Introduction
Novel ways of treating the spinal cord injury (SCI)
have been intensively studied. Among them, the transplantation of
some cell types into the injured area is considered to be a
promising approach (1). The optimal
cellular candidate for the treatment of SCI should be easy to
collect in a non-invasive way, and be both isolated and cultured
without difficulty to reach the number appropriate for
transplantation. Microglial cells appear to be a very attractive
candidate due the possibility of obtaining them during low-invasive
surgical procedures, such as biopsy of the brain, or the
possibility to culture them from bone marrow stem cells (1,2).
The effect of inflammation and immune responses in
the course of damage occurring in the central nervous system (CNS)
gives rise to many controversies. On one hand, it has been
demonstrated that inhibition of the excessive inflammatory reaction
may reduce the extent of damage; on the other hand, there is the
concept of immune neuroprotection, introduced for the first time by
Schwartz et al (1,2). The most important cells involved in
immune neuroprotection are activated microglial cells and
autoreactive cells specific for myelin proteins, including
T-lymphocytes infiltrating the site of injury (3,4). These
cells possess the ability to remove dead cells as well as limit the
size of injury developing during neurodegeneration, thus
stimulating neuronal regeneration (4,5).
Activated microglia produce substances that may stimulate repair
processes in the damaged spinal cord by increasing the survival of
nerve cells and sealing of the blood-brain barrier (5). It has been demonstrated that bone
marrow-derived mesenchymal stromal cells (MSCs), used in MSC
therapy after traumatic brain injury, act as remote ‘bioreactors’
via stimulation of lung macrophages and augmentation of T
regulatory cell production by the spleen, leading to systemic
increases in circulating anti-inflammatory cytokines and alteration
of the locoregional milieu of the CNS (6). The altered intracerebral
microenvironment leads to modulation of the resident microglia
population, stimulating an increase in the ratio of M2
(anti-inflammatory) to M1 (pro-inflammatory) macrophages. This
effect accounts for the observed neuroprotection (6).
Microglial cells were discovered by Pio del
Rio-Hortega (5). There are various
controversies surrounding these cells. It is believed that they
originate from monocyte lines and flow to the brain along with the
development of the vascular system when the blood-brain barrier is
still incomplete and underdeveloped. After reaching the brain
parenchyma, they undergo in situ transformation from an
amebic form into resting microglia (7,8). There
is an alternative theory, ascribing the origin of microglia to a
common progenitor cell for astrocytes and oligodendrocytes present
in the brain (9). Microglial cells
in the embryonic zebrafish brain migrate to an injury site in
response to an SOS signal from damaged neurons. Glutamate is most
likely the strongest inducer of Ca2+-transmitted
microglial attraction to the injury zone (10). Microglial cells are usually uniformly
distributed throughout the whole brain and spinal cord and occur in
an inactive form, representing ~20% of non-neuronal cells in the
brain (7,9). Following a noxious stimulus that may be
of mechanical, chemical or other types, microglial cells are
activated, after which they proliferate and migrate to the site of
injury (4). Activated in a classical
β-amyloid method, microglial cells not only secrete large numbers
of pro-inflammatory and neurotoxic factors, but also produce some
substances of known anti-inflammatory and even neuroprotective
functions, including interleukin (IL)-10, IL-11 and fibroblast
growth factor (11). The
pro-inflammatory factors produced by activated microglia may be
enumerated as: IL-1, tumor necrosis factor (TNF) α, IL-6, IL-12,
IL-15, IL-18, chemokines (IL-8 and interferon γ-induced protein-10)
and cytotoxic compounds (inducible nitric oxide synthase, free
radicals of oxygen and nitrogen) and prostanoids (12).
Currently, it appears that the CNS of adult mammals
is able to initiate signals that alter the function of microglia,
and vice-versa, and these cells in turn release factors that
regulate neuronal function, including neurogenesis (10). Previous research has demonstrated
that activated microglia or blood-born macrophages accumulate in
the lesions of injured spinal cords, and they may influence
survival of neurons in various ways (2). It is understood that a tightly and
timely regulated immune response is required for recovery.
Furthermore, it is now apparent that the phenotype of microglia is
not uniform, and that in response to different stimuli these cells
may acquire either destructive or rather beneficial phenotypes
(2). Therefore, transplantation of
properly activated, stage-dependent microglial cells may have a
crucial role in the process of regeneration of the nervous system.
These ideas could be achieved in different ways. For example, a
study by Yaguchi et al (13)
indicated that IL-12 administration (secreted by dendritic cells)
or direct dendritic cells transplantation into injured adult spinal
cords resulted in functional recovery through the activation of the
microglia/macrophage system in mice. In the present experiment, the
influence of in vitro activated microglial cells on the
injured spinal cords of adult rats was assessed.
Materials and methods
Animals
All procedures used in the present experiment were
performed in accordance with the EU Animal Protection Law and were
approved by the Local Ethics Committee for Experiments on Animals
in Katowice (Katowice, Poland). A total number of 36 adult rats and
45 pups obtained from Experimental Medicine Center of Medical
University of Silesia, (Katowice, Poland) were used in the present
experiment. Animals were housed in individual cages with a 12-h
light/dark cycle, at 22°C and 40% humidity, with access to food and
water ad libitum. Of the adult rats, 30 male Wistar C rats
(weight, ~0.3 kg, age 12–14 weeks) were randomly divided into two
equal (n=15) groups: Control (C) and experimental (M). For magnetic
resonance imaging (MRI) examination, an additional group (n=6) of
healthy male rats (without any procedures) was used as a reference
(R) group for the C and M groups. In addition, 45 2-day old male
Wistar C rat pups (b.w. 7–9 g) were used to prepare glial
cultures.
SCI procedure
The focal SCI was performed using a device that we
designed and created, the pressure impactor, which produces a
precisely controlled air blast, as described previously (14). Briefly, following intraperitoneal
anesthesia with ketamine (100 mg/kg) and xylazine (10 mg/kg; both
POLFA Łódź, Warszawa, Poland), animals were immobilized, Th9-Th11
segments of spinal cord were exposed and, under control of a
stereomicroscope (Nikon SMZ 800; Nikon Corporation, Tokyo, Japan),
a 2-mm hole in the right Th10 vertebral arch was drilled. Special
care was taken not to injure the dura, and for thermal protection,
trepan was cooled down with chilled phosphate-buffered saline
(PBS). Subsequently, the impactor tip was placed within the hole,
contacting the dura mater but exerting no pressure on it. A single
air blast (150 kPa pressure; 0.1 sec duration) was applied. The air
blast shot was observed under the stereomicroscope and recorded by
the attached camera (Invenio 5D; Nikon Corporation). After
completing the procedure, the hole was secured with bone wax,
muscles were sutured in layers, skin was closed and the wound was
treated with a sterile bandage. This procedure was performed on all
adult rats in the C and M groups.
To avoid dehydration, all rats were subcutaneously
injected with 2 ml sterile saline. As the autonomic function of the
urinary bladder was impaired due to the spinal shock, rat bladders
were emptied manually twice daily until the recurrence of bladder
function. To prevent pain, 400 mg paracetamol (suspension 125 mg/5
ml; POLFA Łódź) was dissolved in 100 ml drinking water (average
drug dosage, 200 mg/kg body weight daily).
Cell cultures
Primary mixed glial cultures were prepared from 45
2-day-old Wistar rat pups, as described previously (15). Briefly, following decapitation the
brains were excised aseptically and separated from the blood
vessels and meninges on ice. Cerebral cortical tissue was
dissociated by trituration in ice-cold Dulbecco's modified Eagle
medium (DMEM; 4.5 g glucose/l) supplemented with 10%
heat-inactivated fetal bovine serum (FBS), 2 mM glutamine, 100
UI/ml penicillin, 100 µg/ml streptomycin and 5 µg/ml fungizone (all
Invitrogen; Thermo Fisher Scientific, Inc., Waltham, MA, USA). The
suspension was filtered sequentially through two cell strainers
with 70- and 40-µm meshes (BD Biosciences, Franklin Lakes, NJ,
USA). Dissociated cells were plated (20×106 cells/dish)
on poly-D-lysine-coated 100-mm Petri dishes (BD Biosciences) and
incubated at 37°C in humidified 5% CO2/95% air
(CO2 incubator; Heraeus, Hanau, Germany). The medium was
replenished 1 day after plating and changed every 3 days
thereafter. Subsequent to plating, the cells were cultured for
13–15 days until confluence. Poly-D-lysine was purchased from
Sigma-Aldrich (Merck KGaA, Darmstadt, Germany).
