Introduction
Homocysteine (Hcy) is an intermediate product of
methionine demethylation. Hyperhomocysteinemia (Hhcy) has been
associated with an increased incidence of cardiovascular disease,
including peripheral vascular disease, and has been reported to be
a novel independent risk factor for atherosclerosis (AS) (1). The underlying molecular mechanism by
which Hhcy increases the risk of these diseases remains unknown.
The proposed mechanisms include affecting the expression of various
factors that regulate vascular endothelial cell functions and
processes, including oxidative stress, inflammation and immune
responses (1–3). Hcy can increase oxidative stress, which
subsequently leads to the activation of the nuclear factor (NF)-κB
signaling pathway and inflammation (4). This results in endothelial dysfunction
and promotes the development of AS (5). The combined usage of vitamin B6,
Vitamin B12 and folic acid has been demonstrated to significantly
decrease plasma levels of Hcy (6);
however, the mechanism underlying this effect remains unclear.
Two traditional Chinese medicines, produced from
Codonopsis lanceolate and Gleditsia sinensis Lam, are
widely used in the treatment of various inflammation-associated
diseases, and have been demonstrated to exhibit multiple effects,
including antihaemolysis, antitumorigenesis, anti-inflammation,
anti-infection and lowering cholesterol level (7,8).
Echinocystic acid (EA) is a triterpenoid of the β-amyrin family,
which is primarily derived from the rhizome of C. lanceolate
and the fruits of G. sinensis Lam (9). Previous studies have revealed that EA
exhibits anti-inflammatory effects on lysophosphatidic acid-induced
alveolar macrophages and pneumonia in mice through regulating the
NF-κB and mitogen-activated protein kinase signaling pathways
(10–12). In addition, EA has an antiapoptotic
effect, which allows EA to exert a protective effective against the
ischemia-reperfusion injury of cardiomyocytes (13).
Previous studies have demonstrated that cytochrome
P450 1A1 (CYP1A1) is an important enzyme in drug metabolism, lipid
and steroid synthesis, and the metabolism of toxic molecules
(14–18). CYP1A1 metabolizes endogenous
arachidonic acid into hydroxyeicosatetraenoic acid, which has
detrimental roles in cardiovascular disease, such as affecting the
metabolism of arachidonic acid (19). Thus, CYP1A1 may serve a role in Hcy
metabolism (20). CYP1A1 also serves
an antioxidant/protective role against toxicity in the aorta
(21). The present study
investigated whether CYP1A serves a role in the Hcy-induced
alteration of the aorta, such as the effect of CYP1A on the level
of Hcy and arterial injury, and examined the effect of EA on the
morphology of the aorta and serum Hcy levels, in addition to the
potential molecular mechanisms underlying these effects.
Materials and methods
Animals
A total of 50 male specific pathogen-free Sprague
Dawley rats (5 weeks old, 180–200 g) were housed at the animal
facility of Wuhan University Institute of Comparative Medicine
(Wuhan, China). The rats were kept under conditions of 19–25°C with
a relative humidity of 40–60% and light/dark cycle of 12 h. The
present study was approved by the Animal Care Committee of Wuhan
University.
Reagents
EA monomer was obtained from Nanjing Chunqiu
Biotechnology Co. (Nanjing, China; cat. no. AB2013A0001) with a
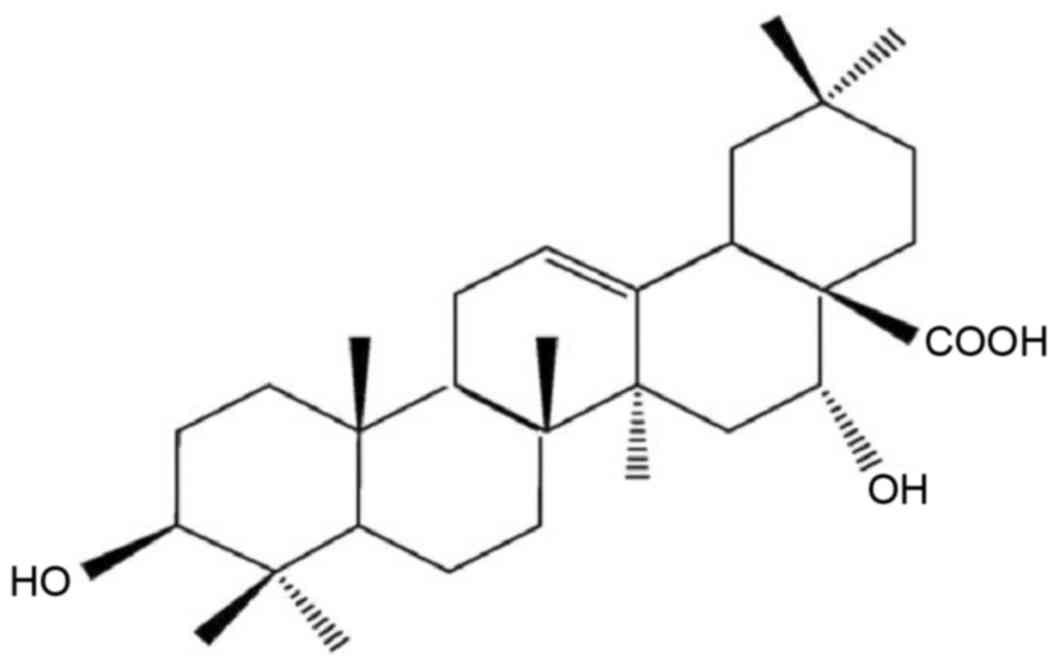
purity of 99.5%. The chemical structure of EA is illustrated in
Fig. 1. Prior to experiments, 0.3%
sodium carboxymethylcellulose (cat. no. F20012203; Sinopharm
Chemical Reagent Co., Ltd, Shanghai, China) was added to the EA
powder to produce a solution of 4 mg/ml EA. High L-methionine diet
was produced by adding 3% L-methionine (Aladdin Reagent Co. Ltd.,
Shanghai, China; batch no. 96934001) to chow diet (purchased from
Wuhan University). All diet was stored in 4°C prior to use.
