Introduction
Hematopoietic stem cell (HSC) transplantation is a
potentially life-saving procedure used to treat a broad spectrum of
disorders, including hematological, immune and genetic diseases
(1). It has been demonstrated that
bone marrow reconstituting HSCs reside within a small subpopulation
of bone marrow or blood-derived mononuclear cells that express the
surface antigen cluster of differentiation (CD)34. The efficacy of
cord blood (CB) transplantation is limited by the low cell dose
available. Low cell doses at transplant are correlated with delayed
engraftment, prolonged neutropenia and thrombocytopenia and
elevated risk of graft failure. The successful ex vivo
culture and amplification of blood-derived CD34+
progenitor cells offers the possibility of HSC transplantation
(2). CB is used as an alternative
for bone marrow or mobilized peripheral blood grafts, particularly
when no matched human leukocyte antigen-related or unrelated donors
are available (3). Under similar
conditions, recipients of CB transplants exhibit a lower incidence
of acute and chronic graft-versus-host disease compared with
recipients of bone marrow transplants (4). The use of CB as a source of HSCs
utilized for transplantation has increased and >3,000 CB
transplants are conducted annually (5).
Allogeneic transplantation with human umbilical cord
blood (hUCB) in adult recipients is mainly limited by a low
CD34+ cell dose (6).
Thus, CBT is generally only used in children and low-weight adults.
Multiple strategies have been investigated to try to overcome these
limitations, one of which involves the ex vivo expansion of
CB units prior to transplantation (7). Previous studies have demonstrated that
HSCs may expand, suggesting that in vitro HSCs should be
exposed to specific factors and signals that promote self-renewal
and amplification (8–10). Furthermore, it has been demonstrated
that cell survival and proliferation in vitro may be
efficiently stimulated by several cytokines, particularly stem cell
growth factor and thrombopoietin (TPO) (11). It has also been indicated that the
fate of HSCs may be chemically modulated by adding small biological
and chemical molecules to the culture media in vitro to
induce cell survival and division, while simultaneously preventing
stem cell differentiation (1). Small
molecules, including all-trans retinoic acid copper chelator,
tetraethylenepentamine, prostaglandin E2 and
6-bromoindirubin-3′-oxime (BIO) all serve a role in HSC (12); for instance, all-trans retinoic acid
serves a role in stem cell differentiation to several lineages,
including myeloid differentiation, and BIO is the first
pharmacological agent demonstrated to maintain self-renewal in
embryonic stem cells (12).
The present study evaluated the effects of three
small-molecule steroid hormones, testosterone, norepinephrine and
epinephrine, on HSCs. As HSCs all express the surface antigen CD34,
CD34+ cells were selected for subsequent
experiments.
Oxygen concentration is an important influence on
the growth of HSCs. In vivo, HSCs are found in
microenvironments, known as niches, in the bone marrow. Throughout
the bone marrow, physiological oxygen concentrations are <4% and
almost 0% in certain areas (13). It
has been hypothesized that the hypoxic environment maintains the
characteristics of HSCs and numerous in vitro studies
investigating the cultivation of HSCs under hypoxic conditions have
been performed (14–17). Thus, the present study included
hypoxia as a condition in the study design, in order to determine
the effects of the three small molecules on CD34+ cell
amplification under hypoxic conditions.
Materials and methods
Collection and purification of
CD34+ cells
Human CB (n=12; male newborns) was obtained from
mothers undergoing full-term deliveries between January and May
2015 in the Department of Obstetrics in Qilu Hospital of Shandong
University (Jinan, China) after informed written consent was
obtained. Maternal age was between 20 to 40 years old (mean, 26±2),
and there was no history of acute, chronic or infectious disease,
neonatal apnea, edema or jaundice. The present study was approved
by the Ethics Committee of Qilu Hospital of Shandong University.
Within 12 h of harvesting the CB. Mononuclear cells (MNCs) were
separated using Ficollpaque medium (density 1.077±0.001 g/ml;
HaoYang company, Tianjin, China) and centrifuging at 1,726 × g at
12°C for 25 min. Isolated MNCs were collected and washed twice in
RPMI 1640 (Gibco, Los Angeles, USA) plus 5% fetal bovine serum
(FBS; Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA).
Cord blood MNCs were incubated with 100 µl of CD34+
micro beads (Miltenyi Biotec GmbH, Bergisch Gladbach, Germany; cat.
no. 130046703) at 4°C for 30 min. Cells were subsequently passed
through an LS MACS column (Miltenyi Biotec GmbH) and enriched
CD34+ cells were collected in 15 ml tubes by flushing
the column. Cells were subsequently suspended in 0.1 M phosphate
buffered saline (PBS; pH 7.4). The purity of CD34+ cells
was detected using a fluorescence-activated cell sorting system
(Guava easyCyte8HT; EMD Millipore, Billerica, MA, USA) and data
were analyzed with Guava Incyte version 2.8 (EMD Millipore).
Hormone screening test
CD34+ cells at a density of
2×103 cells/well were seeded into 24-well plates with
methylcellulose semisolid medium (MethoCult™ GF H4434)
supplemented with stem cell factor (SCF), granulocyte-macrophage
colony-stimulating factor, erythropoietin and interleukin (IL)-3
(all Stem Cell Technologies, Inc., Vancouver, BC, Canada) and used
to perform colony-forming unit (CFU) assays following the
manufacturer's instructions. Cells were cultured under either
normoxic or hypoxic conditions. CD34+ cells were divided
into control, testosterone (4.6×10−8 mol/l; XianJu
company, Zhejiang, China), norepinephrine (5.9×10−5
mol/l; ShuangHe Company, Beijing, China) and epinephrine groups
(2.7×10−6 mol/l; YongKang company, Beijing, China). The
control groups were treated without hormones and at the same oxygen
concentration as other groups. The concentrations of these hormones
were determined as previously described (18–23).
Cells were incubated in an atmosphere containing 20% O2
(normoxic conditions) using a Heal Force Tris-gas incubator (HF240;
Heal Force Bio-meditech Holdings Limited, Shanghai, China) or 1%
O2 (hypoxic conditions; HF100; Heal Force Bio-meditech
Holdings Limited) containing 5% CO2 at 37°C. Following 2
weeks culture, the number and type of CFUs were determined using an
inverted microscope (IX71 Olympus Inverted Microscope; Olympus
Corporation; Tokyo, Japan). The types of colonies identified
included colony-forming units-erythroid (CFU-E), burst-forming
unit-erythroid (BFU-E), colony-forming unit-granulocyte/macrophage
(CFU-GM) and colony forming units-mixed (CFU-Mix; >50 cells) as
previously described (24).
Preparation of feeder
Umbilical cord tissue was also obtained from the
healthy donor mothers who donated CB for the present study.
Informed written consent was received. Umbilical cords were
dissected following thorough washing and blood vessels were
removed. Small fragments (1–2 mm3) were cut and placed
in plates containing low glucose-Dulbecco's modified Eagle's medium
(L-DMEM) supplemented with 10% fetal bovine serum, 100 U/ml
penicillin and 100 µg/ml streptomycin (All Gibco; Thermo Fisher
Scientific, Inc.). Cultures were maintained at 37°C in a humidified
atmosphere containing 5% CO2, as previously described
(25,26). The medium was replenished every 3–4
days. Following 7 days culture, adherent cells were observed
growing from the individual tissue explants. Adherent
fibroblast-like cells became confluent following 2–3 weeks culture.
Subsequently, cells were treated with 0.25% trypsin (Gibco; Thermo
Fisher Scientific, Inc.) and passaged at 1×104
cells/cm2 in L-DMEM. Cells at the 5 and 7th passage were
used following γ-irradiation at a dose of 15 Gy following a
previously described protocol (25).
