Introduction
Identifying effective therapy for the treatment of
malignant tumors is a global challenge, and while the development
of medical technology has led to an increase in cancer therapies, a
treatment strategy of adequate efficacy has not yet been
determined. Strategies employing a combination of surgery,
radiotherapy, chemotherapy and immunotherapy, as well as
clinical-based rehabilitation, have improved the quality of life
for cancer patients and increased the 5-year-survival rate
(1,2). Combination therapies integrate the
research achievements from various specializations, making the
treatment more diversified. Combination therapy may exert the
advantages and avoid the drawbacks of individual treatments,
allowing for the achievement of maximum therapeutic effect.
However, due to the presentation of serious complications, side
effects (including pain, infection, cardiovascular and
gastrointestinal side effects and depression) (3), tumor recurrence and metastasis
following therapy, studies investigating alternative therapeutics
are warranted. Optimal cancer therapies require the capacity to
specifically inhibit tumor cells, while having minimal adverse
effects on normal cells. Therefore, the identification of a
molecular target that specifically inhibits tumor cell growth is
warranted.
Survivin is a member of the inhibitor of apoptosis
family of proteins, which collectively has roles in the inhibition
of caspase activation and regulation of mitosis. Survivin protein
is expressed at high levels in the majority of human tumors and
fetal tissue, though is absent in terminally differentiated cells
(4). In tumor tissue, survivin
promotes the anti-apoptotic activity of tumor cells, while also
contributing to tumor angiogenesis and resistance to
chemoradiotherapy (5–7). Specifically, survivin has been found to
promote the formation and development of tumors by the following
mechanisms: i) interaction with cell division protein kinase 4
leading to acceleration in the S-phase of the cell cycle (8); ii) increasing the expression of human
angiopoietin 1, thus promoting the formation of capillary networks
and proliferation of endothelial cells (9); iii) overexpression in the G2/M phase of
the cell cycle in a cycle-regulated manner and association with
microtubules of the mitotic spindle, thus inducing aberrant mitosis
(10); and iv) suppressing the
release of mitochondrial cytochrome C, thus inhibiting apoptosis
and promoting tumor progression (11).
As a commonly upregulated gene in tumor cells,
survivin is considered to be a potential cancer biomarker and a
novel target for cancer treatment (12). A number of studies have documented
the overexpression of survivin in different human tumor types,
including leukemia, brain tumors, liver, breast, gastric, lung,
colon, prostate, lymph, oesophageal, pancreatic, skin, cervical and
ovarian cancer (13). Therefore,
studies are aiming to identify novel strategies that target the
expression and function of survivin for the treatment of
cancer.
RNA interference (RNAi) is an evolution-conserved
cell defense mechanism mediated by small interfering RNA (siRNA)
(14). Target genes are knocked down
by siRNA inhibition of the corresponding mRNA, resulting in
suppressed gene expression (15).
Compared with conventional antisense strategies, RNAi has
demonstrated greater efficacy in the silencing of specific genes by
allowing low level target gene expression and subsequent protein
function, thus resulting in fewer adverse effects, such as pain,
nausea and vomiting, tiredness and sleeping problems, occurring in
patients (12). However, rapid
degradation of siRNA within the extracellular environment and an
inability to traverse cellular membranes means that the delivery of
siRNA to its intracellular targets remains a challenge (16).
In the present study, liposomes were used for the
intracellular delivery of siRNA against survivin. siRNA liposomal
vehicles were successfully constructed and transfected into cell
lines (MSA-MB-231, SGC-7901, HeLa, A549, SK-OV-3 and Raji, PC-3) of
a number of common human tumor types (breast cancer, gastric
carcinoma, cervical carcinoma, lung carcinoma, ovarian carcinoma,
lymphoma, prostate carcinoma, respectively). Changes in the level
of survivin protein expression and inhibition of tumor cell
proliferation and apoptosis were then determined by reverse
transcription-quantitative polymerase chain reaction (RT-qPCR),
western blot analysis, the MTT method, Hoechst 33258 staining and
Annexin V/PI staining in vitro, in order to determine the
efficacy of survivin as a potential gene target in the treatment of
cancer.
Materials and methods
Cell lines and culture
Tumor cell lines MDA-MB-231 (human breast
carcinoma), SGC-7901 (human gastric carcinoma), HeLa (human
cervical carcinoma) and A549 (human lung carcinoma) were obtained
from Biomics Biotechnologies Co., Ltd. (Nantong, China). SK-OV-3
(human ovarian carcinoma), Raji (human lymphoma) and PC-3 (human
prostate carcinoma) cell lines were purchased from the Cell Banks
of Type Culture Collection at the Chinese Academy of Sciences
(Beijing, China). MDA-MB-231, SGC-7901 and Raji cells were
maintained in RPMI 1640 (Thermo Fisher Scientific, Inc., Waltham,
MA, USA) supplemented with 10% fetal bovine serum (FBS; Hyclone; GE
Healthcare, Logan, UT, USA), and HeLa and A549 cells were
maintained in Dulbecco's modified Eagle medium (DMEM; Thermo Fisher
Scientific, Inc.) supplemented with 10% FBS. SK-OV-3 cells were
maintained in McCoy's 5A modified medium (Life Technologies; Thermo
Fisher Scientific, Inc.) and PC-3 were maintained in DMEM/ nutrient
mixture F12 (Thermo Fisher Scientific, Inc.) supplemented with 10%
FBS. All cell lines were cultured at 37°C in 5% CO2 for
1–3 days.
siRNA constructs
siRNAs were synthesized using the following
sequences: Survivin, forward 5′-GCAUCUCUACAUUCAAGAA-3′ and reverse
5′-UUCUUGAAUGUAGAGAUGC-3′; a scrambled sequence (si-NC), forward
5′-UUCUCCGAACGUGUCACGU-3′ and reverse 5′-ACGUGACACGUUCGGAGAA-3′.
si-NC was used as the negative control and was non-homologous to
all known human DNA sequences.