Reagents for RNA isolation, reverse transcription (RT) and
quantitative polymerase chain reaction (qPCR) were obtained from
Takara Biotechnology Co., Ltd. (Dalian, China), including TRIzol
(cat. no. B4102-1), PrimeScript™ RT reagent kit with gDNA Eraser
(cat. no. AK2501) and SYBR® Premix Ex Taq™ (cat. no.
AK4403). For western blotting, anti-NF-κB (sc-372, 1:1,000),
anti-CYP1A1 (sc-20772, 1:1,000) and anti-β-αctin (sc-130065,
1:1,000) antibodies were obtained from Santa Cruz Biotechnology,
Inc. (Dallas, TX, USA). Secondary antibody horseradish
peroxidase-labeled goat anti-rat IgG (cat. no. GB23302) was
purchased from Wuhan Servicebio Technology Co., Ltd. (Wuhan, China)
and Pierce ECL Western Blotting Substrate was obtained from Thermo
Fisher Scientific, Inc. (Waltham, MA, USA). qPCR primers were
synthesized by Shanghai Shenggong Biology Engineering Technology
Service, Ltd. (Shanghai, China). The sequences of the primers used
for RT and qPCR were as follows: NF-κB forward,
3′-GCTCCTTTTCTCAAGCCGATGT-5′ and reverse,
3′-CGTAGGTCCTTTTGCGTTTTTC-5′ (product size, 220 bp); CYP 1A1
forward, 3′-TCAGGACAGGAGGCTGGACGA-5′ and reverse,
3′-GGATGGTGAATGGGACAAAGGAT-5′ (product size, 293 bp); and β-actin
forward, 3′-TGCTATGTTGCCCTAGACTTCG-5′ and reverse,
3′-GTTGGCATAGAGGTCTTTACGG-5′ (product size, 240 bp).
Groups and establishment of the Hhcy
model
Animals in all groups were fed on a chow diet for 1
week prior to the experiments. Subsequently, a high L-methionine
diet was used to induce Hhcy in the rat as previously described
(22–24). Rats were then randomly divided into
the following groups (n=10/group) for the 8-week-long experiment:
Normal control group (NC; chow diet, daily gastric gavage with
normal saline); model control group (MC; high L-methionine diet,
daily gastric gavage with normal saline); vitamin control group
(VC; high L-methionine diet, daily gastric gavage with folic acid 1
mg/kg + vitamin B2 mg/kg + vitamin B12 10 µg/kg, Aladdin Reagent
Co. Ltd.); EA low dose group (EA1; high L-methionine diet, daily
gastric gavage with 20 mg/kg EA); and EA high dose group (EA2; high
L-methionine diet, daily gastric gavage with 40 mg/kg EA). Body
weight was measured every week in order to adjust the dose of EA
and vitamins accordingly.
Histology
Cardiac tissue samples were fixed at room
temperature with 10% neutral formalin for 5 h. After dehydration
with an increasing series of graded ethanol solutions, specimens
were collected, embedded in paraffin at ~60°C for 5–6 h and then
sectioned (5-µm-thick). The sections were stained with hematoxylin
and eosin (H&E) at 70°C for 30 min. Histological changes were
then observed through a light microscope. Images of five visual
fields were captured for each sample, and the average of thickness
of the media and intima were calculated from five points in each
images.
Determination of plasma Hcy
levels
Animals underwent fasting for 12 h and were weighed,
then 3–5 ml whole blood was drawn from the left ventricle under
anesthesia. The animals were sacrificed and cardiac tissue samples
were harvested. Heparin was used as an anticoagulant after the
blood was drawn (10 IU heparin/1 ml blood). High performance liquid
chromatography was used to measure plasma Hcy levels (25).
RT-qPCR analysis
Total RNA was extracted from the samples using
TRIzol according to the manufacturer's protocol. cDNA was
synthesized using 1 µg of total RNA as the template and specific
primers with the PrimeScript™ RT reagent kit with gDNA Eraser
according to the manufacturer's protocol. qPCR was performed using
the SYBR® Premix Ex Taq™ according to the manufacturer's
protocol (95°C for 30 sec, then 95°C for 5 sec and 60°C for 34 sec,
30 cycles). The PCR products and the concentration of target gene
and reference gene amplification products were obtained by standard
curve analysis software in the computer and calculated using the
2−∆∆Cq method (26). The
expression level of NF-κB and CYP1A1 genes were normalized to the
β-actin.
Western blotting
Total protein was extracted from the aorta using
radioimmunoprecipitation assay buffer [50 mM Tris (pH 7.4), 150 mM
NaCl, 1% Triton X-100, 1% sodium deoxycholate, 0.1% SDS, 2 mM
sodium pyrophosphate, 25 mM β-glycerophosphate, 1 mM EDTA, 1 mM
Na3VO4 and 0.5 µg/ml leupeptin]. Protein
concentrations were determined by BCA protein assay. Samples were
heated at 100°C for 10 min to denature after the addition of 5X
SDS-PAGE sample buffers. Proteins (50 µg/lane) were resolved by 10%
SDS-PAGE gel and transferred to a polyvinylidene difluoride
membrane. After blocking with 5% bovine serum albumin (Shanghai
Liansuo Biological Technology Co. Ltd., Shanghai, China) at 37°C
for 1 h, the membranes were incubated with primary antibodies
anti-NF-κB p65, anti-CYP1A1 (1:1,000) and anti-β-actin (1:5,000) at
4°C overnight. This was followed by a second incubation step with
horseradish peroxidase-conjugated goat anti-rabbit IgG secondary
antibodies (1:12,000) at 37°C for 1 h. Protein bands were then
visualized using the Pierce ECL Western Blotting Substrate
according to the manufacturer's protocol. Quantification of band
density was performed using Quantity One software (version 4.4.0;
Bio-Rad Laboratories, Inc., Hercules, CA, USA).