A total of 5×105 umbilical cord-mesenchymal stem cells
were seeded in a 25-cm2 culture flask and served as the
feeder layer for subsequent experiments.
Co-cultivation of CD34+
cells with feeder
CB CD34+ cells (1.1×105
cells/ml) were co-cultured with feeder in HSC expansion medium
(Stem Cell Technologies, Inc.) supplemented with 10% fetal bovine
serum (Gibco; Thermo Fisher Scientific, Inc.), 70 ng/ml SCF, 30
ng/ml IL-3, 30 ng/ml FMS-like tyrosine kinase 3 ligand, 20 ng/ml of
IL-6, 20 ng/ml bone morphogenetic protein-2 and 20 ng/ml TPO (all
R&D Systems, Inc., Minneapolis, MN, USA). Cultures were
maintained at 37°C in an atmosphere containing 5% CO2
for 7 days. Cytokine concentrations were determined as described
previously (11,27). CD34+ cells were divided
into 4 groups: i) A normoxia testosterone group, consisting of a
co-culture of CD34+ cells and feeder plus testosterone
undecanoate (4.6×10−8 mol/l) in 20% O2; ii) a
normoxia control group, consisting of co-culture of
CD34+ cells and feeder without hormone in 20%
O2; iii) a hypoxia testosterone group, consisting of a
co-culture of CD34+ cells and feeder plus testosterone
undecanoate in 1% O2 and iv) a hypoxia control group,
consisting of a co-culture of CD34+ cells and feeder
without hormone in 1% O2. The medium was replenished
every 3–4 days. On day 7, the total cell suspensions were harvested
for use in subsequent experiments.
CFU assay following liquid
culture
CD34+ cells were amplified 7 days after
co-cultivation with feeder and were seeded into 24-well plates in
the MethoCult GF H4434 medium. The cells were seeded at a density
of 2×103 cells/well following the manufacturer's
instructions for the CFU assay. Each cell group was plated in 6
replicate wells and cultures were maintained at 37°C in a
humidified atmosphere with 5% CO2 and 20% O2.
Following 2 weeks culture, the number and type of CFUs were
determined using an inverted microscope (magnification, ×20; IX71
Olympus Inverted Microscope; Olympus Corporation).
Immunophenotypic analysis
Following co-culture of CD34+ cells for 7
days, cells were confirmed by four-color flow cytometry using a
fluorescence-activated cell sorting Calibur analyzer (Guava Cyte
8HT; EMD Millipore). The cells (1×106) were suspended in
100 µl PBS containing fluorescein isothiocyanate-conjugated
anti-CD34 antibody 10 µl (BioLegend, San Diego, USA; cat. no.
343604), phycoerythrin-conjugated anti-CD71 (BD Biosciences, San
Jose, CA, USA; cat. no. 560981), and phycoerythrin-71-conjugated
anti-CD38 antibodies (BD Biosciences; cat. no. 555537; 1:10) for 15
min at room temperature, following the manufacturer's instructions.
Cells were subsequently washed with PBS and the data was examined
using Guava Incyte (Version 2.8, EMD Millipore).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
CD34+ cells were underwent RT-qPCR
following 7 days co-culture in vitro. Total RNA was
extracted using TRIzol reagent (Ambion; Thermo Fisher Scientific,
Inc.) and cDNA was synthesized using the ReverTra Ace QPCR RT
Master mix kit (Toyobo Co., Ltd., Osaka, Japan) according to the
manufacturer's protocol. Levels of homeobox (HOX)A9, HOXB2, HOXB4,
HOXC4, BMI1, GATA-1, C-MYB, HOXB6, NFE2 and hypoxia inducible
factor α (HIF-1α) were analyzed using qPCR (95°C for 1 min, 95°C
for 15 sec, and 60°C for 1 min for a total of 40 cycles) on an ABI
500 PCR system (Applied Biosystems; Thermo Fisher Scientific, Inc.)
with SYBR green I dye (Toyobo Co., Ltd.). Table I presents the primer sequences used
in RT-qPCR and all primers were purchased from BioSun Technology
Co. Ltd. (Shanghai, China). GAPDH was used as an internal control.
The expression of each gene was determined using the
2−ΔΔCq method (28) and
data were analyzed using Sequence Detection software (version 1.4;
Applied Biosystems; Thermo Fisher Scientific, Inc.). The expression
of mRNA is presented as the fold difference with respect to the
untreated control groups and the control group values were set at a
fold change equal to one.
 | Table I.Primer sequences used for reverse
transcription-quantitative polymerase chain reaction. |
Table I.
Primer sequences used for reverse
transcription-quantitative polymerase chain reaction.
| Gene | Direction | Primer sequence
(5′-3′) |
|---|
| HOXB4 | Forward |
AGCACGGTAAACCCCAATTACG |
|
| Reverse |
GTGTCAGGTAGCGGTTGTAGTG |
| HOXB6 | Forward |
TCGTGCAACAGTTCCTCCTT |
|
| Reverse |
CGCGTCAGGTAGCGATTGTA |
| HOXA9 | Forward |
CCACGCTTGACACTCACACT |
|
| Reverse |
GGGTTATTGGGATCGATGGGG |
| GATA1 | Forward |
GACACTCCCCAGTCTTTCAGG |
|
| Reverse |
CAGTTGAGGCAGGGTAGAGC |
| NFE2 | Forward |
ACTCTGGCCCAGTAGGATGT |
|
| Reverse |
TTGGAGCATTCAGACCCTGC |
| HIF-1α | Forward |
TTCCTTCTCTTCTCCGCGTG |
|
| Reverse |
AACTTATCTTTTTCTTGTCGTTCGC |
| HOXB2 | Forward |
CTAGCCTACAGGGTTCTCTC |
|
| Reverse |
CACAGAGCGTACTGGTGAAAAA |
| BMI-1 | Forward |
TGGACTGACAAATGCTGGAGA |
|
| Reverse |
GAAGATTGGTGGTTACCGCTG |
| C-MYB | Forward |
GAGGTGGCATAACCACTTGAA |
|
| Reverse |
AGGCAGTAGCTTTGCGATTTC |
| HOXC4 | Forward |
GCACCGTCAAGGCTGAGAAC |
|
| Reverse |
TGGTGAAGACGCCAGTGGA |
| GAPDH | Forward |
GCACCGTCAAGGCTGAGAAC |
|
| Reverse |
TGGTGAAGACGCCAGTGGA |
Statistical analysis
Data were analyzed using SPSS software version 14.0
(SPSS Inc., Chicago, IL, USA). Quantitative data are presented as
the mean ± standard deviation. Two-way analysis of variance with
Fisher's least significant difference as a post hoc analysis was
used for comparisons among multiple groups. P<0.05 was
considered to indicate a statistically significant difference.