Preparation of siRNA-liposomes
siRNA was incorporated into liposomes for
intracellular delivery. Disaturated phosphatidylcholine (Avanti
Polar Lipids, Alablaster, AL, USA), cholesterol,
dioctadecyldimethylammonium chloride and
N-palmitoyl-sphingosine-1-succinyl (also known as
methoxypolyethylene glycol 2000; Sigma-Aldrich; Merck Millipore,
Darmstadt, Germany) were mixed in 100% ethanol at a molar ratio of
25:45:25:2.5 to produce pre-formed vesicles (PFV). siRNA (in a 30%
ethanol solution) and PFV's (in a 30% ethanol solution) were warmed
to 35–40°C for 10 min, respectively. Subsequently, siRNA was added
slowly to the PFVs while the solution was stirred continuously to
form siRNA-liposomes. Following the encapsulation, the solution was
concentrated by centrifugal filtration (Merck Millipore) and
sterilized by filtration using a 0.22-mm Supor membrane filter
(Gelman Sciences, Ann Arbor, MI, USA), and the final products were
produced. The average diameters of siRNA-survivin-liposomes and
siRNA-NC-liposomes were 70.7±29.077 and 64.9±26.128 nm,
respectively.
In vitro transfection
Before transfection, the cancer cells at the growth
stage were harvested and a cell suspension was prepared
(1×105 cells/ml) with the use of DMEM-H medium (Life
Technologies; Thermo Fisher Scientific, Inc.). Following this, 1 ml
cell suspension was added in each well. The wells were divided into
three groups: Survivin siRNA group (survivin siRNA nanoliposome),
NC group (NC siRNA nanoliposome) and normal group (without
transfection). The cells were plated and grown to 70–80% confluence
prior to transfection. Survivin and NC siRNA-liposomes were mixed
with Opti-MEM (Life Technologies; Thermo Fisher Scientific, Inc.)
and left to stand for 5 min at room temperature. Subsequently, the
survivin and NC siRNA-liposomes were mixed with an equal volume of
cell suspension and cultured at 37°C in 5% CO2. After
4–6 h, the media were changed to fresh media for growth, as
described, and cells were incubated for an additional 48 h at 37°C.
A total of 24 h after transfection, cells were fixed with 4%
paraformaldehyde for 30 min and washed with phosphate buffered
saline (PBS) three times, 5 min each time, and stained with Hoechst
33258 (Life Technologies; Thermo Fisher Scientific, Inc.) for 10–20
min. Transfection efficiency was observed using a fluorescence
microscope (magnification, ×100; Nikon Corp., Tokyo, Japan).
RNA extraction and RT-qPCR
Total RNA was extracted from the transfected cell
lines with RISO™ RNA Isolation Reagent (Biomics Biotechnologies
Co., Ltd.), according to the manufacturer's instructions. cDNA was
synthesized with a SensiMix™ SYBR-Green One-Step kit (Quantace;
Bioline Reagents, Ltd., London, UK) according to the manufacturer's
instructions. qPCR was performed using an ABI PRISM real-time PCR
system (Applied Biosystems; Thermo Fisher Scientific, Inc.).
Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) was used as an
endogenous control. PCR primers were as follows: survivin, forward
5′-ACGACCCCATAGAGGAACAT-3′ and reverse, 5′-TCCGCAGTTTCCTCAAATTC-3′;
GAP DH, forward 5′-GAAGGTGAAGGTCGGAGTC-3′ and reverse
5′-GAAGATGGTGATGGGATTTC-3′. The RT reaction was performed at 42°C
for 30 min. qPCR was then performed under the following conditions:
The initial PCR incubation was 95°C for 10 min and the
amplification loop (95°C for 20 sec, 58°C for 30 sec and 72°C for
30 sec) was repeated 45 times. Following the reaction, the
amplification and melting curves were analyzed. Relative
quantification was performed using the comparative
2-ΔΔCq method (17),
using GAPDH as a reference gene. All analyses were performed in
triplicate.
Western blot analysis
The expression of survivin protein was determined by
western blotting. Cells were washed with PBS and placed on ice for
4 min following a 3–5 min incubation in a boiling water bath, then
centrifuged at 13,500 × g at 4°C for 15 min. Following
centrifugation, the protein pellet was resuspended and the
resulting cell protein extract samples were analyzed by 8% SDS-PAGE
(15 µl/lane), then transferred to polyvinylidene fluoride membrane
filters (EMD Millipore, Billerica, MA, USA). Membranes were
incubated at room temperature for 2 h in 5% non-fat dry milk, and
rabbit polyclonal anti-survivin antibody (1:500; ab469; Abcam,
Cambridge, UK) and mouse anti-β-actin monoclonal antibody (1:400;
BM0005; Wuhan Boster Biological Technology, Ltd., Wuhan, China)
were added and incubated overnight at 4°C. Following three washes
with Tris-buffered saline with Tween, goat anti-rabbit and
anti-mouse secondary antibodies (both 1:500; BA1054 and BA1051,
respectively; both Wuhan Boster Biological Technology, Ltd.) were
added and incubated at room temperature for 2 h. The gray-scale
values of the blots were quantified with the computer-based system
ImageJ 1.441 (National Institutes of Health, Bethesda, MA, USA).
The results were shown as mean values from three replications.
Cell proliferation assay
Growth of the cancer cell lines was evaluated using
an MTT assay. The assay was performed at 0, 24, 48, 72 and 96 h
post-transfection. Cells were plated in medium supplemented with
serum into 96-well plates at a density of 1×104 cells/ml
(100 µl). Absorbance was determined following incubation with MTT
solution at 37°C without light for 4 h. The supernatant was
discarded and 150 µl DMSO was added. Optical density (OD) values
were measured at a wavelength of 570 nm with a microplate reader,
with normalization to a blank control (dimethyl sulfoxide). The
inhibition rate was calculated relative to the OD value in the
normal control group.
