Introduction
Cardiovascular disease (CVD) is one of the leading
causes of mortality worldwide, and was responsible for ~31.9% of
all mortality in 2010 (1). In the
last 20 years, CVD mortality rates have decreased; however, these
rates are predicted to increase again as a result of global
increases in diabetes and obesity. Additionally, the prevalence of
CVD has increased in developing countries, due to the acquisition
of a westernized lifestyle, which is likely to contribute to the
suspected increase in CVD mortality rates (2). Atherosclerosis can lead to myocardial
infarction, cerebrovascular accident and peripheral vascular
disease, and is now identified as an inflammatory disorder of
medium and large arteries. Factors related to the development of
atherosclerosis include high plasma cholesterol levels and
hypertension (3).
As highly differentiated muscle cells, vascular
smooth muscle cells (VSMCs) comprise the medial layer of the vessel
wall, and they control blood pressure via the contraction and
relaxation of vessels. VSMCs may have an important role in
atherogenesis by actively participating in the control of vascular
remodeling and plaque stabilization (4). Induced by various factors, the aberrant
proliferation of VSMCs has been confirmed to be critical in the
pathogenesis of cardiovascular diseases, such as pulmonary artery
hypertension, post-angioplasty restenosis, vein graft failure and
atherosclerosis (5,6). Distinguishing them from many terminal
differentiated cells, VSMCs have the potential of phenotypic
switching, meaning VSMCs are capable of transferring between
proliferative-synthetic and quiescent-contractile phenotypes,
initiated by changes in the local environment (7,8).
MicroRNA (miRNA) are small, endogenous, non-coding
regulatory molecules of 20–22 nucleotides in length. They are able
to regulate gene expression at the posttranscriptional level by
binding to the 3′-untranslated region (UTR) of messenger RNA (mRNA)
(8). The involvement of miRNA in the
pathogenesis of various human diseases, including cancer,
inflammatory diseases and cardiovascular diseases (9–13), has
been widely investigated. Multiple miRNA are known to control the
different phases of atherogenesis (14); they have an important role in the
proliferation and differentiation of VSMCs under pathological or
physiological conditions, as well as modulating both adaptive and
innate immune responses within VSMCs, influencing every step of
atherogenesis from plaque formation to destabilization and rupture
(15). Although many miRNA have been
linked to atherosclerosis, this association has only been
repeatedly reported for some miRNA, including miR-21, −146a, −150
and −155 (16,17).
Forkhead Box P1 (FoxP1) was first identified by Shu
et al (18), which was
considered a glutamine rich factor that belongs to the FoxP
subfamily of the fox-transcription factors. Bot et al
(19) suggested that FoxP1 modulates
collagen synthesis and proliferation of smooth muscle cell through
downstream target of transforming growth factor (TGF)-β. FoxP1
levels may also be associated with interleukin-2, −4 and −10
expression, which are involved in plaque stability (19).
In the present study, four miRNA (miR-206, −125a,
−876 and −183) that have previously been reported to be
differentially expressed in atherosclerosis (20) were selected. Screening was performed
to compare their ability to affect the proliferation of VSMCs with
the control. It was demonstrated that miR-206 was able to
substantially suppress the proliferation of VSMCs via targeting
forkhead box protein subfamily P (FOXP) 1.
Materials and methods
Ethics statement
All patients involved in the present study provided
their written informed consent. The present study was approved by
the Ethics Committee of Liaocheng People's Hospital (Liaocheng,
China).
Patient samples
A total of 22 human atherosclerotic plaques were
collected from patients (age, 67.63±7.23 years; males, 15; females,
7) who received endarterectomy to reduce the long-term risk of
stroke, and 20 normal control tissue samples were obtained from
patients (age, 65.45±5.43 years; males, 12; females, 8) who
received coronary artery bypass surgery in Liaocheng People's
Hospital. All patients included in the present study were
clinically asymptomatic. Following surgery, all tissue samples were
flash-frozen in liquid nitrogen and, subsequently, stored at −80°C
until the experiments were conducted.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) analysis
RNA isolated from tissue samples were subjected to
an RNeasy kit (Qiagen China Co., Ltd., Shanghai, China) and an
miRNeasy Mini kit (Qiagen China Co., Ltd.) to isolate miRNA and
mRNA, respectively according to the manufacturer's instructions. A
SYBR Green Master Mix kit and a miRCURY LNA Universal RT microRNA
PCR kit (both Exiqon, Vedbæk, Denmark) for cDNA synthesis were
utilized to perform RT-qPCR for miRNA and mRNA in accordance with
the manufacturer's protocols. The following primers were used:
miR-206, forward 5′-CCAAAGCGGAGTCTCGCAT-3′ and reverse
5′-GCCTAGCATCTTGCTTAGCTC-3′; miR-125a, forward
5′-TTGCTCCAGCAGGGTAACTG-3′ and reverse 5′-GTGGTCGAGAAGCTTGTGTGA-3′;
miR-876, forward 5′-ACCTGCACCCGATTCACAG-3′ and reverse
5′-TGGCAGCTCCATACTGACCA-3′; miR-183, forward
5′-GAGAATTGTGGCGTCAAGTCA-3′ and reverse
5′-CAGGTATGTTTTCCAGTGCTCC-3′; FOXP1, forward
5′-CAGCGAAACCACGAAAAGAAG−3′ and reverse
5′-GGTCCACCTCTGTTAGTGATA-3′; and β-actin: Forward
5′-CAGCAAAGAAGGACACGAAAC-3′ and reverse 5′-ACCGGGGATTATTCCTTCTGA-3.
