Introduction
Nonalcoholic fatty liver disease (NAFLD) occurs in a
wide range of histopathological conditions, from asymptomatic
steatosis to severe nonalcoholic steatohepatitis (NASH) (1). The transition from steatosis to
steatohepatitis represents an important step in liver damage
progression, which eventually culminates in hepatic fibrosis and
cirrhosis (2). At present, the
underlying molecular mechanism that promotes this transition
remains unknown and no effective therapy for NASH is available.
A growing body of evidence suggests that neutrophil
infiltration into tissues is associated with the pathogenesis of
inflammatory conditions, including NASH (3,4). A
proteolytic enzyme that is produced by neutrophils, neutrophil
elastase (NE), was recognized as a factor contributing to liver
injury, and a potential target for treatment (5). A previous study by our group
demonstrated that neutrophils serve an essential role in the early
stages of NASH development via the production NE (6). Correspondingly, the serine protease
inhibitor α1-antitrypsin (α1-AT), which is an endogenous inhibitor
of NE, was identified to be dramatically reduced in patients with
NASH (7). A previous study also
identified that an imbalance between levels of NE and α1-AT in
obesity affects insulin sensitivity, inflammation and energy
expenditure (8).
Berberine (BBR) is a natural supplement that is
popular for its potent antimicrobial, antiprotozoal and
antitrachoma effects (9). Previous
studies have suggested that BBR exerts notable anticancer,
antioxidant and anti-inflammatory activities (10), in addition to hepatoprotective
effects in liver fibrosis (11,12).
The present study investigated whether BBR was able
to alleviate the development of NASH development in apolipoprotein
E-deficient (ApoE−/−) mice with experimentally induced
NASH. In addition, the underlying molecular mechanisms of the
effects of BBR in this model were examined.
Materials and methods
Experimental animals and diets
The present study utilized a total of 26 4-week-old
male C57BL/6 J ApoE−/− mice, weighing 16.30±0.45 g
(Model Animal Research Center of Nanjing University, Nanjing,
China). All mice were bred in a specific pathogen-free facility and
maintained in a 12-h light-dark cycle at a controlled temperature
(22±2°C) and humidity (55±5%) and fed ad libitum. The mice
were divided into the following three groups, which were given
different diets: Standard chow diet (SC group, n=8); high-fat
high-cholesterol diet (HFHC group, n=8); and HFHC diet supplemented
with BBR (BBR group, n=10). C57BL/6J ApoE−/− mice
receiving a HFHC diet serves as a typical model of obesity-induced
NASH (13). The chow diet from
Research Diets, Inc. (New Brunswick, NJ, USA; cat. no. D10012 G)
contained 15.8% fat, 20.3% protein, and 63.9% carbohydrate. The
HFHC diet feed (cat. no. D12079B; Research Diets, Inc.) contained
41% fat, 17% protein, 43% carbohydrate and 0.21% cholesterol. BBR
(Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) was administered at
a dosage of 0.2 g/kg daily, starting from the sixth week to the end
of the experiment. Food intake was measured 3 times/week and
individual body weight was measured weekly over a 12-week period.
At the end of the experiments, the animals fasted for 12 h, and
following anesthesia with 3% pentobarbital (Sigma-Aldrich; Merck
KGaA) and sacrifice via cervical dislocation. Blood samples were
rapidly obtained by retro-orbital sampling, as previously described
(14). Liver tissues were harvested
and samples were fixed with 4% formaldehyde at room temperature for
24 h, whilst the remaining liver was snap-frozen in liquid nitrogen
until required. All animal protocols were reviewed and approved by
the Ethics Committee of Hangzhou Normal University Affiliated
Hospital (Hangzhou, China).
Serum biochemical analyses
Serum alanine aminotransferase (ALT), aspartate
aminotransferase (AST), triglyceride and cholesterol levels were
measured using a Hitachi 7180 automatic biochemical analyzer
(Hitachi Ltd., Tokyo, Japan) according to the manufacturer's
protocol. Serum NE and α1-AT levels were examined using a Mouse
Neutrophil Elastase/ELA2 DuoSet ELISA or α 1 Antitrypsin ELISA kit
following the manufacturers' instructions (DY4517-05, R&D
Systems, Inc., Minneapolis, MN, USA; ab205088, Abcam, Cambridge,
UK).
Histopathological analysis
Formalin-fixed liver tissues were embedded in
paraffin, sectioned (5 µm thickness), and stained with hematoxylin
and eosin as previously described (15). Frozen liver sections were cut to 10
µm thickness, fixed with 10% paraformaldehyde for 5 min at room
temperature and stained with Oil Red O for 10 min at room
temperature. Histological scoring was performed by two specialists
blinded to the experimental design and data. Scoring ranges were as
follows: Degree of steatosis (0–3), lobular inflammation (0–3),
hepatocyte ballooning (0–2). In addition, the NAFLD activity score
(NAS) was scored according to the NASH clinical research network
scoring system (16).
Immunohistochemical staining
Immunohistochemical staining was performed as
previously described (6). The
primary antibodies used were as follows: Anti-NE (1:200; cat. no.