Statistics
Data are presented as the mean ± standard deviation
of three independent experiments. All data were analyzed using SPSS
software (version 13.0; SPSS, Inc., Chicago, IL, USA). The data
from experiments with multiple groups were analyzed using one-way
analysis of variance and post hoc analysis (Fisher's least
significant difference or Student-Newman-Keuls method). Data were
compared between groups by least significance difference combined
with Tamhane's test. P<0.05 was considered to indicate a
statistically significant difference.
Results
EA attenuates Hhcy-induced changes in
aortic morphology
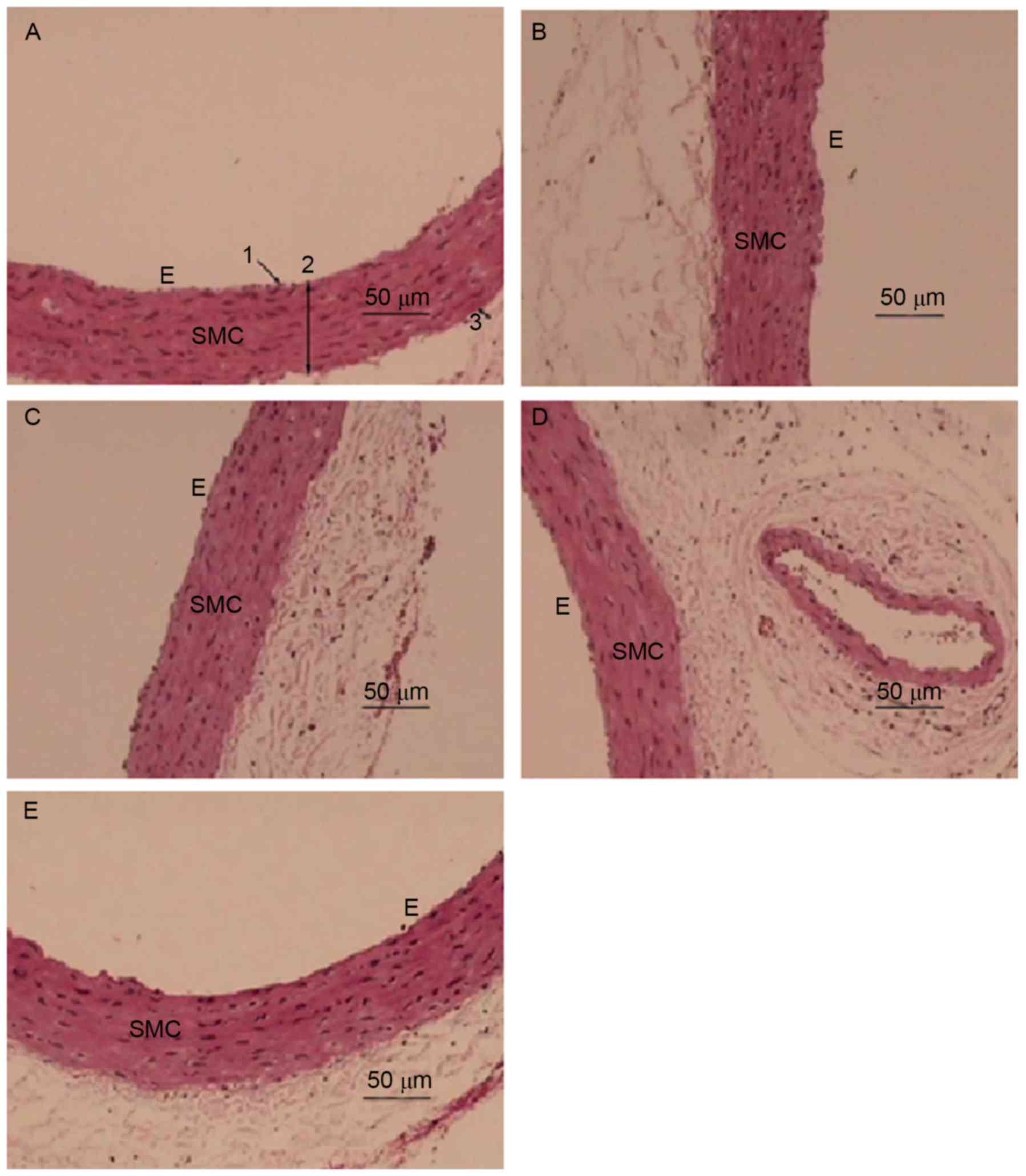
H&E staining was employed to examine the effect
of EA on the morphology of the aorta in Hhcy (Fig. 2). The aorta from the NC group
exhibited an intact endothelial cell lining parallel to the
adventitia (Fig. 2A). The aortic
wall had an equal thickness throughout the entire section, was
lined with smooth muscle cells (SMCs) and possessed endothelial
cells with a normal morphology (Fig.