RT-qPCR for four miRNA (miR-206, −125a, −876 and −183) and FOXP1
was conducted using a HotStart-IT SYBR Green qPCR Master Mix with
UDG (2X) (Affymetrix, Inc., Santa Clara, CA, USA) and a DNA Engine
Opticon 2 Real-Time Cycler (MJ Research, Inc., Waltham, MA, USA).
Subsequently, the 2−ΔΔCq method was used to quantify the
expression levels of each miRNA or mRNA (18). Experiments were repeated at least
three times: β-actin was the internal reference gene.
Cell culture and oligonucleotide
transfection
Human aortic VSMCs (Lonza, Inc., Allendale, NJ, USA)
were cultured in Dulbecco's Modified Eagle's medium (DMEM; Gibco;
Thermo Fisher Scientific, Inc., Waltham, MA, USA) supplemented with
10% fetal bovine serum (FBS) (Bio-Rad Laboratories, Inc., Hercules,
CA, USA). Cells were incubated at 37°C in an atmosphere containing
5% CO2. Mimics for four miRNA (miR-206, −125a, −876 and
−183) were purchased from Ambion (Thermo Fisher Scientific, Inc.)
and transfection was performed using Lipofectamine 2000
(Invitrogen; Thermo Fisher Scientific, Inc.) according to the
manufacturer's protocol.
Construction of FOXP1 plasmid
The coding sequence of FOXP1 was amplified using PCR
and inserted into pcDNA3.0 (Invitrogen; Thermo Fisher Scientific,
Inc.). PCR amplification was performed as follows: 94°C for 3 min,
followed by 30 cycles of 94°C for 40 sec, 56°C for 45 sec and 72°C
for 60 sec, followed by terminal elongation. A DNA Engine Opticon 2
Real-Time Cycler (MJ Research, Inc.) and Taq DNA Polymerase
(Invitrogen; Thermo Fisher Scientific, Inc.) were used. Insertion
accuracy was confirmed via direct Sanger sequencing. The construct
was subsequently transfected into VSMCs using Lipofectamine
2000.
CCK-8 assay
A cell counting kit-8 (CCK-8; Sigma-Aldrich; Merck
Millipore, Darmstadt, Germany) assay was used to measure the
proliferation of cells. Accordingly, 24 h subsequent to producing
equal numbers of VSMCs, the cells were transfected with either the
miR-206 mimic, or both the miR-206 mimic and the FOXP1 plasmid.
Following transfection, DMEM was used to incubate the cells.
Subsequently, 10 µl of the CCK-8 solution was added to each well
where cells were cultured for 2 h at the end of CCK-8 treatment.
Subsequently, the absorbance value was measured at a wavelength of
450 nm using a microplate absorbance reader to confirm the number
of viable cells.
Luciferase reporter assays
The full length of the 3′UTR of FOXP1 was amplified
and cloned into an miR-reporter construct (Ambion; Thermo Fisher
Scientific, Inc.). Subsequently, the potential miR-206 binding site
in the 3′UTR of FOXP1 was replaced with its complementary sequence
using site-directed mutagenesis (210518; Stratagene; Agilent
Technologies, Inc., Santa Clara, CA, USA). For the luciferase
assay, 100 ng/ml of the wild-type or mutant miR-reporter construct
was co-transfected with the 100 nM miR-206 mimics into VSMCs using
Lipofectamine 2000. Following 36 h, cells were lysed and 50 µl of
the lysate was utilized for β-Gal assay (BioVision, Inc., Milpitas,
CA, USA) and 10 µl (2X) was used for the luciferase assay. Assays
were conducted according to the manufacturer's protocol.
Western blot analysis
Following homogenization of the tissue samples or
the cultured cells, proteins were extracted with an extraction
buffer composed of radioimmunoprecipitation assay buffer, protease
inhibitor cocktail and phosphatase inhibitor cocktail (Calbiochem;
Merck Millipore). Total proteins were quantified by the Bradford
method (21). Proteins (30 µg) were
separated by 10% SDS-PAGE and subsequently transferred to a
nitrocellulose membrane (Merck Millipore). For blocking, 5% skimmed
milk was used to block the membrane for 2 hat 37°C and
phosphate-buffered saline was utilized to wash the membrane for 5
min. Membranes were incubated overnight with primary antibodies
against FOXP1 or β-actin (1:5,000 dilution; Abcam, Cambridge, UK)
or rabbit β-actin (1:12,000 dilution; ab8227; Abcam) at 4°C
overnight. Subsequently the membrane was incubated with horseradish
peroxidase-linked secondary antibodies (1:15,000 dilution; ab7090;
Abcam) at 37°C for 2 h. Following this, the membrane was treated
with enhanced chemiluminescent reagent (Amersham ECL detection
system; GE Healthcare Life Sciences, Chalfont, UK). An
Electrochemiluminescence Plus Western Blotting Detection System (GE
Healthcare Bio-Sciences, Pittsburgh, PA, USA) was used to detect
the bound antibodies and high performance chemiluminescence film
(GE Healthcare Bio-Sciences) was used to detect the
chemiluminiscent signals in accordance with the manufacturer's
recommendation. Protein expression was normalized to β-actin and
quantified by densitometry. ImageJ software (version 1.37)
(National Institutes of Health; Bethesda, MD, USA) was utilized to
quantify the expression level of FOXP1.
