Introduction
In 1999, researchers observed that Toll-like
receptor 4 (TLR4) was highly expressed in myocardial cells. In
normal mice and human myocardium, expression of TLR4 in
cardiomyocyte is mainly diffused in cytoplasm. However, expression
of TLR4 was upregulated in the myocardium of rats after infarction
which is far away from the lesion of ischemic injury and in
myocardium of patients with dilated cardiomyopathy. TLR4 staining
showed very strong signal at the junction of two or more than two
cells (1). It has been shown that
lipopolysacchride (LPS) and AngII can synergistically increase
intracellular Ca2+ levels at rest, reduce the transient
increase of Ca2+ level and damage the function of
mitochondria (2). Later on, more
studies have shown that TLR4/nuclear factor-κB (NF-κB), a signal
transduction pathway closely related to immune, inflammatory and
oxidative stress, is closely related to the local renin-angiotensin
system (RAS). TLR4/NF-κB is involved in inflammatory response of
myocardial injury and physiological processes of myocardial
remodeling, and intervention of TLR4/NF-κB signal can attenuate
myocarditis and myocardial damage (3–5).
Ginkgo biloba extract (GBE) is a traditional Chinese
medicine that plays an important role in anti-myocardial injury,
and function of GBE is related to its role in scavenging oxygen
free radical and suppressing immune response (6–8).
However, the involvement of TLR4/NF-κB signaling pathway in
myocardial remodeling prevention through GBE is still unknown. In
this study, TLR4/NF-κB signaling pathway in rat ventricular
myocytes was activated in vitro using LPS, which is a TLR4
specific agonist (9,10). The effects of Ginkgo biloba
leaf extract (GBE50) on TLR4/NF-κB signal and local RAS were
observed, and the mechanism of GBE50 in the prevention and
treatment of myocardial remodeling was also explored.
Materials and methods
Experimental animals and major
reagents
Healthy new born Sprague-Dawley rats (6–24 hour old)
without restriction on sex were provided by Experimental Animal
Center of Dalian Medical University. GBE: Gingko biloba
(GBE50) containing 44.1% of Ginkgo biloba total flavonoids
and 6.4% of ginkgolides was provided by SPH Xingling Science and
Technology Pharmaceutical Co., Ltd. (Dalian, China). LPS (055:B5),
caffeic acid phenethyl ester (CAPE), a specific inhibitor of NF-κB,
type II collagenase, trypsin, and 5-bromo deoxyuridine (Brdu) were
from Sigma (St. Louis, MO, USA). TRIzol and Dulbeccos modified
Eagles medium (DMEM) medium were from Gibco (New York, NY, USA).
Reverse transcription kit was bought from Takara (Dalian, China).
Rabbit anti-rat striated muscle specific sarcomeric α-actin
polyclonal antibody (dilution, 1:200; cat. no. PA5-21396) and SP
immunohistochemistry kit which included biotinylated secondary
antibody, hydrogen peroxide, serum blocker and horseradish
peroxidase labeled streptavidin were from Zymed (San Diego, CA,
USA). Rabbit anti-rat NF-κB p65 polyclonal antibody (dilution,
1:100; cat. no. SC8008) was from Santa Cruz Biotechnology, Inc.
(Dallas, TX, USA). Coomassie Brilliant Blue was from Beijing
Zhongshan Golden Bridge Biotechnology Co., Ltd. (Beijing,
China).
Primary culture of cardiomyocyte and
grouping
Ventricles were collected from Sprague-Dawley rats
aged 6–24 h under sterile conditions and were cut into pieces. 0.1%
trypsin and 0.1% type II collagenase were added to digest the cells
to make single cell suspension. Cardiac fibroblasts and epithelial
cells were removed using differential attachment technique.
Striated muscle specific sagcomeric α-actin monoclonal antibody was
used to identify cardiomyocytes by SABC immunohistochemical
staining. In the first 48 h of culture, 0.1 mmol/l Brdu was added
to inhibit the proliferation of non-cardiomyocytes. DMEM culture
medium without serum was used after culture for 96 h when cells
reached sub-cell-fusion state. After culture for another 24 h,
cells were divided into the following groups: i) control group:
DMEM medium without any intervention factors; ii) LPS group: DMEM
medium containing 1 µg/ml LPS; iii) LPS + GBE50 group: GBE50 was
added first to the final concentration of 80 µg/ml, then LPS was
added to the final concentration of 1 µg/ml 30 min later; iv) LPS +
CAPE group: CAPE was added first to the final concentration of 20
µg/ml, 30 min later, LPS was added to the final concentration of 1
µg/ml. After intervention for 24 h, cells were collected.
