Introduction
Acute respiratory distress syndrome (ARDS) is
reported to be a kind of disease that is characterized by diffuse
inflammatory injury of lung and capillary destruction of alveolar,
which resulted in alveolar extravasation and acute hypoxemic
respiratory failure (1,2). Assorted and numerous factors were
reported to be predisposing factors for ARDS, such as drug overdose
or abuse, pneumonia, lung contusion, multiple blood transfusions,
aspiration of stomach contents and sepsis (3). It has been demonstrated that advanced
strategies and pharmacological interventions have been used to
patients with ARDS, such as noninvasive positive-pressure
ventilation, neuromuscular blockade, β-adrenergic agonists etc.
(4,5). Obviously, in the past 20 years, our
awareness of the risk factors and mechanisms of ARDS has been
improved, but sepsis-associated ARDS is still related to high
mortality (6,7). Several factors, including transforming
growth factor-β1 (TGF-β) and vascular endothelial (VE) cadherin,
were considered to be critical in the pathogenesis of
sepsis-associated ARDS (8,9). Hence, it is important to explore
molecular mechanism for more effective treatment for patients with
sepsis-associated ARDS.
Toll-like receptors (TLRs) are dominating congenital
immune receptors to recognize pathogen-associated molecular
patterns (PAMPs) that not only initiate the primary response to the
invading pathogens but also induce the adaptive immune response
(10). Toll-like receptor 4 (TLR4)
is a pattern recognition receptor (PPR) that induces inflammatory
responses, especially crucial to the progression of
antigen-specific adaptive immune response, following the
identification of some endogenous ligands related to tissue damage
(11). Myeloid differentiation
primary-response protein 88 (MyD88) act as a shared adaptor
molecule for most TLRs which can elicit the production of multiple
inflammatory cytokine genes and the activation of nuclear factor-κB
(NF-κB) and mitogen-activated protein (MAP) kinases involved in
neurotoxicity (12,13). Castoldi et al provided
evidence that the TLR4/MyD88 signaling pathway was crucial to acute
kidney injury (AKI) induced by sepsis (14). The study of Huang et al
exerted efforts to identify the role of monoclonal antibody against
TLR4 in ventilator-induced lung injury in rats through MyD88/NF-κB
signaling (15), which provides some
clues for further studies in ARDS. Additionally, TLR4 is one of the
key receptors associated with entire body and low-level chronic
inflammatory diseases, leading to the macrophage infiltration in
diabetic liver injury, through activating NF-κB and regulating
pro-inflammatory genes (16).
Macrophages are heterogeneous cells of innate immune system,
playing a critical role in the initiation and resolution of
inflammatory response (17).
Interestingly, multiple TLRs, especially, TLR4, are involved in
macrophage activation and secretion of tumor necrosis factor-α
(TNF-α) (18). From all that
mentioned above, it is speculated that TLR4/MyD88 signaling pathway
may be related to the activation of macrophages and inflammatory
factors, and provide a new therapy for sepsis-associated ARDS. On
this regard, this study aims to investigate the role of TLR4/MyD88
signaling pathway in sepsis-associated ARDS via regulating
macrophage activation and inflammatory response.
Materials and methods
Ethics statement
All animal experiments were approved by Ethics
Committee of The First People's Hospital of Changzhou, and
conducted in accordance with Declaration of Helsinki.
Study subjects
A total of 36 specific pathogen free (SPF) male
Sprague-Dawlay (SD) rats (weighted 230±20 g) were purchased from
Hunan Slack King Laboratory Animal Co., Ltd. (Changsha, Hunan,
China). All rats were housed in separate cages at room temperature
of 18–28°C, with relative humidity of 40–70%, and a 12-h day/night
cycle. Rats were fed a standard diet containing 21% protein and
with free access to water. The experiment was carried out after 2
weeks of feeding.
A rat model of sepsis-associated
ARDS
Rats were intraperitoneally injected with 40 mg/kg
phenobarbital sodium, and they were fixed on the operating table in
the supine position after anesthesia. The abdomen was opened to
find the cecum under aseptic condition and the cecum was ligated
tightly with a 4 silk thread at the location of 1/4 proximal cecum.
A 20G sterile needle was used to puncture for 3 times in the
ligated cecum so as to squeeze a small amount of feces, but fecal
pollution should be avoided. The intestinal canal was soaked with 2
ml of 0.9% sodium chloride solution. The cecum was put back into
the abdominal cavity according to its physiological position, and
the abdominal cavity was closed layer by layer. After surgery,
abdominal subcutaneous injection of 0.9% sodium chloride solution
(50 mg/kg) was performed immediately for antishock. In the Sham
group, the abdomen of rats was opened and closed without cecal
ligation and puncture (CLP). No abdominal subcutaneous injection of
0.9% sodium chloride solution was performed after surgery (19). After the CLP, TKR-200C small animal
ventilator and blood gas analyzer (Jiangxi Teli Anesthesia
Breathing Equipment Co., Ltd., Nanchang, Jiangxi, China) were
employed to monitor the respiratory frequency and arterial partial
pressure of oxygen (PaO2) of rats.
Animal grouping and model
establishment
A total of 36 SD rats after 2-week feeding were
randomly divided into 3 groups with 12 rats in each group. The Ab
(anti-TLR4 monoclonal antibody)-CLP group: TLR4 antibody (10 mg/ml;
Cell Signaling Technologies, Inc., Beverly, MA, USA) was injected
into the caudal vein at 1 day before model establishment, and then
the model establishment of sepsis-associated ARDS was conducted.
The CLP group: The same volume of normal saline was injected into
the caudal vein at 1 day before model establishment, and then the
model establishment of sepsis-associated ARDS was conducted. The
Sham group: The same volume of normal saline was injected into the
caudal vein at 1 day before model establishment, and then the
abdomen of rats were opened and closed after anesthesia without
CLP.