Hoechst staining
Apoptotic cells were detected by Hoechst 33258
staining. Cells in survivin siRNA, NC and normal groups were fixed
with 4% paraformaldehyde for 10 min at room temperature and washed
three times with PBS, followed by staining with Hoechst 33258
(Thermo Fisher Scientific, Inc.) for 10–20 min. Apoptotic features
were observed under a fluorescence microscope.
Annexin V/propidium iodide (PI)
staining
To quantify apoptosis, cells were double-stained
with an Annexin V-Fluorescein isothiocyanate Apoptosis Detection
kit (Beyotime Institute of Biotechnology, Haimen, China) and PI,
according to the manufacturer's protocol. The percentages of cells
in different cell cycle phases were then analyzed by FACS Calibur
flow cytofluorometry (BD Biosciences, San Jose, CA USA) for the
presence of viable (Annexin V− and PI−),
early apoptotic (Annexin V+ and PI−) and late
apoptotic (Annexin V+ and PI+) cells, as
described by a previous method (18).
Statistical analysis
Data are presented as the mean ± standard deviation.
Results were statistically analyzed using SPSS 10.0 software (SPSS,
Inc., Chicago, IL, USA). A t-test was performed to determine
statistical differences between the different groups and P<0.05
was considered to indicate a statistically significant
difference.
Results
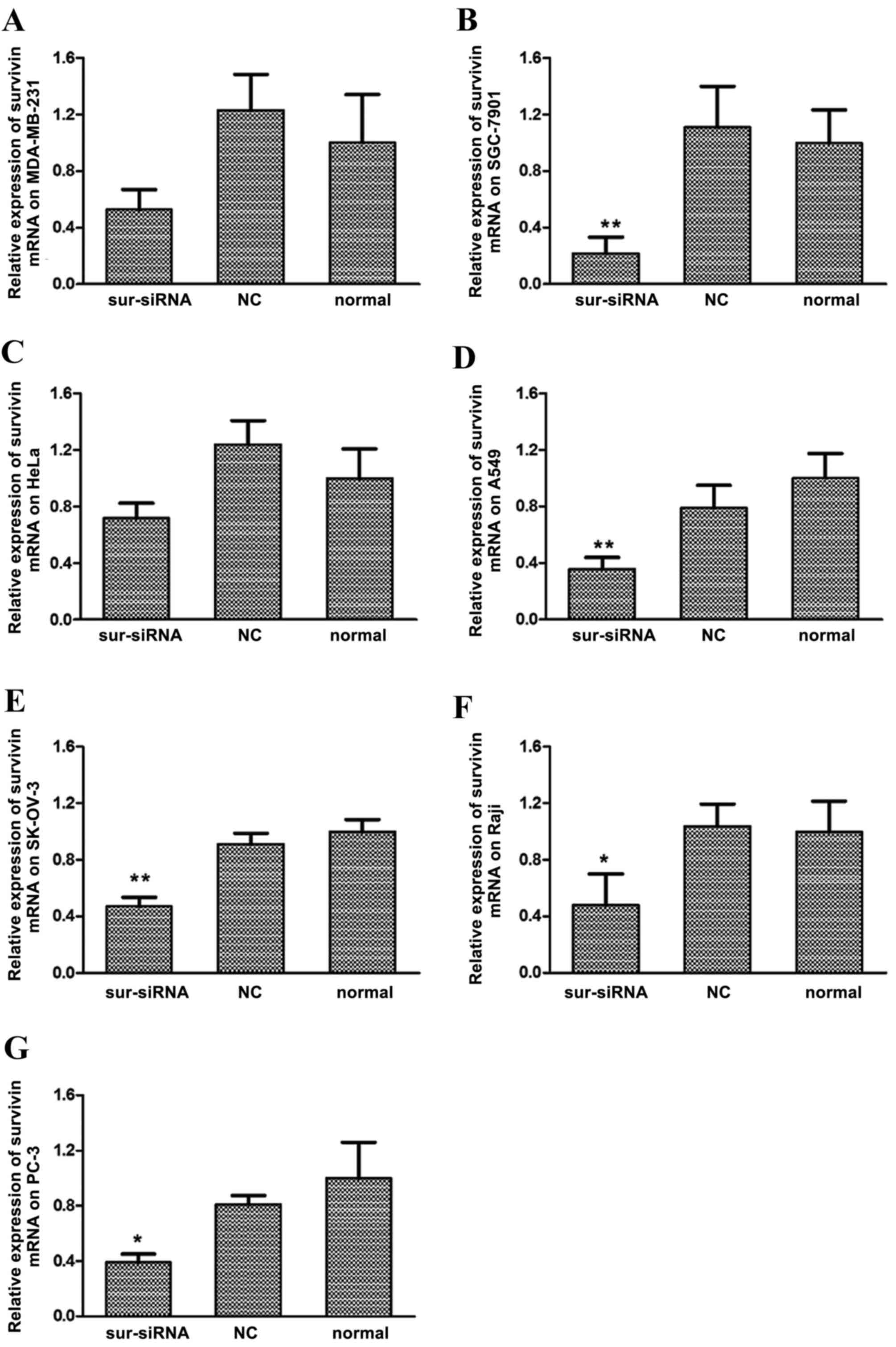
siRNA transfection suppresses survivin
gene expression in human cancer cell lines
Expression of survivin mRNA in the MDA-MB-231,
SGC-7901, HeLa, A549, SK-OV-3, Raji and PC-3 cancer cell lines was
suppressed following siRNA transfection, as indicated by results of
RT-qPCR (Fig. 1A-G, respectively).
Among them, cell line SGC-7901 exhibited the greatest sensitivity
to survivin siRNA, whereby expression of survivin mRNA
significantly decreased to 22% relative to the normal control group
(P<0.01, Fig. 1B). Expression of
survivin mRNA also significantly decreased compared with the
control group in the A549 (36%; P<0.01, Fig. 1D), SK-OV-3 (47%, P<0.01, Fig. 1E) Raji (45%; P<0.05, Fig. 1F) and PC-3 (39%; P<0.05; Fig. 1G) cell lines. Marked decreases in
survivin mRNA were observed in MDA-MB-231 (50%, Fig. 1A) and HeLa (65%, Fig. 1C) cells, relative to their respective
untreated (normal) control groups. For all cell lines, differences
in survivin mRNA levels between the si-NC and normal control groups
were not significant.