Results
Effects of hormone and oxygen
concentrations on CFU
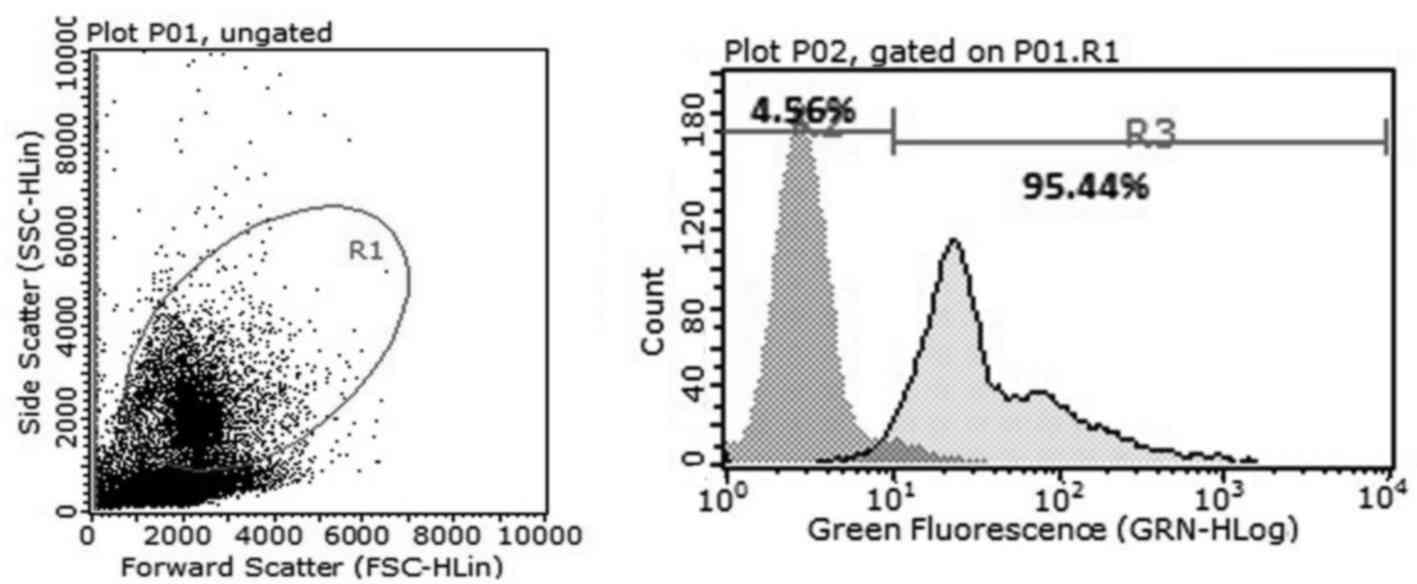
Magnetic activated cell sorting and flow cytometric
analysis demonstrated that >95% cells expressed CD34+
(Fig. 1). Subsequently, the effects
of testosterone, norepinephrine and epinephrine on the colony
formation function of CD34+cells were investigated. The
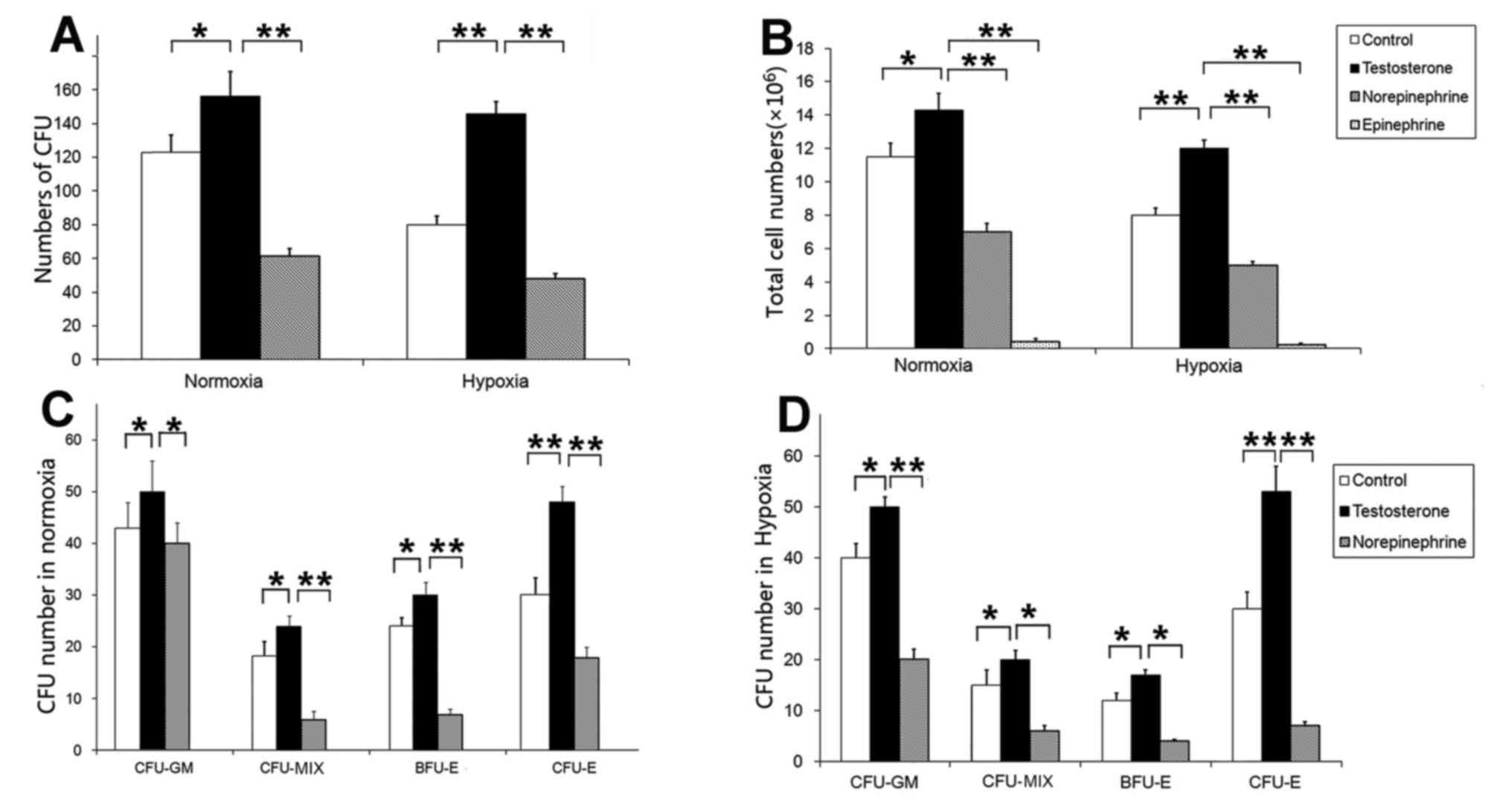
colony number of the testosterone groups were significantly
increased compared with the control and norepinephrine groups under
normoxia (P<0.05 and P<0.01, respectively) and hypoxia (both
P<0.01; Fig. 2A). There was no
evidence of colony formation in the epinephrine group under
normoxic and hypoxic conditions, thus the epinephrine group was
excluded from the analysis. In addition, the total number of cells
in the testosterone group was significantly increased compared with
the control, norepinephrine and epinephrine groups under normoxia
(P<0.05, P<0.01 and P<0.01 respectively) and hypoxia (all
P<0.01; Fig. 2B).
Regarding colony types, the presence of testosterone
significantly increased the number of different CFUs (Fig. 2C). In the testosterone groups, under
normoxic conditions the number of CFU-GMs was significantly
increased (50±8) compared with the control group (43±6; P<0.05)
and the norepinephrine group (39±4; P<0.05). Furthermore, the
number of CFU-MIXs was significantly increased (22±2) compared with
the control group (18±3; P<0.05) and the norepinephrine group
(5±1; P<0.01). In addition, the number of BFU-Es was
significantly increased (30±4) compared with the control group
(26±5; P<0.05) and the norepinephrine group (6±1; P<0.01).
The number of CFU-Es was significantly increased (48±5) compared
with the control group (30±6; P<0.01) and the norepinephrine
group (18±2; P<0.01; Fig. 2C). In
the testosterone groups under hypoxic conditions, the number of
CFU-GM colonies was significantly increased (50±2) compared with
the control group (40±4; P<0.05) and the norepinephrine group
(20±2; P<0.01). The number of CFU-MIX colonies was significantly
increased (19±2) compared with the control group (18±3; P<0.05)
and the norepinephrine group (5±2; P<0.05), and the number of
BFU-E colonies was significantly increased (17±2) compared with the
control group (14±3; P<0.05) and the norepinephrine group (3±1;
P<0.05). Similarly, the number of CFU-E colonies was
significantly increased (54±8) compared with the control group
(28±5; P<0.01) and the norepinephrine group (5±1; P<0.01;
Fig. 2D). The results demonstrated
that norepinephrine and epinephrine may inhibit cell amplification
under normoxic and hypoxic conditions and were therefore not used
in subsequent experiments.