To identify astrocytes, the cultures were fixed in
cold 99.8% methanol for 10 min at 4°C (Sigma-Aldrich; Merck KGaA)
and labeled immunocytochemically with glial fibrillary acidic
protein (rabbit anti-GFAP; catalogue no. ab7260; 1:500 dilution,
overnight at 4°C), which is a specific marker for astrocytes, and
secondary anti-rabbit antibody (IgG, catalogue no. ab7090, dilution
1:10,000 for 1 h at 20°C; both Abcam, Cambridge, MA, USA). Analysis
with light microscope (magnification ×400; Olympus BH2, Olympus
Corporation, Tokyo, Japan) of these mixed glial cultures
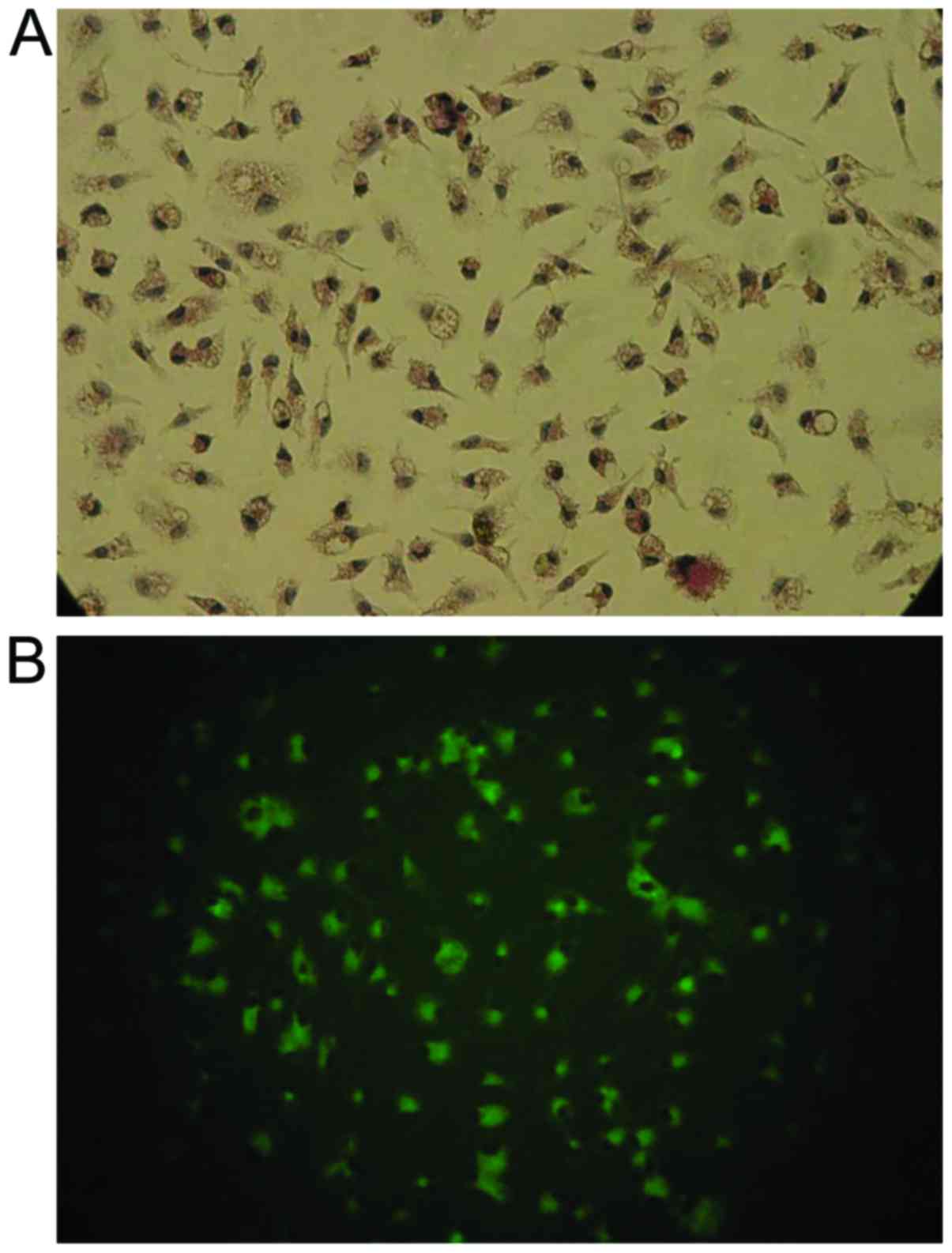
demonstrated that 70–75% of the cells were GFAP-positive. Of the
cultured cells, ~20% reacted with Ricinus communis
agglutinin-1 (RCA-1; Vector Laboratories, Inc., Burlingame, CA,
USA), a marker for microglia (examined under 488 nm fluorescence,
microscope and magnification as above). No neurons were detected,
as confirmed using light microscopy after immunostaining of cells,
fixed as described, with a monoclonal antibody against microtubule
associating protein-2 (rabbit anti-MAP-2; catalogue no. ab32454;
1:500 dilution, overnight at 4°C and IgG anti-rabbit secondary
antibody at a dilution of 1:10,000 for 1 h at 20°C (Abcam).
Rat microglial cultures were obtained by shaking the
primary mixed glial cultures at 200 r/min for 5 h (37°C), with
maximum yields between days 12 and 14. The suspension of floating
cells was filtered through a 40-mm nylon mesh, centrifuged at 200 ×
g for 10 min, suspended in 200 µl DMEM supplemented with 10% FBS,
plated in 96-well tissue culture plates (5×104
cells/well) and incubated at 37°C for 15 min in humidified 5%
CO2/95% air. Subsequently, the wells were vigorously
washed three time with 200 µl culture medium to remove non-adherent
cells. Microglial cells, which firmly adhered to the bottom of the
well, were incubated at 37°C overnight. To stimulate the microglial
cells, 1 µg/ml lipopolysaccharide (LPS; Sigma-Aldrich; Merck KGaA)
was added for 24 h at 37°C. The effectiveness of stimulation was
determined by measuring the concentration of IL-1β (catalogue no.
SRLB00), IL-6 (catalogue no. SR600B) and TNF-α (catalogue no.
SRTA00) in the culture medium using rat ELISA kits (Quantikine
ELISA Kits; R&D Systems, Inc., Minneapolis, MN, USA), according
to manufacturer's recommendations. Absorbance was measured using a
microplate reader at a wavelength of 450 nm. The limit of the assay
was 4.4 pg/ml for IL-1β, 19 pg/ml for IL-6 and 15 pg/ml for TNF-α.
The intra-assay coefficients of variability for all cytokines were
<10%. After 24 h of LPS stimulation, the medium was removed and
the cells were dissociated by trituration in ice-cold PBS.
Microglial suspensions were filtered through 40-µm meshes, adjusted
to 1×106 cells/ml and a constant number of microglia
were thus prepared for implantation.
To differentiate transplanted cells, cells were
labeled with freshly prepared 2 µM/l Cell Tracker CM-DiI solution
(Molecular Probes; Thermo Fisher Scientific, Inc.) at 37°C for 5
min, and at 4°C for 15 min. Excess tracer was washed away with PBS
and cells were suspended in fresh culture medium to obtain ~300
cells in 10 µl. Cell viability was determined by assessing membrane
integrity in the microglial cultures using a 0.1% trypan blue
exclusion test and MTT test with a final MTT concentration of 2.5
µg/ml in PBS. Dimethyl sulfoxide was used as the solvent
(Invitrogen; Thermo Fisher Scientific, Inc.), measured at a
wavelength of 570 nm (xMark Microplate Spectrophotometer; Bio-Rad
Laboratories, Inc., Hercules, CA, USA), as previously described
(16). More than 97–98% of the
cultured cells were living, 95% reacted with RCA-1 (microglial
cells), and only 2–3% were GFAP-positive (astrocytes) (17). All intervals and compound
concentrations used in the present study were within the limits
used in in vitro experiments considering microglia and LPS
(15,18).
Cell transplantation
In group M, cultured LPS-activated and DiI-labeled
microglial cells (~300 cells/injection) were injected twice into
the site of injury. The initial injection was administered
immediately after SCI and the second injection was administered 24
h later. The dura mater was incised with the tip of an injection
needle, exposing the surface of the injured spinal cord. For each
injection, 10 µl microglial cell suspension (group M) or saline
(group C) was slowly injected, at an angle of 45°, into the dorsal
spinal cord next to the middle line at the lesion site using a
spinal stereotaxic frame (manufactured at Department of Physiology,
School of Medicine in Katowice, Katowice, Poland) by means of a
glass pipette connected to a Hamilton syringe with a tip outer
diameter of 100 µm. In order to confirm that transplanted
microglial cells were delivered successfully into the injured
spinal cord, spinal cord sections collected from a small set of
animals (n=2) 24 h post-transplantation were examined using a
fluorescent microscope (Labophot 2; Nikon Corporation) under a 568
nm wavelength to visualize exogenous DiI-labeled microglia cells
present in the spinal cord.
Assessment of locomotor function
During 12 weeks after the surgical procedure,
animals were observed and analyzed regarding development and/or
regression of neurological deficits. Behavioral observations
included gait analysis (foot print test) and open field tests,
according to the Basso, Beattie and Bresnaham (BBB) scale (19).
Foot print tests were conducted once in
postoperative weeks 1, 4, 7 and 12. Animals were tested on a 100-cm
long and 7-cm wide runway with side walls and a transparent bottom.
The walk of a rat through the runway was recorded with a digital
camera and was automatically analyzed frame by frame (Catwalk XT
8.1; Noldus Information Technology, Wageningen, The Netherlands).
Each animal was tested three times, consecutively. Framed foot
images were analyzed regarding the foot rotation angle (the angle
made by two lines connecting the third toe and the stride line at
the center of the paw) of the right hind paw (ipsilateral to the
injury site), and interlimb coordination (smallest distance between
the middle point of the hind paw and forepaw on the same side).