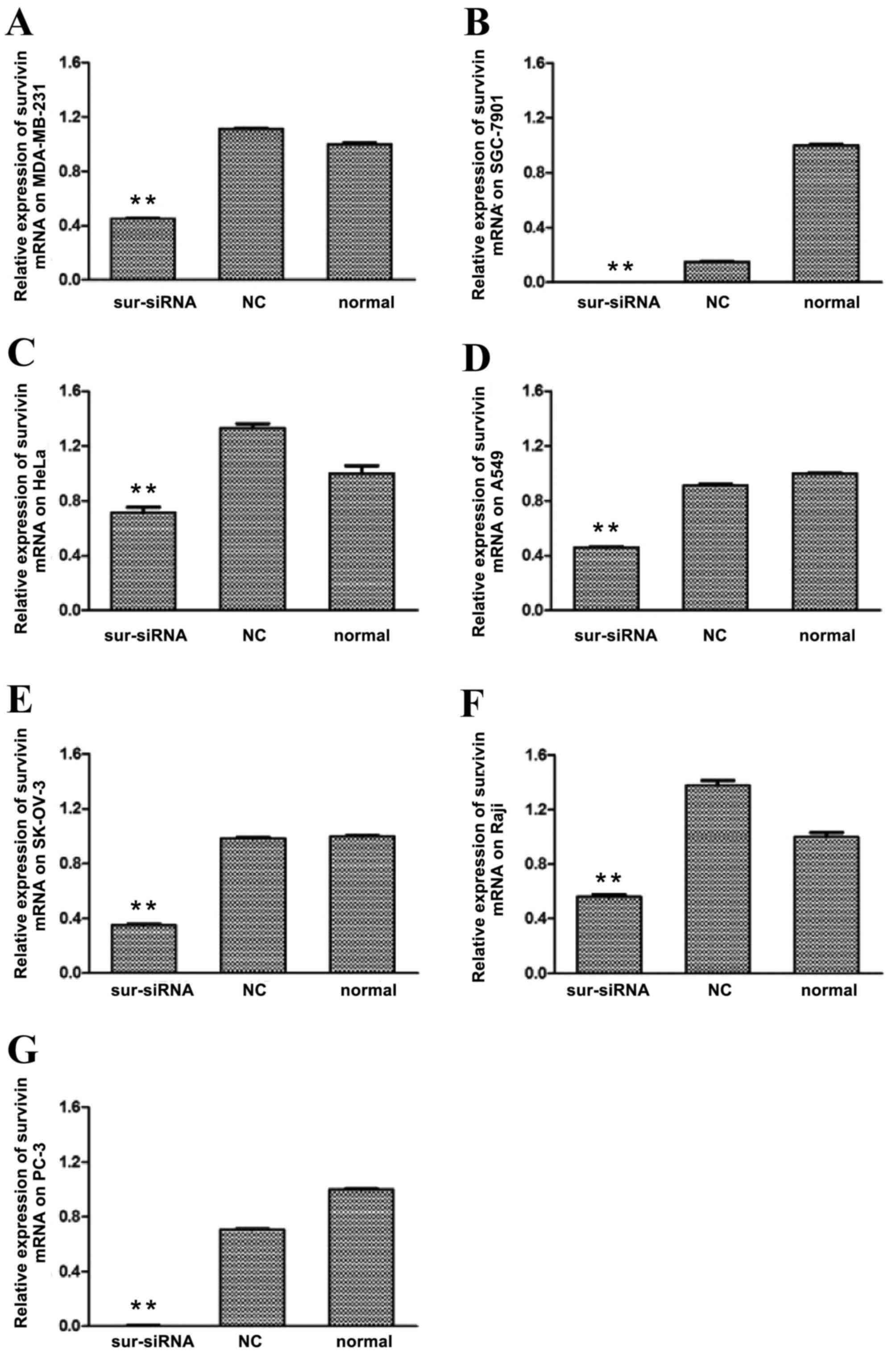
The level of survivin protein expression in each
cell line following survivin siRNA transfection was subsequently
determined by western blot analysis. As depicted in Fig. 2, levels of survivin protein in the
survivin siRNA groups of all cell lines were significantly
(P<0.01) downregulated relative to that in the control groups,
particularly for SGC-7901 and PC-3, where the expression was almost
completely inhibited. The survivin protein expression levels of the
remaining cancer cells were reduced to 40–70% of the level of the
normal control group. For all cell lines, differences in survivin
protein levels between the si-NC and normal control groups were not
significant.
Survivin siRNA inhibits cancer cell
proliferation
The inhibitory effects of survivin siRNA in each
cell line were analyzed by an MTT assay at 24, 48, 72 and 96 h
post-transfection. Inhibition rates are listed in Table I. The data indicates that
proliferation of the cancer cell lines was reduced following
survivin knockdown by siRNA. At 96 h post-transfection, significant
inhibition of cell proliferation was detected in all cell lines
relative to the si-NC group (P<0.05).
 | Table I.Inhibition rate of survivin siRNA on
cancer cell lines. |
Table I.