Expansion of CD34+ cells
and CFU assay
Fresh enriched CB CD34+ cells were
co-cultured for 7 days with feeder. Under normoxic conditions,
during the first 3 days the cells rapidly entered the logarithmic
growth period (testosterone group, 41.53±4.31×105;
control group, 30.65±2.74×105; P>0.05), whereas cells
under hypoxia grew more slowly (testosterone group,
15.22±3.41×105; control group,
13.37±2.10×105; P>0.05; Fig. 3A and B). However, there was an
acceleration in the growth of cells in the hypoxia groups after the
first 3 days (testosterone group, 118.44±17.72×105;
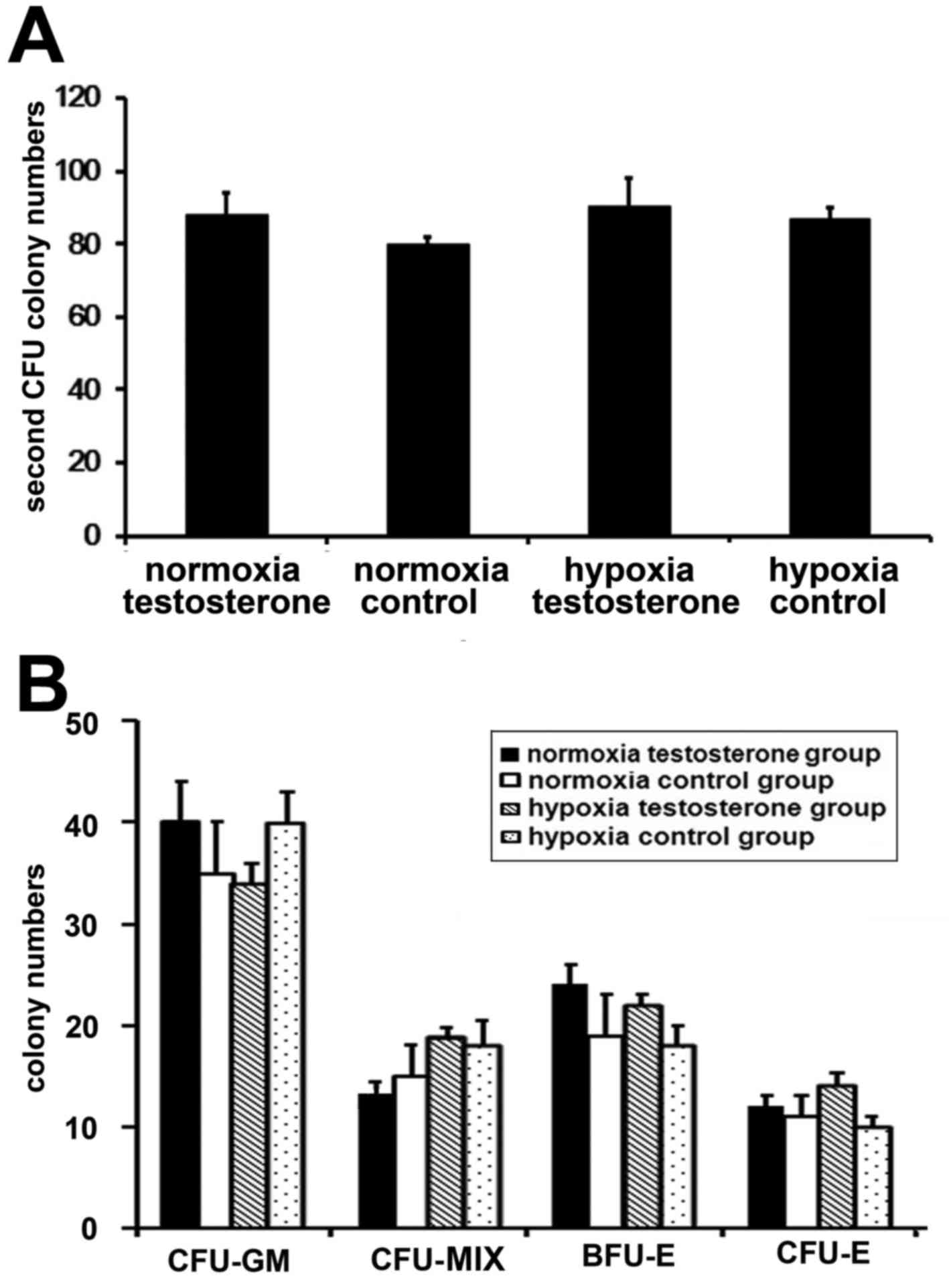
control group, 79.04±10.54×105; P<0.05 Fig. 3B). Following 7 days culture, total
cell numbers in the two normoxia groups were significantly
increased compared with the two hypoxia groups (P<0.05; Fig. 3C). In addition, the highest
amplification efficiency of the cells was observed in the normoxia
testosterone group. Subsequently, the expanded CD34+
cells were cultured in methylcellulose medium for the CFU assay to
observe the differential potential of HPCs. No significant
differences were observed in the number (P>0.05; Fig. 4A) and type (P>0.05; Fig. 4B) of CFUs among the 4 groups.
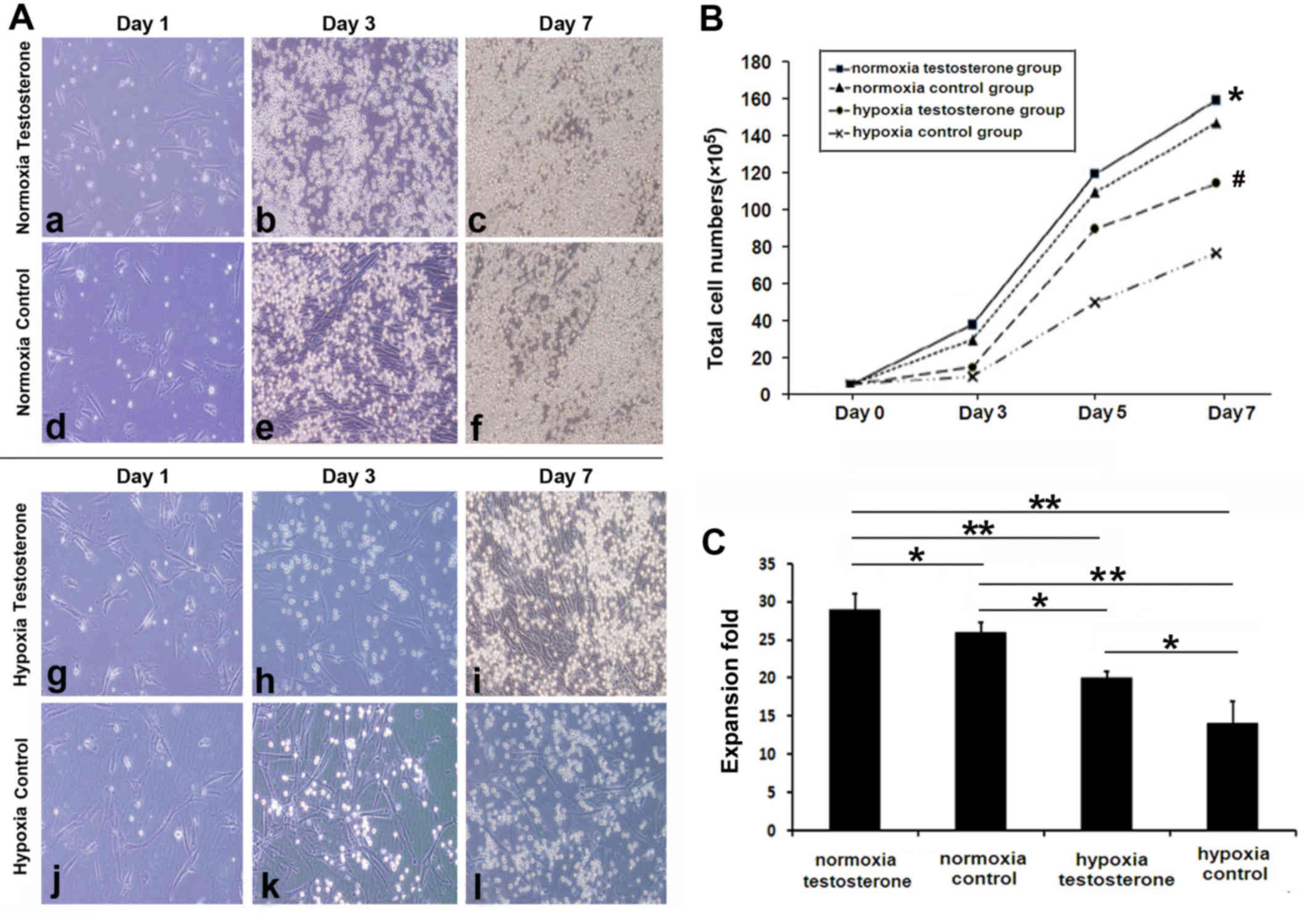 | Figure 3.Cluster of differentiation
CD34+ cell expansion with cytokine cocktail and feeder
(magnification, ×200). (A) Cells in the (a-f) normoxia or (g-l)
hypoxia testosterone and control groups were cultured for (a,d,g,l)
1, (b,e,h,j) 3 and (c,f,i,l) 7 days. Cell proliferation was
markedly higher in the normoxia groups compared with the hypoxia
groups and addition of testosterone promoted cell expansion in the
normoxia and hypoxia groups. (B) Comparison of the total cell
growth rate among all groups. The amplification efficiency was
significantly increased in the normoxia groups compared with the
hypoxia groups and the presence of testosterone increased cell
expansion, particularly during the final 4 days under hypoxia.
