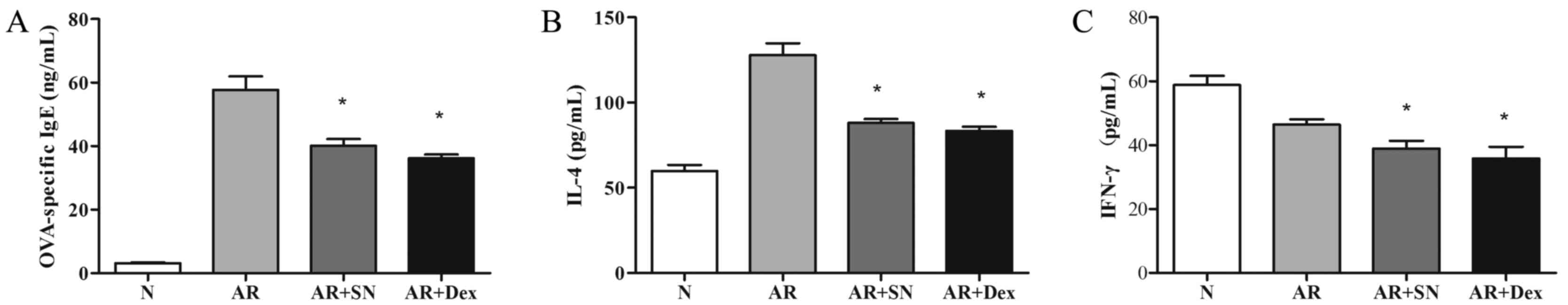 | Figure 5.Protein levels of (A) OVA-specific
IgE, (B) IL-4 and (C) IFN-γ in the various study groups, as
determined by ELISA. Systemic treatment with SN suppressed
OVA-specific IgE, while the levels of systemic Th2 cytokine IL-4
and Th1 cytokine IFN-γ decreased subsequent to SN administration.
*P<0.05 vs. the AR group. OVA, ovalbumin; IL-4, interleukin-4;
IFN, interferon; N, normal; AR, allergic rhinitis; SN, sinomenine;
Dex, dexamethasone. |
Expression of TGF-β in the nasal
mucosa
The level of TGF-β transcriptional activity was
evaluated in the nasal mucosa of each group by RT-qPCR (Fig. 6B). The mRNA expression levels of
TGF-β were significantly increased in the SN treatment group when
compared with the AR group (P<0.05). Furthermore, the protein
levels of TGF-β in the nasal mucosa were also significantly
elevated in the SN treatment group, as determined by western blot
analysis (Fig. 6C and D). The
expression of TGF-β in the Dex group exhibited almost the same
change when compared to the AR group.
Discussion
The majority of the clinical symptoms of AR,
including rhinorrhea, nasal itching, sneezing and nasal congestion,
cause significant discomfort to patients (21). AR is considered to result from an
IgE-mediated allergy associated with a nasal inflammation of
variable intensity (1). Cells,
mediators, cytokines, chemokines, neuropeptides, as well as
adhesion molecules and cells (2–7,22–25), are
all considered to cooperate in a complex network, provoking
specific symptoms and nonspecific nasal hyperreactivity. IgE
production is induced following complex interactions between
B-cells, T-cells, mast cells and basophils, and involves the
presence IL-4, IL-13 and IL-18 cytokines and a physical interaction
between T and B-cells by various surface and adhesion molecules
(2). Eosinophils numbers are
increased and activated in the nasal mucosa of patients with
symptomatic allergic (22). Various
mediators are released in nasal secretions, such as CysLT (23), ECP (24) and histamine (25). CD4+ lymphocytes with a Th2
phenotype serve an important role in the development of AR, and the
suppression of Th2 lymphocytes may be a potential therapeutic
target for the treatment of AR.
Various Chinese traditional medicine formulas have
been used for the treatment of allergic diseases for thousands of
years. The search for appropriate natural products may provide
further treatment options for allergic diseases to the currently
used drugs. In vitro studies have demonstrated that SN is
able to inhibit lymphocyte proliferation and antibody production by
B cells, as well as to potently reduce the production of
inflammatory factors by macrophages (26–28). SN
also inhibits the antigen-presenting capacity of bone
marrow-derived DCs with the decrease of IL-12, TNF-α and IL-1β
production (29). In addition, SN
affects the production of several allergic mediators, including
IL-6, PGD2, LTC4, β-Hex and COX-2 protein (30). In vitro experiments by Shu
et al (31). revealed that
the immunosuppressive activity elicited by SN in CD4+
primary lymphocytes was largely attributed to caspase-3-dependent
apoptosis These findings indicate that SN has the potential for use
in the treatment of allergies.