Normal rotation angle is ~8° and after trauma this increases up to
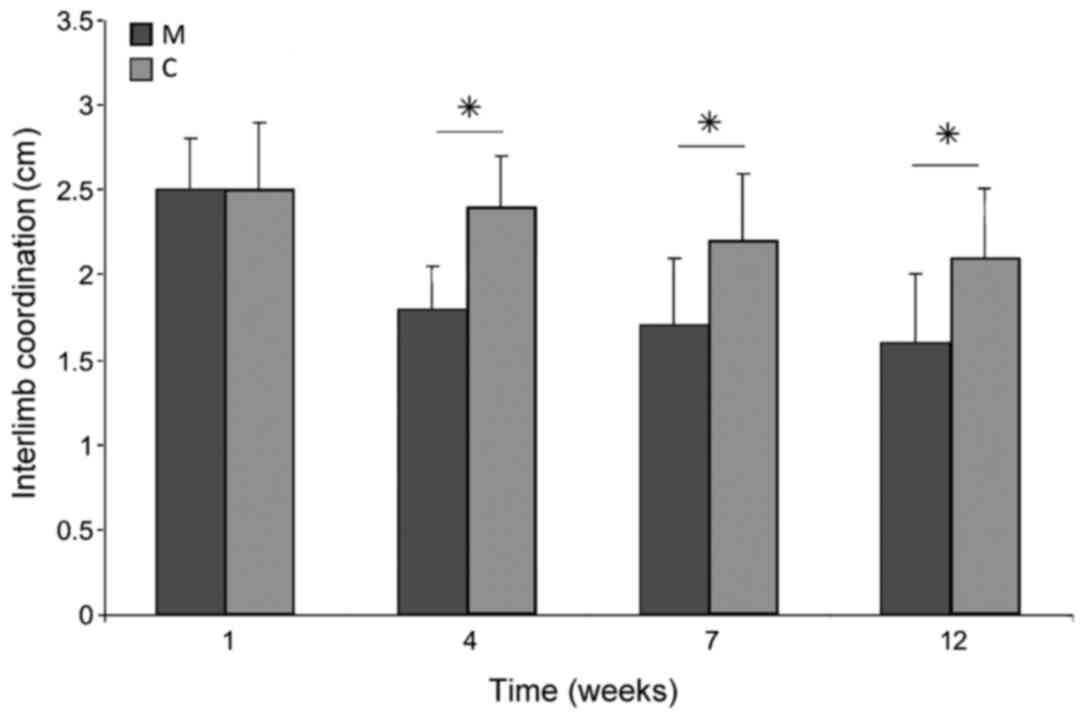
30° (20). Interlimb coordination is
~1.5 cm in healthy rats and increases to 3 cm, or more, following
injury (20).
The BBB open field test was carried out in a
classical way on a plexiglass surface (19). During the test, motor function of
joints was analyzed to assess the stepping ability of the rats.
Additionally, general coordination and stability of the body were
evaluated. The value range was 0–22, where 0 was related to
complete lack of motor capability and 22 indicated the best score
in this aspect. Obtained results were average values from both hind
extremities.
Neuroanatomical tracing
Retrograde neuroanatomical tracing was used to
determine the extent to which supraspinal neurons were able to
regrow axons to reach spinal cord segments caudal to the injury. A
total of 4 rats from each group were randomly selected. One week
before the end of the experiment, rats were anesthetized
intraperitoneally with ketamine (100 mg/kg) and xylazine (10
mg/kg), the spinal cord was exposed by laminectomy below the injury
site and two microcrystals of FluoroGold (FG; Fluorochrome Inc.,
Englewood, NJ, USA) were placed bilaterally inside the spinal cord,
10-mm caudally from the injury site. For quantification of the
number of FG-positive neurons present in the brain stem (red
nucleus) and primary motor cortex, whole brains were carefully
dissected, embedded in TissueTek (Sakura Finetek Europe B.V,
Flemingweg, The Netherlands), frozen at −80°C and then cut
coronally into 10-µm sections. Every sixth slide was analyzed,
giving 18–22 sections per rat. The sections were viewed under a
fluorescent microscope (Labophot 2; Nikon Corporation) at 365 nm
and imaged at a magnification of ×400. FG-positive cells were
counted in five constant sites (center and four corners of the
image). Finally, the number of FG-positive cells in the brain stem
and motor cortex per mm2 was calculated.
MRI analysis
MR images were obtained at the Institute of Nuclear
Physics, Polish Academy of Sciences (Kraków, Poland) using a 4.7 T
research MRI scanner, equipped with MARAN DRX digital console
(Resonance Instruments, Ltd., Witney, UK). A measuring probe-head
with surface radiofrequency coil dedicated for rat spinal cord
investigations was used. Animals were anesthetized with halothane
(2%) mixed with air (60%/1.2 l/min) and oxygen (40%/0.8 l/min).
Physiological conditions of the animal, including temperature,
breathing rate and cardiac rate, were continuously monitored during
examination using an MR-compatible Small Animal Monitoring &
Gating System (SA Instruments, Inc., Stony Brook, NY, USA).
A spin echo pulse sequence was used for acquiring
T2-weighted and diffusion-weighted MR images in sagittal
and axial projections. The following parameters of the spin echo
sequence were applied: Echo time, 32 msec; repetition time, 1.8
sec; field of view, 2.5 cm; image matrix size, 128×128; and slice
thickness, 1.5 mm. The signal was accumulated [number of
acquisitions (NA)=6] in order to improve the signal-to-noise ratio
(SNR).
Diffusion-weighted images were obtained with
additional diffusion sensitive gradients (δ=5 msec and Δ=23 msec)
added to the spin echo sequence in transversal and longitudinal
directions to the spinal cord axis (21,22). In
order to optimize the SNR, thus minimizing the examination time,
different values of b-factor were used to assess the effect
of water diffusion on the echo signal: 750 and 1,100
sec/mm2 for longitudinal and transversal directions,
respectively (14,23). The acquisition was both cardiac- and
respiratory-gated in order to eliminate motion artifacts.
Histomorphological analysis
After 1 (n=5 for each group) and 12 weeks (n=6 for
each group), respectively, rats were re-anesthetized and perfused
transcardially with 100 ml PBS (pH 7.4) followed by 100 ml 4%
paraformaldehyde solution in the same buffer. Fragments of spinal
cord (~2 cm) incorporating the injured area were dissected and
dehydrated in 20% sucrose in PBS for 24 h at 4°C. Spinal cords were
then embedded in TissueTek, frozen at −80°C, cut sagittally or
transversally into 10-µm sections and mounted on SuperFrost Plus
slides (Menzel-Glaser; Thermo Fisher Scientific, Inc.) in a
10-section step manner. Slides were subjected to routine
hematoxylin and eosin (H&E) staining at room temperature:
H&E staining for 5 min, followed by a 15 min tap-water wash,
and 2 min eosin staining; 1% toluidine staining at room temperature
for 15 min; or immunohistochemical labeling. The following anti-rat
antibodies were used (all Chemicon®; EMD Millipore,
Billerica, MA, USA): Rabbit anti-OX-42 (for microglia and
macrophages; catalogue no. CBL1512; dilution 1:100), mouse ED-1
(catalogue no. MAB1435; dilution 1:100) and rabbit GFAP (for
astrocytes; catalogue no. AB5804; dilution 1:500). The sections
were incubated with primary antibodies overnight at 4°C, and after
washing three times with PBS, the sections were treated for 2 h at
room temperature in a dark chamber with secondary goat anti-rabbit
immunoglobulin (Ig G or anti-mouse IgG antibodies conjugated with
AlexaFluor 488 and/or AlexaFluor 568 (Molecular Probes; Thermo
Fisher Scientific, Inc.; goat Anti-Mouse IgG AlexaFluor 488,
catalogue no. A11001; goat Anti-Mouse IgG AlexaFluor 568, catalogue
no. A11004; A11008 for goat Anti-Rabbit IgG AlexaFluor 488,
catalogue no. A11008; goat Anti-Rabbit IgG AlexaFluor 568,
catalogue no. A11011; dilution 1:500). Coverslips were placed over
the sections with VectaShield and DAPI (Vector Laboratories, Inc.,)
and sections were examined under a confocal laser scanning
microscope (FluoView; Olympus Corp., Tokyo, Japan) at a
magnification of ×400. The images were digitally stored and then
analyzed. Maximal distance of expansion of implanted microglia in
the injured spinal cord was measured 1 week after surgery and
survival of transplants (number of cells seen in the epicenter of
the injury) was examined 1 and 12 weeks following surgery,
respectively. Sizes of lesions (cavity area) were also measured 12
weeks after surgery.
Statistical analysis
Analysis of variance (ANOVA), two-way repeated
measures and/or one-way ANOVA with Tukey's post hoc tests [honest
significant difference (HSD) or unequal N HSD, where N is the
number of samples used] or nonparametric Kruskal-Wallis ANOVA were
used to analyze normally distributed data or non-continuous data,
respectively. Differences in foot print, histological and BBB
scores between group means at each time point post injury were
identified using Student's t-tests. STATISTICA, version 10 software
(StatSoft; Dell, Round Rock. TX, USA) was used. P<0.05 was
considered to indicate a statistically significant difference. All
data were expressed as the mean ± standard deviation.