orb1614; Biorbyt Ltd., Cambridge, UK), anti-C-X-C chemokine
receptor type 4 (CXCR4; 1:50; cat. no. ab124824, Abcam) and
anti-C-X-C motif chemokine 12 (CXCL12; 1:50; cat. no. sc-28876;
Santa Cruz Biotechnology, Inc., Dallas, TX, USA). Samples were then
incubated with the anti-rabbit (1:2,000; cat. no. ab97051; Abcam)
secondary antibody for 1 h at room temperature under gentle
agitation. Samples were subsequently visualized under an Eclipse
80i light microscope (Nikon Corporation, Tokyo, Japan) and
representative images were captured for analysis.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) analysis
Hepatic mRNA levels were analyzed by RT-qPCR using
an ABI Prism 7900 HT Fast Real-Time PCR system (Applied Biosystems;
Thermo Fisher Scientific, Inc., Waltham, MA, USA). Total RNA was
isolated from liver tissues using TRIzol (Invitrogen; Thermo Fisher
Scientific, Inc.) followed by DNAse treatment to remove genomic
DNA. cDNA was synthesized using 2 µg of total RNA with
Superscript® II Reverse Transcriptase (Invitrogen; Themo
Fisher Scientific, Inc.) at 42°C for 50 min. Amplification
reactions were performed using a SYBR Select Master Mix
(AmpliTaq® Fast DNA Polymerase, Thermo Fisher
Scientific, Inc.) according to the manufacturer's protocol. PCR
conditions were as follows: 95°C for 10 min, followed by 40 cycles
at 95°C for 10 sec and 60°C for 1 min. PCR products were verified
by melt curve analysis (17). Mouse
β-actin or GAPDH were used as the reference genes to normalize for
differences in the amount of total RNA in each sample. Relative
gene expression levels were analyzed using the 2−ΔΔCq
method (18). PCR murine primers
were obtained from Invitrogen (Thermo Fisher Scientific, Inc.) and
their sequences were as follows: β-actin forward (F),
5′-CTGGCTCCTAGCACCATGAA-3′ and reverse (R),
5′-CGCAGCTCAGTAACAGTCCG-3′; GAPDH F, 5′-AACAGCAACTCCCACTCTTC-3′ and
R, 5′-CCTGTTGCTGTAGCCGTATT-3′; α1-AT F, 5′-GAGCATTGGCACAGCGTTTG-3′
and R, 5′-AAGCGATGGTTGGATGTCAGC-3′; CXCR4 F,
5′-CTACAGCAGCGTTCTCATCC-3′ and R, 5′-TTTCAGCCAGCAGTTTCCTT-3′;
CXCL12 F, 5′-CGACTTCTCAGGCAGGTGA-3′ and R,
5′-GCCATTCCCATAGCATTCAT-3′; phosphatidylinositol-3 kinase (PI3K) F,
5′-AATGTGCCCTCTTTCGTTGT-3′ and R, 5′-TGAATGGTGACTGGCTGACT-3′;
interleukin (IL)-1 F, 5′-CTCTTGCCTGTCATCCCAAC-3′ and R,
5′-ACCATCTGTTCCCAATACGG-3′; IL-6 F, 5′-CGGAGAGGAGACTTCACAGAG-3′ and
R, 5′-CATTTCCACGATTTCCCAGA-3′; IL-8 F, 5′-GGATGGGAACAACGATAGGA-3′
and R, 5′-AGAACGTGGCGGTATCTCTG-3′; protein kinase B (AKT) F,
5′-ACTCATTCCAGACCCACGAC-3′ and R, 5′-ACAATCTCCGCACCATAGA-3′;
nuclear factor-κB (NF-κB) F, 5′-GCTCAAGATCTGCCGAGTAAA-3′ and R,
5′-GTCCCGTGAAATACACCTCAA-3′; tumor necrosis factor (TNF)-α F,
5′-AAGGGAGAGTGGTCAGGTTG-3′ and R, 5′-TCTGTGAGGAAGGCTGTGC-3′;
phospholipase C-β (PLC-β) F, 5′-GCAGGTCCAAGTGTTGATTG-3′ and R,
5′-TTCTTCTCCGCTCAGGTAGC-3′; and monocyte chemoattractant protein-1
(MCP-1) F, 5′-TCTCTCTTCCTCCACCACCAT-3′ and R,
5′-GCTCTCCAGCCTACTCATTGG-3′. Each experiment was performed in
duplicate and repeated three times.
Immunoblotting
Liver tissue (60 mg) was homogenized in RIPA buffer
(P0013B; Beyotime Institute of Biotechnology, Haimen, China),
supplemented with protease inhibitor cocktail (Sigma-Aldrich; Merck
KGaA). Samples (50 µg protein per lane) were separated by 10%
SDS-PAGE and electrotransferred onto polyvinylidene difluoride
membranes. Following blocking with 5% nonfat milk, membranes were
probed with anti-NE antibody (1:200) overnight at 4°C. The
membranes were then incubated with the secondary antibody for 1 h
at room temperature, followed by SuperSignal™ West Pico
Chemiluminescent detection (34077; Thermo Fisher Scientific, Inc.).
Protein band intensities were quantified using Quantity One
software 4.62 (Bio-Rad Laboratories, Inc., Hercules, CA, USA).
NE imaging
The Neutrophil Elastase 680 FAST imaging agent
(PerkinElmer, Inc, Waltham, MA, USA) was used to detect NE in
vivo at the end of the experiment according to the
manufacturer's protocol. This agent is based on a peptide
(PMAVVQSVP) that is specifically recognized by NE. Mice were
administered with 4 nmol of the NE imaging agent intravenously via
the tail vein, and 4 h later an in vivo imaging system
(Molecular Imaging software version 5.3.5; Carestream, CA, USA) was
used to capture the fluorescence and detect NE activity.
Statistical analyses
Statistical analysis was conducted using GraphPad
Prism (version 5; GraphPad Software, Inc., La Jolla, CA, USA). All
data are presented as the mean ± standard deviation. The
statistical significance of differences between groups was assessed
using an unpaired Student's t-test. P<0.05 was considered to
indicate a statistically significant difference.