2A). By contrast, the inner membrane or tunica intima of the
aorta from the MC group was thickened with abundant SMCs lining
(Fig. 2B; Table I). The structure of the aorta media
layer became irregular and thinner in the MC group compared with
that of the control group, with smooth muscle fiber separation and
fracture, characteristic aortic changes of Hhcy, indicating the
successful establishment of a Hhcy model. The inner membrane of the
aorta from the EA1 group still exhibited a slightly disrupted media
layer structure (Fig. 2D). However,
as the dose of EA increased in the EA2 group the inner layer of the
aortic lining exhibited a normal integrity with only slight
thickening and a regular arrangement of SMCs in the media layer
(Fig. 2E), similar to the aorta from
the VC group (Fig. 2C).
 | Table I.Media and intima thickness in the
thoracic aorta. |
Table I.
Media and intima thickness in the
thoracic aorta.
| Group | Media and intima
thickness (µm) |
|---|
| NC | 66.3±1.4 |
| MC |
76.5±2.7a,b |
| VC | 69.1±1.8 |
| EA1 |
66.2±2.2c |
| EA2 |
67.5±1.6c |
EA decreases plasma Hcy levels in
Hhcy
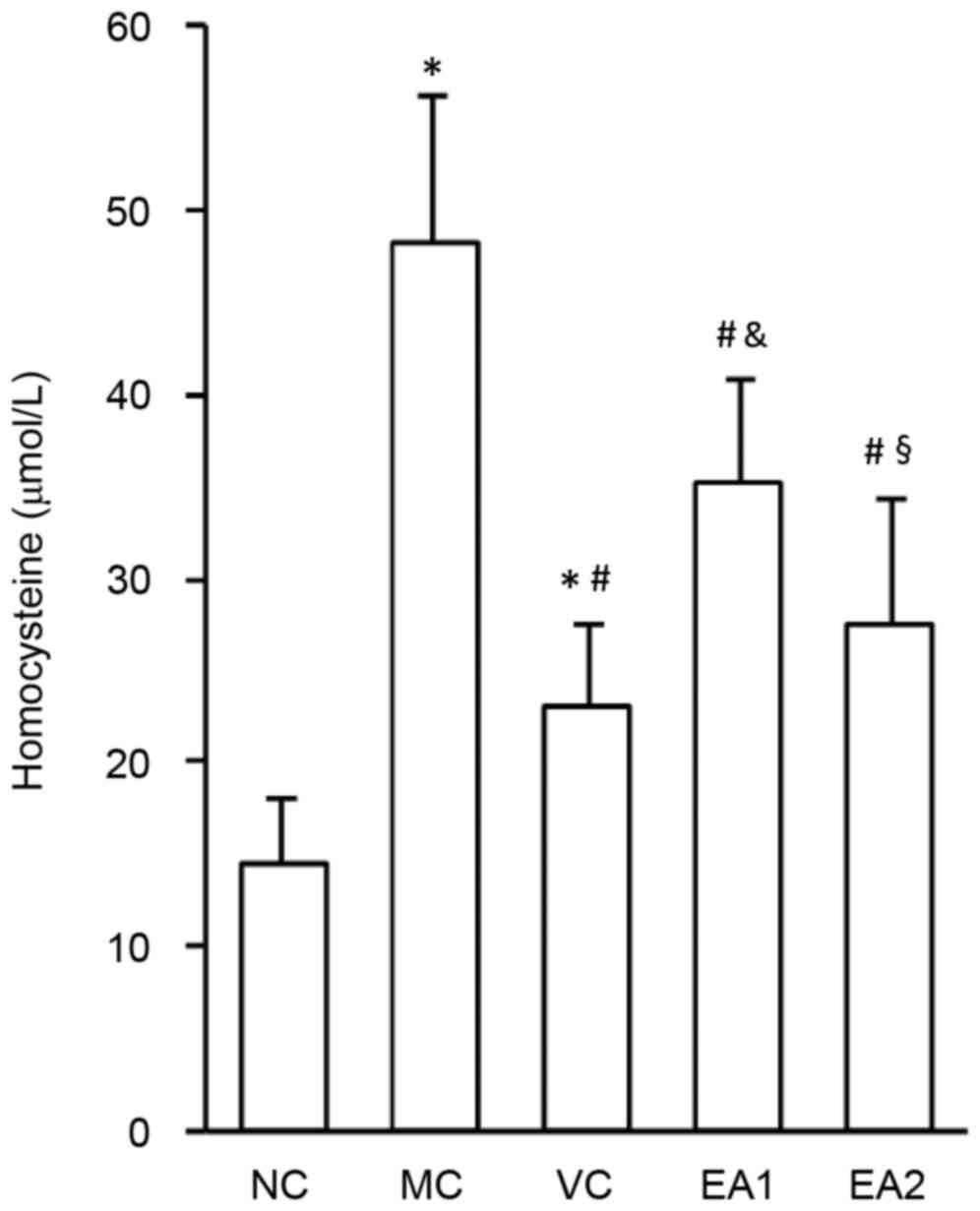
The effect of EA on plasma Hcy levels was
investigated (Fig. 3). Compared with
the NC group, the Hcy levels of rats in the MC, VC, EA1 and EA2
groups were significantly increased (all P<0.05). Serum Hcy
levels in the EA1 were significantly higher compared with that of
the VC group (P<0.05), suggesting a higher efficiency of folic
acid, vitamin B2 and vitamin B12 in lowing plasma Hcy. However, the
effect of EA in lowing plasma Hcy levels was dose-dependent, since
the Hcy level of the EA2 group was significantly lower compared
with that of the EA1 group (P<0.05). There was no significant
difference in Hcy levels between the VC and EA2 groups, indicating
a similar Hcy-lowering effect of high dose EA and vitamins.