Arterial blood gas analysis and
peripheral blood collection
At 6 h after the models were established
successfully, rats were anaesthetized with intraperitoneal
injection of 3% mebumalnatrium (Sigma-Aldrich; Merck KGaA,
Darmstadt, Germany) (30 mg/kg). After successful anesthesia, the
rats in the three groups were fully extended and fixed on the
operating table in the supine position. Hair of neck, chest and
abdomen was shaved using disposable skin preparation knife or
disinfection scissors, followed by disinfection. The abdominal
cavity was opened with midline abdominal incision to expose the
abdominal aorta. Arterial blood (0.3 ml) was collected using 1 ml
disposable syringe which had been washed with heparin. Blood gas
analysis of blood sample was carried out using blood gas analyzer
(Radiometer Medical A/S, Copenhagen, Denmark). The chest was opened
to expose the left atrium. The sterile forcep was used to find the
right atrium and blood was extracted from the right atrium using a
10 ml sterile syringe. The collected blood was centrifuged at low
temperature, and then the supernatant was collected and stored at
−80°C for further use.
Bronchoalveolar lavage fluid (BALF)
collection
The chest was opened immediately along the anterior
midline and the hilum of right lung was ligated with a 2 mm
transverse incision in the trachea. The scalp acupuncture (2 mm
outer diameter) removing the pinpoint was applied as tracheal
catheter and inserted into the left main bronchus. And then the
rats were slowly injected with normal saline (2.5 ml) through the
catheter. When the left lung of rats was gradually swelled and the
color became pale, the normal saline was slowly pumped back. The
lavage step was repeated 5 times with a lavage recovery criterion
of the recovery liquid >2 ml each time. BALF was collected in
the 20 ml plastic centrifuge tube with ice bath at 4°C, and then
centrifuged in low speed centrifuge at a speed of 4,000 × g for 10
min. The supernatant was sub-packed and stored at −80°C for the
detection of inflammatory cytokines using enzyme-linked
immunosorbent assay (ELISA). After centrifugation, the cell
sedimentation was resuspended with a pre-cooled RPMI-l640 medium
for alveolar macrophage extraction and follow-up experiments.
Alveolar macrophage separation
The BALF collected from the Sham, CLP and Ab-CLP
groups was centrifugated, the cell sedimentation of which was added
with DMEM medium containing 15% fetal bovine serum (FBS) in order
to resuspend the cells. Cell suspension was placed in the 6-well
plates with adherent culture for 2 h under 5% CO2 at
37°C. After removal of the cell culture medium, the adherent
alveolar macrophages were collected. After wright staining, cells
with the gray cytoplasm and the purple red nucleus under
microscopic examination were confirmed as macrophage with a purity
of >95%. Cells were observed under the microscope after trypan
blue exclusion test. The dead cells were stained as light blue,
while the living cells were not stained. Cell viability was
calculated in accordance with the following formula: Cell viability
(%)=(the number of total cells-the number of stained cells)/the
number of total cells ×100%. From the above formula, cell viability
(cell viability of normal cells >95%) was calculated, which
indicated that the cells could be used for subsequent experiments
with good growth and strong viability (20).
Lung tissue extraction
After lavage of the left lung alveolar, the upper
lobe of the right lung was immediately resected and fixed in
formaldehyde solution for pathological examination. After the
middle lobe of the right lung was resected, the surface of the lobe
was rinsed immediately with normal saline and dried with a filter
paper, and then it was weighed as wet weight. After that, it was
put in a 65°C oven and baked for at least 48 h. Then, it was
weighed repeatedly to obtain constant dry weight. The ratios of wet
weight/dry weight (W/D) were calculated. After the lower lobe of
the right lung was resected, the blood in the surface was washed
using the ddH2O of RNA-Free and DNA-Free. The lower lobe
of the right lung was cut into two parts, and sub-packed with
tinfoil which was baked in a 260°C oven in order to inactivate
enzyme. After that it was quickly stored in liquid nitrogen. Then,
it was transferred and stored in refrigerator at −80°C. One part
was used for quantitative polymerase chain reaction (qPCR) and the
other for western blotting.
Hematoxylin and eosin (H&E)
staining
The samples were embedded in paraffin and sliced
into small sections of 5 µm thickness. The sections were routinely
dewaxed with xylene for 30 min, followed by gradient ethanol
dehydration with 100% ethanol, 95% ethanol, 85% ethanol and 70%
ethanol, respectively for 2 min. The tissue sections were immersed
in hematoxylin for 15 min. Then the tissue sections were immersed
in 1% hydrochloric alcohol for 6–8 sec and in saturated lithium
carbonate solution for 1 min. Subsequently, the tissue sections
were immersed in eosin staining solution for 4 min (21).
Transmission electron microscope (TEM)
observation
The lung tissue (1.0×1.0×1.0 mm) was washed by
phosphate buffer saline (PBS) pre-cooled at 4°C, and then the
tissue was fixed with 4 ml of 2% glutaraldehyde (PH 7.4; pre-cooled
at 4°C) for 30 min. The lung tissue was rinsed by PBS for 3 times,
10 min each time. The lung tissue was fixed in 1% osmium acid at
4°C for 30 min, following which the lung tissue was rinsed by PBS
for 3 times. The lung tissue was dehydrated at room temperature
using gradient concentrations of acetone: 50% acetone for once with
10 min each time, 70% acetone for once with 10 min each time, 90%
acetone for twice with 10 min each time and 100% acetone for three
times with 10 min each time. After the dehydrating agent in the
bottle was discarded, the lung tissue was embedded with 3 ml of
pure acetone-EPON812 for 30 min at room temperature. After the
removal of the diluted embedding agent, the lung tissue was
embedded with 1 ml of pure embedding agent overnight at room
temperature. Two drops of the mixed embedding medium were dropped
to the bottom of the capsule module hole. After moving the lung
mass to the center of the module, it was filled with the mixed
embedding medium and then baked in a 60°C oven for 24 h. The
embedding block was trimmed for the preparation of semi-thin
sections, and ultrathin sections were prepared after accurate
positioning. The sections were stained using lead acetate uranium,
after which the tissue sections were observed under TEM (Hitachi
High-Technologies Corporation, Tokyo, Japan).