Results
Histopathological findings
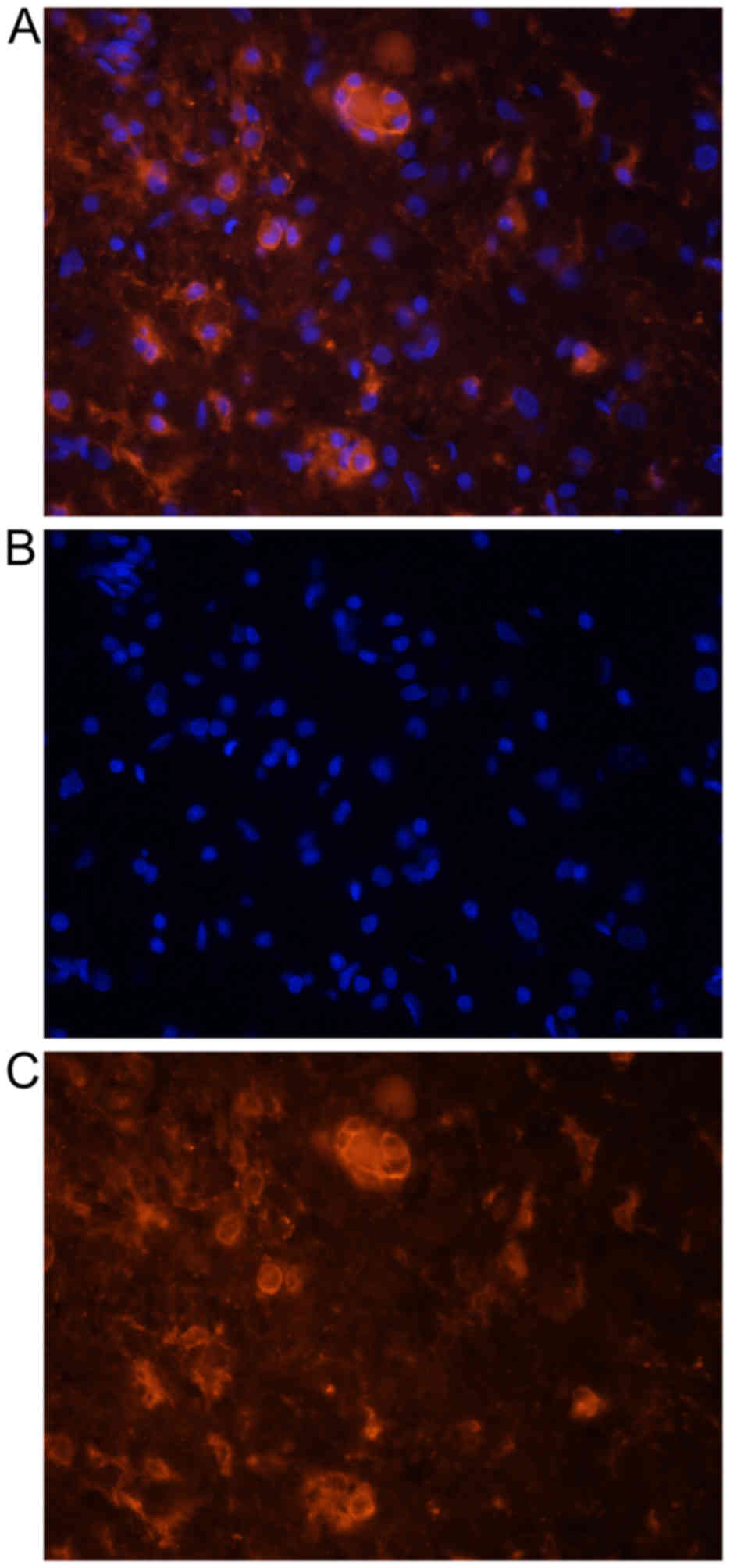
Transplanted activated microglial cells, as
confirmed by double labeling with DiI and OX-2 (Fig. 1), were observed 1 week after
transplantation inside the spinal cord in the closest vicinity of
injury and injection site (expansion, 2.7±1.3 mm). They formed
microsphere-like structures (Fig.
2). Their number rapidly decreased with time; 1 week after
surgery 371.1±63.7 cells/mm2 were observed, while at the
end of the experiment, no cultured microglia were present inside
the spinal cord.
The average length of the lesions measured on
H&E-stained sections in group M (2.9±0.13 mm) was significantly
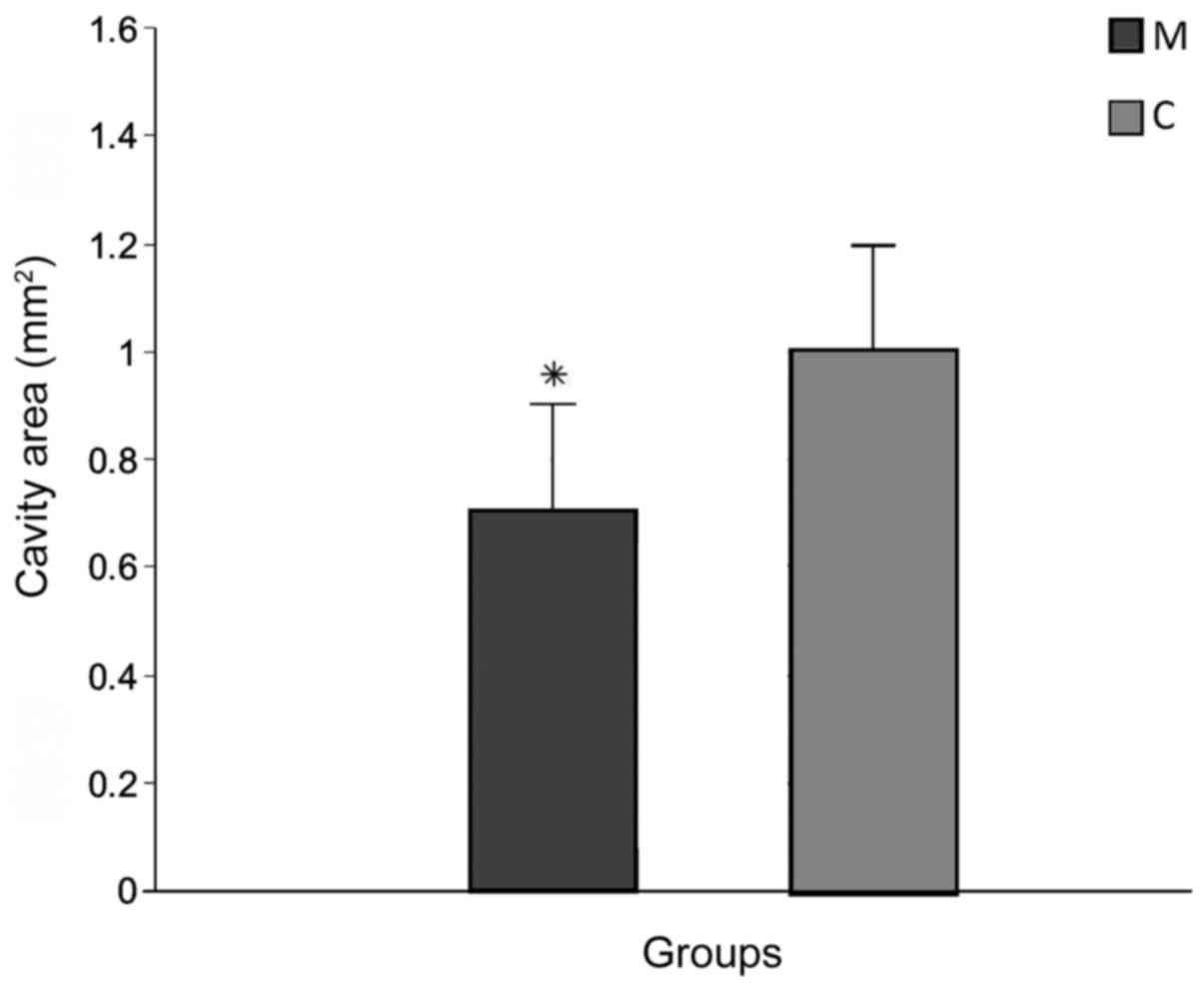
smaller than group C (6.32±2.41 mm; P<0.05; data not shown). The
cavity area of the main lesion was also significantly decreased in
group M (0.7±0.08 mm2) compared with group C (1.02±0.21
mm2; P<0.05; Figs. 3
and 4A).
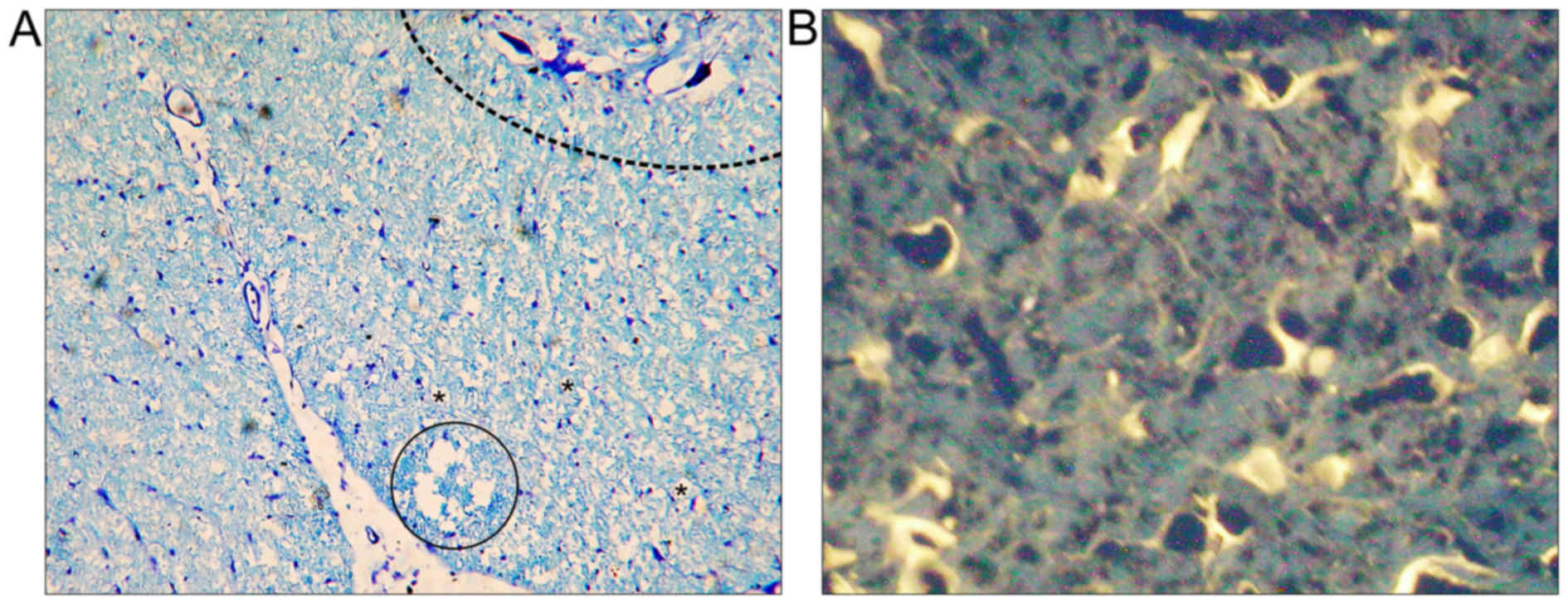
Neuroanatomical tracing
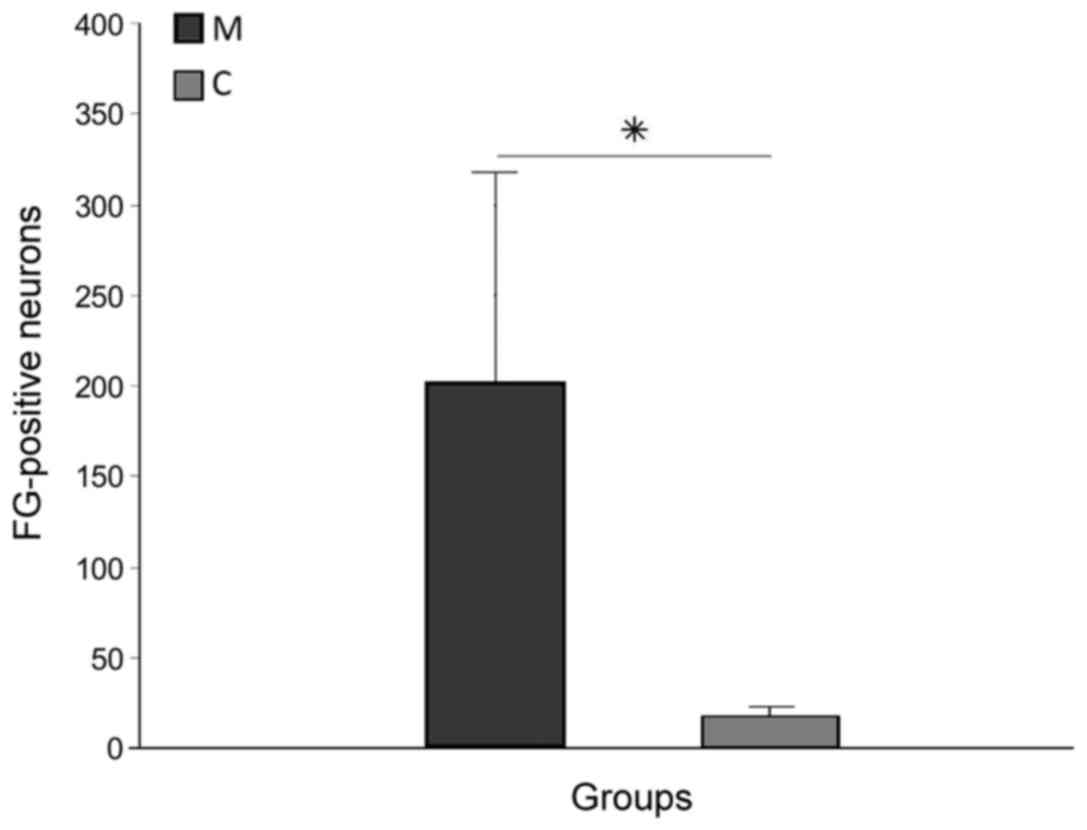
FG-positive cells in the brain stem (Fig. 4B) and primary motor cortex,
demonstrating survival of long-track motor neurons, were more