Statistical analysis
Statistical analysis was performed using SPSS 16.0
(SPSS Inc, Chicago, IL, USA). Normal distribution of data was
tested using the Shapiro-Wilk test. If a normal distribution was
not confirmed, the Mann-Whitney U test was used to examine
differences between groups. If a normal distribution was confirmed,
Student's t-test or one-way analysis of variance was used to
compare the parameter between two or more groups, respectively.
Categorical variables were compared using the Chi-square test.
P<0.05 was considered to indicate a statistically significant
difference.
Results
miR-206 suppresses the survival of
VSMCs
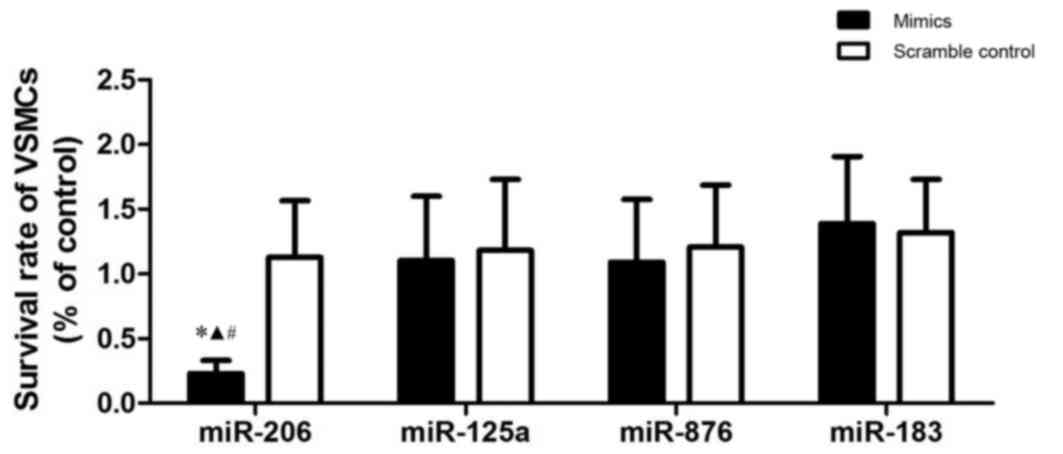
To explore the pathogenesis of atherosclerosis, the
survival rate of VSMCs transfected with several candidate miRNA
that were previously reported to be differentially expressed in
atherosclerosis (including miR-206, −125a, −876 and −183) (22), was investigated. Results demonstrated
that miR-206 was the only miRNA that was able to significantly
suppress the growth of the VSMCs. A significantly reduced survival
rate was observed in comparison with the other miRs (P<0.05;
Fig. 1), indicating that miR-206 may
be the one miRNA out of the four miRNA investigated that suppresses
the survival of VSMCs.
miR-206 targets the conserved 3′UTR of
FOXP1
Due to the evident effect of miR-206 upon VSMC
survival rates, it was hypothesized that the downstream miR-206
target genes may be associated with the control of cell
proliferation. Bioinformatics tools (TargetScan; http://www.targetscan.org/vert_71) were utilized
to identify the potential target genes of miR-206. Two putative
target sites (820–826 and 3447–3455) in the 3′UTR of the FOXP1 gene
were identified as potential binding sites for miR-206 (Fig. 2), indicating that FOXP1 may be a
target of miR-206.
To verify whether FOXP1 is a direct target gene of
miR-206 and to determine the exact miR-206 binding site on the
FOXP1 gene, two wild-type FOXP1 3′UTR segments, and two different
mutants (wild-type site 820–826 and mutated site 3447–3455, defined
as Mutant1 in Fig. 2; mutated site
820–826 and wild-type site 3447–3455, defined as Mutant2 in
Fig. 2) of the FOXP1 3′UTR segments
were cloned into different miR-luciferase reporter constructs.
Following treatment with miR-206 in the luciferase reporter assay,
cells co-transfected with plasmids containing the Mutant2 FOXP1
3′UTR segments exhibited similar luciferase activity when compared
with the scramble controls (Fig. 3).
In contrast, cells co-transfected with Mutant1 of the FOXP1 3′UTR
segments demonstrated significantly reduced relative luciferase
activity compared with the scramble controls, and exhibited no
significant differences with cells co-transfected with wild-type
FOXP1 3′UTR segments (P<0.05; Fig.
3). This indicated that miR-206 downregulated FOXP1 expression
by targeting the 820–826 putative binding site on FOXP1 3′UTR
segments.