Results
BBR has a hepatoprotective role
The effect of BBR on weight gain and liver metabolic
features in C57BL/6J ApoE−/− mice receiving a HFHC diet
was observed (Fig. 1). Mice fed a
HFHC diet had significantly increased body weights, associated with
hypertriglyceridemia and hypercholesterolemia, compared with
regular chow-fed mice in the SC group (P<0.05; Fig. 1A). This weight gain was also
accompanied by a significantly higher relative liver:body weight
ratio (liver index) in the HFHC group compared with the SC group
(P<0.01; Fig. 1B). Furthermore,
mice in the HFHC group had significantly enhanced increased ALT and
AST levels, indicative of liver injury, compared with the SC group
(P<0.01; Fig. 1E and F).
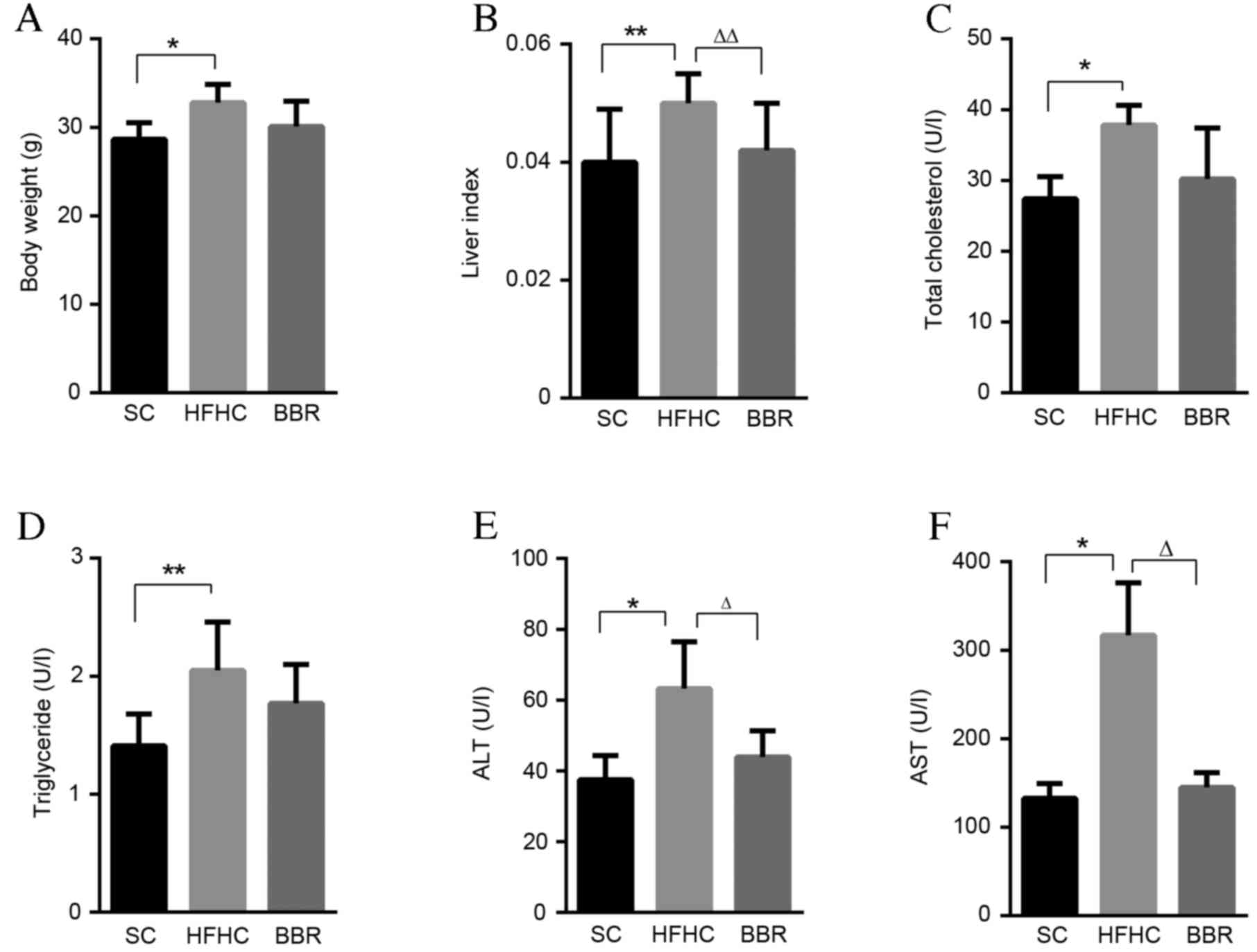 | Figure 1.BBR treatment protects against liver
injury. (A) Body weight, (B) liver-to-body weight ratio, (C) total
cholesterol, (D) serum triglyceride, (E) ALT and (F) AST levels
from the three experimental groups at the end of the study.
*P<0.05, **P<0.01 vs. the HFHC group; ∆P<0.05,
∆∆P<0.01 vs. the BBR group. BBR, berberine; ALT,
alanine transaminase; AST, aspartate transaminase; SC, standard
chow; HFHC, high-fat high-cholesterol. |
BBR supplementation markedly reduced body weight,
serum total cholesterol and triglyceride levels in
ApoE−/− mice, though the results were not statistically
significant compared with the SC group (Fig. 1A, C and D). Thus, treatment with BBR
did not notably impact the global metabolic profile of the mice.
However, the liver index and serum ALT and AST levels were
significantly lower in the BBR group compared with the HFHC group,
demonstrating a hepatoprotective role of BBR against liver injury
(P<0.05; Fig. 1E and F).