Inhibition rate of survivin siRNA on
cancer cell lines.
|
|
| Inhibition rate (mean
± standard deviation, %) |
|---|
|
|
|
|
|---|
| Cell lines | Groups | 24 h | 48 h | 72 h | 96 h |
|---|
| MDA-MB-231 | siRNA | 19.2±4.8a | 17.5±1.4a | 16.6±3.7a |
9.9±2.2a |
|
| si-NC |
9.1±5.1 |
4.1±2.2 |
4.6±1.9 |
−1.5±3.7 |
| SGC-7901 | siRNA |
9.0±0.8 | 15.5±5.6 |
5.3±1.8 |
6.8±4.8a |
|
| si-NC |
5.2±3.5 |
8.3±5.0 |
1.1±4.1 |
1.1±3.7 |
| HeLa | siRNA |
7.1±2.2 | 18.6±2.2 |
25.5±4.6a |
46.3±1.8a |
|
| si-NC |
8.6±3.6 | 18.3±3.4 |
5.8±4.2 | 33.2±4.3 |
| A549 | siRNA |
4.0±1.0 |
1.4±2.2 |
25.4±2.5a |
29.4±1.2a |
|
| si-NC |
2.8±2.4 |
2.3±2.1 |
2.9±3.7 |
3.3±3.1 |
| SK-OV-3 | siRNA |
1.2±2.1 |
2.8±4.0 |
−0.9±4.3 |
7.0±0.9a |
|
| si-NC |
−1.7±3.3 |
3.0±3.2 |
−2.4±3.8 |
0.4±1.8 |
| Raji | siRNA |
2.0±0.4 |
3.0±0.02 |
1.7±5.0 |
10.8±4.0a |
|
| si-NC |
1.4±0.3 |
0.5±0.7 |
0.5±3.1 |
7.4±7.0 |
| PC-3 | siRNA |
1.2±1.8 |
0.4±1.5 |
4.7±2.1 |
5.5±2.7a |
|
| si-NC |
0.2±2.5 |
1.0±0.9 |
1.4±1.9 |
−0.5±4.1 |
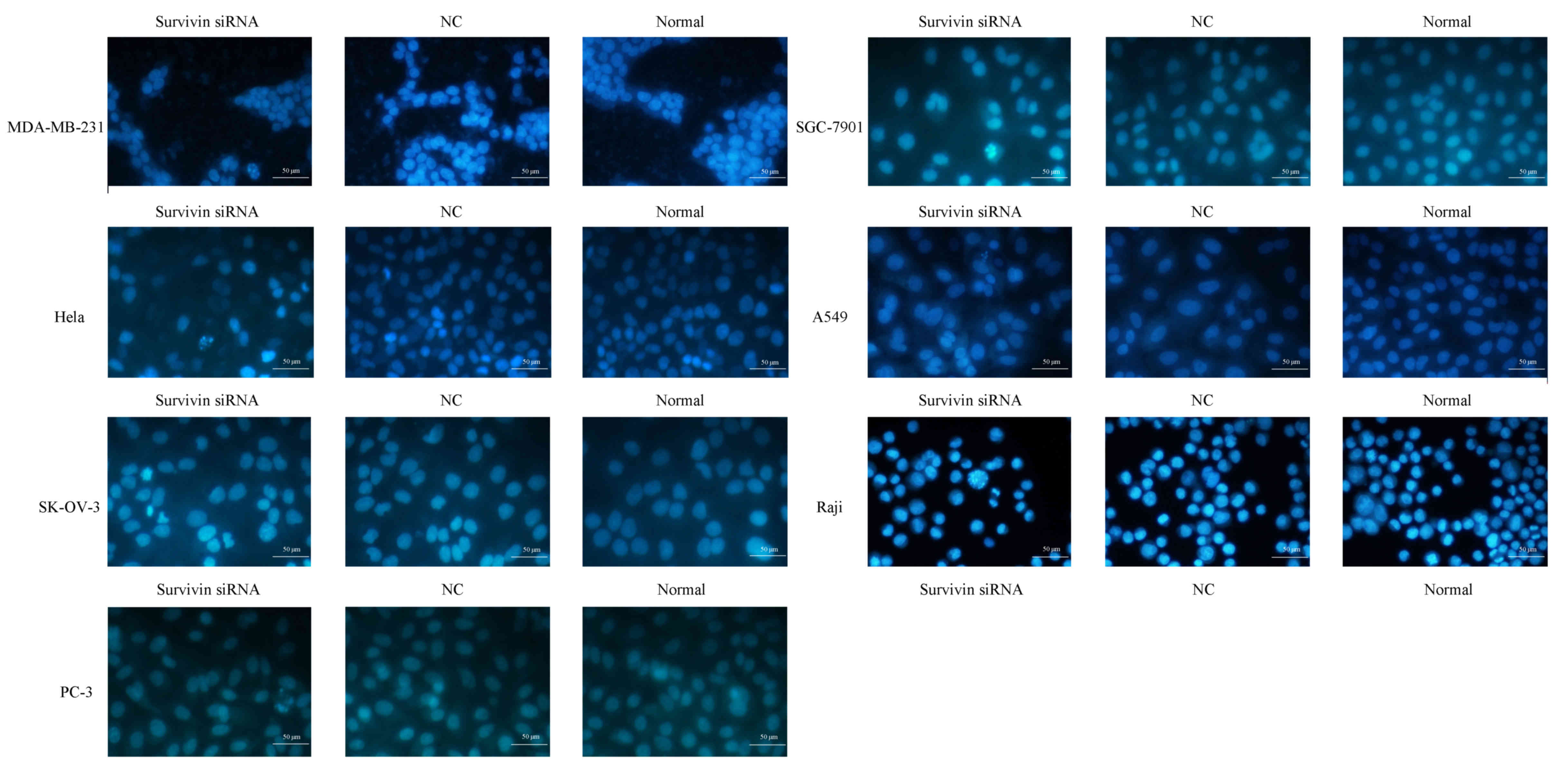
Survivin knockdown induces apoptosis
of cancer cell lines
Hoechst 33258 staining was performed in order to
evaluate the levels of cell apoptosis following survivin mRNA
knockdown (Fig. 3). Cell nuclei were
invariably stained blue by Hoechst 33258, however the nuclei
morphology and extent of staining were distinct between normal and
apoptotic cells. Normal cells with intact nuclei appeared light
blue in color, while apoptotic cells with condensed or fragmented
nuclei exhibited bright blue, fragmented nuclei of variable sizes.
As depicted in Fig. 3, the nuclei of
cancer cell lines transfected with survivin siRNA exhibited
granular bright blue fluorescence, reflecting the typical
morphological characteristics of apoptosis. These indicators of
apoptosis were not observed in the si-NC and normal control
groups.