*P<0.05 vs. normoxia control, hypoxia testosterone and hypoxia
control groups. #P<0.05 vs. hypoxia control group.
(C) Comparison of total cell numbers. Cell numbers were
significantly increased in the normoxia groups compared with the
hypoxia groups. The addition of testosterone increased
amplification in normoxia and hypoxia groups. Data are presented as
the mean + standard deviation. *P<0.05 and **P<0.01. |
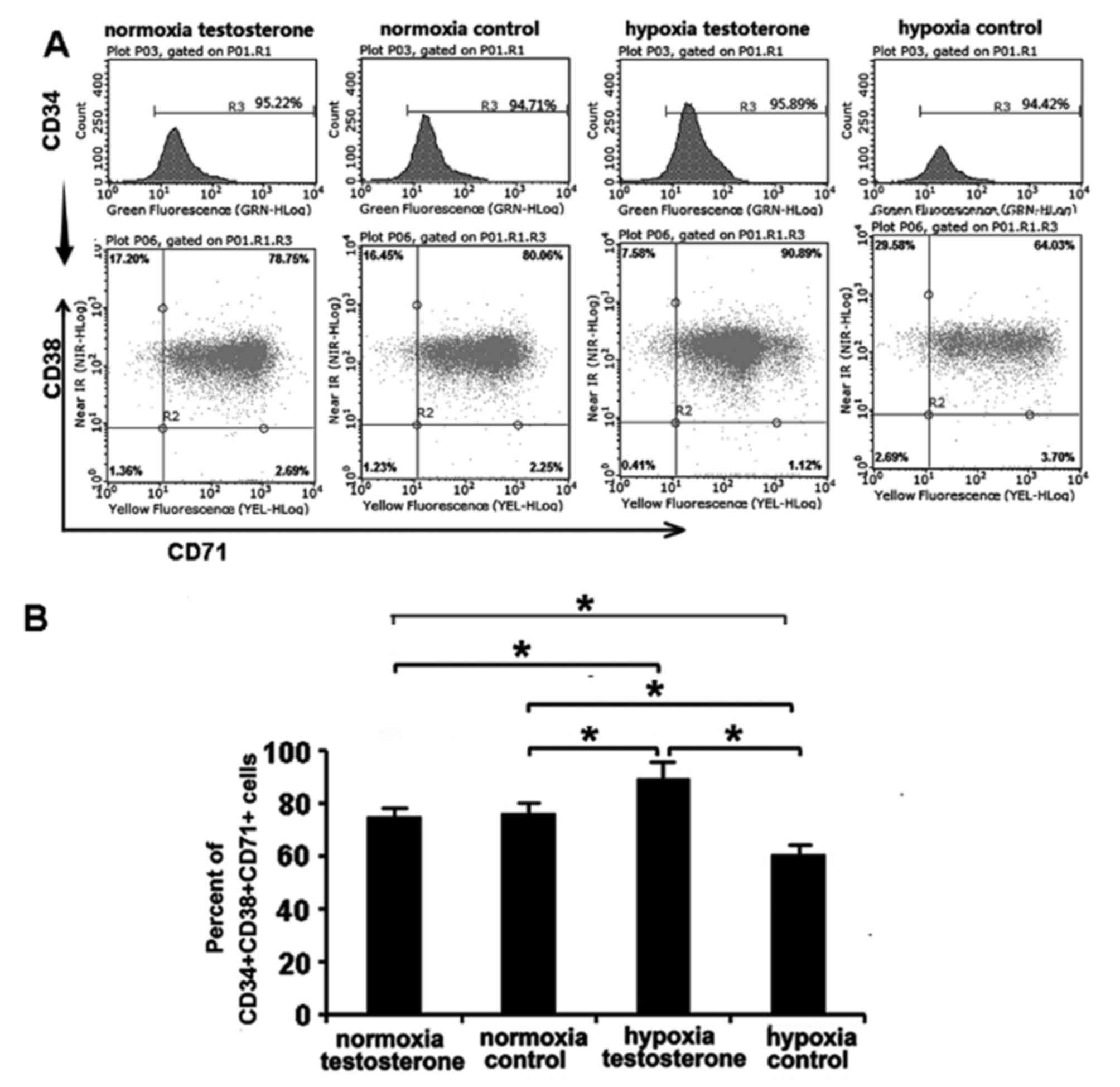
Analysis of cell phenotypes
Following culture of CD34+ cells for 7
days, CD34, CD38 and CD71 antigen phenotypes were analyzed using
flow cytometry (Fig. 5). CD34 and
CD38 are the surface markers currently used to identify HSC/HPC and
CD71 is the surface marker of erythroid progenitor cells (29,30). The
results demonstrated that the proportion of
CD34+CD38+CD71+ cells in the
normoxia testosterone and control groups were similar (74.98±8.79%
vs. 75.82±9.50%, respectively; P>0.05; Fig. 5). However, there were significant
differences between the hypoxia testosterone and control groups
(87.15±10.13% vs. 60.45±6.58%; P<0.05; Fig. 5). Furthermore, the proportion of
CD34+CD38+CD71+ cells in the
hypoxia testosterone group was significantly higher compared with
the other groups (87.15±10.13% vs. 74.98±8.79%, 75.82±9.50% and
60.45±6.58%; P<0.05). The proportion of
CD34+CD38+CD71+ cells in the
hypoxia control group was lowest compared with the other groups
(60.45±6.58%; P<0.05; Fig. 5).
These results suggest that hypoxia may be beneficial in maintaining
CD34+ cells in an undifferentiated state and the
addition of testosterone may promote the differentiation of
CD34+ cells into erythroid hematopoietic progenitor
cells (HPCs).