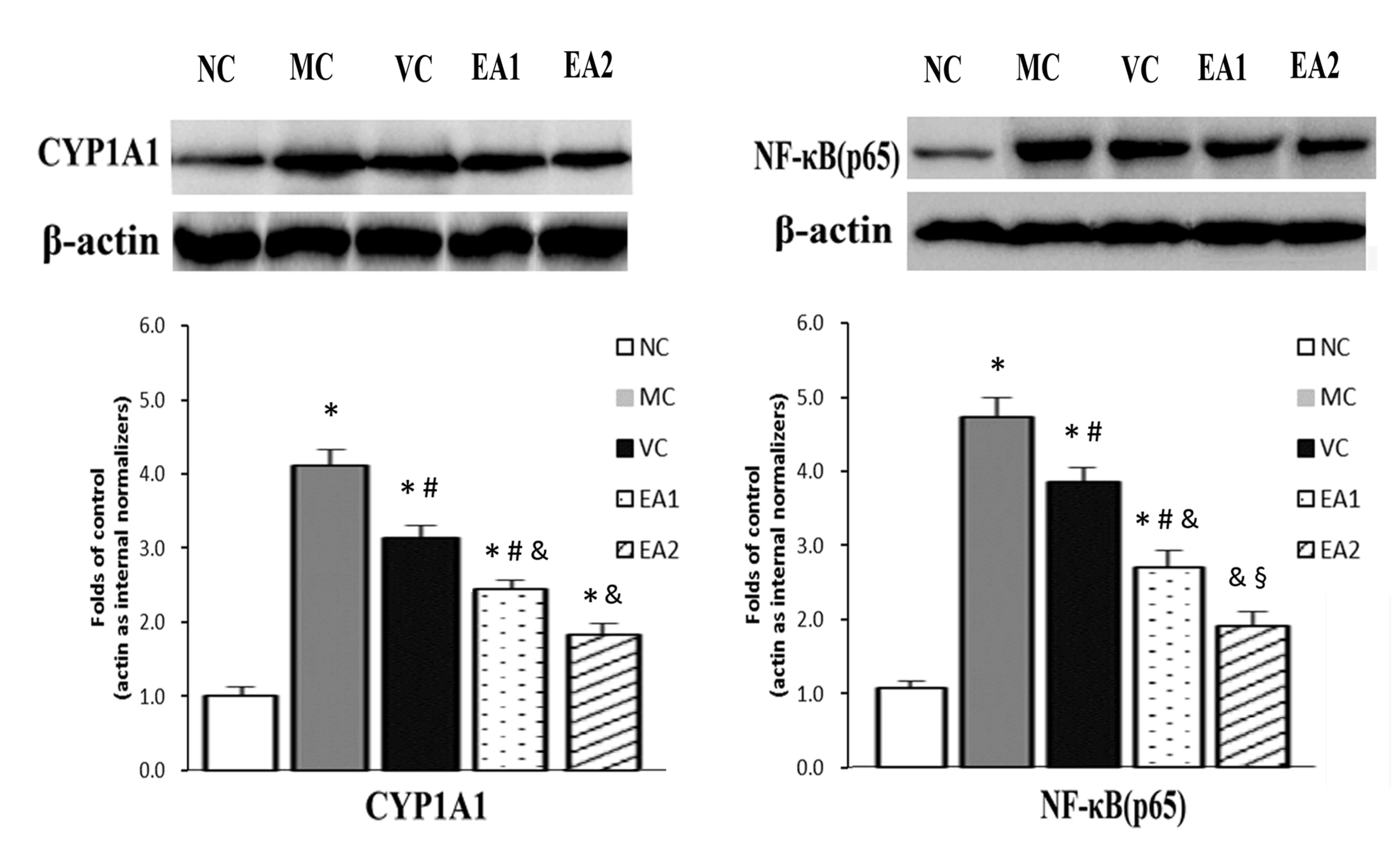
EA decreases the mRNA and protein
expression of NF-κB and CYP1A1 in the aorta in Hhcy
A previous study identified that EA had an
anti-inflammatory effect through regulating NF-κB (8). CYP1A1 is an important enzyme in
catalyzing the metabolism of drugs and toxic molecules, and lipid
and steroid synthesis (13). The
present study investigated the influence of EA on the expression of
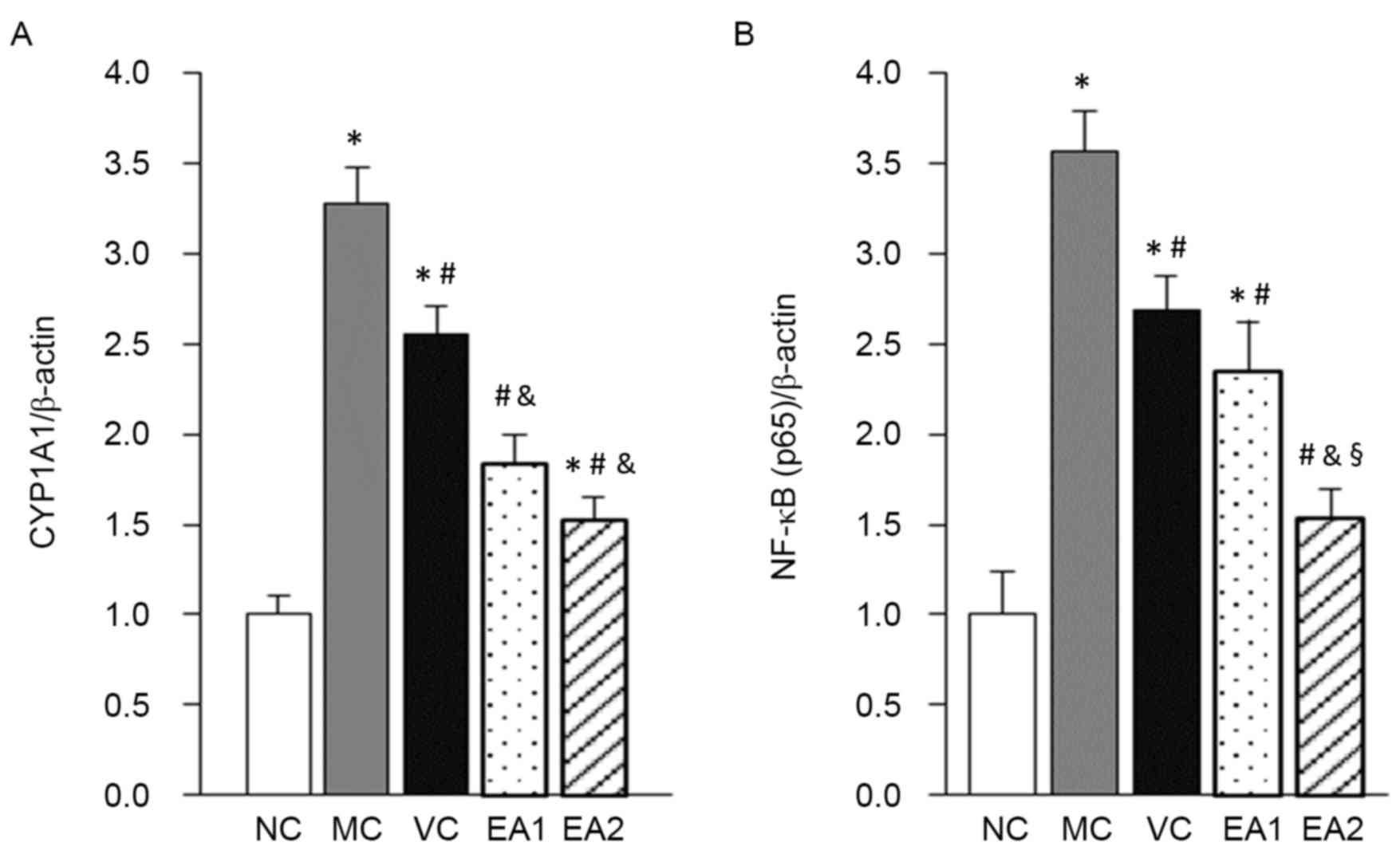
NF-κB and CYP1A1 mRNA using RT-qPCR (Fig. 4). This demonstrated that NF-κB and
CYP1A1 mRNA levels in the MC group were significantly higher
compared with those in the NC group (both P<0.05). The VC, EA1
and EA2 groups had significantly decreased NF-κB and CYP1A1 mRNA
expression levels compared with the MC group (all P<0.05).
Compared with the VC group, the EA1 and EA2 groups had
significantly decreased CYP1A1 mRNA expression (P<0.05). By
contrast, only the EA2 group had a significant decrease in NF-κB
mRNA compared with the VC group (P<0.05).
The protein expression of NF-κB and CYP1A1 was
investigated in the Hhcy model with and without EA treatment
(Fig. 5). Similar to the mRNA
levels, NF-κB and CYP1A1 protein levels were significantly higher
in the MC group compared with the NC group (P<0.05). The VC, EA1
and EA2 groups exhibited significantly lower NF-κB and CYP1A1
protein levels compared with the MC group (P<0.05). EA was
significantly more effective at lowering NF-κB and CYP1A1 protein
levels compared with the vitamin treatment (EA1 vs. the VC group;
EA2 vs. the VC group; both P<0.05). This effect of EA was
dose-dependent, with the EA2 group exhibiting significantly lower
levels of NF-κB and CYP1A1 protein compared with the EA1 group
(P<0.05). In addition, NF-κB protein levels in the EA2 group
were normalized to the NC group level.
Discussion
Hcy is one of the intermediates produced by
demethylation during methionine metabolism and 1% of Hcy exists
freely in the circulation. The remaining 99% of Hcy combines with
different proteins, primarily albumin, in vivo (1). Hhcy refers to a condition of increased
plasma Hcy level, reaching a level that is associated with an
increased risk of cardiovascular disease, including ischemic heart