Immunocytochemical analysis of the
activation of NF-κB
Density of neonatal rat ventricular myocytes (NRVMs)
was adjusted to 1×106/ml and inoculated on cell culture
plate with coverglass in each well. After treatment for 24 h,
coverglasses were collected and washed with PBS. Fixation was
performed with 1% paraformaldehyde (pH 7.0) at room temperature for
30 min, followed by washing with 0.1% Triton X-100 (pH 7.0) at room
temperature for 5 min. Rabbit anti-mouse NF-κB p65 antibody (1:100)
was added, followed by incubation at 37°C for 60 min. Biotin-avidin
reaction system was used. After color development with DAB and
hematoxylin re-staining, slides were sealed. Determination of
immunocytochemical results were as follows: because NF-κB is in
cytoplasm under normal condition, after activation, it will enter
into nucleus, nuclear-positivity was used as the criteria for the
determination of the activation of NF-κB. Under light microscope
(200-fold; Olympus, Tokyo, Japan), 5 visual fields were randomly
selected. The activation of NF-κB was assessed by calculating the
percentage of positive cells.
Detection of the expression of TLR4,
AT1a receptor, angiotensinogen (ATG) and β-myosin heavy
chain (β-MHC) mRNA in cardiomyocytes
Primers were synthesized by
Takara
Total RNA was extracted using TRIzol reagent
according to the instruction. RNA sample with
OD260/OD280=1.6–1.8 were used for reverse
transcription. Reverse transcription conditions: 30°C for 10 min,
50°C for 30 min and 99°C 5 min. RT-PCR was performed to detect the
expression of TLR4, AT1a receptor, ATG and β-MHC mRNA.
TLR4 primer sense, 5′-CGCTTTCAGCTTTGCCTTCATTAC-3′ and antisense,
5′-AGCTACTTCCTTGTGCCCTGTGAG-3′, length of amplified fragment was
555 bp; ATG primer sense, 5′-TTCAGGCCAAGACCTCCC-3′ and antisense,
5′-CCAGCCGGGAGGTGCAGT-3′, length of amplified fragment was 308 bp;
AT1a primer sense, 5′-GCACACTGGCAATGTAATGC-3′ and
antisense, 5′-GTTGAACAGAACAAGTGACC-3′, length of amplified fragment
was 385 bp; β-MHC primer sense, 5′-GTGGACGTTTATTGACTTCGG-3′ and
antisense, 5′-TTCTTTGCTTTGCCTTTGC-3′, length of amplified fragment
was 399 bp; endogenous control β-actin primer sense,
5′-AACCCTAAGGCCAACCGTGAAAAG-3′ and antisense,
5-TCATGAGGTAGTCTGTCAT-3′, length of amplified fragment was 241 bp.
PCR reaction conditions for TLR4, 94°C for 2 min, followed by 30
cycles of 94°C for 30 sec, 60°C for 60 sec and 72°C for 1.5 min,
and 72°C for 10 min; PCR reaction conditions for ATG, 94°C for 2
min, followed by 30 cycles of 94°C for 30 sec, 63°C for 60 sec and
72°C for 1.5 min, and 72°C for 10 min; PCR reaction conditions for
AT1a receptor, 94°C for 2 min, followed by 30 cycles of
94°C for 30 sec, 58°C for 45 sec and 72°C for 1.5 min, and 72°C for
10 min; PCR reaction conditions for β-MHC, 94°C for 2 min, followed
by 35 cycles of 94°C for 30 sec, 55°C for 45 sec and 72°C for 1
min, and 72°C for 7 min. PCR products were subjected to 1.5%
agarose gel electrophoresis. A gel imaging system was used to take
the images. The ratio of the absorbance of the target gene band to
the absorbance of the β-actin band was taken as the relative
expression level of target mRNA.
Determination of protein content in
cardiomyocytes
Cells were digested with 0.125% trypsin, and 0.1%
SDS (0.1 ml) was added to lysis cells, followed by centrifugation
at 2,000 × g for 5 min to collect supernatant. Total protein
content of cardiomyocytes was measured by Coomassie Brilliant Blue
method.
Statistical analysis
Data were processed using SPSS 11.5 statistical
software (version X; IBM, Armonk, NY, USA). Each sample was
measured three times in parallel, and the data were expressed as
mean ± standard deviation (mean ± SD). Comparisons among multiple
samples were performed by variance analysis. Comparisons between
two samples were performed by Q test. P<0.05 was considered to
be statistically significant.
Results
Culture and identification of
NRVMS
After fibroblasts were removed by differential
attachment technique, cells were cultured for 24 h. Cardiomyocytes
were long rod-, polygonal or spindle-shaped with concentric or
radial growth. After culture for 48 h, synchronized pulsation
(12–80 bpm) was observed in cardiomyocytes. Immunocytochemical
identification of striated muscle specific sarcomeric α-actin
showed that the purity of cardiomyocytes was >95%.