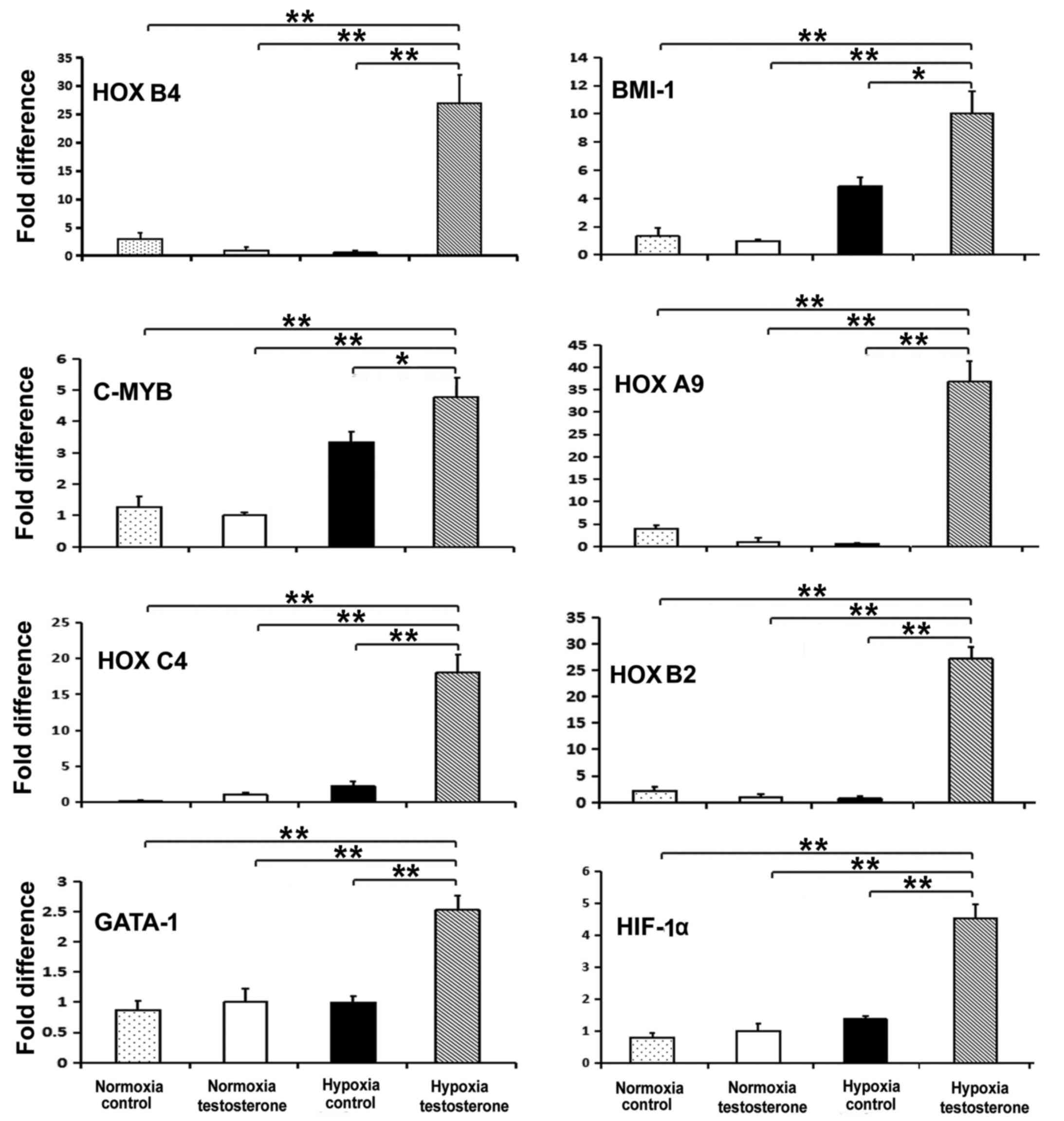
Expression of hematopoiesis-related
genes
RT-qPCR was performed to detect the expression of
the stem cell-specific genes HOXB4, BMI-1 and C-MYB, the
erythroid-specific genes HOXB2 and HOXB6, the lymphocyte
lineage-related gene HOXC4, the granulocyte lineage-related gene
HOXA9, the megakaryocyte lineage-related genes GATA-1 and NFE2, and
the hypoxia-related gene HIF-α to determine the properties of
self-renewal and multi-differentiation of CD34+ cells.
Following 7 days culture, the expression of the HOXA9, HOXB2,
HOXB4, BMI1, HOXC4, GATA-1, HIF-1α and C-MYB genes were
significantly increased in the hypoxia testosterone group compared
with all other groups (P<0.05; Fig.
6). However, the expression of HOXB6 and NFE2 did not
significantly differ among the hypoxia control group, normoxia
testosterone group and normoxia control group (data not shown).
Discussion
To promote efficient HSC proliferation and to mimic
the microenvironment in the bone marrow, small molecules were
introduced in the culture medium. In the present study,
testosterone, norepinephrine and epinephrine small-molecule steroid
hormones were selected and the effects of these hormones on
CD34+ cells were investigated. The results demonstrated
that the number of CFUs and total cells in the testosterone group
were significantly increased under normoxic and hypoxic conditions
compared with the corresponding control groups. Furthermore, the
results indicated that norepinephrine and epinephrine significantly
inhibited colony formation and cell amplification under normoxic
and hypoxic conditions. As a result, testosterone was selected for
use in subsequent experiments.
It has been reported that androgens significantly
reduce the quiescence ratio and promote HSC proliferation (31). Previous studies have also
demonstrated that testosterone significantly enhances colony
formation and the expansion of HSCs (32–34). Kim
et al (20) revealed that
androgens exhibit a modest growth- and survival-enhancing effect on
CFU-E, but not on CFU-GM or BFU-E. Similarly, the present study
demonstrated that testosterone significantly increased the number
of different types of CFUs, particularly CFU-E, under normoxic and
hypoxic conditions. These results indicate that androgens have an
effect on HPC that is restricted to mature erythroid progenitors.
Norepinephrine and epinephrine also affected HSCs. Norepinephrine
signaling controls HSC/HPC mobilization and the sympathetic nervous
system to regulate the attraction of stem cells to their niche
(35). However, increasing the
concentrations of norepinephrine and epinephrine may reduce HSC
cloning, thereby inhibiting hematopoiesis (18,23,30).
Previous studies have reported that hypoxia is
beneficial in maintaining the self-renewal properties of HSCs
(36–38). Furthermore, a number of studies have
suggested that hypoxia may promote HSC amplification (14,39,40).
Ivanovic et al (14)
identified that 3% was the lowest O2 concentration that
resulted in the same rate of colony-forming cell expansion as when
O2 concentration was 20%. In the present study, the
number of CFUs and total cells in all hypoxia groups were
significantly decreased compared with those in the normoxia groups.
These results were consistent with the results of a study by
Eliasson et al (16).
However, in the present study, the addition of testosterone
promoted CD34+ cell amplification in hypoxia and
normoxia groups, indicating that hypoxia and testosterone may be
important factors regulating the growth of CD34+
cells.
The results of the present study suggested that in
the normoxia groups, CD34+ cells rapidly entered the
logarithmic growth period within the first 3 days in liquid
culture. Cell amplification in the normoxia testosterone group was
increased, whereas cells grew slowly in the hypoxia groups.
Notably, cell amplification in the hypoxia testosterone groups
gradually accelerated between days 3 and 7 in culture. Although
hypoxia was not beneficial to CD34+ cell proliferation,
treatment with testosterone promoted CD34+ cell
proliferation in normoxic and hypoxic conditions. Flow cytometry
determined that the proportion of
CD34+CD38+CD71+cells was lowest in
the hypoxic control group, indicating that hypoxic conditions were
beneficial in maintaining the characteristics of CD34+
cells and delaying differentiation. Additionally, the highest
proportion of CD34+CD38+CD71+
cells was detected in the hypoxic testosterone group. These results
suggest that a combination of hypoxia and testosterone may promote
the differentiation of CD34+ cells into erythroid HPCs.