disease, stroke and peripheral vascular disease (1–3). Hhcy is
determined by various factors, including genetics, nutrition,
medication, disease status, smoking and age. Several large-scale
clinical and epidemiological studies have revealed that Hhcy is an
independent risk factor for AS (27). However, the pathogenesis and
underlying mechanism by which Hhcy increases the risk of AS remains
unclear. Proposed mechanisms include oxidative stress, inflammation
and immune responses, which directly influence vascular endothelial
cell function (28). Therefore,
agents that lower Hcy are potential treatments to prevent the
development of AS.
AS is broadly regarded as a process of chronic
inflammation (29–31). NF-κB is an important transcription
factor in the regulation of the immune response and inflammation,
which affect endothelial function. Endothelial dysfunction
initiates the development of AS (32,33).
Previous results have demonstrated that Hcy can increase oxidative
stress (4), leading to the
subsequent activation of the NF-κB signaling pathway, which induces
inflammation. This causes endothelial dysfunction and promotes the
development of AS (2). Therefore,
regulating the NF-κB signaling pathway could inhibit inflammation
and prevent the development of AS. CYP is a heme-containing
monooxygenase superfamily. CYP family proteins serve a role in the
oxidative metabolism of endogenous and exogenous molecules, which
maintains cardiovascular hemostasis (34). CYP1A1 is highly expressed in vascular
endothelial cells, and there are interactions between PAHs and
CYP1A1 (35). The present results
indicated that the expression levels of Hcy and CYP1A1 of Hhcy rats
were increased, while Hcy level and CYP1A1 in EA group were
decreased, suggesting that there may be some interaction between
Hcy and CYP1A1, and EA could reduce the level of them. CYP1A1
metabolizes endogenous arachidonic acids, including
hydroxyeicosatetraenoic acid, which serves a role in cardiovascular
disease, renal disease, pulmonary sclerosis and anti-platelet
activity (36). A previous study
demonstrated that NF-κB directly regulates CYP1A1 activity together
with a heavy metal ion, indicating that there is an association
between NF-κB and CYP1A1 (37).