Measurement of lung wet weight/dry
weight (W/D) ratios
Part of tissues in the middle lobe of the right lung
was selected and the exudate and blood on the surface of which was
absorbed using filter paper. The wet weight of the lung was weighed
using the electronic balance (Mettler-Toledo International Inc.,
Zurich, Switzerland) and then baked in a 70°C electrothermal
constant-temperature dry box (Shanghai Jinghong Laboratory
Instrument Co., Ltd., Shanghai, China) until it weighed to constant
weight as dry weight. The lung coefficient was calculated in
accordance with the following formula: W/D=wet weight of lung/dry
weight of lung, lung water content=(net wet weight of lung-net dry
weight of lung)/net wet weight of lung × 100%.
Enzyme-linked immunosorbent assay
(ELISA)
The BALF and peripheral blood that collected and
stored separately in the refrigerator was defrosted on the ice. The
ELISA kit was taken out from the 4°C refrigerator, and maintained
at room temperature for 30 min. The ELISA kit for TNF-α and
interleukin-1β (IL-1β) was purchased from Cusabio Biotech Co., Ltd.
(Wuhan, China), and the experimental procedures were carried out in
strict accordance with the instructions of kit. The blank well was
applied for zero calibration and the optical density (OD) value of
each well was measured successively using a microplate reader
(Bio-Tek Instruments Inc., Winooski, Vermont, USA) at a wavelength
of 450 nm.
Reverse transcription-qPCR
(RT-qPCR)
The TLR4, TLR9, MyD88 and NF-κΒ mRNA expressions in
alveolar macrophage were measured by qRT-PCR. The total RNA was
extracted from alveolar macrophage using trizol method (Takara
Biotechnology Co., Ltd., Dalian, Liaoning, China), and the
concentration and purity of RNA were detected. Reverse
transcription kit (DRR047S; Takara Biotechnology Co., Ltd.) was
used to reversely transcribe RNA into cDNA with 10 µl of reverse
transcription system. The cDNA obtained from above reversion was
diluted with 65 µl of diethyl pyrocarbonate (DEPC) solution with
sufficient mixing. Reaction system: 5 µl of SsoFast EvaGreen
Supermix (1708882; Bio-Rad Laboratories, Inc., Hercules, CA, USA),
0.5 µl of Forward primer (10 µM), 0.5 µl of Reverse primer (10 µM)
and 4 µl of cDNA. PCR amplification conditions: Pre-denaturation at
95°C for 1 min, 30 cycles of denaturation at 95°C for 30 sec and
annealing at 58°C for 5 sec, and final extension at 72°C for 5 sec.
Primers were synthesized by Beijing Genomics Institute (BGI,
Shenzhen, China) (Table I). β-actin
was used as an internal reference. Each gene in the samples was
measured in triplicates. The reliability of the PCR results was
verified using the solubility curve. The Cquantification
cycle (Cq) value was obtained. Formula: ∆Cq=Cqtarget
gene-Cqinternal reference, ∆∆Cq=∆Cqthe
experimental group-∆Cqthe control group. And the
gene expressions were calculated using 2−∆∆Cq method
(22). The experiment was repeated
three times.
 | Table I.Primer sequences for quantitative
polymerase chain reaction. |
Table I.
Primer sequences for quantitative
polymerase chain reaction.
| Gene | Primer sequences
(5′-3′) |
|---|
| TLR4 |
|
| F |
CGCTCTGGCATCATCTTCAT |
| R |
CTCCTCAGGTCAAAGTTGTTGC |
| TLR9 |
|
| F |
CCTGGCACACAATGACATTCA |
| R |
TAAAGGTCCTCCTCGTCCCA |
| MyD88 |
|
| F |
GAGATCCGCGAGTTTGAGAC |
| R |
TTGTCTGTGGGACACTGCTC |
| NF-κB |
|
| F |
GAGGACTTGCTGAGGTTGG |
| R |
TGGGGTGGTTGATAAGGAGTG |
| GAPDH |
|
| F |
AACGGATTTGGTCGTATTGGG− |
| R |
TCGCTCCTGGAAGATGGTGAT |
Western blotting
Alveolar macrophages were collected and digested
with trypsin. After centrifugation, alveolar macrophages were
washed by PBS for two times, and then cracked with pre-cooled cell
lysate on ice for 30 min. Then, they were centrifuged at 12,000 × g
for 20 min at 4°C, after which the supernatant was collected and
stored at −20°C. The 2 µg/µl of bovine serum albumin (BSA) was
diluted with PBS into following concentrations: 20, 15, 10, 5, 2.5,
0 µg/ml. bicinchoninic acid (BCA) protein assay kit (Thermo Fisher
Scientific, Inc., Waltham, MA, USA) was used to detect protein
concentration in accordance with the procedure of instructions
combined with sample number. Protein electrophoresis was performed
in a 4°C chromatography cabinet. The electrophoresis was performed
with compressed gels at the voltages of 80 V and then separation
gels at 120 V. After electrophoresis, the proteins were transferred
onto polyvinylidene fluoride (PVDF) membrane using wet transfer,
and then they were blocked using 5% skimmed milk-tris-buffered
saline (TBST) and incubated at room temperature for 2 h. Protein
samples were incubated at 4°C for overnight after being added with
primary antibodies against TLR4 (1:500; Cell Signaling
Technologies, Inc.), TLR9 (1:800), MyD88 (1:900), NF-κB (1:1,000),
and GAPDH (1:8,000; Proteintech Group, Inc. Chicago, IL, USA).
Samples were rinsed by TBST three times, 10 min each time. Samples
were added with the second antibody (1:10,000), and incubated at
room temperature for 1 h. Then, samples were rinsed by TBST three
times, 10 min each time. Finally, the samples were stained using
chemiluminescence and tabulated by X-ray, followed by film
development and fixation, and finally results were analyzed.