FOXP1 expression is downregulated by
miR-206 in atherosclerosis tissues
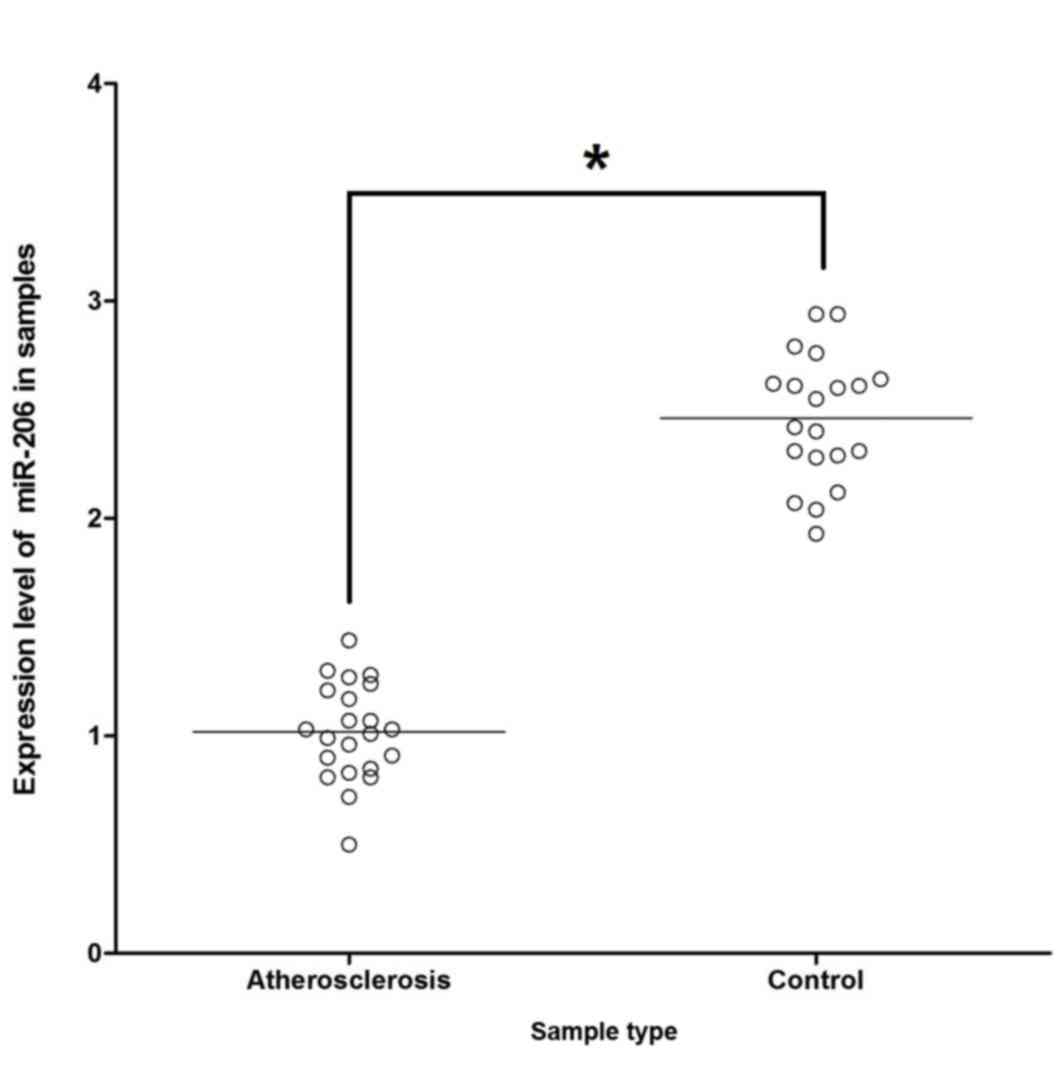
To explore the mechanism by which miR-206 influences
the development of atherosclerosis via targeting the predicted
target FOXP1 gene, the mRNA expression level of miR-206 among the
22 atherosclerosis samples and the 20 control samples was
investigated. It was demonstrated that expression of miR-206 was
markedly suppressed in the atherosclerosis samples compared with
control samples (Fig. 4).
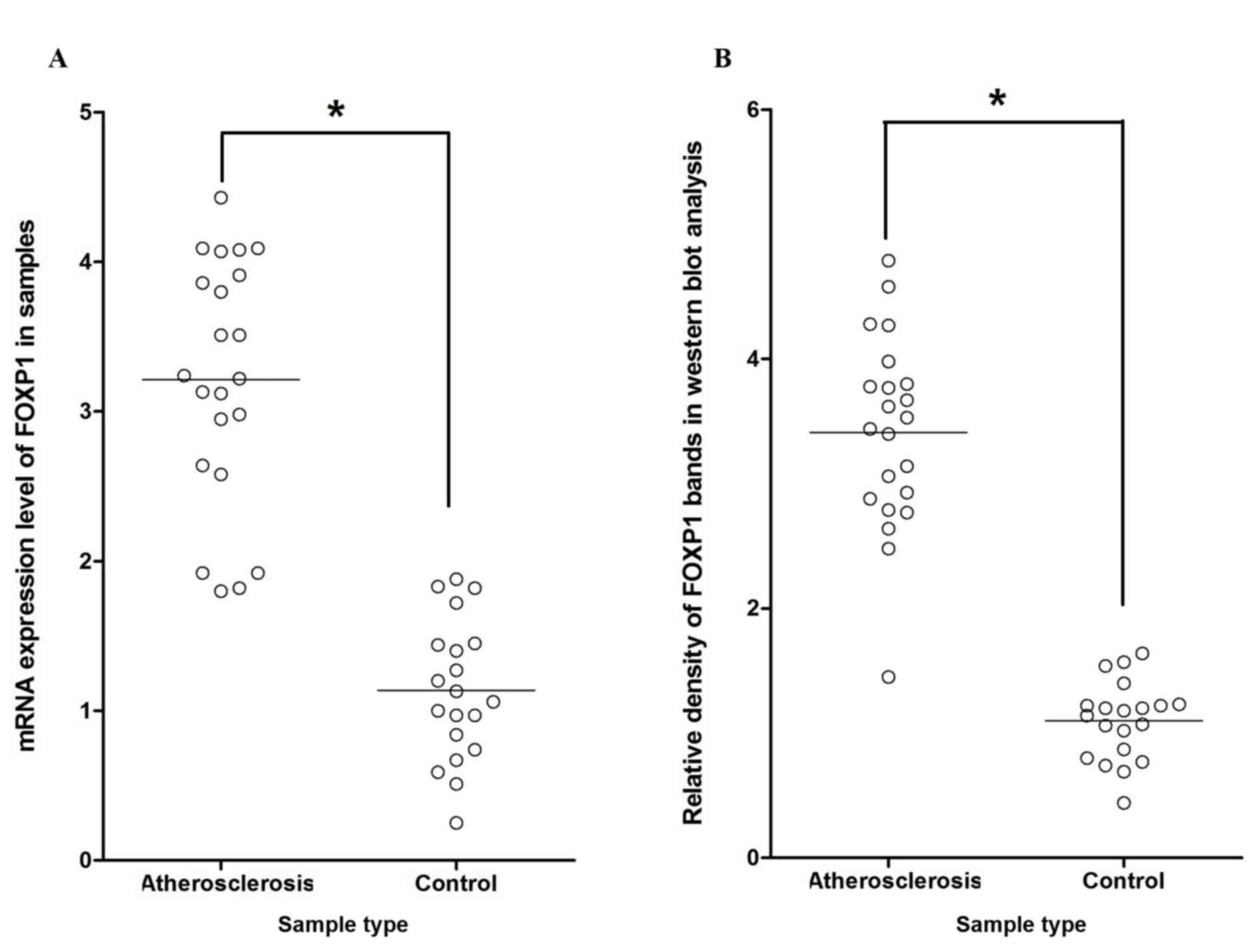
Additionally, RT-qPCR and western blot analysis were performed
separately on atherosclerosis groups and control groups to
investigate the regulatory association between miR-206 and FOXP1.
This revealed significantly increased mRNA/protein expression
levels of FOXP1 among atherosclerosis tissue samples compared with
the normal control samples (P<0.05; Fig. 5A and B, respectively), indicating the
negatively correlated relationship between miR-206 and FOXP1.
FOXP1 overexpression has a potential
rescue effect upon miR-206
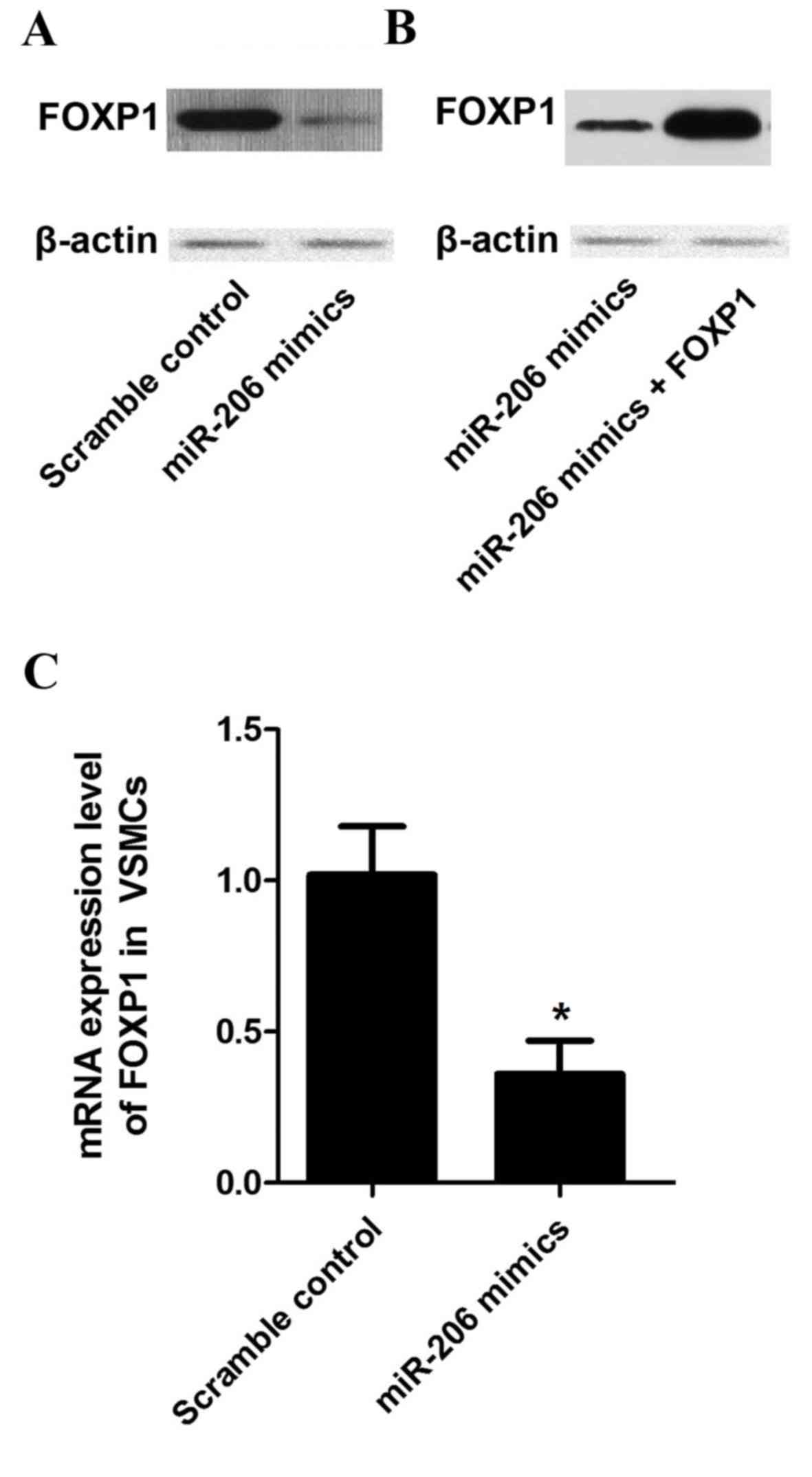
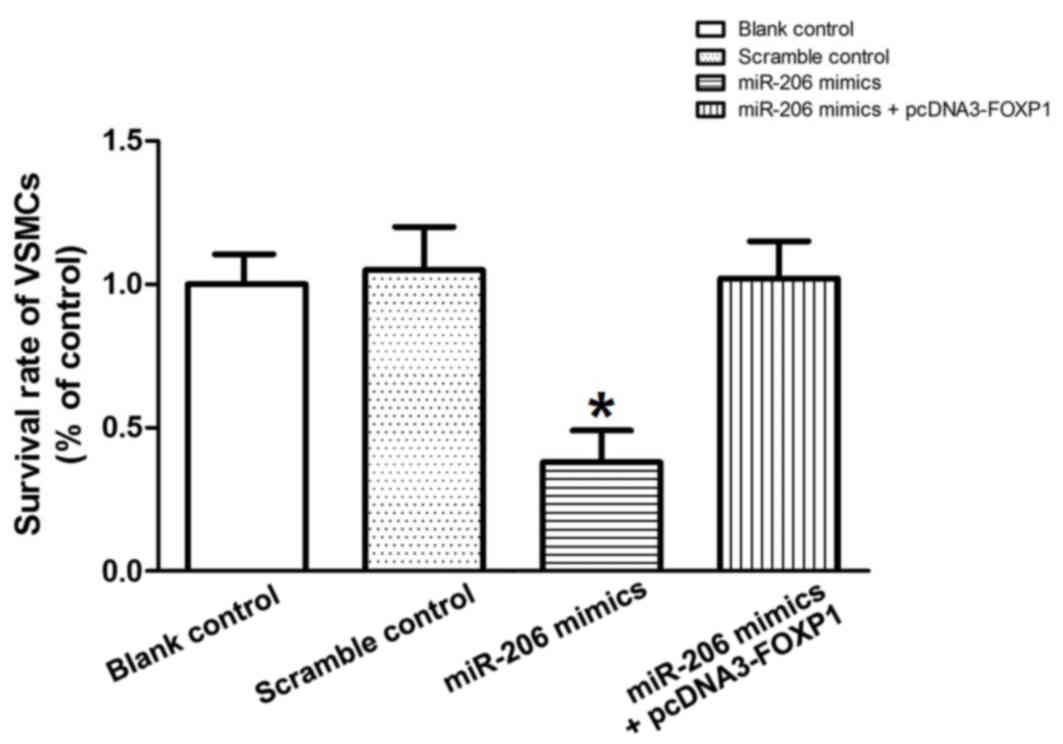
To explore the regulatory relationship between