The present study identified that plasma Hcy levels
were increased in the Hhcy rat model, indicating successful model
establishment. Hhcy was accompanied with vascular endothelial cell
injury in this model. EA treatment could significantly reduce
plasma Hcy levels, although it was not typically as effective as
the vitamin treatment (vitamin B2, vitamin B12 and folic acid). Van
Mil et al (38) reported that
folic acid reduced levels of the proinflammatory chemokines
monocyte chemoattractant protein 1 and interleukin 8 through
decreasing plasma Hcy levels; however, the authors concluded that
folic acid did not protect against AS development. The current
study identified that EA significantly decreased NF-κB and CYP1A1
protein and mRNA levels in a dose-dependent manner. This indicates
that EA has a protective effect on endothelial cells in Hhcy. In
addition, EA decreased Hcy levels and had an antioxidant effect
through decreasing NF-κB and CYP1A1 expression. However, the
underlying molecular mechanisms through which EA decreases Hcy,
NF-κB and CYP1A1 requires further investigation.
In conclusion, the results of the present study
indicate that EA exerts a protective role on vascular endothelial
cells in Hhcy. EA likely exerts this effect through decreasing
plasma Hcy levels and thus NF-κB and CYP1A1 expression. Future
studies should investigate the underlying molecular mechanisms of
these effects.
Acknowledgements
The present study was supported by the Science and
Technology Development Project of Nanyang City (grant no.
2011GG041).
References
|
1
|
Fowler B: Homocystein-an independent risk
factor for cardiovascular and thrombotic diseases. Ther Umsch.
62:641–646. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lentz SR: Mechanisms of
homocysteine-indued atherothrombosis. Thromb Haemost. 3:1646–1654.
2005. View Article : Google Scholar
|
|
3
|
De Bree A, Verschuren WM, Kromhout D,
Kluijtmans LA and Blom HJ: Homocysteine determinants and the
evidence to what extent homocysteine determines the risk of
coronary heart disease. Pharmacol Rev. 54:599–618. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hidiroglou N, Gilani GS, Long L, Zhao X,
Madere R, Cockell K, Belonge B, Ratnayake WM and Peace R: The
influence of dietary vitamin E, fat, and methionine on blood
cholesterol profile, homocysteine levels, and oxidizability of low
density lipoprotein in the gerbil. J Nutr Biochem. 15:730–740.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Lin CP, Chen YH, Chen JW, Leu HB, Liu TZ,
Liu PL and Huang SL: Cholestin (Monascus purpureus rice) inhibits
homocysteine-induced reactive oxygen species generation, nuclear
factor-kappaB activation, and vascular cell adhesion molecule-1
expression in human aortic endothelial cells. J Biomed Sci.
15:183–196. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Remacha AF, Souto JC, Piñana JL, Sardà MP,
Queraltó JM, Martí-Fabregas J, García-Moll X, Férnandez C,
Rodriguez A and Cuesta J: Vitamin B12 deficiency,
hyperhomocysteinemia and thrombosis: A case and control study. Int
J Hematol. 93:458–464. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Deng YT, Kang WB, Zhao JN, Liu G and Zhao
MG: Osteoprotective effect of echinocystic acid, a triterpone
component from eclipta prostrata, in ovariectomy-induced
osteoporotic rats. PLoS One. 10:e01365722015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Xu LP, Wang H and Yuan Z: Triterpenoid
saponins with anti-inflammatory activity from Codonopsis
lanceolata. Planta Med. 74:1412–1425. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Joh EH, Gu W and Kim DH: Echinocystic acid
ameliorates lung inflammation in mice and alveolar macrophages by
inhibiting the binding of LPS to TLR4 in NF-κB and MAPK pathways.
Biochem Pharmacol. 84:331–340. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Joh EH, Gu W and Kim DH: Echinocystic acid
ameliorates lung inflammation in mice and alveolar macrophages by
inhibiting the binding of LPS to TLR4 in NF-kB and MAPK pathways.
Biochem Pharmacol. 84:331–340. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Joh EH, Jeong JJ and Kim DH: Inhibitory
effect of echinocystic acid on
12-O-tetradecanoylphorbol-13-acetate-induced dermatitis in mice.
Arch Pharm Res. 37:225–231. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ryu S, Shin JS, Jung JY, Cho YW, Kim SJ,
Jang DS and Lee KT: Echinocystic acid isolated from Eclipta
prostrata suppresses lipopolysaccharide-induced iNOS, TNF-α and
IL-6 expressions via NF-κB inactivation in RAW 264.7 macrophages.
Planta Med. 79:1031–1037. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wu J, Li J, Zhu Z, Li J, Huang G, Tang Y
and Gao X: Protective effects of echinocystic acid isolated from
Gleditsia sinensis Lam. against acute myocardial ischemia.
Fitoterapia. 81:8–10. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zou JG, Ma YT, Xie X, Yang YN, Pan S, Adi
D, Liu F and Chen BD: Erratum to: The association between CYP1A1
genetic polymorphisms and coronary artery disease in the Uygur and
Han of China. Lipids Health Dis. 14:1182015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wang XL, Greco M, Sim AS, Duarte N, Wang J
and Wilcken DE: Effect of CYP1A1 MspI polymorphism on cigarette
smoking related coronary artery disease and diabetes.
