Introduction
Nonsyndromic cleft lip with or without cleft palate
(NSCLP) is one of the most common human craniofacial malformations.
The incidence is 1.82/1,000 live births in China, and the worldwide
overall incidence of oral clefts is estimated to be 1.43/1,000,
with a wide variability among ethnic groups and regions (1). NSCLP is a complex disorder that does
not follow normal Mendelian patterns of inheritance; rather, NSCLP
exhibits etiological heterogeneity wherein genetic and
environmental factors may predispose patients to orofacial clefts
(2–4). In addition, gene-gene and/or
gene-environment interactions may contribute to clefting (5–7). The
etiopathogenesis of clefts has been widely studied but remains
poorly understood.
Mature microRNAs (miRNAs), which are small,
naturally occurring non-coding RNAs, inhibit messenger RNA and act
as negative regulators of gene expression. miRNAs are crucial for
the proper spatiotemporal development of various tissues and organs
(8–11). The roles of miRNAs in cellular
proliferation and differentiation as well as embryonic development
are gradually becoming clear (12).
In addition, there is evidence that miRNAs may be required for
craniofacial development (13), and
Eberhart et al (14)
demonstrated that the microRNA Mirn140 negatively regulates
platelet-derived growth factor (PDGF) signaling during palatal
development, and provided a mechanism for how the disruption of
PDGF signaling causes palatal clefting in zebrafish.
Despite the acknowledged function of miRNAs in
development and their importance in disease, as well as the fact
that miRNAs are a unique means of modulating signaling pathways
such as Wnt and transforming growth factor-β (15–18),
little is known with regard to the involvement of miRNAs in
regulating NSCLP. Research on the above issue is still in the early
stages, at which the problem of selecting appropriate samples and
screening miRNAs in human tissues require to be solved. It
represents a novel approach to elucidating the pathogenesis of
NSCLP with regard to miRNAs. Certain general principles require
establishment as follows: i) Tissue was selected over blood, as
tissue provides RNA of higher quality and quantity. This is likely
to reduce the stress and potential side effects associated with
invasive sample collection and thus, greatly facilitate participant
recruitment for the study. ii) Umbilical samples were used as
control tissue. NSCLP is a congenital deformity caused by abnormal
facial development during gestation and forms in embryos at 6–8
gestational weeks (19). The
umbilical cord develops from and contains remnants of the yolk sac
and allantois (and is therefore derived from the zygote), which is
involved in the whole embryonic process. Healthy umbilical cords
were used in the present study. This not only takes into
consideration the fact that NSCLP tissues collected were more than
one year apart from the timing when clefts were forming in
utero, but also allows the miRNA to be synchronized with
spatiotemporal characteristics in development. iii) Patients aged
3–24 months were selected as these patients can endure plastic
surgery according to Chinese standards (20).
The expression of 1,800 unique human miRNAs was
profiled in four NSCLP and 4 normal umbilical cord samples using
locked nucleic acid-based oligonucleotide microarrays and several
miRNAs that may be associated with the Wnt signal pathway were
screened.
Materials and methods
Specimen collection
The NSCLP samples used in the present study were
obtained from four patients with NSCLP from the Department of Oral
and Maxillofacial Surgery (Hospital of Stomatology, the First
Affiliated Hospital, Harbin Medical University, Harbin, China) and
were tissues that were not re-used in plastic surgery. The 4
umbilical cord tissues (used as the control) were obtained from the
Department of Obstetrics and Gynecology at the First Affiliated
Hospital of Harbin Medical University from July 2012 to August
2012. Control umbilical cord tissues were donated by healthy
gravidae without systemic disease, clefts or family history
thereof. The samples were flash-frozen in liquid nitrogen and
stored at −80°C until RNA extraction. Of these samples, 8 were used
for miRNA microarray analysis and 8 were used for reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
analysis. The NSCLP cases were classified as bilateral NSCLP,
unilateral NSCLP, right unilateral NSCLP and left unilateral NSCLP.
The clinical data of the patients are shown in Table I. Written consent for tissue donation
(for research purposes) was obtained from the patients or their
parents prior to tissue collection and the protocol was approved by
the Institutional Review Board of the First Affiliated Hospital of
Harbin Medical University (Harbin, China).
 | Table I.Clinical background of the 4 Han
Chinese NSCLP patients. |
Table I.