numerous in group M (203.7±123.3) than in group C (14.3±3.3), and
this difference was significant (P<0.05; Fig. 5). Tracer spread through the blood was
absent in all rats, as confirmed by microscope observation during
the application procedure.
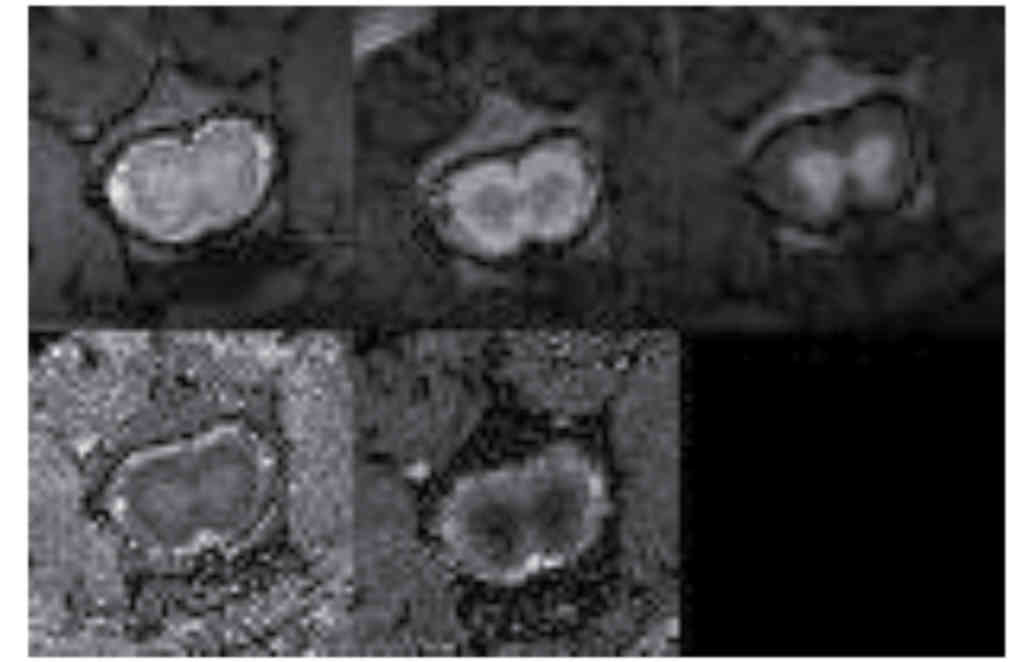
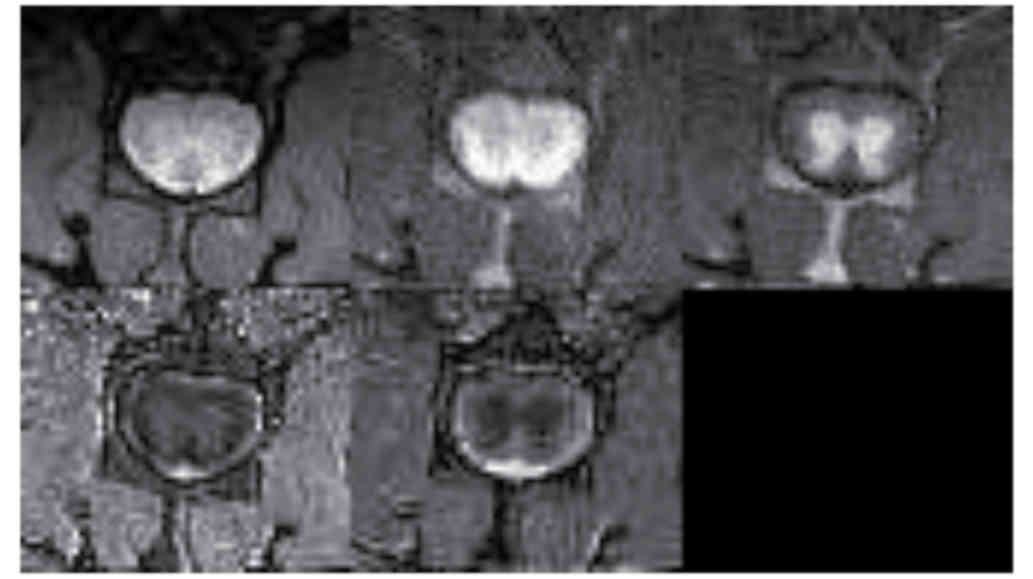
MRI analysis
Fig. 6 demonstrates
MRI images and reconstructed parametric maps from representative
spinal cords in group R. Fig. 7
demonstrated corresponding images from a representative injured
spinal cord treated with microglia from group M. The average values
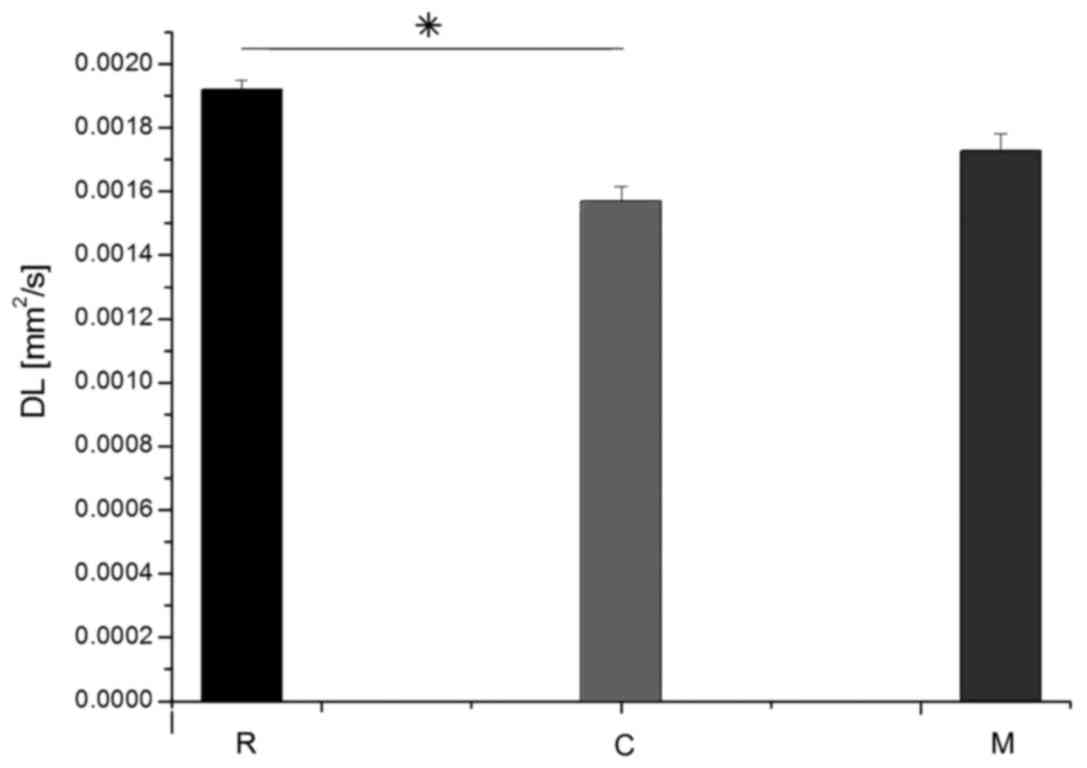
of longitudinal and transverse diffusion coefficients (DL and DT,
respectively) for the M, C and R groups are demonstrated in
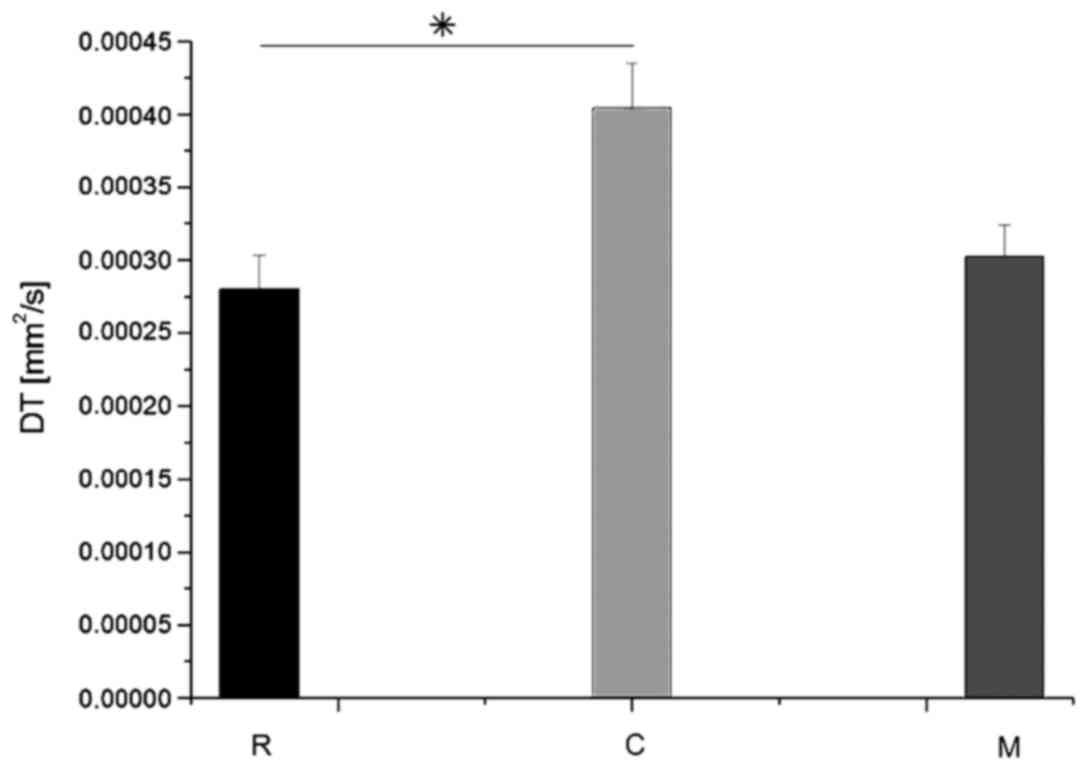
Figs. 8 and 9. The average DL value in group C (Fig. 8) was significantly lower than in the
R group (Tukey's unequal N HSD test; P=0.0012). No significant
differences were observed between the average DL value in the M and
C groups or between the average DL value in the M and R groups. The
average DT value (Fig. 9) in group C
was significantly higher (Tukey's unequal N HSD; P=0.027) than in
group R, while the remaining comparisons demonstrated no