Statistical analysis
Data were analyzed statistically using SPSS 22.0
software (IBM Corp, Armonk, NY, USA). Measurement data were
presented as the mean ± standard deviation (SD). The comparisons
among multiple groups were conducted by the one-way analysis of
variance (ANOVA), and the comparisons between two groups were
tested by the t-tests. P<0.05 was considered to indicate a
statistically significant difference.
Results
Respiratory frequency and arterial
PaO2 of rats in three groups
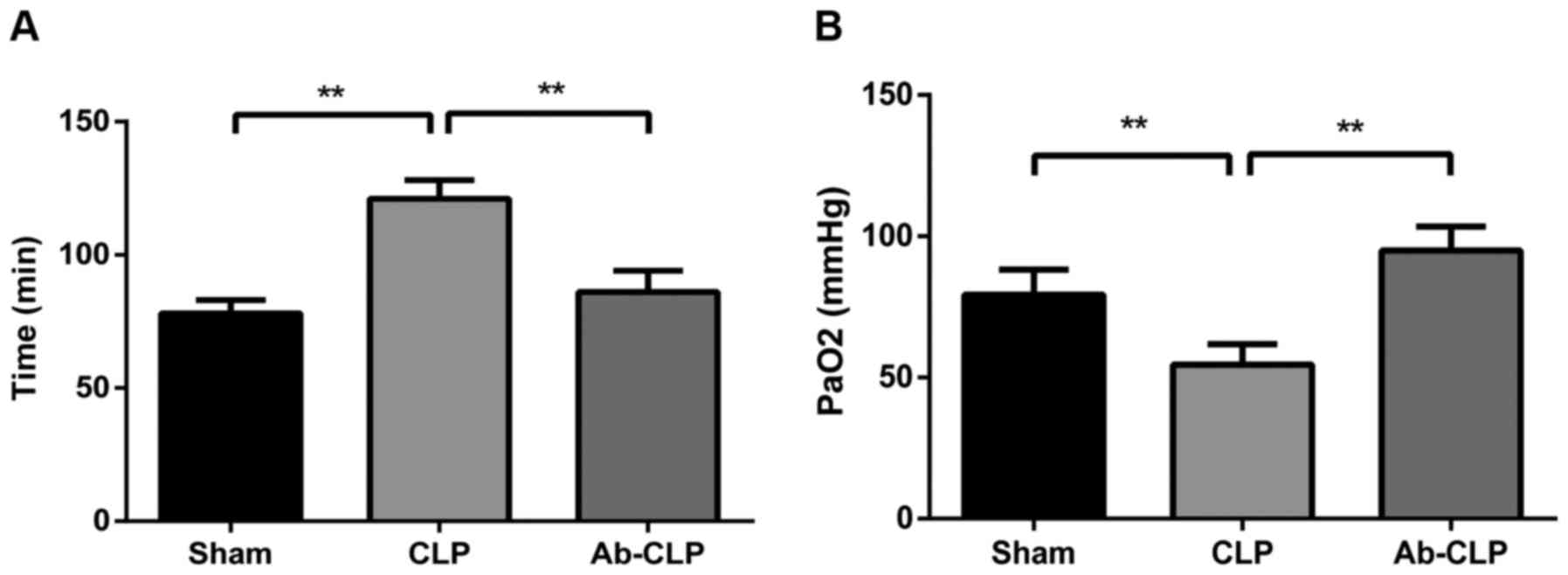
The respiratory frequencies of rats in the Sham, CLP
and Ab-CLP groups were 78±5, 121±7 and 86±8 breaths/min,
respectively. The respiratory frequencies of rats in the CLP group
were significantly higher than that in the Sham and Ab-CLP groups
(P<0.05). Arterial PaO2 in the Sham, CLP
and Ab-CLP groups were 79.3±8.9, 54.6±7.2 and 74.9±6.9 mmHg,
respectively. In comparison to the CLP group, the Sham and Ab-CLP
groups showed decreased arterial PaO2
(P<0.05) (Fig. 1).
Pathological changes of lung tissue of
rats in three groups
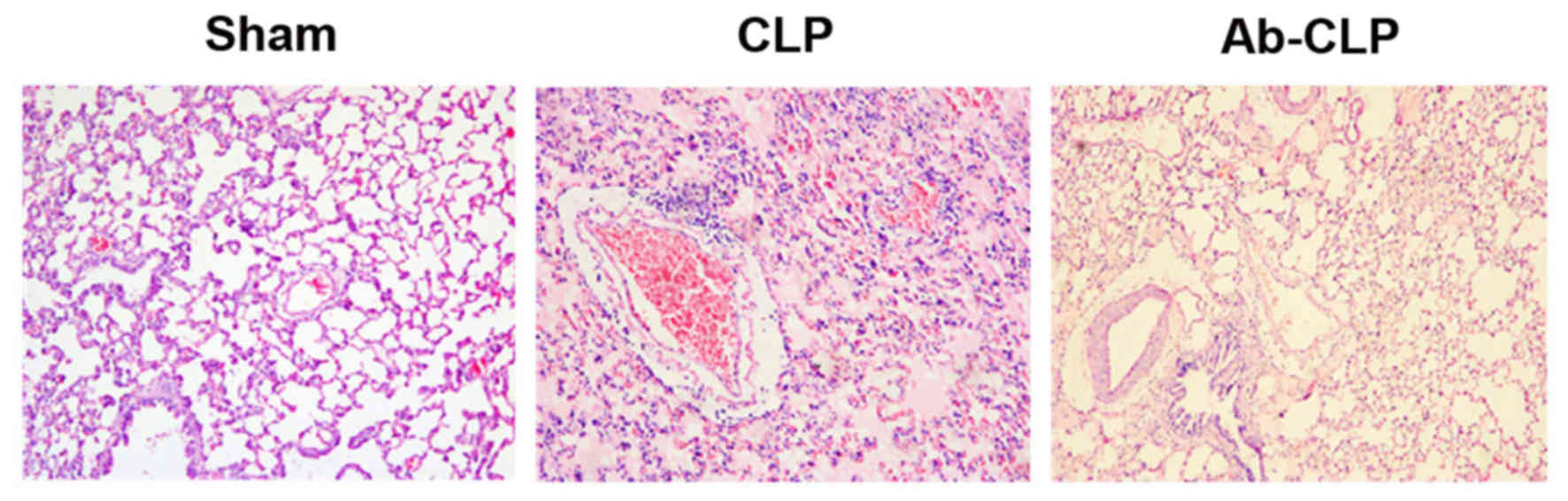
Lung tissue samples were observed under the light
microscope. In the Sham group, the lung tissue structure was
complete and clear, and less interstitial inflammatory cell
infiltration was visible in alveolar without obvious congestion and
hemorrhage. Besides, the alveolar septum in the Sham group was not
widened, and the alveolar wall was complete without edema in the
cavity. In the CLP group, a large area of alveolar wall was
destructed in the lung tissue of rats. Additionally, the alveolar
cavity was filled with pink edema fluid with widened alveolar
septum and edema. A large number of inflammatory cells were
infiltrated and red blood cells (RBCs) were leaked out with
pulmonary vascular congestion. In the Ab-CLP group, the lung tissue
of rats still had the same pathological changes as that in the CLP
group, but the size of damaged area, alveolar edema fluid, alveolar
interstitial edema, inflammatory cell infiltration and leakage of
RBCs were significantly reduced (Fig.