Clinical background of the 4 Han
Chinese NSCLP patients.
| Gender | Age (months) | Classification | Tissue type |
|---|
| Female | 3 | Right unilateral
NSCLP | Lip mucosa |
| Male | 3 | Left unilateral
NSCLP | Lip mucosa |
| Female | 24 | Bilateral
NSCLP | Palatine
mucosa |
| Male | 18 | Left unilateral
NSCLP | Palatine
mucosa |
miRNA microarray
The 7th generation miRCURY™ LNA Array (v.18.0;
Exiqon, Inc, Woburn, MA, USA) contains 3,100 capture probes,
covering all the human miRNAs annotated in miRBase 18.0 as well as
all viral miRNAs associated with these. In addition, this array
contains capture probes for 25 miRPlus™ human miRNAs.
RNA extraction
Total RNA was isolated using TRIzol (Invitrogen;
Thermo Fisher Scientific, Inc., Waltham, MA, USA) and the miRNeasy
Mini kit (Qiagen, Hilden, Germany) according to the manufacturers'
instructions, efficiently recovering all RNA species including
miRNA. RNA quality and quantity was measured using a NanoDrop
spectrophotometer (ND-1000; Thermo Fisher Scientific, Inc.) and RNA
integrity was determined by gel electrophoresis.
RNA labeling
After RNA isolation from the samples, the miRCURY™
Hy3™/Hy5™ Power labeling kit (Exiqon, Inc.) was used according to
the manufacturer's instructions for miRNA labeling. One microgram
of each sample was 3′-end-labeled with the Hy3™ fluorescent label,
using T4 RNA ligase and the following procedure: RNA in 2.0 µl
water was combined with 1.0 µl CIP buffer and CIP (Exiqon). The
mixture was incubated for 30 min at 37°C and the reaction was
terminated by incubating at 95°C for 5 min. Subsequently, 3.0 µl
labeling buffer, 1.5 µl fluorescent label (Hy3™), 2.0 µl dimethyl
sulfoxide and 2.0 µl labeling enzyme were added to the mixture. The
labeling reaction was then incubated for 1 h at 16°C and terminated
by incubating at 65°C for 15 min.
Array hybridization
After stopping the labeling procedure, the
Hy3™-labeled samples were hybridized to the miRCURY™ LNA Array
(v.18.0; Exiqon, Inc.) according to the instructions from the array
manual. The total 25-µl mixture containing the Hy3™-labeled samples
was added to 25 µl hybridization buffer, denatured for 2 min at
95°C and then incubated on ice for 2 min. Subsequently, the mixture
was hybridized to the microarray for 16–20 h at 56°C in a 12-Bay
Hybridization System (Nimblegen Systems Inc., Madison, WI, USA),
which provided an active mixing action and constant incubation
temperature to improve hybridization uniformity and enhance the
signal. Following hybridization, the slides were washed several
times using a Wash Buffer kit (Exiqon, Inc.) and dried by
centrifugation for 5 min at 127.44 × g. The slides were then
scanned using an Axon GenePix 4000B microarray scanner (Axon;
Molecular Devices, LLC, Sunnyvale, CA, USA).
miRNA target prediction
MicroCosm Targets Version5 (http://www.ebi.ac.uk/enright-srv/microcosm/htdocs/targets/v5/),
formerly miRBase Targets, Miranda (http://www.microrna.org/microrna/home.do), which
contains the target information for the hg19, mm9 and RN34 miRNAs,
and TargetScan version 6.2 (http://www.targetscan.org/vert_60/), which provides
information about human and mouse miRNA, were used to analyze
potential target genes for the deregulated miRNAs.
RT-qPCR
To confirm the results obtained by miRNA profiling,
the expression of selected miRNAs was measured using RT-qPCR
according to standard protocols on an Applied Biosystems 7700 HT
Sequence Detection System (Thermo Fisher Scientific, Inc.). In
brief, complementary (c)DNA was synthesized from total RNA using
gene-specific primers (Table II).
Reverse transcriptase reactions contained 800 ng RNA, 1 µmol/l
stem-loop RT primer, 10X RT buffer, 2.5 mmol/l of each of the
dNTPs, 200 U/µl MultiScribe reverse transcriptase and 40 U/µl RNase
inhibitor. The 20-µl reactions were incubated for 30 min at 16°C,
40 min at 42°C, and 5 min at 85°C and then were held at 4°C. The
10-µl PCR reaction mixture included 2 µl RT product, 2 µl 2X SYBR
Green PCR master mix (Thermo Fisher Scientific, Inc.) and 1 µl
primers (Table II). Reactions were
incubated in a 384-well optical plate at 95°C for 10 min, followed
by 40 cycles of 95°C for 10 sec and 60°C for 1 min. U6 small
nuclear RNA was used as an internal control to normalize RNA input.