miR-206 and FOXP1 in vitro, the mRNA and protein expression
levels of FOXP1 in VSMCs treated with miR-206 mimics were
investigated. Marked decreases in protein expression (Fig. 6A) and significant decreases in mRNA
expression levels of FOXP1 (P<0.05; Fig. 6B) were detected among the miR-206
mimics treatment group compared with the scramble controls. To
perform the rescue test, recombinant plasmids expressing the
complete coding sequence of the FOXP1 gene were constructed and,
subsequently, transfected into cells treated with miR-206 mimics.
The viability of the VSMCs transfected with the scramble control,
the miR-206 mimics and the miR-206 mimics with the FOXP1-expressing
plasmid was determined. miR-206 transfection significantly
decreased (P<0.05) the survival rate of the VSMCs compared with
the control, while overexpression of FOXP1 almost reversed the
survival decrease induced by miR-206 (Fig. 7), indicating that the miR-206
decreases VSMC viability and this is, at least partially, mediated
by the target of miR-206, FOXP1.
Discussion
miR-206 is a functional miRNA in both physiological
and pathological conditions. As a differentiation-related miRNA,
miR-206 has been demonstrated to have a crucial role in the process
of cellular differentiation. A study by Dey et al (23) demonstrated that differentiation and
proliferation of myoblasts was modulated by miR-206 through its
direct target, PAX7. A study by Anderson et al (24) identified miR-206 as a key regulator
of osteoblast differentiation, by inhibiting the expression of the
connexin-43 gene. Jalali et al (25) demonstrated that miR-206 may be a
potential regulator of the differentiation and proliferation of
VSMCs in pulmonary arterial hypertension. As an anti-oncogenic
miRNA, miR-206 has been demonstrated to inhibit both tumorigenesis
and tumor progression. A study by Song et al (26) exhibited that miR-206 is a
pro-apoptotic activator of cell death that is associated with the
regulation of Notch3 function and, thus, the suppression of tumor
growth. In various types of cancer, miR-206 demonstrates decreased
regulation (27,28). By inhibiting the expression of Notch3
protein, miR-206 overexpression in HeLa cancer cells increases
apoptosis (29). When compared with
controls, decreased expression levels of miR-206 in mice with
pulmonary hypertension induced by hypoxia has been demonstrated in
experiments in vivo.