significant differences, similarly as for the DL coefficient. For
both parameters, average values demonstrated in group M were closer
to the group R values.
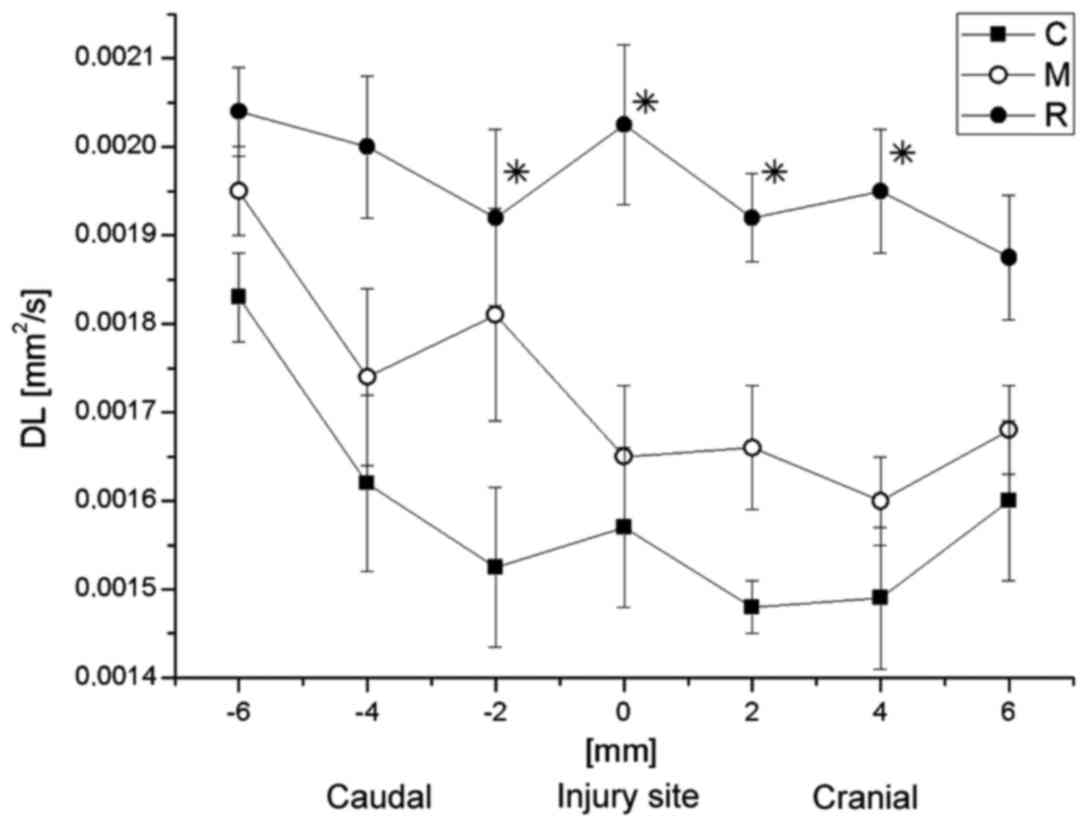
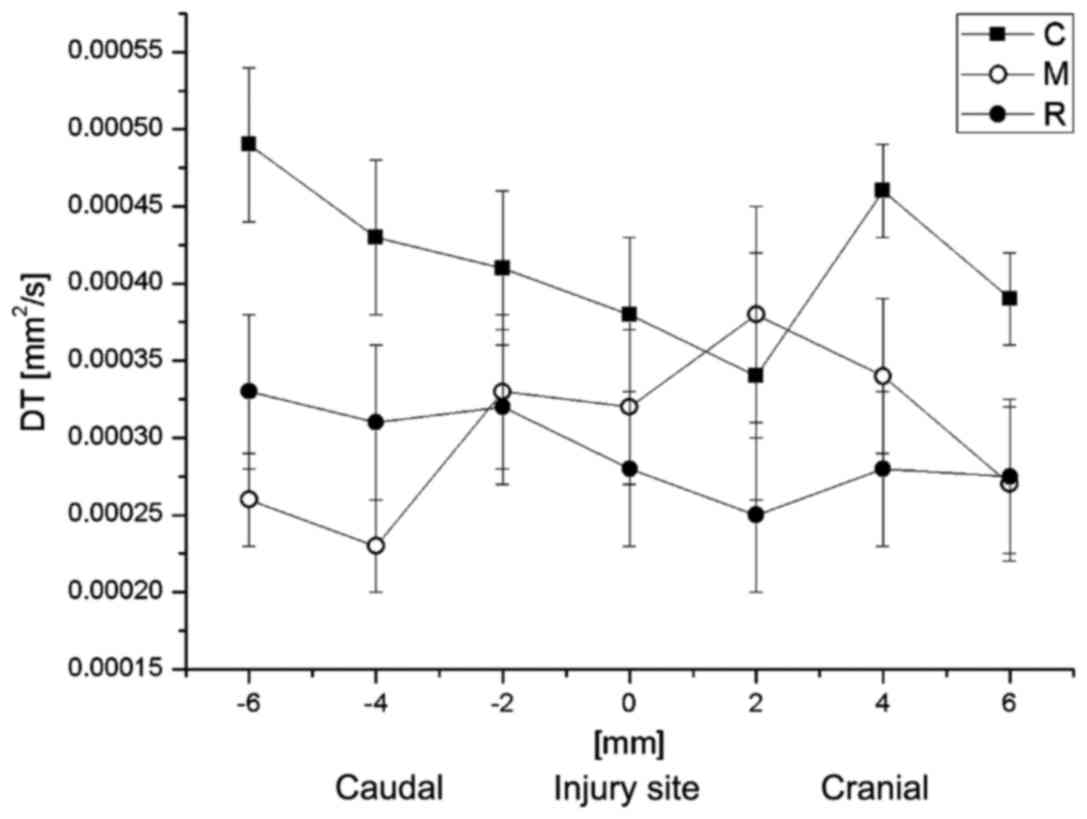
Subsequently, the correlation between DT or DL with
the distance from the injury site was analyzed. The average values
of DL and DT at the particular spinal cord levels are demonstrated
in Figs. 10 and 11, respectively. DL values in group C were
significantly lower than in the group R 2-mm caudally to the injury
site, at the injury site, and at 2 and 4-mm cranially to the injury
site (Fisher's least significant difference test; P=0033, 0.039,
0.023 and 0.027, respectively), while no significant difference was
observed between them at the caudal side from the injury. In
general, the DL values in group M were closer to the group R values
(Fig. 10). At the caudal side,
results for group C were also nearer those of group R. DT values
for all groups did not demonstrate any significant differences
between each other at any spinal cord level. However, the DL and DT
values for group M were generally closer to the respective values
for group R (Fig. 11).