2).
Cellular structure of type II alveolar
epithelial cells under transmmision electron microscope
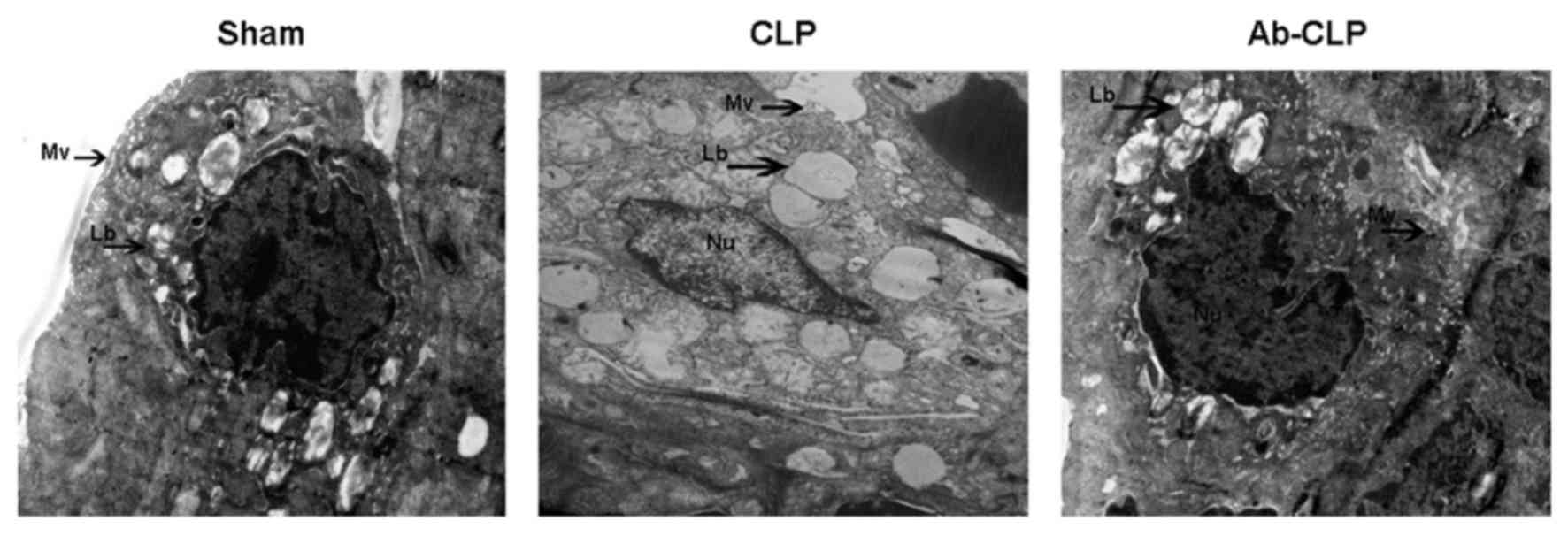
The results of high resolution transmission electron
microscope are shown in Fig. 3. In
the CLP group, the nucleus and its boundary was clear with
karyopyknosis and obvious shape changing; the nucleolus and the
cell membrane were broken with disordered cytoplasm structure and a
large number of vacuoles; the microvillus almost disappeared with
badly damaged organelle, vague shape of nucleus and obvious
perinuclear space; the nucleolus presented apoptosis status with
obvious exudation in alveolar space. The type II alveolar
epithelial cells have integrated cell membrane in the Sham and
Ab-CLP groups. Meanwhile, the cytoplasm and the nuclear structure
were clear with a small amount of cytoplasmic cavitation and slight
perinuclear space but no obvious cell damage in the Sham and Ab-CLP
groups. The chromatin is evenly aligned with continuous and
integrated cell membrane.