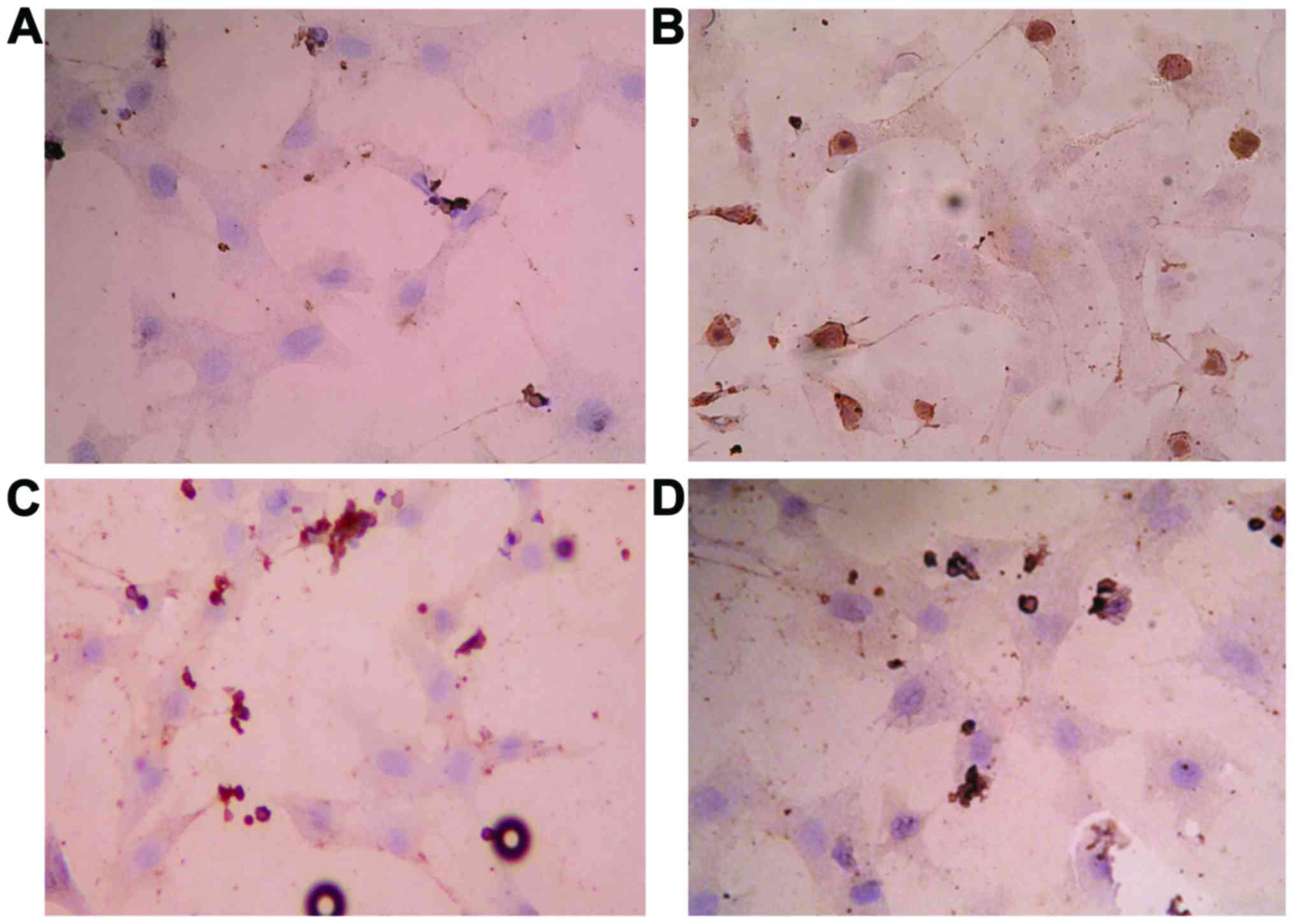
Effects of GBE50 and NF-κB inhibitor
CAPE on activation of NF-κB induced by LPS in NRVMs
Results of immunocytochemical analysis showed that
NF-κB was activated 24 h after LPS (1 µg/ml) stimulation, and
nuclear staining was significantly increased (P<0.01). After
treatment with GBE50 (80 µg/ml) and CAPE (20 µg/ml), almost no
nuclear staining was observed (P<0.01) (Fig. 1).