The fold-change was calculated using the 2−ΔΔCq method
(21), where Cq is defined as the
fractional cycle number at which the fluorescence passes the fixed
threshold, and presented as the fold-expression change in NSCLP
tissues relative to their corresponding normal tissues after
normalization to the endogenous control. All experiments were
performed in triplicate.
| Table II.Oligonucleotide primers used in the
present study. |
Table II.
Oligonucleotide primers used in the
present study.
| Primer set
name |
Reverse-transcriptase reaction primer
(5′-3′) | PCR primer
(5′-3′) |
|---|
| U6 |
CGCTTCACGAATTTGCGTGTCAT | F:
GCTTCGGCAGCACATATACTAAAAT |
|
|
| R:
CGCTTCACGAATTTGCGTGTCAT |
| hsa-miR-1260b |
GTCGTATCCAGTGCGTGTCGTGGAGTC | GSP:
GGAATCCCACCACTGC |
|
|
GGCAATTGCACTGGATACGACATGGTG | R:
TGCGTGTCGTGGAGTC |
| hsa-miR-24-3p |
GTCGTATCCAGTGCGTGTCGTGGAGTC | GSP:
GGGTGGCTCAGTTCAGC |
|
|
GGCAATTGCACTGGATACGACCTGTTC | R:
CAGTGCGTGTCGTGGAG |
| hsa-miR-27b-3p |
GTCGTATCCAGTGCGTGTCGTGGAGTC | GSP:
GGGGGTTCACAGTGGCTAAG |
|
|
GGCAATTGCACTGGATACGACGCAGAA | R:
GTGCGTGTCGTGGAGTCG |
| hsa-miR-205-5p |
GTCGTATCCAGTGCGTGTCGTGGAGTCG | GSP:
CGTCCTTCATTCCACCG |
|
|
GCAATTGCACTGGATACGACCAGACT | R:
CAGTGCGTGTCGTGGAGT |
Data analysis
Scanned images were imported into the GenePix Pro
6.0 software (Axon; Molecular Devices, LLC) for grid alignment and
data extraction. Replicated miRNAs were averaged, and miRNAs with
intensities ≥30 in all samples were chosen for use in calculating
the normalization factor. Median normalization was used to
normalize the expression data. After normalization, miRNAs with
significant differences in expression were identified using volcano
plot filtering. Hierarchical clustering was performed using MEV
software (v4.6, TIGR; The J. Craig Venter Institute, La Jolla, CA,
USA).
Group data are expressed as the mean ± standard
error of the mean. Statistical comparisons (performed using
analysis of variance followed by Dunnett's post hoc test) were
performed using Microsoft Excel 2007 (Microsoft Corp., Redmond, WA,
USA). A two-tailed P<0.05 was considered to indicate a
statistically significant difference.
Results
RNA quantity and quality
The quantity and quality of the RNA samples in each
group were checked by denaturing agarose gel electrophoresis and
absorbance at 260 nm (A260)/280 and A260/230 ratios (Fig. 1). The A260/280 ratio, A260/230 ratio
and gel electrophoresis results confirmed that the quality of the
RNA isolated was good.
Differential expression of miRNAs in
NSCLP and control tissues
After normalization of the raw data, 254 miRNAs were
found to be differentially expressed. Compared to the control, 181
miRNAs were found to be upregulated and 73 downregulated in the
NCLP samples. Of these, based on fold-changes and P-values, 10
miRNAs were selected for further analysis, including 7 miRNAs that
were upregulated (hsa-miR-24-3p, hsa-miR-27b-3p, hsa-miR-720,
hsa-miR-205-5p, hsa-miR-1260b, hsa-miR-3676-5p and hsa-miR-5701)
and 3 miRNAs that were downregulated (hsa-miR-564, hsa-miR-4799-3p
and hsa-miR-4703-3p) (Table
III).
| Table III.miRNAs selected for bioinformatics
analysis. |
Table III.
miRNAs selected for bioinformatics
analysis.
| Name | Fold-change (exp
vs. con) | P-value (exp vs.