Cells in each group were subsequently double-stained
with Annexin V/PI and analyzed with flow cytometry (Fig. 4). The apoptosis rate of MDA-MB-231
and PC-3 cells was significantly increased in the survivin
siRNA-transfected groups compared with the apoptosis rate in the
si-NC and normal control groups (all P<0.05; Table II). The apoptosis rate was
significantly increased in the SGC-7901, HeLa and A549 survivin
siRNA-transfected groups compared with the apoptosis rate in the
normal control group (all P<0.05; Table II); however, the differences for
SK-OV-3 and Raji were not significant.
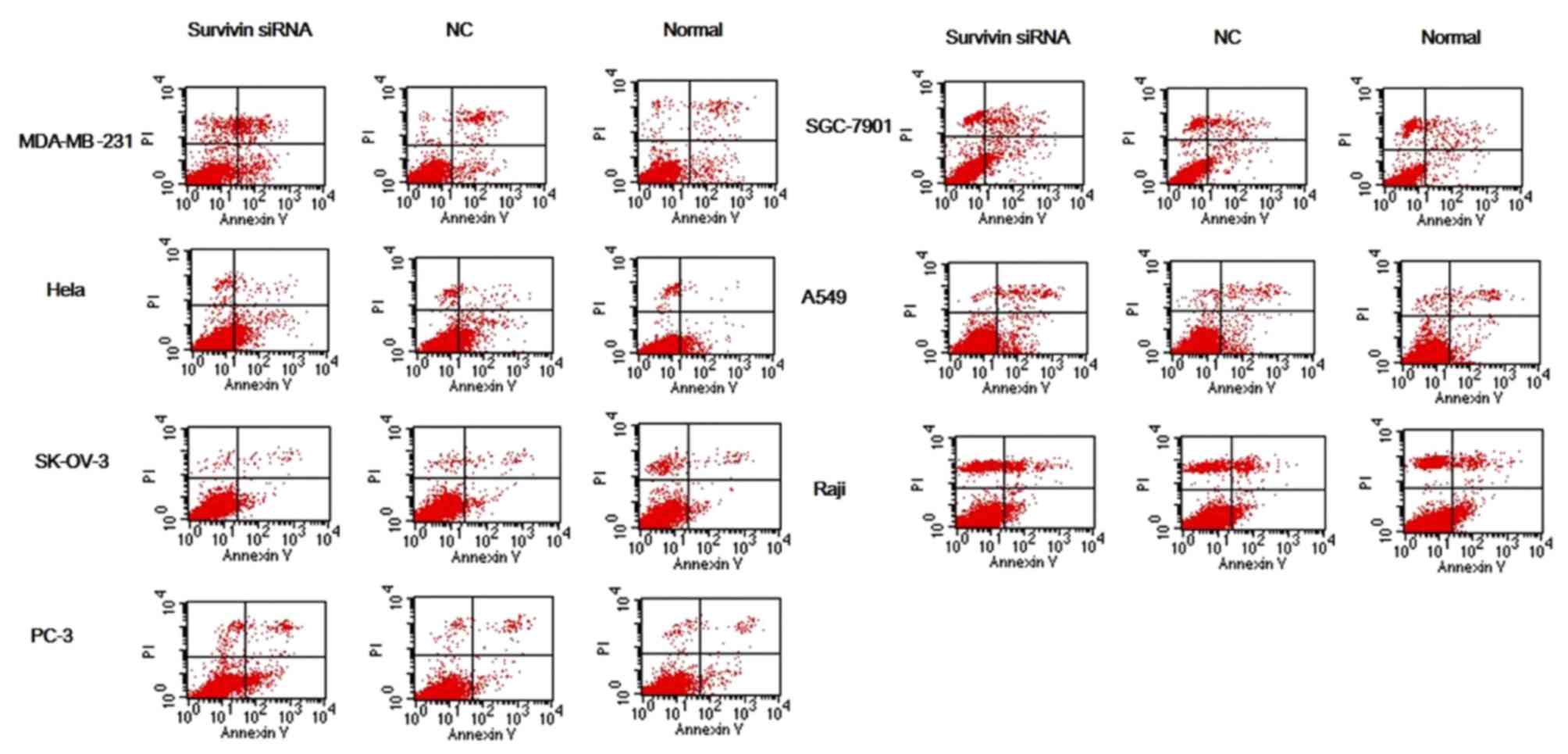 | Figure 4.Apoptosis of cancer cell lines
transfected with survivin siRNA was detected by Annexin V/PI double
staining. Following siRNA knockdown of survivin in cancer cell
lines (MDA-MB-231, SGC-7901, HeLa, A549, SK-OV-3, Raji and PC-3),
levels of cell apoptosis were evaluated by Annexin V/PI double
staining followed by flow cytofluorometry analysis. Early and late
stage apoptosis (Annexin V+/PI− and Annexin
V+/PI+, respectively) in the survivin siRNA
groups were increased to some extent relative to the si-NC and
normal control groups. siRNA, small interfering RNA; NC, scrambled
negative control siRNA; normal, untreated negative control. |
| Table II.Effect of survivin siRNA on the
apoptosis rate of different cancer cells detected by flow cytometry
after Annexin-V/propidium iodide staining. |
Table II.