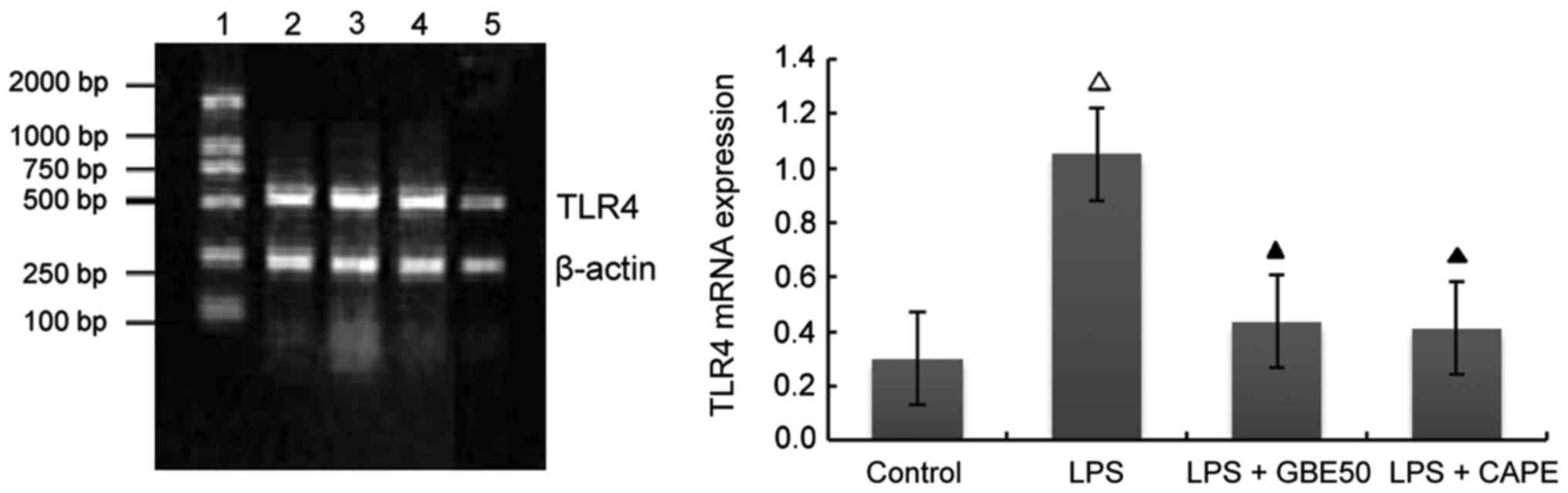
Effects of GBE50 and NF-κB inhibitor
CAPE on TLR4 mRNA expression induced by LPS in NRVMs
Compared with control group (expression level of
TLR4 mRNA was 0.301±0.112), expression level of TLR4 mRNA in NRVMs
were significantly increased at 24 h after LPS (1 µg/ml)
stimulation (expression level of TLR4 mRNA was 1.052±0.227,
P<0.01); compared with LPS group, GBE50 (80 µg/ml) and CAPE (20
µg/ml) significantly reduced the expression of TLR4 mRNA in NRVMs
induced by LPS (expression levels of TLR4 mRNA were 0.436±0.145 and
0.411±0.123, respectively, P<0.01). Compared with LPS + GBE50
group, expression level of TLR4 mRNA was slightly reduced in LPS +
CAPE group, but the difference was not significant (Fig. 2).
Effects of GBE50 and NF-κB inhibitor
CAPE on LPS-induced expression of ATG mRNA in NRVMs
Compared with control group (expression level of ATG
mRNA was 0.652±0.227), expression level of ATG mRNA was
significantly increased in NRVMs at 24 h after LPS (1 µg/ml)
stimulation (expression level of ATG mRNA was 0.652±0.227,
P<0.01). GBE50 (80 µg/ml) and CAPE (20 µg/ml) could inhibit the
increased expression level of ATG mRNA induced by LPS (expression
levels of ATG mRNA were 0.171±0.102 and 0.141±0.097, respectively,
P<0.01). No significant differences in the expression level of
ATG mRNA were found between LPS + GBE50 group, LPS + CAPE group and
control group. Compared with LPS + GBE50 group, expression level of
ATG mRNA was slightly reduced in LPS + CAPE group, but the
difference was not significant (Fig.
3).
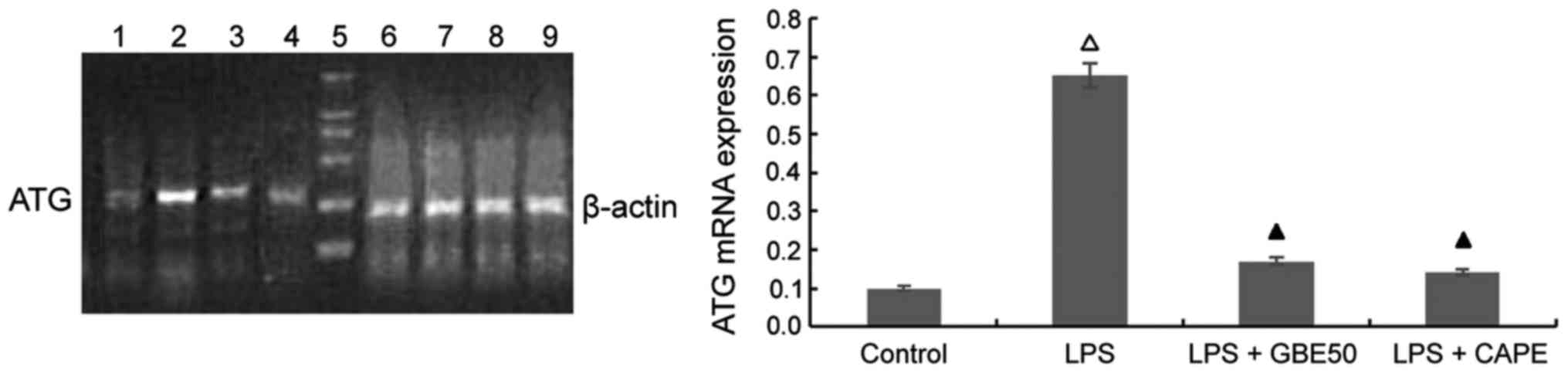 | Figure 3.RT-PCR amplification of ATG.