con) |
|---|
| hsa-miR-24-3p | 12.54569 | 0.02068 |
| hsa-miR-27b-3p | 12.24550 | 0.040829 |
| hsa-miR-720 | 10.49021 | 0.000145 |
| hsa-miR-205-5p | 12.08784 | 0.010106 |
| hsa-miR-1260b | 12.25486 | 0.011225 |
|
hsa-miR-3676-5p | 15.98345 | 0.006023 |
| hsa-miR-5701 | 23.22485 | 0.049594 |
| hsa-miR-564 | 0.098898 | 0.036984 |
|
hsa-miR-4799-3p | 0.099580 | 0.002004 |
| Down 3 |
|
|
|
hsa-miR-4703-3p | 0.080486 | 0.003453 |
miRNA target prediction and RT-qPCR
validation of microarray results
MicroCosm, Miranda and TargetScan were used to
identify potential target genes for the dysregulated miRNAs. The
results revealed that hsa-miR-1260b, hsa-miR-205-5p, hsa-miR-24-3p,
hsa-miR-27b-3p and hsa-miR720 may be associated with Wnt signaling
genes, including WNT10B, WNT5A, WNT5B, WNT10A, WNT9B, WNT4, WNT8B,
WNT2B, WNT3A, WNT3 (NSF), AXIN2, glycogen synthase kinase 3β
(GSK3B), golgi SNAP receptor complex member 2, secreted frizzled
related protein 1 (SFRP1), dickkopf WNT signaling pathway inhibitor
2 (DKK2), low-density lipoprotein receptor-related protein (LRP)5,
LRP6 and Frizzled class receptor 7 (Table IV). The dysregulation of
hsa-miR-1260b, hsa-miR-205-5p, hsa-miR-24-3p and hsa-miR-27b-3p,
for which predicted target information was supported by information
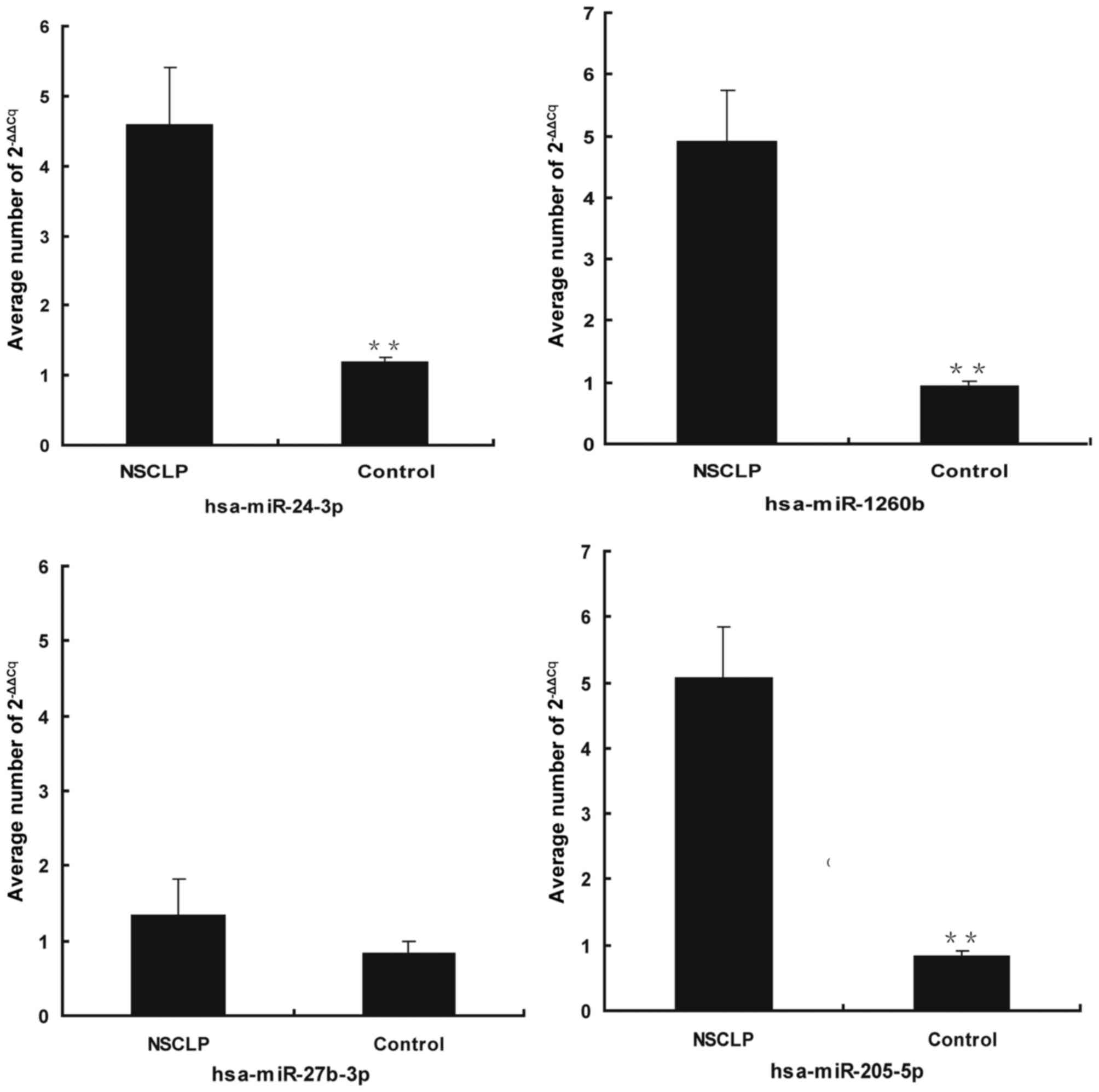
from at least two or more databases, was confirmed by RT-qPCR. The
results of the RT-qPCR analysis were consistent with the microarray
data for three of the four miRNAs examined and showed that,
although expression of hsa-miR-27b-3p was not significantly
different between NSCLP and control (P=0.09), hsa-miR-1260b
(P=0.001), hsa-miR-205-5p (P=0.001) and hsa-miR-24-3p (P=0.003)
were significantly upregulated in NSCLP tissues (Fig. 2).