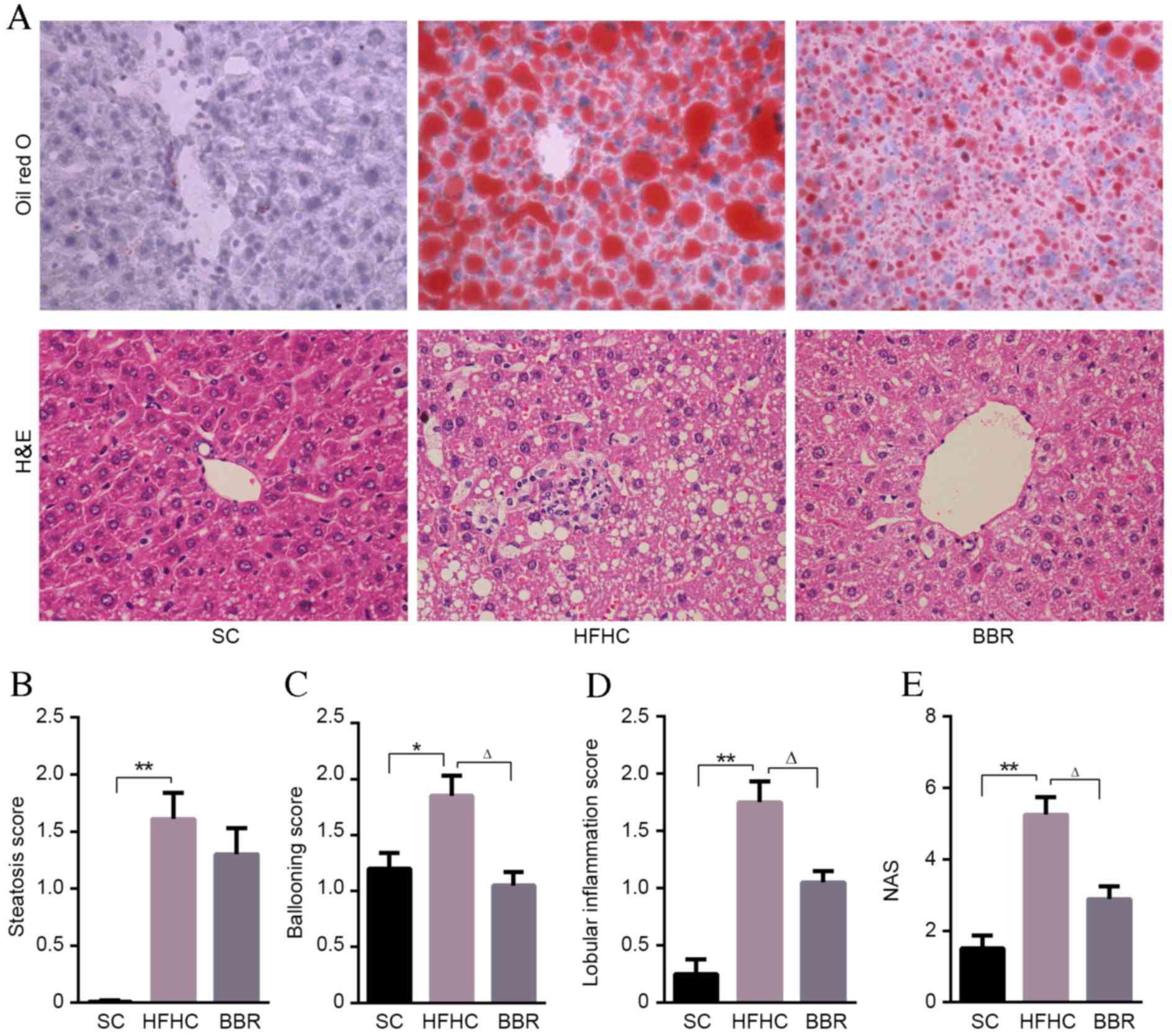
BBR alleviates hepatic steatosis and
inflammation in ApoE−/− mice with NASH
Compared with mice in the SC group, significant
pathological findings of NASH, including marked steatosis
(P<0.01), ballooning (P<0.05), lobular inflammation
(P<0.01) and NAS (P<0.01) were observed in the HFHC group
(Fig. 2). However, no significant
fibrosis was observed in the mouse liver tissues.
BBR-treated mice had a lower steatosis score
compared with HFHC-fed mice, although this difference was not
statistically significant (Fig. 2B).
Lobular inflammation and ballooning scores were significantly lower
in the BBR group compared with the HFHC group (P<0.05; Fig. 2C and D). Indeed the overall NAS score
indicated that BBR treatment had a protective effect, being
significantly lower in the BBR group compared with the HFHC group
(P<0.05; Fig. 2E). These data
indicate that inflammation is significantly associated with HFHC
diet-induced liver damage, and that treatment with BBR
significantly ameliorates this inflammation and liver injury.