Lipopolysaccharide (LPS) upregulated expression level of ATG mRNA
in neonatal rat ventricular myocytes (NRVMs), whereas Ginkgo
biloba leaf extract (GBE50) and caffeic acid phenethyl ester
(CAPE) reduced LPS-induced increase in expression level of ATG. 5,
DNA marker, bands from bottom to top are 100, 250, 500, 750, 1,000
and 2,000 bp; 1–4, angiotensinogen (ATG); 6–9, β-actin; 1,6,
control group; 2,7, LPS group; 3,8, LPS + GBE50 group; 4,9, LPS +
CAPE group. △P<0.01 compared with control group;
▲P>0.05 compared with contol group. |
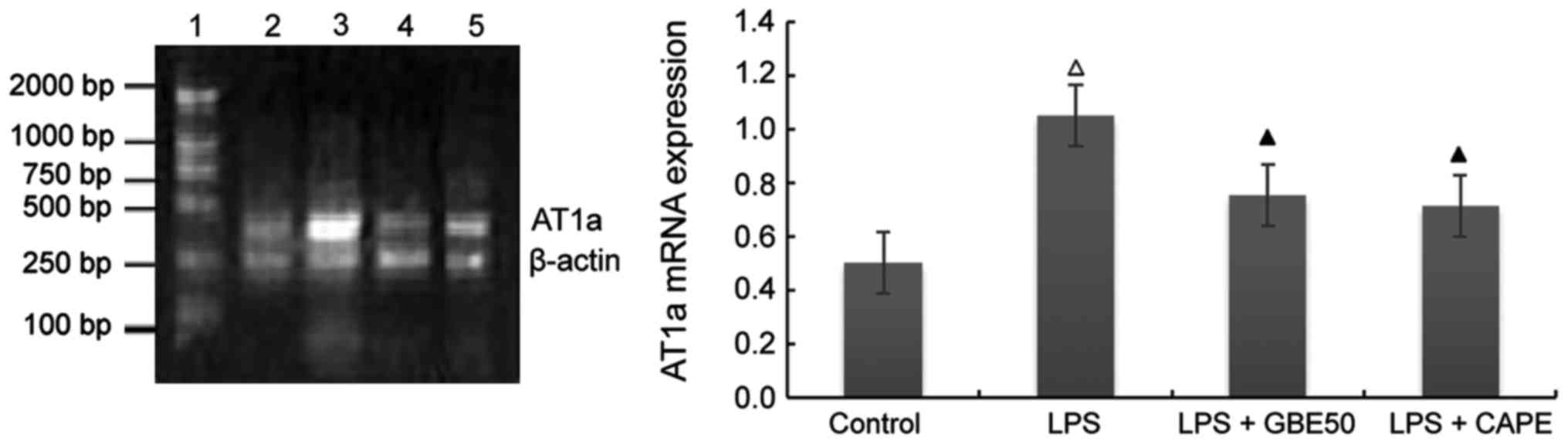
Effect of GBE50 and NF-κB inhibitor
CAPE on expression of AT1a receptor mRNA induced by LPS
in NRVMs
Compared with control group (expression level of
AT1a receptor mRNA was 0.501±0.112), expression level of
AT1a receptor mRNA in NRVMs were significantly increased
at 24 h after LPS (1 µg/ml) stimulation (expression level of TLR4
mRNA was 1.05±0.227, P<0.01); compared with LPS group, GBE50 (80
µg/ml) and CAPE (20 µg/ml) significantly reduced the expression of
AT1a mRNA in NRVMs induced by LPS (expression levels of
AT1a receptor mRNA were 0.751±0.257 and 0.711±0.223,
respectively, P<0.01). No significant difference in the
expression level of AT1a receptor mRNA was found between
LPS + GBE50 group, LPS + CAPE group and control group. Compared
with LPS + GBE50 group, expression level of AT1a
receptor mRNA was slightly reduced in LPS + CAPE group, but the
difference was not significant (Fig.
4).