| Table IV.Wnts associated with upregulated
miRNAs. |
Table IV.
Wnts associated with upregulated
miRNAs.
| miRNA | Gene symbol | Miranda | MicroCosm | TargetScan | Num statistics |
|---|
| hsa-miR-1260b | WNT10B | 1 | 0 | 1 | 2 |
| hsa-miR-1260b | WNT5A | 1 | 0 | 0 | 1 |
| hsa-miR-1260b | WNT5B | 1 | 0 | 0 | 1 |
| hsa-miR-205-5p | WNT10A | 1 | 0 | 0 | 1 |
| hsa-miR-205-5p | WNT5A | 1 | 1 | 0 | 2 |
| hsa-miR-205-5p | WNT5B | 0 | 1 | 0 | 1 |
| hsa-miR-205-5p | WNT9B | 0 | 1 | 0 | 1 |
| hsa-miR-205-5p | WNT3
(NSF) | 1 | 0 | 1 | 2 |
| hsa-miR-205-5p | AXIN2 | 1 | 0 | 1 | 2 |
| hsa-miR-205-5p | SMAD4 | 1 | 0 | 1 | 2 |
| hsa-miR-24-3p | WNT4 | 0 | 0 | 1 | 1 |
| hsa-miR-24-3p | WNT8B | 1 | 1 | 0 | 2 |
| hsa-miR-24-3p | GSK3B | 1 | 0 | 1 | 2 |
| hsa-miR-27b-3p | WNT2B | 1 | 0 | 0 | 1 |
| hsa-miR-27b-3p | WNT3A | 0 | 1 | 1 | 2 |
| hsa-miR-27b-3p | GOSR2 | 1 | 0 | 1 | 2 |
| hsa-miR-27b-3p | SFRP1 | 1 | 0 | 1 | 2 |
| hsa-miR-27b-3p | DKK2 | 1 | 0 | 1 | 2 |
| hsa-miR-27b-3p | LRP5 | 1 | 1 | 0 | 2 |
| hsa-miR-27b-3p | LRP6 | 1 | 0 | 1 | 2 |
| hsa-miR-27b-3p | FZD7 | 1 | 0 | 1 | 2 |
| hsa-miR720 | WNT5B | 1 | 0 | 0 | 1 |
Discussion
The present study detected 254 miRNAs that were
differentially expressed in NSCLP tissues as compared to those in
normal umbilical cord tissues. The NSCLP miRNA signature revealed
an altered biological phenotype that is similar to the miRNA
profiles of various growth-associated diseases: It was
characterized by significant upregulation of miRNAs (181/254).
Based on fold-change and P-values, 10 of these miRNAs, including 8
hsa-miRNAs that were upregulated and 2 hsa-miRNAs that were
downregulated, were selected for further analysis. Of note,
hsa-miR-27b-3p, hsa-miR-720, hsa-miR-1260b, hsa-miR-24-3p and
hsa-miR-205-5p were found to be highly expressed in tissues from
NSCLP patients. Furthermore, bioinformatics analysis showed that
various Wnt genes such as WNT2B and WNT10B were
associated with these hsa-miRNAs that we screened. Accordingly,
various miRNAs in control and NSCLP tissues were identified by
RT-qPCR, including hsa-miR-720, hsa-miR-1260b, hsa-miR-24-3p and
hsa-miR-205-5p.
A growing body of evidence showed that the Wnt
family of genes and their associated signaling pathways have
critical roles in various processes of growth and development,
including embryonic induction, epithelial and mesenchymal cellular
polarity, cell fate determination, cytoskeletal organization and
cell proliferation (22–24). Wnt expression is observed in the
upper lip and primary and secondary palates, and Wnts are involved
in regional specification of the vertebrate face.