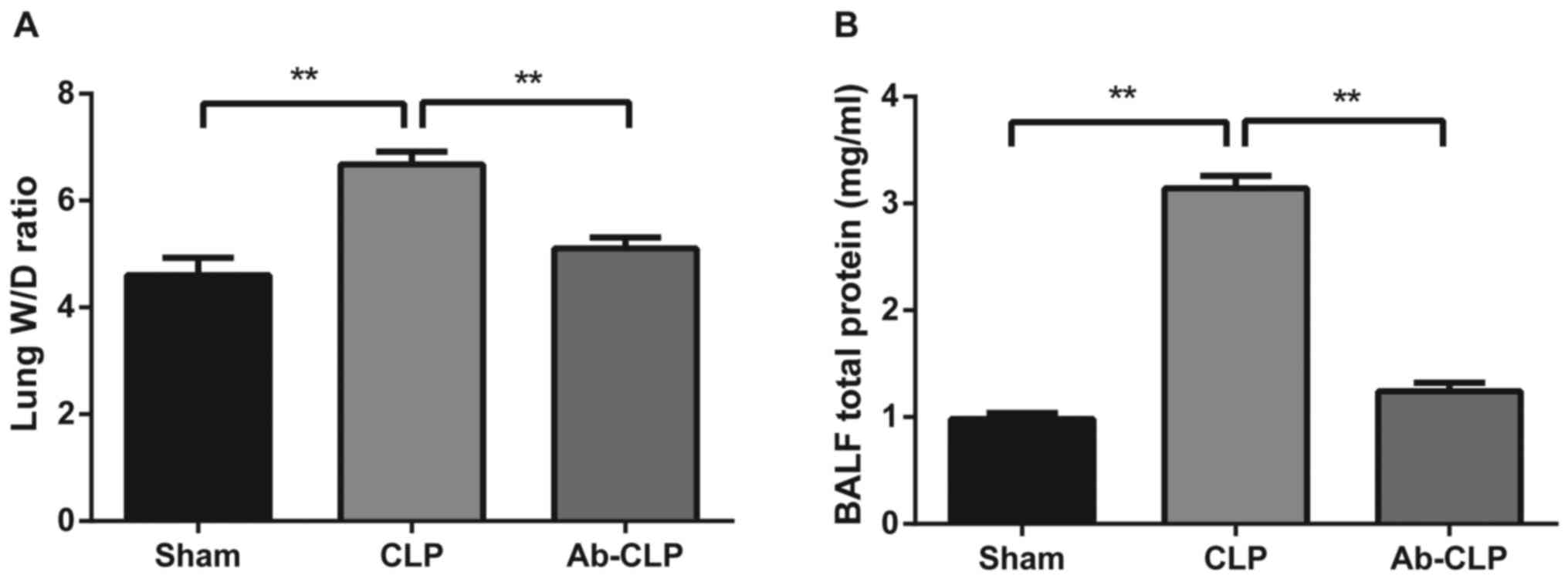
Comparisons of lung W/D ratios and
total protein content in BALF of rats in three groups
In the Sham, CLP and Ab-CLP groups, lung W/D ratios
were 4.214±0.324; 6.678±0.241 and 5.901±0.212, respectively. The
lung W/D ratios in the CLP group were significantly higher than
that in the Sham and Ab-CLP groups (P<0.05; Fig. 4A). In consistent with the results of
pathological examination, it was suggested that the lung tissue of
rats in the CLP group had obvious pulmonary edema. The W/D ratios
of lung tissue were reduced after pretreatment with anti-TLR4
monoclonal antibody (anti-TLR4 mAb), which indicated that CLP could
reduce the degree of pulmonary edema after pretreatmentm of
anti-TLR4 mAb. In comparison to the CLP group, the total protein
contents in BALF in the Sham and Ab-CLP groups were evidently
decreased (P<0.05; Fig. 4B). It
was suggested that the total protein content in BALF was
significantly increased in the CLP group, but that was decreased
after pretreatment with anti TLR-4 mAb.
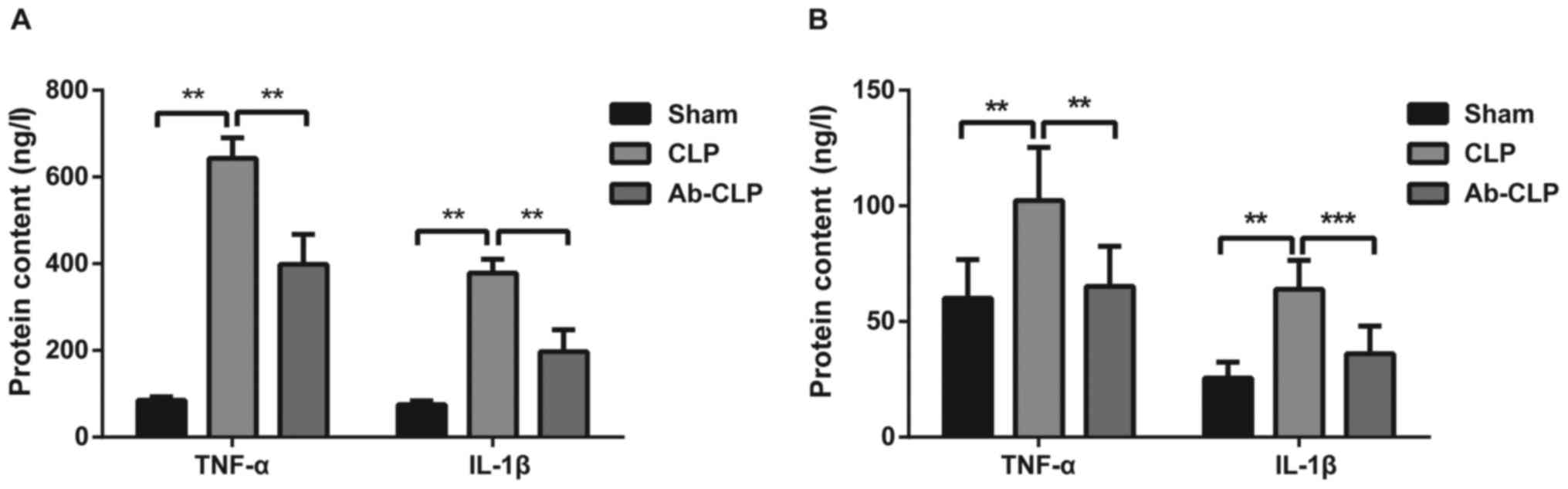
Pretreatment with anti TLR-4 mAb
reduced the protein contents of inflammatory factors (TNF-α and
IL-1β) in BALF and peripheral blood
The protein contents of TNF-α and IL-1β in BALF and
peripheral blood of rats in three groups were measured using ELISA
(Fig. 5). The results showed that
the protein contents of TNF-α and IL-1β in the Sham group were
85.14±7.82 and 75.16±8.62, respectively. The protein contents of
TNF-α and IL-1β in the CLP group were 643.15±47.18 and
378.06±32.19, respectively. The protein contents of TNF-α and IL-1β
in the Ab-CLP group were 398.46±69.12 and 197.15±50.37,
respectively. The protein contents of TNF-α and IL-1β in the CLP
group were significantly higher than these in the Sham and Ab-CLP
groups (P<0.05). It was suggested that the pretreatment with
anti-TLR4 mAb reduced the synthesis and secretion of inflammatory
factors induced by sepsis-associated ARDS, and alleviated the
inflammatory response.