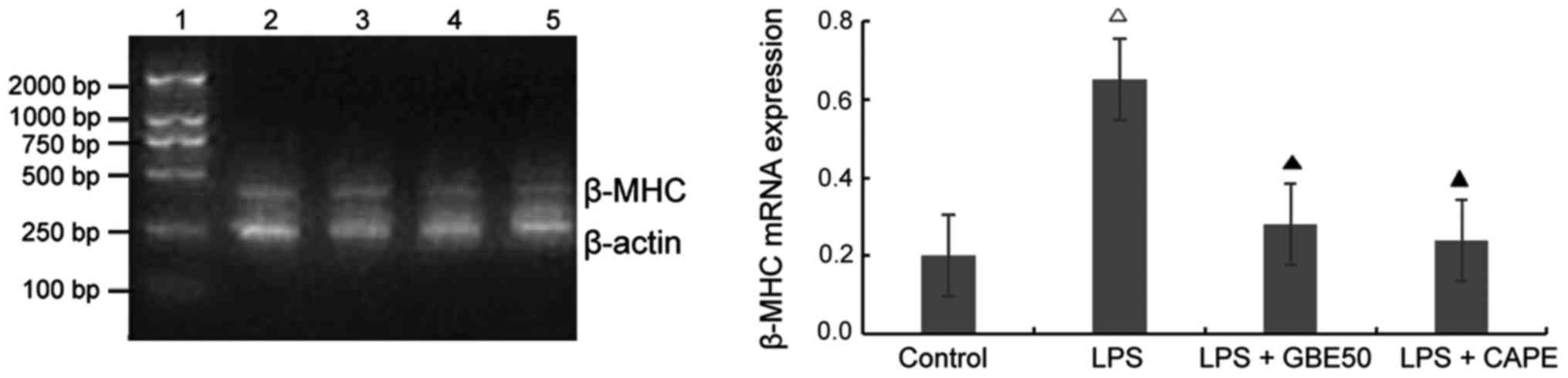
Effect of GBE50 and NF-κB inhibitor
CAPE on expression of β-MHC mRNA induced by LPS in NRVMs
Compared with control group (expression level of
β-MHC mRNA was 0.201±0.098), expression level of β-MHC mRNA in
NRVMs were significantly increased at 24 h after LPS (1 µg/ml)
stimulation (expression level of β-MHC mRNA was 0.653±0.228,
P<0.01); compared with LPS group, GBE50 (80 µg/ml) and CAPE (20
µg/ml) significantly reduced the expression of β-MHC mRNA in NRVMs
induced by LPS (expression levels of β-MHC mRNA were 0.201±0.098
and 0.191±0.089, respectively, P<0.01). No significant
difference in the expression level of β-MHC mRNA was found between
LPS + GBE50 group, LPS + CAPE group and control group. Compared
with LPS + GBE50 group, expression level of β-MHC mRNA was slightly
reduced in LPS + CAPE group, but the difference was not significant
(Fig. 5).
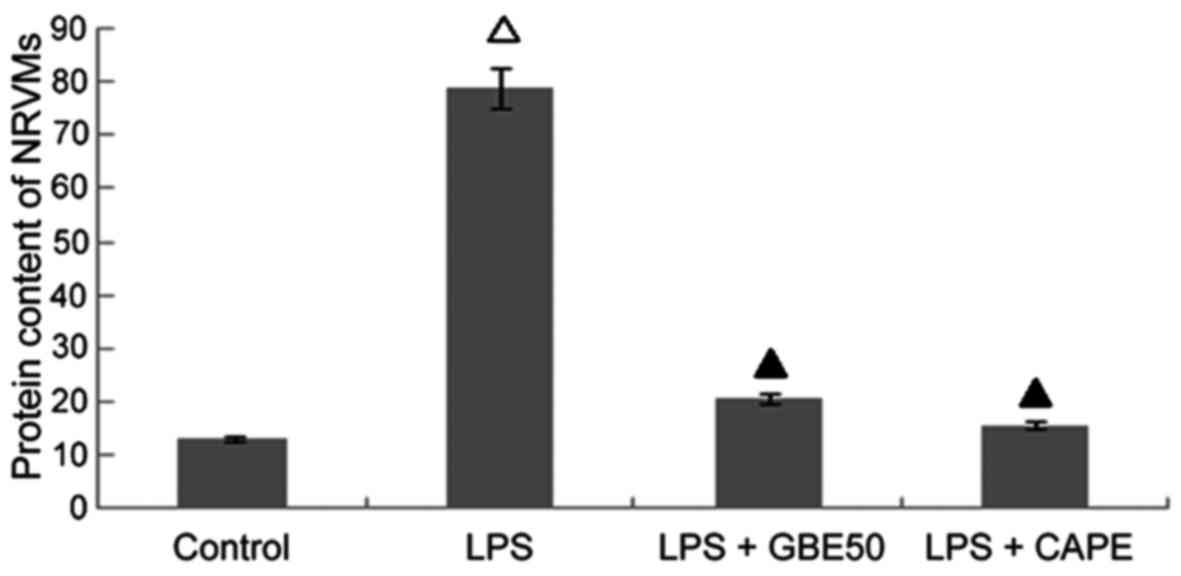
Effects of GBE50 and NF-κB inhibitor
CAPE on increased protein content in NRVMs cells induced by
LPS
Compared with control group (protein content was
13.18±2.51 mg/l), protein content in NRVMs were significantly
increased at 24 h after LPS (1 µg/ml) stimulation (protein content
was 78.81±8.51 mg/l, P<0.01); compared with LPS group, GBE50 (80
µg/ml) and CAPE (20 µg/ml) significantly reduced the increased
protein content in NRVMs induced by LPS (protein contents were
20.80±3.42 and 15.78±3.02 mg/l, respectively, P<0.01). No
significant difference in protein content was found between LPS +
GBE50 group, LPS + CAPE group and control group. Compared with LPS
+ GBE50 group, protein content was slightly reduced in LPS + CAPE
group, but the difference was not significant (Fig. 6).