WNT4, targeted by hsa-miR-24-3p (25), is a member of the canonical Wnt
signaling pathway and is extensively expressed in palatal
epithelium. He et al (26)
found that WNT4 is expressed in the palatal epithelium in
the anterior as well as posterior palate in a microarray survey of
gene expression profile in E13.5 mouse palatal shelves. Yu et
al (27) demonstrated miR-205
suppresses the expression of GSK-3β, the protein encoded by
GSK3B, through binding to its 3′-untranslated region. It has
been shown that GSK-3β is a direct target of miR-205 in 3T3-L1
cells and suppression of the expression of GSK-3β by miR-205 led to
activation of the Wnt pathway. GSK3B is part of the Wnt
signaling pathway and has a major role in epithelial cell
homeostasis (28). A role for
GSK3B in craniofacial defects has also been demonstrated,
since homozygous null mice display incomplete fusion of the ribs at
the midline and bifid sternum, delayed sternal ossification and
cleft palate (29).
However, no direct evidence has illustrated how
miR-1260b regulates Wnt genes and affects lip fusion and palatal
fusion. Hirata et al (30)
found that miRNA-1260b can silence SFRP1, DKK2 and
SMAD4 genes, which affects cancer cell proliferation and
invasion as well as the percentage of apoptotic cells. Iwata et
al (31) revealed that Smad4 can
synergistically regulate the fate of the medial edge epithelium
during palatal fusion in mice. Accordingly, it was speculated that
miRNA-1260b may also regulate other genes associated with orofacial
clefts.
In fact, individuals born with a cleft have a higher
mortality rates at all stages of life (32). Individuals with a cleft as well as
their family members also have a higher risk of developing various
cancer types. Andrade Filho et al (33) found that GSK-3β and Axin2 belong to
the Wnt pathway and increase susceptibility to oral squamous cell
carcinoma and colon cancer, respectively. The present study showed
that hsa-miR-205-5p and hsa-miR-24-3p regulates the levels of
GSK-3β and Axin2. Axin2 is a negative regulator of Wnt-β-catenin,
which has a critical and evolutionarily conserved role in directing
cell fate during craniofacial morphogenesis, and Axin2 mRNA is
expressed throughout palatogenesis stages (34).
As Wnt-regulating miRNAs were found to be associated
with NSCLP in the present study for the first time, they hold
potential value for further research on the association of miRNAs
with NSCLP. Wnt5A deficiency has been found to lead to a complete
cleft of the secondary palate, which exhibits distinct phenotypic
alterations at the histological, cellular and molecular level in
the anterior as well as posterior regions (35). Alterations in Wnt5A function may
perturb formation and/or fusion of the facial processes and
predispose to NSCLP (36).
WNT9B may also be involved in the clefting phenotype and is
therefore an excellent candidate gene for NSCLP. Wnt9b can activate
the Wnt/β-catenin response in first branchial arch mesenchyme. When
A/WySn mice were bred with WNT9B−/− mice, there
was a higher prevalence of clefting in the progeny (67%) than in
the founders (37). WNT10A
and WNT10B have not previously been identified as being
associated with NSCLP. However, Lin et al (38) showed that WNT10a and
WNT10b transcripts in palatal epithelium were involved in
the formation of palatal rugae, with Wnt10a being more strongly
expressed in the rugae. Although it has not yet been determined
whether WNT8B and WNT5B are directly involved in
NSCLP, Kawakami et al (39)
found that limb bud formation was initiated by Wnt molecules (Wnt2b
and Wnt8), which are expressed in the lateral plate mesoderm and
which signal through β-catenin to restrict fibroblast growth factor
10 expression in the presumptive limb mesoderm. Kim et al
(40) also showed that Wnt8b can
suppress Frizzled 8a expression in the anterior neuroectoderm and
potentially affect the level and/or range of Wnt signaling.
In the present study, hsa-miR-27b-3p was also
identified by microarray analysis as being upregulated in NSCLP,
although the RT-qPCR results indicated that the difference between
NSCLP and control tissues was not significant (P=0.09). However,
its involvement in NSCLP cannot be ruled out until more samples are
collected and tested.
It should be noted that the present study only
revealed a disparity in the expression of miRNAs, which may be
associated with the Wnt family of genes and the associated
signaling pathways, between 4 NSCLP tissue samples and 4 normal
umbilical cord samples. One of the limitations of the present study
is the relatively modest number of NSCLP cases examined, which may
explain certain results. It remains to be determined whether the
miRNAs identified in the present study actually perturb the Wnt
signaling pathway, even though bioinformatics support this
hypothesis. In the future, a larger number of samples should be
collected and tested in a follow-up study to elucidate the various
factors that contribute to NSCLP.