Atherosclerosis. 162:391–397. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Jarvis MD, Palmer BR, Pilbrow AP, Ellis
KL, Frampton CM, Skelton L, Doughty RN, Whalley GA, Ellis CJ,
Yandle TG, et al: CYP1A1 MSPI (T6235C) gene polymorphism is
associated with mortality in acute coronary syndrome patients. Clin
Exp Pharmacol Physiol. 37:193–198. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Achour B, Barber J and Rostami-Hodjegan A:
Expression of hepatic drug-metabolizing cytochrome P450 enzymes and
their intercorrelations: A meta-analysis. Drug Metab Dispos.
42:1349–1356. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Morel Y and Barouki R: Down-regulation of
cytochrome P450 1A1 gene promoter by oxidative stress. Critical
contribution of nuclear factor 1. J Biol Chem. 273:26969–26976.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Chu ZM, Croft KD, Kingsbury DA, Falck JR,
Reddy KM and Beilin LJ: Cytochrome P450 metabolites of arachidonic
acid may be important mediators in angiotensin II-induced
vasoconstriction in the rat mesentery in vivo. Clin Sci (Lond).
98:277–282. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhou B, He S, Wang XI, Zhen X, Su X and
Tan W: Metabolism of arachidonic acid by the cytochrome P450 enzyme
in patients with chronic Keshan disease and dilated cardiomyopathy.
Biomed Rep. 4:251–255. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Uno S, Sakurai K, Nebert DW and Makishima
M: Protective role of cytochrome P450 1A1 (CYP1A1) against
benzo[a]pyrene-induced toxicity in mouse aorta. Toxicology.
316:34–42. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhang R, Ma J, Xia M, Zhu H and Ling W:
Mild hyperhomocysteinemia induced by feeding rats diets rich in
methionine or deficient in folate promotes early atherosclerotic
inflammatory processes. J Nutr. 134:825–830. 2014.
|
|
23
|
Song S, Kertowidjojo E, Ojaimi C,
Martin-Fernandez B, Kandhi S, Wolin M and Hintze TH: Long-term
methionine-diet induced mild hyperhomocysteinemia associated
cardiac metabolic dysfunction in multiparous rats. Physiol Rep.
3:pii: e12292. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
da Cunha AA, Ferreira AG, da Cunha MJ,
Pederzolli CD, Becker DL, Coelho JG, Dutra-Filho CS and Wyse AT:
Chronic hyperhomocysteinemia induces oxidative damage in the rat
lung. Mol Cell Biochem. 358:153–160. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Pfeiffer CM, Huff DL and Gunter EM: Rapid
and accurate HPLC assay for plasma total homocysteine and cysteine
in a clinical laboratory setting. Clin Chem. 45:290–292.
1999.PubMed/NCBI
|
|
26
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Tamadon MR, Jamshidi L, Soliemani A,
Ghorbani R, Malek F and Malek M: Effect of different doses of folic
acid on serum homocysteine level in patients on hemodialysis. Iran
J Kidney Dis. 5:93–96. 2011.PubMed/NCBI
|
|
28
|
Yang RX, Huang SY, Yan FF, Lu XT, Xing YF,
Liu Y, Liu YF and Zhao YX: Danshensu protects vascular endothelia
in a rat model of hyperhomocysteinemia. Acta Pharmacol Sin.
31:1395–1400. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Ross R: Atherosclerosis-an inflammatory
diseas. N Engl J Med. 340:115–126. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Berliner JA, Navab M, Fogelman AM, Frank
JS, Demer LL, Edwards PA, Watson AD and Lusis AJ: Atherosclerosis:
Basic mechanisms. Oxidation, inflammation, and genetics.
Circulation. 91:2488–2496. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Hossain GS, van Thienen JV, Werstuck GH,
Zhou J, Sood SK, Dickhout JG, de Koning AB, Tang D, Wu D, Falk E,
et al: TDAG51 is induced by homocysteine, promotes
detachment-mediated programmed cell death, and contributes to the
cevelopment of atherosclerosis in hyperhomocysteinemia. J Biol
Chem. 278:30317–30327. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Baird WM, Hooven LA and Mahadevan B:
Carcinogenic polycyclic aromatic hydrocarbon-DNA adducts and
mechanism of action. Environ Mol Mutagen. 45:106–114. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Hockley SL, Arlt VM, Brewer D, Te Poele R,
Workman P, Giddings I and Phillips DH: AHR- and DNA-damage-mediated
gene expression responses induced by benzo(a)pyrene in human cell
lines. Chem Res Toxicol. 20:1797–1810. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Au-Yeung KK, Woo CW, Sung FL, Yip JC and
Siow YLOK: Hyperhomocysteinemia activities nuclear factor-kappaB in
endothelial cells via oxidative stresss. Circ Res. 94:28–36. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Gao M, Li Y, Xue X, Long J, Chen L, Shah W
and Kong Y: Impact of AhR, CYP1A1 and GSTM1 Genetic Polymorphisms
on TP53 R273G Mutations in Individuals Exposed to Polycyclic
Aromatic Hydrocarbons. Asian Pac J Cancer Prev. 15:2699–2705. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Demirdöğen BC, Adali AÇ, Bek S, Demirkaya
Ş and Adali O: Cytochrome P4501A1 genotypes and smoking- and
hypertension-related ischemic stoke risk. Hum Exp Toxicol.
32:483–491. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zordoky BN and El-Kadi AO: Role of
NF-kappaB in the regulation of cytochrome P450 enzymes. Curr Drug
Metab. 10:164–178. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
van Mil NH, Oosterbaan AM and
Steegers-Theunissen RP: Teratogenicity and underlying mechanisms of
homocysteine in animal models: A review. Reprod Toxicol.
30:520–531. 2010. View Article : Google Scholar : PubMed/NCBI
|