In the present study, AR mice treated with SN or Dex
had lower symptoms scores for nose rubbing and sneezing. Meanwhile,
the infiltration of eosinophils, the proliferation of goblet cells
and the loss of ciliated cells in the nasal epithelium are common
histopathological changes of the nasal mucosa in AR mice (32). Subsequent to treatment with SN or
Dex, the number of eosinophils was reduced when compared with that
in the nasal mucosa of untreated AR mice. Thus, it can be concluded
that SN treatment, as well as Dex treatment, decreased changes in
the nasal mucosa and alleviated the symptoms in AR mice.
OVA-specific IgE and IL-4 levels were found to
increase in AR mice compared with those in the normal group.
However, SN significantly inhibited the expression of OVA-specific
IgE and IL-4 in AR mice in the current study experiments.
Similarly, Feng et al (18)
observed that SN treatment suppressed the production of antibodies,
including anti-OVA IgG2a, IgG1 and IgE, as well as the secretion of
cytokines, such as IFN-γ and IL-5. In addition, this previous study
demonstrated that SN enhanced the secretion of TGF-β (18). SN also serves an important role in
the Th1/Th2 cell balance by regulating the expression levels of
T-bet and GATA-3, which are the transcription factors of Th1 and
Th2, respectively (33). All these
aforementioned results suggest that SN appears to have a
suppressive effect on AR. The regulatory T cells (Tregs) have been
shown to suppress Th1 and Th2 responses in vitro (34–36), and
TGF-β was considered to serve an important role in the development
and differentiation of Tregs (37–39). In
the present study, treatment with SN was followed by an enhancement
of TGF-β secretion. Thus, the downregulation of Th2 responses and
allergic symptom scores by SN treatment may be associated with the
role of this Treg cytokine in these immune responses.
In conclusion, the present study demonstrated that
SN had an inhibitory efficacy on AR, by alleviating the symptoms
and inhibiting the expression of OVA-specific IgE and Th2 cytokines
in OVA-induced AR mice. SN treatment also reduced the eosinophil
infiltration. These results may depend on the induction of the
local and systemic TGF-β expression, which is an important cytokine
in Treg cells, by SN treatment. Therefore, the results suggest that
SN may have a good potency in AR treatment.
Acknowledgements
The authors are grateful to the staff in the
laboratory of Renmin Hospital of Wuhan University (Wuhan, China)
who fully cooperated and provided important support to the current
project. The authors would specifically like to thank Dr Liu Ying
(College Of Nursing, The Second Military Medical University,
Shanghai, China) for correcting the manuscript.
References
|
1
|
Bousquet J, Khaltaev N, Cruz AA, Denburg
J, Fokkens WJ, Togias A, Zuberbier T, Baena-Cagnani CE, Canonica
GW, van Weel C, et al: Allergic rhinitis and its impact on asthma
(ARIA) 2008 update (in collaboration with the World Health
Organization, GA(2)LEN and AllerGen). Allergy. 63:(Suppl 86).
S8–S160. 2008. View Article : Google Scholar
|
|
2
|
Punnonen J, Aversa GG, Vandekerckhove B,
Roncarolo MG and de Vries JE: Induction of isotype switching and Ig
production by CD5+ and CD10+ human fetal B
cells. J Immunol. 148:3398–3404. 1992.PubMed/NCBI
|
|
3
|
Romagnani S: Immunologic influences on
allergy and the TH1/TH2 balance. J Allergy Clin Immunol.
113:395–400. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Romagnani S: Regulatory T cells: Which
role in the pathogenesis and treatment of allergic disorders?
Allergy. 61:3–14. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Allam JP and Novak N: Immunological
mechanisms of sublingual immunotherapy. Curr Opin Allergy Clin
Immunol. 14:564–569. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Akdis CA, Barlan IB, Bahceciler N and
Akdis M: Immunological mechanisms of sublingual immunotherapy.