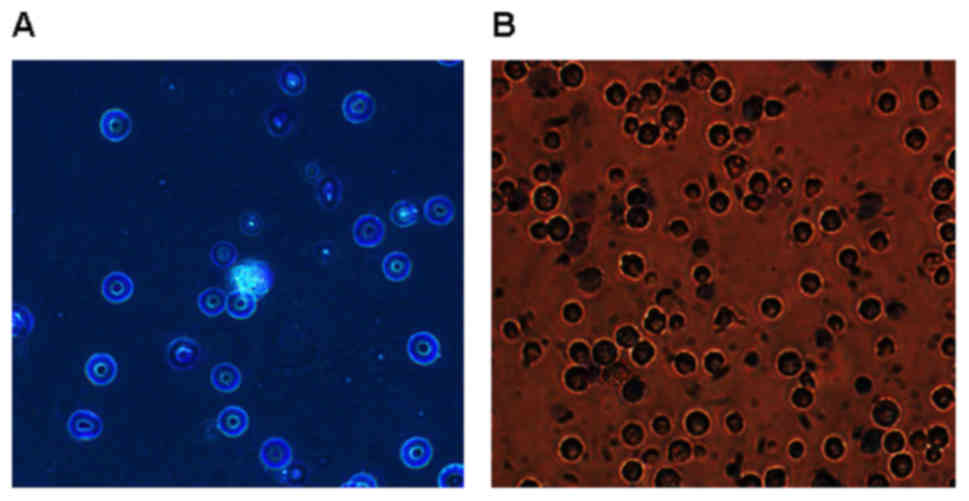
The purity and cell viability of
alveolar macrophage
Alveolar macrophages were obtained by
bronchoalveolar lavage with a purity over 95%. The isolated
alveolar macrophages were round or oval with clear and neat edge,
the cytoplasm of which was slightly gray with pale purple red
cytoplasmic granules and deep purple red nucleus in the cells. The
purple-red nucleus and the pale red cytoplasm were occasionally
visible with pale purple red neutrophile granulocyte. As shown in
Fig. 6, cell viability is >95%
after trypan blue staining.
Pretreatment with anti-TLR4 mAb inhibited TLR4/MyD88
signaling pathway in alveolar macrophage of rats with
sepsis-associated ARDS. The mRNA and protein expressions of
TLR4/MyD88 signaling pathway-related factors (TLR4, TLR9, MyD88 and
NF-κΒ P65) in alveolar macrophage of rats with sepsis-associated
ARDS were measured by qRT-PCR and western blotting (Fig. 7). The results showed that TLR4/MyD88
signaling pathway was activated after CLP, and the expressions of
TLR4, MyD88, NF-κΒ P65 mRNAs and proteins in the CLP group were
significantly higher than these in the Sham group (P<0.05). In
comparison to the CLP group, the Ab-CLP group showed the
inactivation of TLR4/MyD88 signaling pathway after pretreatment
with anti-TLR4 mAb. There was no significant difference in the TLR9
protein and mRNA expressions between the CLP and Ab-CLP groups. But
TLR9 protein and mRNA expressions in the CLP and Ab-CLP groups were
significantly higher than that in the Sham group (P<0.05). It
was suggested that pretreatment with anti-TLR4 mAb inhibited the
TLR4/MyD88 signaling pathway in alveolar macrophage pf rats with
sepsis-associated ARDS.