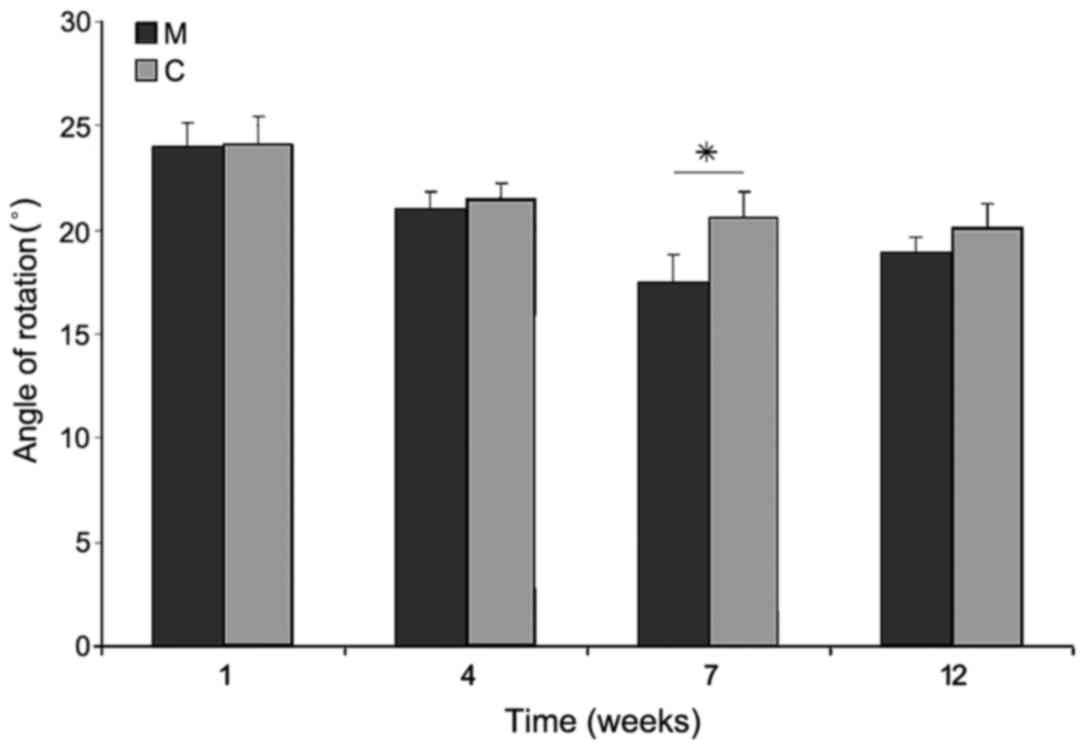
Behavioral study
Immediately after SCI, the majority of rats were
monoplegic; 14 rats in M group and all in C group. The hind paw
angle of rotation increased in all rats in the first week after
surgery, and reached 20.3±1.1° in group C rats at the end of
experiment, indicating marked deterioration of locomotor function
following injury. In group M, at week 12 after surgery, this
parameter had decreased; however, this decrease was not significant
compared with group C (18.9±0.8°). The differences appeared in week
7 of observation, where the angle of rotation in group M was
significantly lower than that in group C (P<0.05; Fig. 12). Interlimb coordination improved
significantly in animals treated with microglia compared with those
in group C at weeks 4, 7 and 12 of observation (P<0.05),
reaching a value of 1.61±0.36 cm at week 12, compared with
2.17±0.41 cm in group C (Fig.
13).
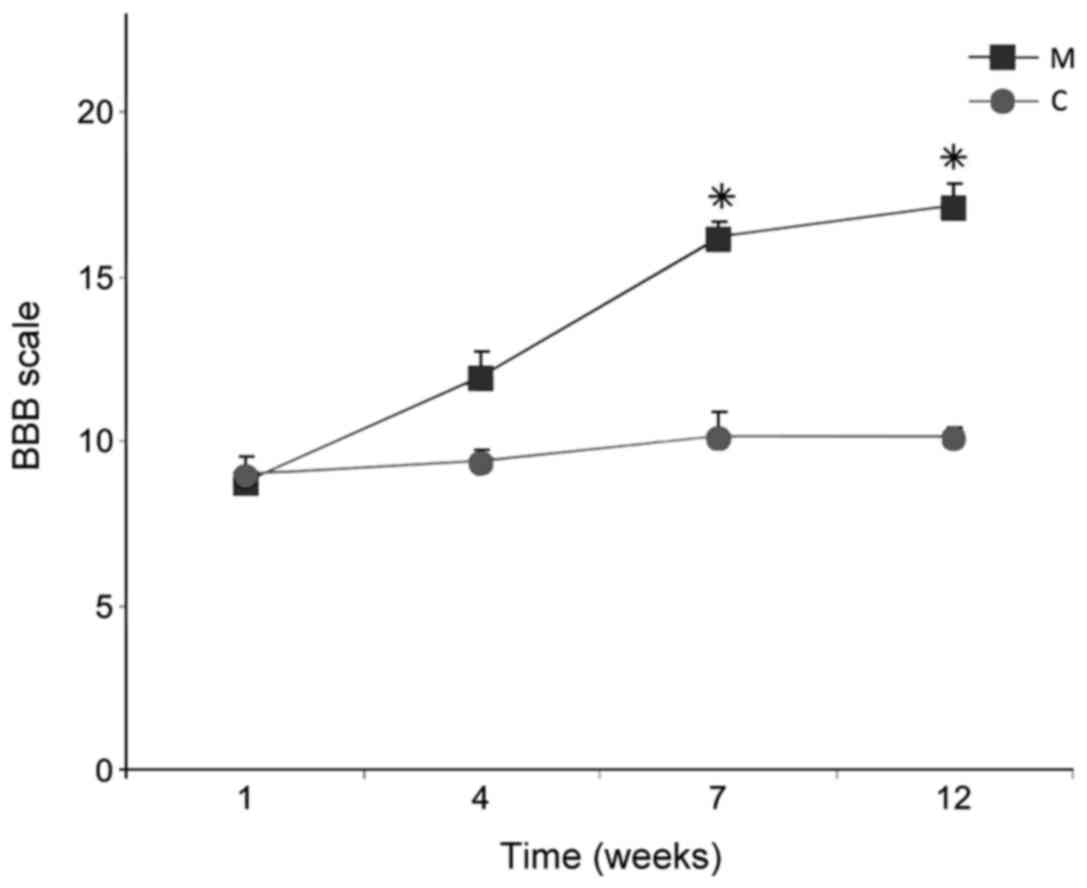
BBB tests revealed improved dynamics of recovery of
function in group M compared with group C. The index value, which
was <10 1 week after SCI in all rats, did not improve after 12
weeks in rats in group C (10.1±0.3), while in group M the index
increased significantly by weeks 7 and 12, reaching 17.5±0.3 at
week 12 (P<0.05; Fig. 14).
Discussion
On the basis of the present results, a potentially
beneficial role of activated microglia in the treatment of SCI
cannot be excluded. Both morphological features of injury and
functional status were improved in rats that received implantations
of microglial cells in the vicinity of injury. The predominant idea
of delivery of microglia into the injured spinal cord was to
scavenge the remnants of damaged neurons, which should improve
regeneration by suppressing the inflammatory reaction (2). In the classical experiment, Schwartz
et al (1) used cultured and
activated autologous blood-derived macrophages to stimulate
regeneration of injured spinal cord. Such a procedure has also been
attempted in human therapy (20,24). The
present study focused on another population of phagocytic cells,
microglia, which naturally occur in the brain and spinal cord.
The present study utilized glial cells from immature
rats. From the studies of newborn infant rats, it has been
demonstrated that there is no development of post-traumatic
scarring at all, and cell response of microglia greatly differs
from adults. It was observed that, in newborn rats, infiltration of
microglial cells at the site of SCI was reduced and they
disappeared quickly (after just a few days), covering only the
nearest region of post-traumatic changes (25). Only single reports exist that
demonstrate that intraspinal transplanted microglial cells were
capable of stimulating regeneration of sensory neurons (26). Furthermore, in cell culture
conditions, convincing evidence was obtained that a mixed
population of microglia and Schwann cells stimulated the
regeneration of sensory cells more effectively than either of these
populations used separately (27).
Thus, these studies indicated that microglia are able to improve
the regeneration of selected populations of neurons and interact
with other cell types in the spinal cord. A study by Yu et
al for the first time demonstrated that delayed transplantation
(7 days after SCI) of activated microglial cell suspension directly
into the injured spinal cord area improved the hind limb motor
function recovery and reduced the size of the liquefaction necrosis
area. The extent of lower limb motor function improvement has a
positive correlation with the number of aggregated microglia.
Furthermore, this demonstrated that the functions of microglial
cells may be modified pharmacologically, by neutralizing IL-6
action by blocking the availability of its specific receptor
(28). As a result, in the injured
spinal cord, level and activity of granulocyte macrophage
colony-stimulating factor, a potent mitogen factor for microglial
cells, increased and in turn led to a rapid decrease in the amount
of degradation products of myelin and Nogo-A protein, a well-known
inhibitor of axonal outgrowth (29).
Consequently, this resulted in increased re-growth and sprouting of
axons, and the final result was the improvement of motor function
(29).