On the foundation of previous evidence, the role of
miR-206 in modulating the differentiation, proliferation and
apoptosis of VSMCs in vitro was investigated. In the present
study, the survival rates of VSMCs transfected with several
candidate microRNA, which were previously reported to be
differentially expressed in atherosclerosis (including miR-206,
−125a, −876 and −183) (20), were
investigated. miR-206 was identified as the only miRNA, out of the
four miRNA investigated, that could markedly suppress the growth of
VSMCs. Furthermore, bioinformatics tools were utilised to identify
FOXP1 as a target of miR-206 and luciferase reporter analysis was
performed to confirm this relationship. Luciferase reporter
analysis revealed that the binding site for miR-206 was in the
3′UTR of FOXP1. A negatively correlated relationship between
miR-206 and FOXP1 was confirmed by the observation that expression
of miR-206 was suppressed in the atherosclerosis samples, whereas
mRNA/protein expression levels of FOXP1 were upregulated, compared
with control samples.
FOXP1, a member of the FOXP subfamily of the
FOX-transcription factors, was identified in a study by Shu et
al (18) as a glutamine rich
transcription factor, detectable in a wide variety of fetal and
adult tissue cell types. Additionally, the expression of FOXP1 has
been detected in several malignant neoplasms, including B-cell
lymphomas and prostate cancer (30).
FOXP1 has been reported to be involved in the control of
differentiation and proliferation of cardiac muscle cells, and
differential effects of FOXP1 on the early and late stage of
cardiac development have been demonstrated (31). Different types of cells in
atherosclerotic lesions express FOXP1, which is a downstream target
of TGF-β (19). FOXP1 is
predominantly expressed by smooth muscle cells (19) in atherosclerotic plaques. The role of
FOXP1 in some other cell types relevant to atherosclerosis has been
studied (32); however, its effect
on VSMCs remains largely unknown.
In the present study, it was hypothesized that FOXP1
may be a downstream target of miR-206, and its regulatory effect on
the proliferation of VSMCs may be mediated by its downstream
effectors such as fibroblast growth factor and TGF-β (33). In the present study, miR-206
transfection markedly decreased the survival rate of VSMCs compared
with the controls. However, the overexpression of FOXP1 in VSMCs
treated with miR-206 almost restored the survival rates, indicating
that the survival-suppressing effect of miR-206 is, at least
partially, mediated by its target, FOXP1.
In conclusion, the present study has provided a
mechanistic model that may demonstrate the role of miR-206 and
FOXP1 in the development of atherosclerotic plaques. It was
demonstrated that the miR-206/FOXP1 pathway is functionally
involved in the control of the proliferation of VSMCs. These
results suggest that miR-206 inhibits the expression of FOXP1,
resulting in decreased viability and increased apoptosis of VSMCs.
Decreased levels of miR-206 release the physiologically-inhibited
expression of FOXP1, resulting in the increased expression of FOXP1
and, thereby, activation of the downstream TGF-β signalling
pathway. High mRNA/protein expression levels of FOXP1 were detected
in atherosclerosis samples compared with controls; therefore,
miR-206 may be a protective factor in atherosclerosis by inhibiting
the expression of FOXP1. FOXP1 may be a potential target for the
treatment and/or prevention of acute cardiovascular events and
atherosclerotic plaque formation.
References
|
1
|
Go AS, Mozaffarian D, Roger VL, Benjamin
EJ, Berry JD, Blaha MJ, Dai S, Ford ES, Fox CS, Franco S, et al:
Heart disease and stroke statistics-2014 update: A report from the
American Heart Association. Circulation. 129:e28–e292. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Poole-Wilson P: The prevention of
cardiovascular disease worldwide: Whose task and WHO's task? Clin
Med (Lond). 5:379–384. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
McLaren JE, Michael DR, Ashlin TG and
Ramji DP: Cytokines, macrophage lipid metabolism and foam cells:
Implications for cardiovascular disease therapy. Prog Lipid Res.
50:331–347. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Doran AC, Meller N and McNamara CA: Role
of smooth muscle cells in the initiation and early progression of
atherosclerosis. Arterioscler Thromb Vasc Biol. 28:812–819. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Lemmens K, Doggen K and De Keulenaer GW:
Role of neuregulin-1/ErbB signaling in cardiovascular physiology
and disease: Implications for therapy of heart failure.
Circulation. 116:954–960. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Marx SO, Totary-Jain H and Marks AR:
Vascular smooth muscle cell proliferation in restenosis. Circ
Cardiovasc Interv. 4:104–111. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Rensen SS, Doevendans PA and van Eys GJ:
Regulation and characteristics of vascular smooth muscle cell
phenotypic diversity. Neth Heart J. 15:100–108. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Li L, Zhang HN, Chen HZ, Gao P, Zhu LH, Li
HL, Lv X, Zhang QJ, Zhang R, Wang Z, et al: SIRT1 acts as a
modulator of neointima formation following vascular injury in mice.