Effect of survivin siRNA on the
apoptosis rate of different cancer cells detected by flow cytometry
after Annexin-V/propidium iodide staining.
|
| Apoptosis rate
(mean ± standard deviation, %) |
|---|
|
|
|
|---|
| Cell lines | Survivin siRNA
group | NC group | Normal group |
|---|
| MDA-MB-231 |
17.21±2.13a,b |
4.29±1.47 |
3.27±1.32 |
| SGC-7901 |
19.45±2.58c | 13.77±2.71 | 12.72±2.83 |
| HeLa |
5.37±1.04c |
4.45±1.01 |
2.62±0.91 |
| A549 |
19.89±3.18c | 15.29±2.86 | 13.38±2.15 |
| SK-OV-3 |
4.87±0.84 |
4.66±0.84 |
4.53±0.81 |
| Raji |
8.98±1.03 |
8.65±1.02 |
7.89±0.92 |
| PC-3 |
7.38±1.56b,d |
3.77±1.19 |
3.72±0.89 |
Discussion
Malignant tumors are characterized by uncontrolled
and accelerated proliferation of cancer cells and are associated
with high morbidity and mortality. In 2012, 14.1 million new cancer
cases occurred globally, and cancer resulted 8.2 million deaths
(19,20). In the treatment of cancer, combined
therapy (21), which includes
surgery, chemotherapy, radiotherapy and biological therapy, is
generally adopted. Of these, biological therapy is considered to
have the greatest efficacy at each stage of tumor development, as
well as the least side effects (22). The occurrence and development of
tumors is in part due to synergistic and antagonistic interactions
between multiple genes, including oncogenes, tumor suppressor
genes, apoptosis inhibiting genes and reverse transcriptases.
Therefore, along with uncontrolled proliferation of tumor cells,
tumorigenesis is also associated with restricted tumor cell
apoptosis (23,24).
Survivin is an intracellular protein that belongs to
the inhibitor of apoptosis family (5). The protein is considered to be a
potential target of cancer therapy, due to its observed
overexpression in the majority of malignancies. Therefore, the
potential mechanisms of survivin regarding its inhibition of
apoptosis have been evaluated (25).
As survivin is considered to serve a key regulatory role in cancer
progression, methods that suppress survivin expression may promote
tumor cell apoptosis and thus have anticancer effects.
RNAi is a post-transcriptional gene silencing
mechanism that uses siRNA, and may be used to analyze cancer
biology (26). In the present study,
RNAi was used to determine the potential role of survivin in human
cancer. Specific siRNA against survivin was transfected into a
number of human cancer cell lines and its effect on cell
proliferation and apoptosis was assessed using RT-qPCR, western
blot analysis, MTT assay and Hoechst and Annexin V/PI staining.
Using RT-qPCR and western blotting, it was determined that the
expression of survivin was suppressed following transfection with
survivin siRNA, In addition, data from the MTT assay indicated that
proliferation of cancer cells lines was reduced. Subsequent
staining assays also demonstrated that siRNA knockdown of survivin
led to higher rates of apoptosis in cancer cells. Collectively,
these data suggest that survivin may serve a key role in cancer
progression.
siRNA technology may be a potential therapeutic
method of treating tumors, however the difficulties in delivering
siRNA into target cells has limited its use (27). Thus, studies investigating effective
delivery systems for siRNA are warranted. A previous study
documented the intracellular delivery of siRNA by lipid-based
agents and carriers, including atelocollagen, protamine-antibody
fusion protein and polyethyleneimine (28). Prior to initiation of the present
study, a cationic liposome was optimized and used as an siRNA
transfer vehicle. This liposome was chosen due to its ability to
efficiently traverse the cell membrane, relatively high stability
and biological compatibility.
The distinct distribution of survivin in cancer
tissues makes it a suitable target for RNAi-based antitumor
therapy, and downregulation of survivin by RNAi has been documented
in gastric cancer (29), rectal
cancer (30), bladder cancer
(31) and lung cancer cells
(14). The results of these studies
indicated that RNAi-targeting successfully inactivated the
anti-apoptotic function of survivin and inhibited tumor growth.
Furthermore, expression of survivin in tissues and organs is
considered to be an indicator of precancerosis (32), while also being associated with tumor
invasion and metastasis (33),
sensitivity to chemotherapy (34)
and degree of lymphatic metastasis (35). Therefore, survivin is a potential
biomarker for the early diagnosis, treatment and prognosis of
cancer.
In conclusion, the present study achieved
downregulation of survivin in a number of common human cancer cell
lines by RNAi. It was observed that siRNA knockdown of survivin
inhibited the growth of tumor cells and induced their apoptosis,
demonstrating that survivin is a potential gene target in cancer
therapy. However, due to differences between in vitro
experiments and clinical therapy, the present study was unable to
simulate an in vivo environment completely, which made the
obtained results biased to some degree. Thus, further studies in
animal models are required in the future.
Acknowledgements
The present study was supported by the National High
Technology Research and Development Program of China (grant no.
2012AA020804).
References
|
1
|
Sydorak RM and Applebaum H: Soft Tissue
TumorsFundamentals of Pediatric Surgery. Mattei P: Springer-Verlag;
New York, NY: pp. 755–760. 2011, View Article : Google Scholar
|
|
2
|
Li W: The new concept of cancer supportive
treatment. Electronic Journal of Metabolism and Nutrition of
Cancer. 1:2014.
|
|
3
|
Horiot JC: At last-progress in the
assessment of the adverse effects of cancer treatments. Lancet
Oncol. 8:568–570. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Barrett PT, Petrides K, Eysenck SB and
Eysenck HJ: The Eysenck personality questionnaire: An examination
of the factorial similarity of PE, N and L across 34 countries.
Personality Individual Differences. 25:805–819. 1998. View Article : Google Scholar
|
|
5
|
Ambrosini G, Adida C and Altieri DC: A
novel anti-apoptosis gene, survivin, expressed in cancer and
lymphoma. Nat Med. 3:917–921. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zaffaroni N and Daidone MG: Survivin
expression and resistance to anticancer treatments: Perspectives
for new therapeutic interventions. Drug Resistance Updates.