Discussion
In 1997, Medzhitov et al (11) found the first human Toll
protein-TLR4, confirming the presence of TLR4/NF-κB signaling
pathway in human body. TLR4/NF-κB is the primary pathway that
mediates LPS signaling, and LPS from Escherichia coli can
specifically activate TLR4 without affecting other members of
Toll-like receptor families (9,10). Our
results showed that TLR4 was expressed in NRVMs, and LPS increased
the expression level of TLR4, ATG and AT1a receptor, and
also increased the indexes of cardiomyocyte hypertrophy - protein
content in cardiomyocytes and expression level of β-MHC expression,
suggesting that TLR4/NF-κB signaling pathway may be involved in the
occurrence and development of cardiomyocyte hypertrophy by
activating local renin-angiotensin system. NF-κB is a group of
multidirectional nuclear transcriptional regulators. Under normal
conditions, p50 and p65, which are two subunits of NF-κB, can form
heterodimers in cytoplasm, in the inactive form of NF-κB.
Inhibitors-κB (IκBs), which can bind to NF-κB dimer to mask the
nuclear localization sequence of NF-κB, controls the activation of
NF-κB. Phosphorylation and activation of inhibitor-κB kinase (IKK)
can be induced by a variety of stimulatory factors, and
phosphorylated IκBs will replease and activate NF-κB, then NF-κB
will translocate from cytoplasm to nucleus to bind the target
genes, which in turn regulate the expression of a series of target
genes related to the pathophysiological processes of cardiovascular
disease and promote protein synthesis in cardiomyocytes. The target
genes include cytokines, angiotensinogen, AT1 receptor and
embryonic genes such as β-MHC (12).
In vivo and in vitro experiments have shown that
NF-κB is the essential nuclear transcriptional regulator for the
development of cardiac hypertrophy (13,14). In
this study, 24 h after LPS stimulation, NF-κB p65 was activated,
while NF-κB inhibitor CAPE inhibited the activation of NF-κB p65
and reduced the expression levels of angiotensinogen,
AT1a receptor, β-MHC and increased protein content in
cardiomyocytes. Therefore, TLR4/NF-κB is involved in the
LPS-induced changes in cardiomyocytes, and NF-κB may play a key
role.
GBE50 is the concentrated and purified active
ingredient of Ginkgo biloba. GBE50 contains 44.1% of
Ginkgo biloba total flavonoids, 26.4% of flavonoid
glycosides and 6.4% of ginkgolides. Ginkgolides can effectively
inhibit the activation of platelet and flavonoids can remove oxygen
free radicals to achieve antioxidant effect. It is reported that
GBE can remove oxygen free radicals and reduce the expression
levels of p53 and Bcl-2, so GBE is widely used in the treatment of
cardiovascular disease (6–8). Myocardial ischemia and reperfusion
experiments found that GBE can increase antioxidant activity in
plasma, which in turn protects the heart (15). In animal models of
doxorubicin-induced chronic cardiotoxicity, Naidu et al
(16) showed that GBE could reduce
the adipokine-induced increase in myocardial lipid peroxidation and
restored the antioxidant activity of superoxide dismutase and
reduced glutathione, which in turn protects the heart from
adriamycin toxicity and reduces mortality. GBE50 inhibited the
activation of TLR4/NF-κB and local RAS in NRVMs induced by LPS, and
also reduced the expression of β-MHC and decreased the protein
content and reversed cardiac hypertrophy. TLR4/NF-κB and RAS are
also closely related to myocardial remodeling, suggesting that
GBE50 may have an important role in the prevention and treatment of
myocardial remodeling. Results of this study suggest that GBE50 and
NF-κB inhibitor CAPE have similar effects, and they both can
significantly inhibit the activation of NF-κB. Studies have shown
that reactive oxygen species are involved in the activation of
NF-κB induced by LPS, and various antioxidants can inhibit NF-κB
activation (1,12). So the inhibitory effects of GBE50 on
NF-κB activation may be related to the function of GBE50 in
scavenging oxygen free radicals.
In this study, LPS upregulated expression level of
TLR4 in NRVMs, while GBE50 reduced the increased expression level
of TLR4. Compared with LPS + GBE50 group, expression level of TLR4
mRNA was slightly reduced in LPS + CAPE group, but the difference
was not significant. In vitro studies performed by Frantz
et al (1) suggested that
NF-κB activation can not only mediate downstream signal
transduction of TLR4, but also is necessary for maintaining TLR4
expression. Results of this study showed that expression level of
TLR4 was increased after the activation of NF-κB, while expression
level of TLR4 was decreased with the inhibited activation of NF-κB,
suggesting that the regulatory role of GBE50 in TLR4 expression may
also be associated with the inhibition of NF-κB activation. Through
decreasing the expression level of TLR4, GBE50 indirectly inhibited
the activation of NF-κB, which in turn reduced TLR4/NF-κB signal,
so as to inhibit the activation of RAS, reduce expression level of
β-MHC and increase protein content in cardiomyocytes with activated
TLR4/NF-κB pathway.