Circ Res. 108:1180–1189. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Bartel DP: MicroRNAs: Target recognition
and regulatory functions. Cell. 136:215–233. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Nana-Sinkam SP and Croce CM: MicroRNAs as
therapeutic targets in cancer. Transl Res. 157:216–225. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Dai R and Ahmed SA: MicroRNA, a new
paradigm for understanding immunoregulation, inflammation, and
autoimmune diseases. Transl Res. 157:163–179. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Santovito D, Mezzetti A and Cipollone F:
MicroRNAs and atherosclerosis: New actors for an old movie. Nutr
Metab Cardiovasc Dis. 22:937–943. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Santovito D, De Nardis V, Marcantonio P,
Mandolini C, Paganelli C, Vitale E, Buttitta F, Bucci M, Mezzetti
A, Consoli A and Cipollone F: Plasma exosome microRNA profiling
unravels a new potential modulator of adiponectin pathway in
diabetes: Effect of glycemic control. J Clin Endocrinol Metab.
99:E1681–E1685. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Nazari-Jahantigh M, Wei Y and Schober A:
The role of microRNAs in arterial remodelling. Thromb Haemost.
107:611–618. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kang H and Hata A: MicroRNA regulation of
smooth muscle gene expression and phenotype. Curr Opin Hematol.
19:224–231. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ma X, Ma C and Zheng X: MicroRNA-155 in
the pathogenesis of atherosclerosis: A conflicting role? Heart Lung
Circ. 22:811–818. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Raitoharju E, Oksala N and Lehtimäki T:
MicroRNAs in the atherosclerotic plaque. Clin Chem. 59:1708–1721.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Shu W, Yang H, Zhang L, Lu MM and Morrisey
EE: Characterization of a new subfamily of winged-helix/forkhead
(Fox) genes that are expressed in the lung and act as
transcriptional repressors. J Biol Chem. 276:27488–27497. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Bot PT, Grundmann S, Goumans MJ, de Kleijn
D, Moll F, de Boer O, van der Wal AC, van Soest A, de Vries JP, van
Royen N, et al: Forkhead box protein P1 as a downstream target of
transforming growth factor-β induces collagen synthesis and
correlates with a more stable plaque phenotype. Atherosclerosis.
218:33–43. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Mandolini C, Santovito D, Marcantonio P,
Buttitta F, Bucci M, Ucchino S, Mezzetti A and Cipollone F:
Identification of microRNAs 758 and 33b as potential modulators of
ABCA1 expression in human atherosclerotic plaques. Nutr Metab
Cardiovasc Dis. 25:202–209. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ernst O and Zor T: Linearization of the
bradford protein assay. J Vis Exp. 12:pii: 1918. 2010.
|
|
22
|
Abeyesinghe SM, Nicol CJ, Wathes CM and
Randall JM: Development of a raceway method to assess aversion of
domestic fowl to concurrent stressors. Behav Processes. 56:175–194.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Dey BK, Gagan J and Dutta A: miR-206 and
−486 induce myoblast differentiation by downregulating Pax7. Mol
Cell Biol. 31:203–214. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Anderson C, Catoe H and Werner R: MIR-206
regulates connexin43 expression during skeletal muscle development.
Nucleic Acids Res. 34:5863–5871. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Jalali S, Ramanathan GK, Parthasarathy PT,
Aljubran S, Galam L, Yunus A, Garcia S, Cox RR Jr, Lockey RF and
Kolliputi N: Mir-206 regulates pulmonary artery smooth muscle cell
proliferation and differentiation. PLoS One. 7:e468082012.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Song G, Zhang Y and Wang L: MicroRNA-206
targets notch3, activates apoptosis, and inhibits tumor cell
migration and focus formation. J Biol Chem. 284:31921–31927. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Chen X, Yan Q, Li S, Zhou L, Yang H, Yang
Y, Liu X and Wan X: Expression of the tumor suppressor miR-206 is
associated with cellular proliferative inhibition and impairs
invasion in ERα-positive endometrioid adenocarcinoma. Cancer Lett.
314:41–53. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kondo N, Toyama T, Sugiura H, Fujii Y and
Yamashita H: miR-206 Expression is down-regulated in estrogen
receptor alpha-positive human breast cancer. Cancer Res.
68:5004–5008. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Chatterjee A and Hage FG: Guidelines in
review: 2014 ACC/AHA guideline on perioperative cardiovascular
evaluation and management of patients undergoing noncardiac
surgery: A report of the American College of Cardiology/American
Heart Association Task Force on practice guidelines. J Nucl
Cardiol. 22:158–161. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Banham AH, Beasley N, Campo E, Fernandez
PL, Fidler C, Gatter K, Jones M, Mason DY, Prime JE, Trougouboff P,
et al: The FOXP1 winged helix transcription factor is a novel
candidate tumor suppressor gene on chromosome 3p. Cancer Res.
61:8820–8829. 2001.PubMed/NCBI
|
|
31
|
Zhang Y, Li S, Yuan L, Tian Y, Weidenfeld
J, Yang J, Liu F, Chokas AL and Morrisey EE: Foxp1 coordinates
cardiomyocyte proliferation through both cell-autonomous and
nonautonomous mechanisms. Genes Dev. 24:1746–1757. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Caruso P, MacLean MR, Khanin R, McClure J,
Soon E, Southgate M, MacDonald RA, Greig JA, Robertson KE, Masson
R, et al: Dynamic changes in lung microRNA profiles during the
development of pulmonary hypertension due to chronic hypoxia and
monocrotaline. Arterioscler Thromb Vasc Biol. 30:716–723. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Bonnet S and Archer SL: Potassium channel
diversity in the pulmonary arteries and pulmonary veins:
Implications for regulation of the pulmonary vasculature in health
and during pulmonary hypertension. Pharmacol Ther. 115:56–69. 2007.
View Article : Google Scholar : PubMed/NCBI
|