5:65–72. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kong LJ, Zhao J and Fan WW: Survivin and
Tumor Angiogenesis. Zhong Liu Xue Za Zhi. 15:950–952. 2009.(In
Chinese).
|
|
8
|
Suzuki A, Hayashida M, Ito T, Kawano H,
Nakano T, Miura M, Akahane K and Shiraki K: Survivin initiates cell
cycle entry by the competitive interaction with Cdk4/p16 (INK4a)
and Cdk2/cyclin E complex activation. Oncogene. 19:3225–3234. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Mesri M, Morales-Ruiz M, Ackermann EJ,
Bennett CF, Pober JS, Sessa WC and Altieri DC: Suppression of
vascular endothelial growth factor-mediated endothelial cell
protection by survivin targeting. Am J Pathol. 158:1757–1765. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Li F, Ambrosini G, Chu EY, Plescia J,
Tognin S, Marchisio PC and Altieri DC: Control of apoptosis and
mitotic spindle checkpoint by survivin. Nature. 396:580–584. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kayaselcuk F, Nursal T, Polat A, Noyan T,
Yildirim S, Tarim A and Seydaoglu G: Expresion of survivin, Bcl-2,
P53 and bax in breast carcioma and ductal intraepithelial neoplasia
(DIN 1a). J Exp Clin Cancer Res. 23:105–112. 2004.PubMed/NCBI
|
|
12
|
Ryan BM, O'Donovan N and Duffy MJ:
Survivin: A new target for anti-cancer therapy. Cancer Treat Rev.
35:553–562. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Altieri DC: Survivin, versatile modulation
of cell division and apoptosis in cancer. Oncogene. 22:8581–8589.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Chen XQ, Yang S, Li ZY, Lu HS, Kang MQ and
Lin TY: Effects and mechanism of downregulation of survivin
expression by RNA interference on proliferation and apoptosis of
lung cancer cells. Mol Med Rep. 5:917–922. 2012.PubMed/NCBI
|
|
15
|
Baker B, Eldrup A, Manoharan M, et al:
RNA-induced silencing complexes; RNA interference; gene expression
inhibition US patent 10/700,884. October 4–2003, issued February
17. 2005
|
|
16
|
Aliabadi H Montazeri, Landry B, Mahdipoor
P and Uludag H: Induction of apoptosis by survivin silencing
through siRNA delivery in a human breast cancer cell line. Mol
Pharm. 8:1821–1830. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Mirandola P, Sponzilli I, Gobbi G,
Marmiroli S, Rinaldi L, Binazzi R, Piccari GG, Ramazzotti G,
Gaboardi GC, Cocco L and Vitale M: Anticancer agents sensitize
osteosarcoma cells to TNF-related apoptosis-inducing ligand
downmodulating IAP family proteins. Int J Oncol. 28:127–133.
2006.PubMed/NCBI
|
|
19
|
Stewart B and Wild C: World cancer report
2014. International Agency for Research on Cancer; Lyon: 2014
|
|
20
|
Organization WH: ES Fact sheet 310: Top
causes of death. Centro De Prensa. 2014.
|
|
21
|
Arnold LD, Ji QS, Buck E, Haley JD and
Mulvihill MJ: Combination cancer therapy U.S. Patent 8,575,164.
2013
|
|
22
|
Yang Y and Huang S: Discuss the strengths
and weaknesses between tumour biotherapy and chemoradiotherapy.
Medical Information. 24:3503–3504. 2011.
|
|
23
|
Hengartner MO: The biochemistry of
apoptosis. Nature. 407:770–776. 2000. View
Article : Google Scholar : PubMed/NCBI
|
|
24
|
Reed JC: Dysregulation of apoptosis in
cancer. J Clin Oncol. 17:2941–2953. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Chiou SK, Jones MK and Tarnawski AS:
Survivin-an anti-apoptosis protein: Its biological roles and
implications for cancer and beyond. Med Sci Monit. 9:PI25–PI29.
2003.PubMed/NCBI
|
|
26
|
Merritt WM, Bar-Eli M and Sood AK: The
dicey role of Dicer: Implications for RNAi therapy. Cancer Res.
70:2571–2574. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Semple SC, Akinc A, Chen J, Sandhu AP, Mui
BL, Cho CK, Sah DW, Stebbing D, Crosley EJ, Yaworski E, et al:
Rational design of cationic lipids for siRNA delivery. Nat
Biotechnol. 28:172–176. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
28
|
Medarova Z, Pham W, Farrar C, Petkova V
and Moore A: In vivo imaging of siRNA delivery and silencing in
tumors. Nat Med. 13:372–377. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
29
|
Wenying Z, Zhaoning J, Zhimin Y, Dongyun C
and Lili S: Survivin siRNA inhibits gastric cancer in nude mice.
Cell Biochem Biophys. 62:337–341. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Rödel F, Hoffmann J, Distel L, Herrmann M,
Noisternig T, Papadopoulos T, Sauer R and Rödel C: Survivin as a
radioresistance factor, and prognostic and therapeutic target for
radiotherapy in rectal cancer. Cancer Res. 65:4881–4887. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Ning S, Fuessel S, Kotzsch M, Kraemer K,
Kappler M, Schmidt U, Taubert H, Wirth MP and Meye A:
siRNA-mediated down-regulation of survivin inhibits bladder cancer
cell growth. Int J Oncol. 25:1065–1071. 2004.PubMed/NCBI
|
|
32
|
Endoh A, Asanuma K, Moriai R, Yamada M,
Koyanagi Y, Sato T, Yagihasi A, Nakamura M, Kobayashi D and
Watanabe N: Expression of survivin mRNA in CD34-positive cells.
Clin Chim Acta. 306:149–151. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Lee GH, Joo YE, Koh YS, Chung IJ, Park YK,
Lee JH, Kim HS, Choi SK, Rew JS, Park CS and Kim SJ: Expression of
survivin in gastric cancer and its relationship with tumor
angiogenesis. Eur J Gastroenterol Hepatol. 18:957–963. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Smith SD, Wheeler MA, Plescia J, Colberg
JW, Weiss RM and Altieri DC: Urine detection of survivin and
diagnosis of bladder cancer. JAMA. 285:324–328. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Yao X, Liu F, Qi X, Wu B, Yin HL, Ma HH,
Shi QL, Zhou XJ and Li JS: Expression of survivin in human gastric
adenocarcinomas: Correlation with proliferation and apoptosis.
Zhonghua Wai Ke Za Zhi. 42:145–148. 2004.(In Chinese). PubMed/NCBI
|