Acknowledgements
This study was supported by grants from the Li Ka
Shing Foundation.
Glossary
Abbreviations
Abbreviations:
|
NSCLP
|
nonsyndromic cleft lip with or without
cleft palate
|
|
miRNAs
|
microRNAs
|
References
|
1
|
Meng T, Shi B, Zheng Q, Wang Y and Li S:
Clinical and epidemiologic studies of nonsyndromic cleft lip and
palate in china: Analysis of 4268 cases. Ann Plast Surg.
57:264–269. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Maestri NE, Beaty TH, Hetmanski J, Smith
EA, McIntosh I, Wyszynski DF, Liang KY, Duffy DL and VanderKolk C:
Application of transmission disequilibrium tests to nonsyndromic
oral clefts: Including candidate genes and environmental exposures
in the models. Am J Med Genet. 73:337–344. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Schliekelman P and Slatkin M: Multiplex
relative risk and estimation of the number of loci underlying an
inherited disease. Am J Hum Genet. 71:1369–1385. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Wyszynski DF, Duffy DL and Beaty TH:
Maternal cigarette smoking and oral clefts: A meta-analysis. Cleft
Palate Craniofac J. 34:206–210. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Lidral AC and Moreno LM: Progress toward
discerning the genetics of cleft lip. Curr Opin Pediatr.
17:731–739. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Lidral AC and Murray JC: Genetic
approaches to identify disease genes for birth defects with cleft
lip/palate as a model. Birth Defects Res A Clin Mol Teratol.
70:893–901. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Shi M, Wehby GL and Murray JC: Review on
genetic variants and maternal smoking in the etiology of oral
clefts and other birth defects. Birth Defects Res C Embryo Today.
84:16–29. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Bernstein E, Kim SY, Carmell MA, Murchison
EP, Alcorn H, Li MZ, Mills AA, Elledge SJ, Anderson KV and Hannon
GJ: Dicer is essential for mouse development. Nat Genet.
35:215–217. 2003. View
Article : Google Scholar : PubMed/NCBI
|
|
9
|
Chen JF, Murchison EP, Tang R, Callis TE,
Tatsuguchi M, Deng Z, Rojas M, Hammond SM, Schneider MD, Selzman
CH, et al: Targeted deletion of Dicer in the heart leads to dilated
cardiomyopathy and heart failure. Proc Natl Acad Sci USA.
105:2111–2116. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Murchison EP, Stein P, Xuan Z, Pan H,
Zhang MQ, Schultz RM and Hannon GJ: Critical roles for Dicer in the
female germline. Genes Dev. 21:682–693. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Shalgi R, Brosh R, Oren M, Pilpel Y and
Rotter V: Coupling transcriptional and post-transcriptional miRNA
regulation in the control of cell fate. Aging (Albany NY).
1:762–770. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Alvarez-Garcia I and Miska EA: MicroRNA
functions in animal development and human disease. Development.
132:4653–4662. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wienholds E, Koudijs MJ, van Eeden FJ,
Cuppen E and Plasterk RH: The microRNA-producing enzyme Dicer1 is
essential for zebrafish development. Nat Genet. 35:217–218. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Eberhart JK, He X, Swartz ME, Yan YL, Song
H, Boling TC, Kunerth AK, Walker MB, Kimmel CB and Postlethwait JH:
MicroRNA Mirn140 modulates Pdgf signaling during palatogenesis. Nat
Genet. 40:290–298. 2008. View
Article : Google Scholar : PubMed/NCBI
|
|
15
|
Hornstein E and Shomron N: Canalization of
development by microRNAs. Nat Genet. 38 Suppl:S20–S24. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Lee CT, Risom T and Strauss WM: MicroRNAs
in mammalian development. Birth Defects Res C Embryo Today.
78:129–139. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Shalgi R, Lieber D, Oren M and Pilpel Y:
Global and local architecture of the mammalian
microRNA-transcription factor regulatory network. PLoS Comput Biol.
3:e1312007. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Song L and Tuan RS: MicroRNAs and cell
differentiation in mammalian development. Birth Defects Res C
Embryo Today. 78:140–149. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Qiu WL: Oral and maxillofacial
surgeryCongenital cleft lip, facial cleft and palate cleft. 4th
edition. Beijing People's Medical Publishing House Press; Beijing:
pp. 3692000
|
|
20
|
Qiu WL: Oral and maxillofacial
surgeryCongenital cleft lip, facial cleft and palate cleft. 4th
edition. Beijing People's Medical Publishing House Press; Beijing:
pp. 374–398. 2000
|
|
21
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Cadigan KM and Nusse R: Wnt signaling: A
common theme in animal development. Genes Dev. 11:3286–3305. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Bejsovec A: Wnt pathway activation: New
relations and locations. Cell. 120:11–14. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Dale RM, Sisson BE and Topczewski J: The
emerging role of Wnt/PCP signaling in organ formation. Zebrafish.