To further identify the ability of hematopoietic reconstitution, a
CFU assay was performed. A variety of colonies formed,
demonstrating that the multi-differentiation ability of the cells
was unaffected by hypoxia or testosterone.
The expression of hematopoiesis-related genes in
CD34+ cells was detected using RT-qPCR and it was
determined that there were significant differences in the levels of
gene expression between the hypoxia testosterone group and the
other groups. In the hypoxia testosterone group, the expression of
hematopoiesis-related genes, including HSC-specific and
differentiated, erythroid-specific, lymphocyte lineage-related
granulocyte lineage-related, megakaryocyte lineage-related and
hypoxia-related genes, were significantly higher compared with all
other groups. This increase in the expression of genes may have
been caused in part by the differentiation of CD34+
cells into HPCs. The results of the secondary CFU assay and flow
cytometry suggested that an increased number of
CD34+cells were differentiated into erythroid HPCs under
the combined effects of testosterone and hypoxia. This indicates
that the combination of hypoxia and testosterone was advantageous
in promoting the expression of hematopoietic genes.
Androgens promote the amplification of HSCs but
cannot effectively maintain the self-renewal characteristics of
stem cells. Huang et al (41)
demonstrated that bone marrow mesenchymal stem cells in androgen
receptor-knockdown mice exhibited enhanced self-renewal ability.
Nilutamidie is an anti-androgen agent that blocks the effects of
androgen and promotes the self-renewal of ESCs (42). Given the common characteristics of
stem cells, it was speculated that androgens may promote HSC
proliferation rather than maintain the self-renewal of HSCs.
Hypoxia (1%) is conducive to maintaining the undifferentiated HSC
state but does not promote amplification (16). The results of the present study
demonstrated that a combination of hypoxia and testosterone in
vitro may increase CD34+ cell differentiation and
prolong the HPC stage. This phenomenon may be caused by a number of
mechanisms. Firstly, hypoxia may not be conducive to HSC expansion,
potentially due to the action of HIF-1α, which may decrease HSC
proliferation and block cells in the G0 phase (16,32). The
results of the present study indicated that there was a significant
increase in the expression of HIF-1α in the hypoxia testosterone
compared with the control groups. Another potential mechanism to
consider is that hypoxia may directly suppress the proliferation
and differentiation of erythroid progenitor/precursor cells. These
effects may be reduced or counter-balanced by increasing
erythropoietin (EPO) levels (43)
and one acknowledged mechanism involved in the hematological
effects of androgens is the promotion of erythroid progenitor
expansion by increasing EPO levels (44). Furthermore, androgen and androgen
receptor signals may promote the proliferation of HSCs/HPCs and
stimulate hematopoietic lineage differentiation (45). Another potential mechanism may be
that the addition of testosterone may enhance the G1-S transition
rate in the cell cycle and the survival of HSCs (32). Additionally, androgens may stimulate
telomerase-related gene expression and enzymatic activity in bone
marrow CD34+ cells and extend the lifespan of the
CD34+ stem/progenitor cells (46).
In conclusion, CB ex vivo expansion is a
promising approach to deliver high doses of cells and improve the
outcomes of CBT. Careful selection of optimal CB units for
transplantation may improve the efficiency of the source of HSCs
and HPCs in adult transplantation and reduce the cost of processing
(47). The present results may
provide a means to improve HSC/HPC culture conditions in
vitro. Future studies investigating this technique may require
a larger sample size, however the present study demonstrated that
the combination of hypoxia and androgen in vitro may be a
promising condition of cultivation.
Acknowledgements
The present study was supported by grants from the
Major State Basic Research Development Program (grant no.
2012CB966504), Jinan Natural Science Foundation (grant no.
201403010) and the Basic Scientific Fund of Shandong University
(grant no. 2014QLKY02).
References
|
1
|
Walasek MA, van Os R and de Haan G:
Hematopoietic stem cell expansion: Challenges and opportunities.
Ann N Y Acad Sci. 1266:138–150. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Möbest D, Mertelsmann R and Henschler R:
Serum-free ex vivo expansion of CD34(+) hematopoietic progenitor
cells. Biotechnol Bioeng. 60:341–347. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ballen KK, Gluckman E and Broxmeyer HE:
Umbilical cord blood transplantation: The first 25 years and
beyond. Blood. 122:491–498. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Rocha V, Wagner JE Jr, Sobocinski KA,
Klein JP, Zhang MJ, Horowitz MM and Gluckman E: Graft-versus-host
disease in children who have received a cord-blood or bone marrow
transplant from an HLA-identical sibling. Eurocord and
International Bone Marrow Transplant Registry Working Committee on
Alternative Donor and Stem Cell Sources. N Engl J Med.
342:1846–1854. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Foeken LM, Green A, Hurley CK, Marry E,
Wiegand T and Oudshoorn M: Monitoring the international use of
unrelated donors for transplantation: The WMDA annual reports. Bone
Marrow Transplant. 45:811–818. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Gao L, Chen X, Zhang X, Liu Y, Kong P,
Peng X, Liu L, Liu H and Zeng D: Human umbilical cord blood-derived
stromal cell, a new resource of feeder layer to expand human
umbilical cord blood CD34+ cells in vitro. Blood Cells,
Molecules and Diseases. 36:322–328. 2006. View Article : Google Scholar
|
|
7
|
Horwitz ME and Frassoni F: Improving the
outcome of umbilical cord blood transplantation through ex vivo
expansion or graft manipulation. Cytotherapy. 17:730–738. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Pawliuk R, Eaves C and Humphries RK:
Evidence of both ontogeny and transplant dose-regulated expansion
of hematopoietic stem cells in vivo. Blood. 88:2852–2858.
1996.PubMed/NCBI
|
|
9
|
Iscove NN and Nawa K: Hematopoietic stem
cells expand during serial transplantation in vivo without apparent
exhaustion. Curr Biol. 7:805–808. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Osawa M, Hanada K, Hamada H and Nakauchi
H: Long term lymp hematopoietic reconstitution by a single
CD34-low/negative hematopoietic stem cell. Science. 273:242–245.
1996. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Sauvageau G, Iscove NN and Humphries RK:
In vitro and in vivo expansion of hematopoietic stem cells.
Oncogene. 23:7223–7232. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Schugar RC, Robbins PD and Deasy BM: Small
molecules in stem cell self-renewal and differentiation. Gene
Therapy. 15:126–135. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Hermitte F, de la Grange Brunet P, Belloc
F, Praloran V and Ivanovic Z: Very low O2 concentration (0. 1%)
favors G0 return of dividing CD34+ cells. Stem Cells.
24:65–73. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ivanovic Z, Hermitte F, de la Grange
Brunet P, Dazey B, Belloc F, Lacombe F, Vezon G and Praloran V:
Simultaneous maintenance of human cord blood SCID-repopulating
cells and expansion of committed progenitors at low O2
concentration (3%). Stem Cells. 22:716–724. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ivanovic Z, Dello Sbarba P, Trimoreau F,
Faucher JL and Praloran V: Primitive human HPCs are better
maintained and expanded in vitro at 1 percent oxygen than at 20
percent. Transfusion. 12:1482–1488. 2000. View Article : Google Scholar
|
|
16
|
Eliasson P, Rehn M, Hammar P, Larsson P,
Sirenko O, Flippin LA, Cammenga J and Jönsson JI: Hypoxia mediates
low cell-cycle activity and increases the proportion of long-term
reconstituting hematopoietic stem cells during in vitro culture.