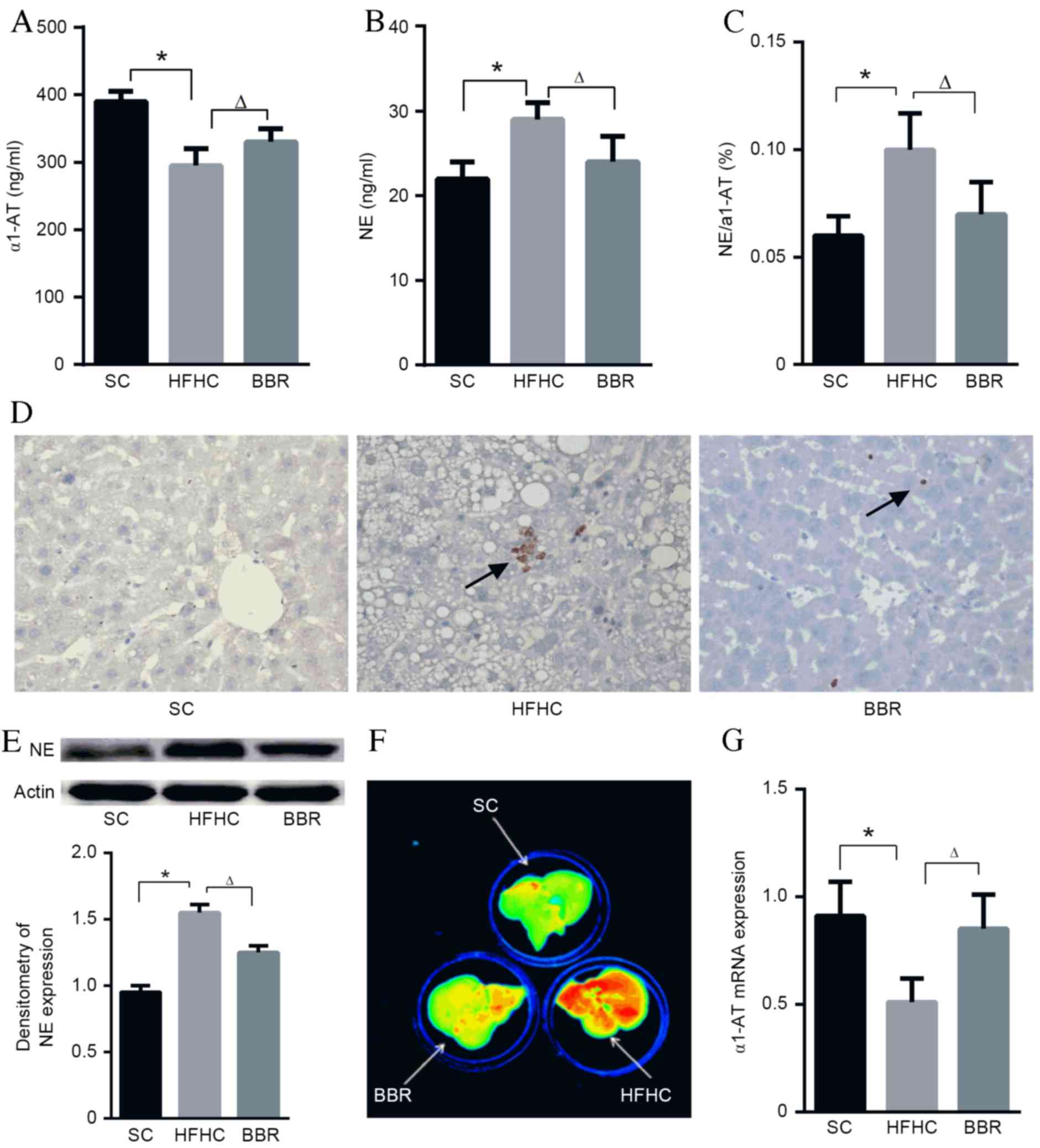
An imbalance between NE and α1-AT is
associated with NASH development in ApoE−/− mice fed a
HFHC diet
Feeding a high-fat diet to mice induces an increase
in neutrophil recruitment to tissues (19), which may initiate inflammatory
signaling pathway cascades. Particularly, NE serves an important
role in host defense and inflammation (20), whereas its counterpart, α1-AT,
protects tissues from serine protease-induced damage (8). To validate whether the expressions of
α1-AT and NE were altered in the present model, ELISA assays were
performed. The results demonstrated that serum α1-AT levels were
significantly lower in the HFHC group compared with the SC or BBR
groups (P<0.05; Fig. 3A), whereas
NE levels and the NE/α1-AT ratio were significantly higher in the
HFHC group compared with the SC and BBR groups (P<0.05; Fig. 3B and C).
Histopathological and immunoblot examinations of
liver specimens demonstrated that NE protein expression was
significantly elevated in the HFHC group compared with the SC group
(P<0.05), and BBR treatment significantly reduced NE expression
in liver tissues (P<0.05) (Fig. 3D
and E). It was also observed that NE enzymatic activity was
markedly associated with NASH (the HFHC group) and inhibited by BBR
treatment (Fig. 3F). In contrast to
NE, α1-AT protein expression in the HFHC group was significantly
lower compared with that in the SC group (P<0.05), and α1-AT
levels were significantly upregulated in the BBR group compared
with the HFHC group (P<0.05) (Fig.
3G). This NE/α1-AT balance is consistent with the reduced
inflammatory foci and ALT levels observed in the ApoE−/−
mice following BBR treatment.
BBR hepatoprotection is associated
with the CXCR4/CXCL12 signaling pathway
Localization of inflammatory cells to the liver is
an essential step in the progression of NASH (21). The active form of NE is externalized
during neutrophil activation at inflammatory sites, thus helping to
promote inflammatory and immune responses (20,22). The
present study investigated whether NE elevation affected the
expression of inflammatory chemokines and cytokines. The HFHC diet
stimulated the expression of TNF-α, MCP-1, IL-6, CXCR7 and CXCL11
in the mouse liver compared with mice fed a standard diet (data not
shown). However, although BBR treatment reduced the expression of
these cytokines and chemokines, no statistically significant
differences were observed between the HFHC and BBR group (data not
shown).
Previous studies have suggested that the
CXCR4/CXCL12 signaling pathways serves a key role in maintaining
neutrophil homeostasis (23). Liver
mRNA levels of CXCR4 and CXCL12 were significantly lower in the BBR
group compared with the HFHC group (P<0.05; Fig. 4A and B). Immunochemistry staining
yielded similar results for the protein levels of CXCR4 and CXCL12
(Fig. 4C). Gene expression analysis
found no significant differences in the expression of
mitogen-activated protein kinase 14, extracellular signal-regulated
kinase 1/2 or and B-cell lymphoma 2-associated agonist of cell
death between the HFHC and BBR groups (data not shown). However,
RAS, PLC-β, PI3K, AKT, NF-κB, IL-1 and IL-8 expression was
significantly attenuated by BBR treatment (P<0.05; Fig. 4D-J).