6:9–14. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Chhabra R, Dubey R and Saini N:
Cooperative and individualistic functions of the microRNAs in the
miR-23a~27a~24-2 cluster and its implication inhuman diseases. Mol
Cancer. 9:2322010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
He F, Xiong W, Wang Y, Li L, Liu C,
Yamagami T, Taketo MM, Zhou C and Chen Y: Epithelial Wnt/β-catenin
signaling regulates palatal shelf fusion through regulation of
Tgfβ3 expression. Dev Biol. 350:511–519. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Yu J, Chen Y, Qin L, Cheng L, Ren G, Cong
P, Mo D and He Z: Effect of miR-205 on 3T3-L1 preadipocyte
differentiation through targeting to glycogen synthase kinase 3
beta. Biotechnol Lett. 36:1233–1243. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kim M, Datta A, Brakeman P, Yu W and
Mostov KE: Polarity proteins PAR6 and aPKC regulate cell death
through GSK-3beta in 3D epithelial morphogenesis. J Cell Sci.
120:2309–2317. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Liu KJ, Arron JR, Stankunas K, Crabtree GR
and Longaker MT: Chemical rescue of cleft palate and midline
defects in conditional GSK-3beta mice. Nature. 446:79–82. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Hirata H, Ueno K, Nakajima K, Tabatabai
ZL, Hinoda Y, Ishii N and Dahiya R: Genistein downregulates
onco-miR-1260b and inhibits Wnt-signalling in renal cancer cells.
Br J Cancer. 108:2070–2078. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Iwata J, Suzuki A, Pelikan RC, Ho TV,
Sanchez-Lara PA, Urata M, Dixon MJ and Chai Y: Smad4-Irf6 genetic
interaction and TGFβ-mediated IRF6 signaling cascade are crucial
for palatal fusion in mice. Development. 140:1220–1230. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Christensen K, Juel K, Herskind AM and
Murray JC: Long term follow up study of survival associated with
cleft lip and palate at birth. BMJ. 328:14052004. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
de Filho PA Andra, Letra A, Cramer A,
Prasad JL, Garlet GP, Vieira AR, Ferris RL and Menezes R: Insights
from studies with oral cleft genes suggest associations between
WNT-pathway genes and risk of oral cancer. J Dent Res. 90:740–746.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Parr BA, Shea MJ, Vassileva G and McMahon
AP: Mouse Wnt genes exhibit discrete domains of expression in the
early embryonic CNS and limb buds. Development. 119:247–261.
1993.PubMed/NCBI
|
|
35
|
He F, Xiong W, Yu X, Espinoza-Lewis R, Liu
C, Gu S, Nishita M, Suzuki K, Yamada G, Minami Y and Chen Y: Wnt5a
regulates directional cell migration and cell proliferation via
Ror2-mediated noncanonical pathway in mammalian palate development.
Development. 135:3871–3879. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Chiquet BT, Blanton SH, Burt A, Ma D, Stal
S, Mulliken JB and Hecht JT: Variation in WNT genes is associated
with non-syndromic cleft lip with or without cleft palate. Hum Mol
Genet. 17:2212–2218. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Juriloff DM, Harris MJ, McMahon AP,
Carroll TJ and Lidral AC: Wnt9b is the mutated gene involved in
multifactorial nonsyndromic cleft lip with or without cleft palate
in A/WySn mice, as confirmed by a genetic complementation test.
Birth Defects Res A Clin Mol Teratol. 76:574–579. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Lin C, Fisher AV, Yin Y, Maruyama T, Veith
GM, Dhandha M, Huang GJ, Hsu W and Ma L: The inductive role of
Wnt-β-Catenin signaling in the formation of oral apparatus. Dev
Biol. 356:40–50. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Kawakami Y, Capdevila J, Büscher D, Itoh
T, Rodríguez Esteban C and Izpisúa Belmonte JC: WNT signals control
FGF-dependent limb initiation and AER induction in the chick
embryo. Cell. 104:891–900. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Kim SH, Shin J, Park HC, Yeo SY, Hong SK,
Han S, Rhee M, Kim CH, Chitnis AB and Huh TL: Specification of an
anterior neuroectoderm patterning by Frizzled8a-mediated Wnt8b
signalling during late gastrulation in zebrafish. Development.
129:4443–4455. 2002.PubMed/NCBI
|