Allergy. 61:(Suppl 81). S11–S14. 2006. View Article : Google Scholar
|
|
7
|
Wilson MS, Taylor MD, Balic A, Finney CA,
Lamb JR and Maizels RM: Suppression of allergic airway inflammation
by helminth-induced regulatory T cells. J Exp Med. 202:1199–1212.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hu G, Walls RS, Bass D, Ramon B, Grayson
D, Jones M and Gebski V: The Chinese herbal formulation biminne in
management of perennial allergic rhinitis: A randomized,
double-blind, placebo-controlled, 12-week clinical trial. Ann
Allergy Asthma Immunol. 88:478–487. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Makino T: Pharmacological properties of
Gyokuheifusan, a traditional Kampo medicinal formula. Yakugaku
Zasshi. 125:349–354. 2005.(In Japanese). View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Yang SH and Yu CL: Antiinflammatory
effects of Bu-zhong-yi-qi-tang in patients with perennial allergic
rhinitis. J Ethnopharmacol. 115:104–109. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Yang SH, Yu CL, Chen YL, Chiao SL and Chen
ML: Traditional Chinese medicine, Xin-yi-san, reduces nasal
symptoms of patients with perennial allergic rhinitis by its
diverse immunomodulatory effects. Int Immunopharmacol. 10:951–958.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wei Y, Lyu Y, Li M, Luo Q, Sun J, Liu F,
Lin Y, Chen M, Nurahmat M, Abduwaki M and Dong J: Comparison of
effect of granules and herbs of Bu-Shen-Yi-Qi-Tang on airway
inflammation in asthmatic mice. Chin Med J (Engl). 127:3957–3962.
2014.PubMed/NCBI
|
|
13
|
Chen HY, Lin YH, Huang JW and Chen YC:
Chinese herbal medicine network and core treatments for allergic
skin diseases: Implications from a nationwide database. J
Ethnopharmacol. 168:260–267. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Yamasaki H: Pharmacology of sinomenine, an
anti-rheumatic alkaloid from Sinomenium acutum. Acta medica
Okayama. 30:1–20. 1976.PubMed/NCBI
|
|
15
|
Lodge D, Headley PM, Duggan AW and Biscoe
TJ: The effects of morphine, etorphine and sinomenine on the
chemical sensitivity and synaptic responses of Renshaw cells and
other spinal neurones in the rat. Eur J Pharmacol. 26:277–284.
1974. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Lee SS, Won TB, Kim JW, Rhee CS, Lee CH,
Hong SC and Min YG: Effects of dexamethasone on the expression of
transforming growth factor-beta in the mouse model of allergic
rhinitis. Laryngoscope. 117:1323–1328. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Mo JH, Kang EK, Quan SH, Rhee CS, Lee CH
and Kim DY: Anti-tumor necrosis factor-alpha treatment reduces
allergic responses in an allergic rhinitis mouse model. Allergy.
66:279–286. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Feng H, Yamaki K, Takano H, Inoue K,
Yanagisawa R and Yoshino S: Suppression of Th1 and Th2 immune
responses in mice by Sinomenine, an alkaloid extracted from the
chinese medicinal plant Sinomenium acutum. Planta Med.
72:1383–1388. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Okano M, Nishizaki K, Abe M, Wang MM,
Yoshino T, Satoskar AR, Masuda Y and Harn DA Jr: Strain-dependent
induction of allergic rhinitis without adjuvant in mice. Allergy.