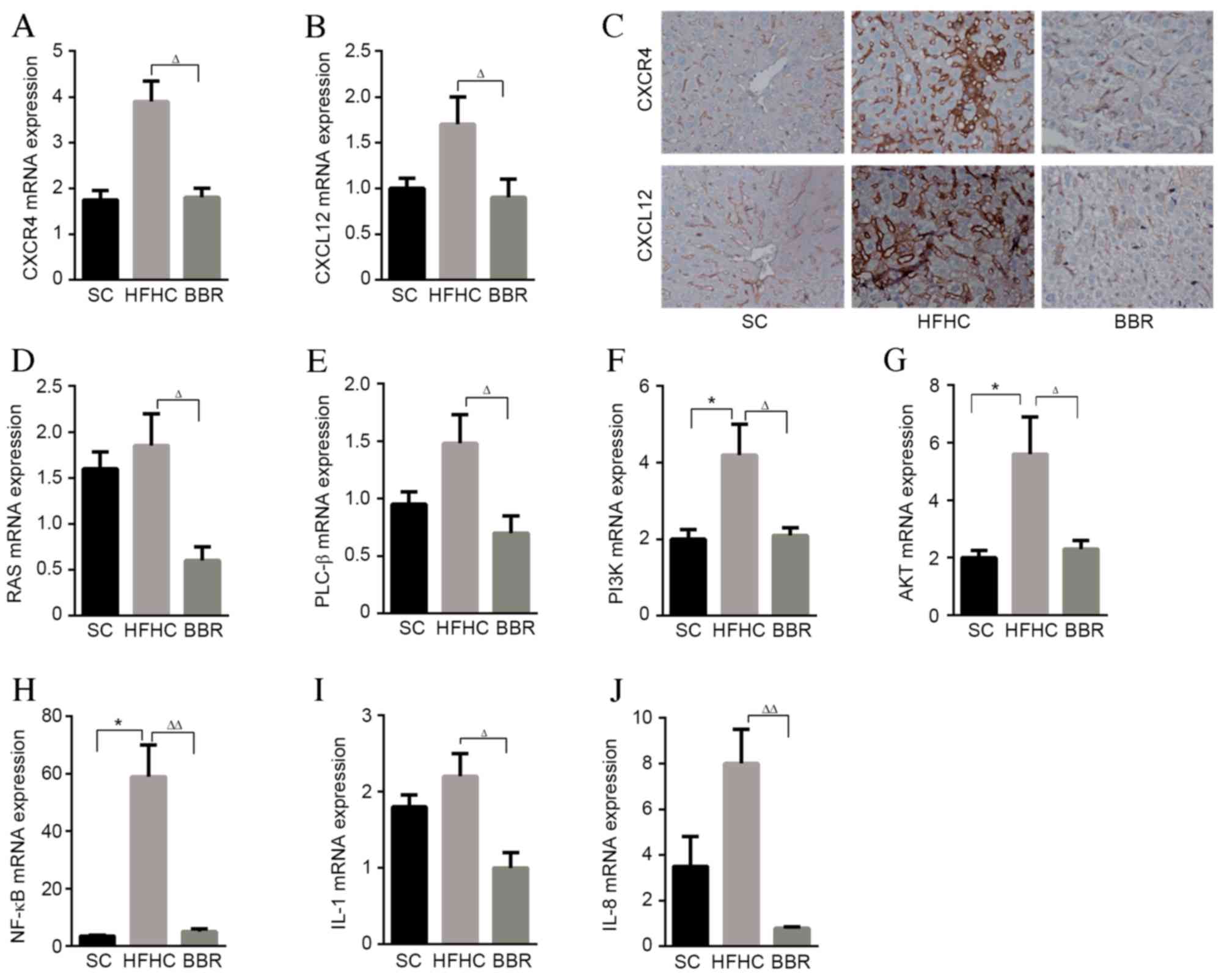 | Figure 4.BBR reducing the expression of
inflammatory cytokine, chemokines and associated genes. Hepatic
expression of (A) CXCR4 and (B) CXCL12 mRNA. (C) Representative
slides illustrating CXCR4 and CXCL12-stained liver sections from
the three experimental groups. Hepatic expression of (D) RAS, (E)
PLC-β, (F) PI3K, (G) AKT, (H) NF-κβ, (I) IL-1 and (J) IL-8 mRNA was
determined by quantitative polymerase chain reaction analysis and
normalized to GAPDH. *P<0.05 vs. the HFHC group;
∆P<0.05, ∆∆P<0.01 vs. the BBR group.
CXCR4, C-X-C chemokine receptor type 4; CXCL12, C-X-C motif
chemokine 12; BBR, berberine; HFHC, high-fat high-cholesterol; SC,
standard chow; PLC-β, phospholipase C-β; PI3K,
phosphatidylinositol-3 kinase; AKT, protein kinase B; NF-κβ,
nuclear factor-κβ; IL, interleukin. |
Discussion
ApoE−/− mice are well-established and
typically used for studies of NAFLD (15). In the present study,
ApoE−/− mice fed a HFHC diet presented with hepatic
steatosis, ballooning and increased hepatic inflammation, similar
to the clinical findings of human NASH. The principal findings of
the present study demonstrate that BBR improves the histological
and biochemical effects of NASH in mice.
BBR is known to improve lipid metabolism disorders
via multipathway mechanisms (24).
In the present study, BBR treatment reduced weight gain,
hypertriglyceridemia and hypercholesterolemia in ApoE−/−
mice fed a HFHC diet; however, the decrease was not statistically
significant, which suggests that lipid metabolism signaling
pathways are not the primary target of BBR. Histological
examination demonstrated that the hepatoprotective effect of BBR in
the ApoE−/− mouse NASH model was associated with
anti-inflammatory activities.