There are also works that have indicated that
suppression of the inflammatory reactions of microglia in the
injured spinal cord through the application of hypothermia may have
positive influences on the structure of the spinal cord, as well as
on the recovery of lost motor function (30). It has been also been demonstrated
that T- and B-lymphocytes restrict functional recovery from SCI by
the increasing numbers of microglia/macrophages, as well as
decreasing axonal sprouting and myelination (31). A study by Emmetsberger and Tsirka
(32) suggested that suppressing
microglia/macrophage activation with specific tripeptide
macrophage/microglia inhibitory factor reduced secondary injury
around the lesion epicenter in a dorsal hemisection model of SCI.
It decreased the hypertrophic change of astrocytes and induced an
increase in the number of axons present within the lesion epicenter
(32). Furthermore, timely
inhibition of microglial/macrophage activation prevented
demyelination and axonal dieback by modulating oligodendrocyte
survival and oligodendrocyte precursor maturation (32).
To speed up the treatment of SCI, the present study
applied microglia already pre-activated in culture. However, one
must consider the side effects of the use of microglia in an active
form, which is the possibility of their ‘hyperstimulation’, leading
to the origin of so-called hyperreactive cells. Such cells secrete
many substances toxic to neurons, such as TNF-α, prostaglandin E2
or nitric oxide, in contrast to non-stimulated microglial cells
that produce rather neurotrophic factors (33). Fortunately, such a situation requires
specific conditions. In the microglial cell culture used in the
present study, a relatively low concentration of LPS was chosen,
empirically based on previous tests (unpublished data), to avoid
hyperstimulation of microglia. The results of the present study
indicated a potential positive influence of activated microglia on
the regeneration of spinal cord white matter.
In conclusion, the present study demonstrated that
delivery of activated microglia to the injured spinal cord may be
an effective method for the repair of white matter. Activated
microglia may be one of the optimal candidates for the repair of
spinal cord after injury. Nevertheless, this issue requires further
investigation, particularly if its future implementation into human
therapy is concerned.
Acknowledgements
The present study was supported by the Medical
University of Silesia (grant nos. KNW-1-030/P/1/0 and
KNW-1-026/P/2/0). The results of the current study were presented
on July 15, 2012, at 8th FENS Forum in Barcelona 14-18.07.2012
(presentation code 12.26, Abstract Number 3472, Poster Board Number
C111).
References
|
1
|
Schwartz M, Cohen I, Lazarov-Spiegler O,
Moalem G and Yoles E: The remedy may lie in ourselves: Prospects
for immune cell therapy in central nervous system protection and
repair. J Mol Med (Berl). 77:713–717. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Schwartz M: Macrophages and microglia in
central nervous system injury: Are they helpful or harmful? J Cereb
Blood Flow Metab. 23:385–394. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Moalem G, Leibowitz-Amit R, Yoles E, Mor
F, Cohen IR and Schwartz M: Autoimmune T cells protect neurons from
secondary degeneration after central nervous system axotomy. Nat
Med. 5:49–55. 1999. View
Article : Google Scholar : PubMed/NCBI
|
|
4
|
Loane DJ and Byrnes KR: Role of microglia
in neurotrauma. Neurotherapeutics. 7:366–377. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Del Río Hortega P: La neuroglía normal.
Conceptos de neurogliona y angiogliona. Arch Histol Norm Patol.
1:5–71. 1942.
|
|
6
|
Walker PA, Shah SK, Jimenez F, Aroom KR,
Harting MT and Cox CS Jr: Bone marrow-derived stromal cell therapy
for traumatic brain injury is neuroprotective via stimulation of
non-neurologic organ systems. Surgery. 152:790–793. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Miyake T, Tsuchihashi Y, Kitamura and
Fujita S: Immunohistochemical studies of blood monocytes
infiltrating into the neonatal rat brain. Acta Neuropathol.
62:291–297. 1984. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Perry VH, Hume DA and Gordon S:
Immunohistochemical localization of macrophages and microglia in
the adult and developing mouse brain. Neuroscience. 15:313–326.
1985. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
David S and Kroner A: Repertoire of
microglial and macrophage responses after spinal cord injury. Nat
Rev Neurosci. 12:388–399. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
10
|
Fujita S and Kitamura T: Origin of brain
macrophages and the nature of the so-called microglia. Acta
Neuropathol Suppl. Suppl 6:S291–S296. 1975.
|
|
11
|
Li Y, Li D and Raisman G: Transplanted
schwann cells, not olfactory ensheathing cells, myelinate optic
nerve fibres. Glia. 55:312–316. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Aloisi F: Immune function of microglia.
Glia. 36:165–179. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Yaguchi M, Ohta S, Toyama Y, Kawakami Y
and Toda M: Functional recovery after spinal cord injury in mice
through activation of microglia and dendritic cells after IL-12
administration. J Neurosci Res. 86:1972–1980. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Marcol W, Slusarczyk W, Gzik M,
Larysz-Brysz M, Bobrowski M, Grynkiewicz-Bylina B, Rosicka P,
Kalita K, Węglarz W, Barski JJ, et al: Air gun impactor-a novel
model of graded white matter spinal cord injury in rodents. J
Reconstr Microsurg. 28:561–568. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Labuzek K, Kowalski J, Gabryel B and
Herman ZS: Chlorpromazine and loxapine reduce interleukin-1beta and
interleukin-2 release by rat mixed glial and microglial cell
cultures. Eur Neuropsychopharmacol. 15:23–30. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Mosmann T: Rapid colorimetric assay for
cellular growth and survival: Application to proliferation and
cytotoxicity assays. J Immunol Methods. 65:55–63. 1983. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Satoh J, Lee YB and Kim SU: T-cell
costimulatory molecules B7-1 (CD80) and B7-2 (CD86) are expressed
in human microglia but not in astrocytes in culture. Brain Res.
704:92–96. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Giri S, Nath N, Smith B, Viollet B, Singh
AK and Singh I:
5-aminoimidazole-4-carboxamide-1-beta-4-ribofuranoside inhibits
proinflammatory response in glial cells: A possible role of
AMP-activated protein kinase. J Neurosci. 24:479–487. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Basso DM, Beattie MS and Bresnahan JC: A
sensitive and reliable locomotor rating scale for open field
testing in rats. J Neurotrauma. 12:1–21. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Karimi-Abdolrezaee S, Eftekharpour E, Wang
J, Morshead CM and Fehlings MG: Delayed transplantation of adult
neural precursor cells promotes remyelination and functional
neurological recovery after spinal cord injury. J Neurosci.
26:3377–3389. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Węglarz WP, Jasiński A, Krzyżak AT,
Kozłowski P, Adamek D, Sagnowski P and Pindel J: MR microscopy of
water diffusion tensor in biological systems. Appl Magn Reson.
15:333–341. 1998. View Article : Google Scholar
|
|
22
|
Weglarz WP, Adamek D, Markiewicz J, Skórka
T, Kulinowski P and Jasiński A: Analysis of the diffusion weighted
MR microscopy data of excised spinal cord of a rat on the basis of
model of restricted diffusion. Solid State Nucl Magn Reson.
25:88–93. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Minati L and Węglarz WP: Physical,
foundations, models, and methods of diffusion magnetic resonance
imaging of the brain: A review. Concepts Magnetic Resonance.
30A:278–307. 2007. View Article : Google Scholar
|
|
24
|
Knoller N, Auerbach G, Fulga V, Zelig G,
Attias J, Bakimer R, Marder JB, Yoles E, Belkin M, Schwartz M and
Hadani M: Clinical experience using incubated autologous
macrophages as a treatment for complete spinal cord injury: Phase I
study results. J Neurosurg Spine. 3:173–181. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Fujimoto Y, Yamasaki T, Tanaka N,
Mochizuki Y, Kajihara H, Ikuta Y and Ochi M: Differential
activation of astrocytes and microglia after spinal cord injury in
the fetal rat. Eur Spine J. 15:223–233. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Prewitt CM, Niesman IR, Kane CJ and Houlé
JD: Activated macrophage/microglial cells can promote the
regeneration of sensory axons into the injured spinal cord. Exp
Neurol. 148:433–443. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Hynds DL, Rangappa N, Ter Beest J, Snow DM
and Rabchevsky AG: Microglia enhance dorsal root ganglion outgrowth
in Schwann cell cultures. Glia. 46:218–223. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Yu TB, Cheng YS, Zhao P, Kou DW, Sun K,
Chen BH and Wang AM: Immune therapy with cultured microglia
grafting into the injured spinal cord promoting the recovery of
rat's hind limb motor function. Chin J Traumatol. 12:291–295.
2009.PubMed/NCBI
|
|
29
|
Mukaino M, Nakamura M, Yamada O, Okada S,
Morikawa S, Renault-Mihara F, Iwanami A, Ikegami T, Ohsugi Y, Tsuji
O, et al: Anti-IL-6-receptor antibody promotes repair of spinal
cord injury by inducing microglia-dominant inflammation. Exp
Neurol. 224:403–414. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Morino T, Ogata T, Takeba J and Yamamoto
H: Microglia inhibition is a target of mild hypothermic treatment
after the spinal cord injury. Spinal Cord. 46:425–431. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wu B, Matic D, Djogo N, Szpotowicz E,
Schachner M and Jakovcevski I: Improved regeneration after spinal
cord injury in mice lacking functional T- and B-lymphocytes. Exp
Neurol. 237:274–285. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Emmetsberger J and Tsirka SE: Microglial
inhibitory factor (MIF/TKP) mitigates secondary damage following
spinal cord injury. Neurobiol Dis. 47:295–309. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Klusman I and Schwab ME: Effects of
pro-inflammatory cytokines in experimental spinal cord injury.
Brain Res. 762:173–184. 1997. View Article : Google Scholar : PubMed/NCBI
|