54:593–601. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Stokes JR, Romero FA Jr, Allan RJ,
Phillips PG, Hackman F, Misfeldt J and Casale TB: The effects of an
H3 receptor antagonist (PF-03654746) with fexofenadine on reducing
allergic rhinitis symptoms. J Allergy Clin Immunol. 129:409–412,
412.e1-2. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Eliashar R and Levi-Schaffer F: The role
of the eosinophil in nasal diseases. Curr Opin Otolaryngol Head
Neck Surg. 13:171–175. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Skoner DP, Lee L, Doyle WJ, Boehm S and
Fireman P: Nasal physiology and inflammatory mediators during
natural pollen exposure. Ann Allergy. 65:206–210. 1990.PubMed/NCBI
|
|
24
|
Rasp G, Thomas PA and Bujía J: Eosinophil
inflammation of the nasal mucosa in allergic and non-allergic
rhinitis measured by eosinophil cationic protein levels in native
nasal fluid and serum. Clin Exp Allergy. 24:1151–1156. 1994.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wilson SJ, Lau L and Howarth PH:
Inflammatory mediators in naturally occurring rhinitis. Clin Exp
Allergy. 28:220–227. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Liu L, Resch K and Kaever V: Inhibition of
lymphocyte proliferation by the anti-arthritic drug sinomenine. Int
J Immunopharmacol. 16:685–691. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Hojo H, Kondo Y, Umeda H, Tahira T and
Hashimoto Y: Effect of sinomenine on antibody responses in mice. J
Immunopharmacol. 7:33–42. 1985. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kim HM, Moon PD, Chae HJ, Kim HR, Chung
JG, Kim JJ and Lee EJ: The stem of Sinomenium acutum
inhibits mast cell-mediated anaphylactic reactions and tumor
necrosis factor-alpha production from rat peritoneal mast cells. J
Ethnopharmacol. 70:135–141. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Wang Y, Ma D, Jie Y, Wu Y and Pan Z:
Sinomenine can prolong high-risk corneal graft survival in a rat
model. Immunotherapy. 4:581–586. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Oh YC, Kang OH, Choi JG, Brice OO, Lee YS,
Keum JH, Kim SB, Shin DW, Ma J, Jeong GH and Kwon DY: Anti-allergic
effects of sinomenine by inhibition of prostaglandin D2
and leukotriene C4 in mouse bone marrow-derived mast
cells. Immunopharmacol Immunotoxicol. 33:266–270. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Shu L, Yin W, Zhang J, Tang B, Kang YX,
Ding F and Hua ZC: Sinomenine inhibits primary CD4+
T-cell proliferation via apoptosis. Cell Biol Int. 31:784–789.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Miyata M, Hatsushika K, Ando T, Shimokawa
N, Ohnuma Y, Katoh R, Suto H, Ogawa H, Masuyama K and Nakao A: Mast
cell regulation of epithelial TSLP expression plays an important
role in the development of allergic rhinitis. Eur J Immunol.
38:1487–1492. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Cheng Y, Zhang J, Hou W, Wang D, Li F,
Zhang Y and Yuan F: Immunoregulatory effects of sinomenine on the
T-bet/GATA-3 ratio and Th1/Th2 cytokine balance in the treatment of
mesangial proliferative nephritis. Int Immunopharmacol. 9:894–899.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Xu G, Mou Z, Jiang H, Cheng L, Shi J, Xu
R, Oh Y and Li H: A possible role of
CD4+CD25+ T cells as well as transcription
factor Foxp3 in the dysregulation of allergic rhinitis.
Laryngoscope. 117:876–880. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Lee SM, Gao B, Dahl M, Calhoun K and Fang
D: Decreased FoxP3 gene expression in the nasal secretions from
patients with allergic rhinitis. Otolaryngol Head Neck Surg.
140:197–201. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Malmhäll C, Bossios A, Pullerits T and
Lötvall J: Effects of pollen and nasal glucocorticoid on
FOXP3+, GATA-3+ and T-bet+ cells
in allergic rhinitis. Allergy. 62:1007–1013. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Dardalhon V, Awasthi A, Kwon H, Galileos
G, Gao W, Sobel RA, Mitsdoerffer M, Strom TB, Elyaman W, Ho IC, et
al: IL-4 inhibits TGF-beta-induced Foxp3+ T cells and,
together with TGF-beta, generates IL-9+
IL-10+ Foxp3(−) effector T cells. Nat Immunol.
9:1347–1355. 2008. View
Article : Google Scholar : PubMed/NCBI
|
|
38
|
Dardalhon V, Korn T, Kuchroo VK and
Anderson AC: Role of Th1 and Th17 cells in organ-specific
autoimmunity. J Autoimmun. 31:252–256. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Taylor A, Verhagen J, Blaser K, Akdis M
and Akdis CA: Mechanisms of immune suppression by interleukin-10
and transforming growth factor-beta: The role of T regulatory
cells. Immunology. 117:433–442. 2006. View Article : Google Scholar : PubMed/NCBI
|