In conclusion, GBE50 can inhibit the activation of
NF-κB and expression of TLR4 induced by LPS, which in turn
attenuate TLR4/NF-κB signaling pathway and reduce the increased
expression levels of angiotensinogen and AT1a receptors.
TLR4/NF-κB signaling pathway intervention may be one of the
mechanisms of GBE in preventing myocardial remodeling.
Acknowledgements
We are grateful for the support of the National
Natural Science Foundation of China (grant no. 30371568).
References
|
1
|
Frantz S, Kobzik L, Kim YD, Fukazawa R,
Medzhitov R, Lee RT and Kelly RA: Toll4 (TLR4) expression in
cardiac myocytes in normal and failing myocardium. J Clin Invest.
104:271–280. 1999. View
Article : Google Scholar : PubMed/NCBI
|
|
2
|
Yasuda S and Lew WY: Angiotensin II
exacerbates lipopolysaccharide-induced contractile depression in
rabbit cardiac myocytes. Am J Physiol. 276:H1442–H1449.
1999.PubMed/NCBI
|
|
3
|
Li HL, Suzuki J, Bayna E, Zhang FM, Dalle
Molle E, Clark A, Engler RL and Lew WY: Lipopolysaccharide induces
apoptosis in adult rat ventricular myocytes via cardiac AT(1)
receptors. Am J Physiol Heart Circ Physiol. 283:H461–H467. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Li HB, Li X, Huo CJ, Su Q, Guo J, Yuan ZY,
Zhu GQ, Shi XL, Liu JJ and Kang YM: TLR4/MyD88/NF-κB signaling and
PPAR-γ within the paraventricular nucleus are involved in the
effects of telmisartan in hypertension. Toxicol Appl Pharmacol.
305:93–102. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Dange RB, Agarwal D, Masson GS, Vila J,
Wilson B, Nair A and Francis J: Central blockade of TLR4 improves
cardiac function and attenuates myocardial inflammation in
angiotensin II-induced hypertension. Cardiovasc Res. 103:17–27.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Liu AH, Bao YM, Wang XY and Zhang ZX:
Cardio-protection by Ginkgo biloba extract 50 in rats with
acute myocardial infarction is related to
Na+-Ca2+ exchanger. Am J Chin Med.
41:789–800. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Lu S, Guo X and Zhao P: Effect of
Ginkgo biloba extract 50 on immunity and antioxidant enzyme
activities in ischemia reperfusion rats. Molecules. 16:9194–9206.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Liu TJ, Yeh YC, Ting CT, Lee WL, Wang LC,
Lee HW, Wang KY and Lai HC and Lai HC: Ginkgo biloba extract
761 reduces doxorubicin-induced apoptotic damage in rat hearts and
neonatal cardiomyocytes. Cardiovasc Res. 80:227–235. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Takada H, Yokoyama S and Yang S:
Enhancement of endotoxin activity by muramyldipeptide. J Endotoxin
Res. 8:337–342. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wang J, Zhang Y, Guo LL, Wu GJ and Liu RH:
Salvianolic acid B inhibits the TLR4-NFκB-TNFα pathway and
attenuates neonatal rat cardiomyocyte injury induced by
lipopolysaccharide. Chin J Integr Med. 17:775–779. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Medzhitov R, Preston-Hurlburt P and
Janeway CA Jr: A human homologue of the Drosophila Toll
protein signals activation of adaptive immunity. Nature.
388:394–397. 1997. View
Article : Google Scholar : PubMed/NCBI
|
|
12
|
Jones WK, Brown M, Ren X, He S and
McGuinness M: NF-kappaB as an integrator of diverse signaling
pathways: The heart of myocardial signaling? Cardiovasc Toxicol.
3:229–254. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Purcell NH, Tang G, Yu C, Mercurio F,
DiDonato JA and Lin A: Activation of NF-kappa B is required for
hypertrophic growth of primary rat neonatal ventricular
cardiomyocytes. Proc Natl Acad Sci USA. 98:pp. 6668–6673. 2001,
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Shi Y, Pan H, Zhang HZ, Zhao XY, Jin J and
Wang HY: Lipoxin A4 mitigates experimental autoimmune myocarditis
by regulating inflammatory response, NF-κB and PI3K/Akt signaling
pathway in mice. Eur Rev Med Pharmacol Sci. 21:1850–1859.
2017.PubMed/NCBI
|
|
15
|
Carini M, Aldini G, Rossoni G, Morazzoni P
and Facino RM: Complexation of Ginkgo biloba extract with
phosphatidylcholine improves cardioprotective activity and
increases the plasma antioxidant capacity in the rat. Planta Med.
67:326–330. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Naidu MU, Kumar KV, Mohan IK, Sundaram C
and Singh S: Protective effect of Gingko biloba extract
against doxorubicin-induced cardiotoxicity in mice. Indian J Exp
Biol. 40:894–900. 2002.PubMed/NCBI
|