Previous studies have identified that fat and
cholesterol-induced inflammatory molecules are associated with
liver injury (25). Neutrophils are
typically the first immune cells to initiate the inflammatory
signaling pathway cascade in response to obesity and are able to
exacerbate chronic inflammation by recruiting other immune cells
(26). A previous study by our group
demonstrated that NASH mice with neutrophil depletion had lower
serum ALT levels, liver inflammation and mRNA levels of
proinflammatory genes (6). It is now
generally accepted that, as an important enzyme produced by
neutrophils, NE is able to promote inflammatory responses in
several disease models (26). Our
group previously demonstrated that the NE inhibitor sivelestat was
able to recapitulate the effects of neutrophil depletion in
ApoE−/− mice given a HFHC diet (6). In the present study, a significant
increase was observed in NE expression and activation in the serum
and liver tissues of mice receiving a HFHC diet. These data suggest
that inflammation may be associated with NE-dependent liver injury
in mice with HFHC diet-induced NASH, and that treatment with BBR
reverses this effect. An imbalance between NE and its natural
inhibitor α1-AT contributes to the development of obesity and
subsequent inflammation (8). The
present study demonstrated that HFHC diet-induced NASH was
associated with a significant increase in the NE/α1-AT ratio in a
mouse model. BBR treatment increased α1-AT expression in the serum
and liver tissues of the NASH mice, suggesting that the protective
effect of BBR against NASH development is associated with
alleviation of the positive feed-forward loop between the imbalance
of NE and α1-AT.
The results of the present study suggest that there
is crosstalk between the NE and CXCR4/CXCL12 signaling pathway in
NASH mice. It has previously been suggested that neutrophil
homeostasis is tightly regulated by the CXCR4/CXCL12 signaling
pathway (23). It has also been
reported that CXCR4 was markedly upregulated in patients with NASH
(27), and that the CXCL12/CXCR4
signaling pathway contributes to the enhanced recruitment of
cluster of differentiation 4+ T-cells in NASH (21). In addition, a previous study
demonstrated that NE directly interacts with CXCL12 in vitro
(28). In the present study, it was
observed that NE and CXCR4/CXCL12 expression were simultaneously
blocked by BBR treatment. Furthermore, a significant induction of
PI3K/AKT/NF-κB gene expression in HFHC-fed mice was observed in the
present study, and this was significantly reduced by BBR treatment.
This is consistent with previous observations that activation of
the PI3K/AKT/NF-κB signaling pathways occurs in patients and mice
with NAFLD (29,30), which may contribute to the increased
expression of inflammatory, chemotaxis and cell proliferation genes
(31). Notably, a previous study
reported that NE expression resulted in overexpression of IL-8
(28). This is in accordance with
the findings of the present study, which demonstrated an
upregulation of IL-8 and IL-1 in the HFHC-fed mice, and that this
upregulation was significantly attenuated by BBR treatment.
In conclusion, the results of the present study
suggest that BBR has a hepatoprotective effect and highlighted a
potential molecular mechanism underlying this effect. In the murine
model of HFHC-diet induced NASH used in the present study, NE may
trigger a proinflammatory cascade via the CXCR4/CXCL12 signaling
pathway, which further increases hepatocyte inflammation and
damage. BBR was identified to suppress NE overexpression by
restoring the balance between NE and α1-AT levels, and inhibiting
the inflammatory CXCR4/CXCL12 signaling pathway. These results
indicate that BBR is a candidate agent for the treatment of
NASH.
Acknowledgements
The present study was supported by the Natural
Science Foundation of Zhejiang, China (grant nos. LY14H070004,
LY15H070004, LY17H030009 and LQ17H070002) and the Traditional
Chinese Medical Science and Technology Project of Zhejiang Province
(grant nos. 2015ZA067 and 2015ZA141).
References
|
1
|
Farrell GC, van Rooyen D, Gan L and
Chitturi S: NASH is an inflammatory disorder: Pathogenic,
prognostic and therapeutic implications. Gut Liver. 6:149–171.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Tiniakos DG, Vos MB and Brunt EM:
Nonalcoholic fatty liver disease: Pathology and pathogenesis. Annu
Rev Pathol. 5:145–171. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Gadd VL, Skoien R, Powell EE, Fagan KJ,
Winterford C, Horsfall L, Irvine K and Clouston AD: The portal
inflammatory infiltrate and ductular reaction in human nonalcoholic
fatty liver disease. Hepatology. 59:1393–1405. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Mócsai A: Diverse novel functions of
neutrophils in immunity, inflammation, and beyond. J Exp Med.
210:1283–1299. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Uchida Y, Freitas MC, Zhao D, Busuttil RW
and Kupiec-Weglinski JW: The protective function of neutrophil
elastase inhibitor in liver ischemia/reperfusion injury.
Transplantation. 89:1050–1056. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zang S, Wang L, Ma X, Zhu G, Zhuang Z, Xun
Y, Zhao F, Yang W, Liu J, Luo Y, et al: Neutrophils play a crucial
role in the early stage of nonalcoholic steatohepatitis via
neutrophil elastase in mice. Cell Biochem Biophys. 73:479–487.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zang S, Ma X, Zhuang Z, Liu J, Bian D, Xun
Y, Zhang Q, Zhao F, Yang W, Liu J, et al: Increased ratio of
neutrophil elastase to α1-antitrypsin is closely associated with
liver inflammation in patients with nonalcoholic steatohepatitis.