Experimental Hematology. 38:301–310. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Rehn M, Olsson A, Reckzeh K, Diffner E,
Carmeliet P, Landberg G and Cammenga J: Hypoxic induction of
vascular endothelial growth factor regulates murine hematopoietic
stem cell function in the low-oxygenic niche. Blood. 118:1534–1543.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Penn A, Mohr AM, Shah SG, Sifri ZC, Kaiser
VL, Rameshwar P and Livingston DH: Dose-response relationship
between norepinephrine and erythropoiesis: Evidence for a critical
threshold. J Surg Res. 163:85–90. 2010. View Article : Google Scholar
|
|
19
|
Kröpfl JM, Stelzer I, Mangge H, Pekovits
K, Fuchs R and Allard N: Exercise-induced norepinephrine decreases
circulating hematopoietic stem and progenitor cell colony-forming
capacity. PLoS One. 9:106–120. 2014. View Article : Google Scholar
|
|
20
|
Kim SW, Hwang JH, Cheon JM, Park NS, Park
SE, Park SJ, Yun HJ, Kim S and Jo DY: Direct and indirect effects
of androgens on survival of hematopoietic progenitor cells in
vitro. J Korean Med Sci. 20:409–416. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Beckman B and Fisher JW: Decreased
erythroidcolony-forming cell response of XTfm/Y mice to
testosterone and 5 betadihydrotestosterone. Endocrinology.
107:1587–1592. 1980. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Huang CK, Luo J, Lee SO and Chang C:
Concise review: Androgen receptor differential roles in
stem/progenitor cells including prostate, embryonic, stromal and
hematopoietic lineages. Stem Cells. 32:2299–2308. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chen C, Cao J, Song X, Zeng L, Li Z, Li Y
and Xu K: Adrenaline administration promotes the efficiency of
granulocyte colony stimulating factor-mediated hematopoietic stem
and progenitor cell mobilization in mice. Int J Hematol. 97:50–57.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Duchez P, Chevaleyre J, de la Grange
Brunet P, Vlaski M, Boiron JM, Wouters G and Ivanovic Z:
Cryopreservation of hematopoietic stem and progenitor cells
amplified ex vivo from cord blood CD34+ cells.
Transfusion. 53:2012–2019. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Dumont N, Boyer L, Émond H, Celebi-Saltik
B, Pasha R, Bazin R, Mantovani D, Roy DC and Pineault N: Medium
conditioned with mesenchymal stromal cell-derived osteoblasts
improves the expansion and engraftment properties of cord blood
progenitors. Exp Hematol. 42:741–752. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhuang Y, Li D, Fu J, Shi Q, Lu Y and Ju
X: Comparison of biological properties of umbilical cord-derived
mesenchymal stem cells from early and late passages:
Immunomodulatory ability is enhanced in aged cells. Mol Med Rep.
11:166–174. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Robinson S, Niu T, de Lima M, Ng J, Yang
H, McMannis J, Karandish S, Sadeghi T, Fu P and del Angel M: Ex
vivo expansion of umbilical cord blood. Cytotherapy. 3:243–250.
2005. View Article : Google Scholar
|
|
28
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2 (-Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Bock TA, Ziegler BL, Bühring HJ, Scheding
S, Brugger W and Kanz L: Characterization of purified and ex vivo
manipulated human hematopoietic progenitor and stem cells in
xenograft recipients. Ann N Y Acad Sci. 872:200–207. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Frascoli M, Proietti M and Grassi F:
Phenotypic analysis and isolation of murine hematopoietic stem
cells and lineage-committed progenitors. J Vis Exp. 65:pii: 3736.
2012.
|
|
31
|
Zhang QS, Benedetti E, Deater M, Schubert
K, Major A, Pelz C, Impey S, Marquez-Loza L, Rathbun RK, Kato S, et
al: Oxymetholone therapy of fanconi anemia suppresses osteopontin
transcription and induces hematopoietic stem cell cycling. Stem
Cell Reports. 4:90–102. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Gallien-Lartigue O: Differential effects
of external agents on the G1-S transit rate of murine pluripotent
hemopoietic stem cells (CFUs) after their release from G0. Stem
Cells. 2:218–228. 1982.PubMed/NCBI
|
|
33
|
Freedman MH and Saunders EF: Factors
affecting erythroid colony growth (CFU-E) from human marrow. Exp
Hematol. 5:250–253. 1977.PubMed/NCBI
|
|
34
|
Reissmann KR, Udupa KB and Kawada K:
Effects of erythropoietin and androgens on erythroid stem cells
after their selective suppression by BCNU. Blood. 44:649–657.
1974.PubMed/NCBI
|
|
35
|
Katayama Y, Battista M, Kao WM, Hidalgo A,
Peired AJ, Thomas SA and Frenette PS: Signals from the sympathetic
nervous system regulate hematopoietic stem cell egress from bone
marrow. Cell. 124:407–421. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Ivanović Z, Bartolozzi B, Bernabei PA,
Cipolleschi MG, Rovida E, Milenković P, Praloran V and Dello Sbarba
P: Incubation of murine bone marrow cells in hypoxia ensures the
maintenance of marrow-repopulating ability together with the
expansion of committed progenitors. Br J Haematol. 108:424–429.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Ivanović Z, Dello Sbarba P, Trimoreau F,
Faucher JL and Praloran V: Primitive human HPCs are better
maintained and expanded in vitro at 1 percent oxygen than at 20
percent. Transfusion. 40:1482–1488. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Ivanovic Z, Belloc F, Faucher JL,
Cipolleschi MG, Praloran V and Dello Sbarba P: Hypoxia maintains
and interleukin-3 reduces the pre-colony-forming cell potential of
dividing CD34(+) murine bone marrow cells. Experimental Hematology.
30:67–73. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Ishikawa Y and Ito T: Kinetics of
hemopoietic stem cells in a hypoxic culture. Eur J Haematol.
40:126–129. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Lu L and Broxmeyer HE: Comparative
influences of phytohemagglutinin-stimulated leukocyte conditioned
medium, hemin, prostaglandin E and low oxygen tension on colony
formation by erythroid progenitor cells in normal human bone
marrow. Exp Hematol. 13:989–993. 1985.PubMed/NCBI
|
|
41
|
Huang CK, Tsai MY, Luo J, Kang HY, Lee SO
and Chang C: Suppression of androgen receptor enhances the self
renewal of mesenchymal stem cells through helevated expression of
EGFR. Biochim Biophys Acta. 1833:1222–1234. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Chang CY, Hsuuw YD, Huang FJ, Shyr CR,
Chang SY, Huang CK, Kang HY and Huang KE: Androgenic and
antiandrogenic effects and expression of androgen receptor in mouse
embryonic stem cells. FertilSteril. 85:1195–1203. 2006.
|
|
43
|
Rogers HM, Yu X, Wen J, Smith R, Fibach E
and Noguchi CT: Hypoxia alters progression of the erythroid
program. Exp Hematol. 1:17–27. 2008. View Article : Google Scholar
|
|
44
|
Fried W and Morley C: Effects of
androgenic steroids on erythropoiesis. Steroids. 46:799–826. 1985.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Huang CK, Luo J, Lee SO and Chang C:
Concise review: Androgen receptor differential roles in
stem/progenitor cells including prostate, embryonic, stromal and
hematopoietic lineages. Stem Cells. 9:2299–2308. 2014. View Article : Google Scholar
|
|
46
|
Calado RT, Yewdell WT, Wilkerson KL, Regal
JA, Kajigaya S, Stratakis CA and Young NS: Sex hormones, acting on
the TERT gene, increase telomerase activity in human primary
hematopoietic cells. Blood. 114:2236–2243. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Lane TA: Umbilical cord blood grafts for
hematopoietic transplantation in adults: A cup half empty or half
full? Transfusion. 45:1027–1034. 2005. View Article : Google Scholar : PubMed/NCBI
|