Clin Exp Pharmacol Physiol. 43:13–21. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Mansuy-Aubert V, Zhou QL, Xie X, Gong Z,
Huang JY, Khan AR, Aubert G, Candelaria K, Thomas S, Shin DJ, et
al: Imbalance between neutrophil elastase and its inhibitor
α1-antitrypsin in obesity alters insulin sensitivity, inflammation,
and energy expenditure. Cell Metab. 17:534–548. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kulkarni SK and Dhir A: Berberine: A plant
alkaloid with therapeutic potential for central nervous system
disorders. Phytother Res. 24:317–324. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Battu SK, Repka MA, Maddineni S,
Chittiboyina AG, Avery MA and Majumdar S: Physicochemical
characterization of berberine chloride: A perspective in the
development of a solution dosage form for oral delivery. AAPS Pharm
Sci Tech. 11:1466–1475. 2010. View Article : Google Scholar
|
|
11
|
Li J, Pan Y, Kan M, Xiao X, Wang Y, Guan
F, Zhang X and Chen L: Hepatoprotective effects of berberine on
liver fibrosis via activation of AMP-activated protein kinase. Life
Sci. 98:24–30. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Sun X, Zhang X, Hu H, Lu Y, Chen J, Yasuda
K and Wang H: Berberine inhibits hepatic stellate cell
proliferation and prevents experimental liver fibrosis. Biol Pharm
Bull. 32:1533–1537. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ye D, Li FY, Lam KS, Li H, Jia W, Wang Y,
Man K, Lo CM, Li X and Xu A: Toll-like receptor-4 mediates
obesity-induced non-alcoholic steatohepatitis through activation of
X-box binding protein-1 in mice. Gut. 61:1058–1067. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Van Herck H, Baumans V, Brandt CJ, Boere
HA, Hesp AP, van Lith HA, Schurink M and Beynen AC: Blood sampling
from the retro-orbital plexus, the saphenous vein and the tail vein
in rats: Comparative effects on selected behavioural and blood
variables. Lab Anim. 35:131–139. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Jeon S, Park YJ and Kwon YH: Genistein
alleviates the development of nonalcoholic steatohepatitis in
ApoE(−/−) mice fed a high-fat diet. Mol Nutr Food Res. 58:830–841.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kleiner DE, Brunt EM, Van Natta M, Behling
C, Contos MJ, Cummings OW, Ferrell LD, Liu YC, Torbenson MS,
Unalp-Arida A, et al: Design and validation of a histological
scoring system for nonalcoholic fatty liver disease. Hepatology.
41:1313–1321. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wienken CJ, Baaske P, Duhr S and Braun D:
Thermophoretic melting curves quantify the conformation and
stability of RNA and DNA. Nucleic Acids Res. 39:e522011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Elgazar-Carmon V, Rudich A, Hadad N and
Levy R: Neutrophils transiently infiltrate intra-abdominal fat
early in the course of high-fat feeding. J Lipid Res. 49:1894–1903.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Pham CT: Neutrophil serine proteases:
Specific regulators of inflammation. Nat Rev Immunol. 6:541–550.
2006. View
Article : Google Scholar : PubMed/NCBI
|
|
21
|
Boujedidi H, Robert O, Bignon A,
Cassard-Doulcier AM, Renoud ML, Gary-Gouy H, Hemon P, Tharinger H,
Prévot S, Bachelerie F, et al: CXCR4 dysfunction in non-alcoholic
steatohepatitis in mice and patients. Clin Sci (Lond). 128:257–267.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Snelgrove RJ, Jackson PL, Hardison MT,
Noerager BD, Kinloch A, Gaggar A, Shastry S, Rowe SM, Shim YM,
Hussell T and Blalock JE: A critical role for LTA4H in limiting
chronic pulmonary neutrophilic inflammation. Science. 330:90–94.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Eash KJ, Means JM, White DW and Link DC:
CXCR4 is a key regulator of neutrophil release from the bone marrow
under basal and stress granulopoiesis conditions. Blood.
113:4711–4719. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhang Q, Xiao X, Feng K, Wang T, Li W,
Yuan T, Sun X, Sun Q, Xiang H and Wang H: Berberine moderates
glucose and lipid metabolism through multipathway mechanism. Evid
Based Complement Alternat Med. 2011:pii:9248512011. View Article : Google Scholar
|
|
25
|
Bartelt A, Orlando P, Mele C, Ligresti A,
Toedter K, Scheja L, Heeren J and Di Marzo V: Altered
endocannabinoid signalling after a high-fat diet in Apoe(−/−) mice:
Relevance to adipose tissue inflammation, hepatic steatosis and
insulin resistance. Diabetologia. 54:2900–2910. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Talukdar S, Oh DY, Bandyopadhyay G, Li D,
Xu J, McNelis J, Lu M, Li P, Yan Q, Zhu Y, et al: Neutrophils
mediate insulin resistance in mice fed a high-fat diet through
secreted elastase. Nat Med. 18:1407–1412. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Liu H, Li J, Tillman B, Morgan TR, French
BA and French SW: TLR3/4 signaling is mediated via the NFκB-CXCR4/7
pathway in human alcoholic hepatitis and non-alcoholic
steatohepatitis which formed Mallory-Denk bodies. Exp Mol Pathol.
97:234–240. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Korkmaz B, Horwitz MS, Jenne DE and
Gauthier F: Neutrophil elastase, proteinase 3, and cathepsin G as
therapeutic targets in human diseases. Pharmacol Rev. 62:726–759.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Sharma M, Mitnala S, Vishnubhotla RK,
Mukherjee R, Reddy DN and Rao PN: The riddle of nonalcoholic fatty
liver disease: Progression from nonalcoholic fatty liver to
nonalcoholic steatohepatitis. J Clin Exp Hepatol. 5:147–158. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Pisonero-Vaquero S, Martínez-Ferreras Á,
García-Mediavilla MV, Martínez-Flórez S, Fernández A, Benet M,
Olcoz JL, Jover R, González-Gallego J and Sánchez-Campos S:
Quercetin ameliorates dysregulation of lipid metabolism genes via
the PI3K/AKT pathway in a diet-induced mouse model of nonalcoholic
fatty liver disease. Mol Nutr Food Res. 59:879–893. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Teicher BA and Fricker SP: CXCL12
(SDF-1)/CXCR4 pathway in cancer. Clin Cancer Res. 16:2927–2931.
2010. View Article : Google Scholar : PubMed/NCBI
|