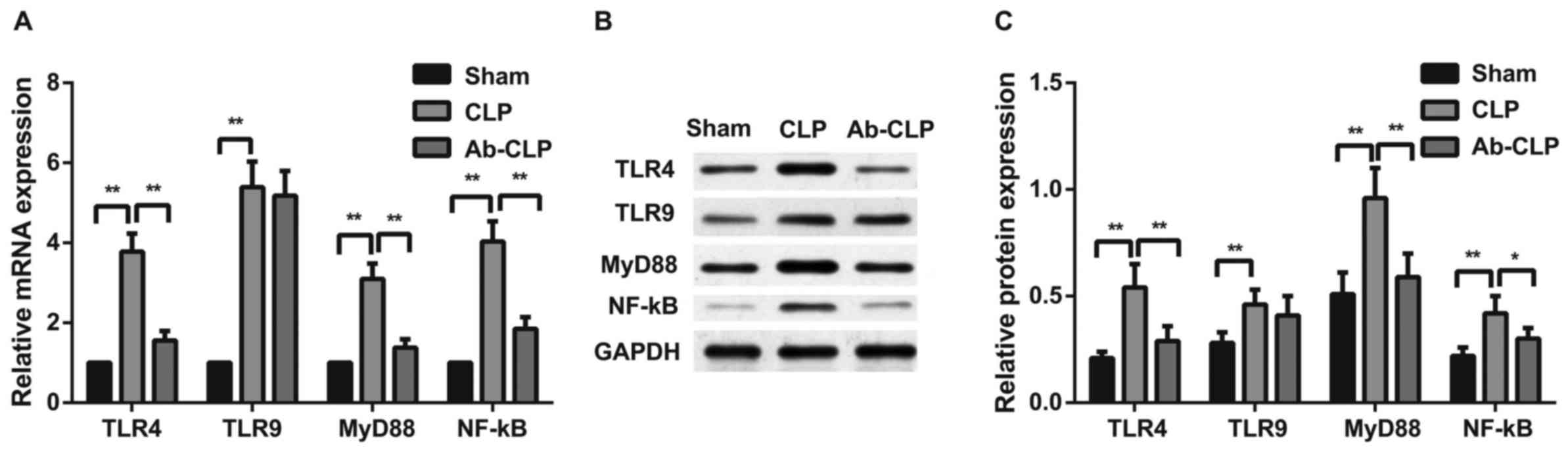 | Figure 7.Comparison of the mRNA and protein
expressions of TLR4/MyD88 signaling pathway-related factors in
alveolar macrophage among the three groups. (A) Comparison of TLR4,
TLR9, MyD88 and NF-κΒ P65 mRNA expressions in alveolar macrophage
of rats among the Sham, CLP and Ab-CLP groups; (B) protein bands of
TLR4, TLR9, MyD88 and NF-κΒ P65 in the Sham, CLP and Ab-CLP
detected by western blotting; (C) comparison of TLR4, TLR9, MyD88
and NF-κΒ P65 protein expressions in alveolar macrophage of rats
among the Sham, CLP and Ab-CLP groups; **P<0.005 compared with
the CLP group; *P<0.05 compared with the CLP group. CLP, cecal
ligation puncture; Ab, anti-TLR-4 monoclonal antibody; TLR4,
Toll-like receptor 4; TLR9, Toll-like receptor 9; MyD88, myeloid
differentiation primary-response protein 88; NF-κB, nuclear
factor-κB; GAPDH, glyceraldehyde-3-phosphate dehydrogenase. |
Discussion
Over the past few years, TLR4/MyD88 signaling
pathway plays a crucial role in various diseases including cerebral
ischemia/reperfusion injury, diabetic liver injury and
ventilator-induced lung injury (16,23,24). In
this study, we further investigated the effects of TLR4/MyD88
signaling pathway on rats with sepsis-associated ARDS by regulating
macrophage activation, and found that the inhibition of TLR4/MyD88
signaling pathway may relieve sepsis-associated ARDS through
regulating macrophage activation and inflammatory response.
One of the main findings in our study was that in
comparison to the CLP group, the Ab-CLP group showed significantly
decreased damaged area, alveolar edema fluid, alveolar interstitial
edema, inflammatory cell infiltration and leakage of red blood
cells (RBCs) which indicated that the pretreatment with anti-TLR4
mAb alleviated lung injury, pulmonary edema and inflammatory
infiltration of lung tissue in a rat model of sepsis-associated
ARDS. Rats in the Ab-CLP group were treated with anti-TLR4 mAb
before CLP. Importantly, mAbs account for a large proportion of
medicines in effective treatments for the patients with a variety
of inflammatory diseases, and mAbs are created to inhibit TLR4
signaling pathway with a FcγR-binding mechanism (25). In line with our study, Huang et
al also demonstrated that anti-TLR4 mAb attenuated the lung
injury caused by mechanical ventilation, inflammation and edema in
rats by inhibiting TLR4/MyD88 signaling pathway (15), but our study indicated that
inhibition of TLR4/MyD88 signaling pathway may relieve
sepsis-associated ARDS in rats through regulating macrophage
activation and inflammatory response. Thus, the inhibition of
TLR4/MyD88 signaling pathway might contribute to less lung injury,
inflammation and edema in rats with sepsis-associated ARDS.
Additionally, in our study, the results of ELISA
indicated the expressions of TNF-α and IL-1β in the CLP group was
significantly higher than that in the Sham and Ab-CLP groups,
suggesting that pretreatment of anti-TLR4 mAb reduced the synthesis
and secretion of inflammatory factors in sepsis-associated ARDS,
and alleviated the inflammatory response. Macrophage responses are
coordinated through classical (or M1) and alternative (or M2)
activation programs (26). The
functions of macrophage are greatly determined by their activation
states, exerting a causal role of macrophage activation in the
initiation of inflammation (27).
TNF-α and IL-1β were major pro-inflammatory cytokines and both were
mainly created by the activated macrophages (28). A study conducted by Lin et al
once demonstrated that the inhibition of TLR4/MyD88 signaling
pathway reduced the expressions of proinflammatory cytokines IL-1β
and TNF-α, thereby ameliorating the proinflammatory phenotype
(29), which is in consistent with
our investigation. Inflammatory cytokines, including IL-2, IL-4,
IL6, IL8 and IL-1β, correlates with the mortality of ARDS (30,31).
From the results above, we see that the inhibition of the
TLR4/MyD88 signaling pathway may have protective effects on
sepsis-associated ARDS through suppressing inflammatory response,
which could be regulated by macrophage activation.
Surprisingly, the results revealed that TLR4/MyD88
signaling pathway was activated after CLP, the mRNA and protein
expressions of TLR4, MyD88, NF-κΒ P65 in the CLP group was
significantly higher than that in the Sham group. While in
comparison to the CLP group, the Ab-CLP group showed the
inactivation of TLR4/MyD88 signaling pathway after pretreatment
with anti-TLR4 mAb. TLR4 signaling in macrophages regulates
hundreds of gene expressions stimulating anti-microbial activity
and induces inflammatory signaling pathways driving inflammatory
responses (32). Dai et al
revealed that the high expression of TLR4 and TLR9 in alveolar
macrophage played a critical role in ventilator-induced lung
injury, through the recruitment of MyD88, which could activate
NF-κΒ inducing the transcription of various pro-inflammatory genes
(33). NF-κΒ acts as a nuclear
factor, the secretion of pro-inflammatory cytokines, such as IL-6,
IL-8, and TNF-α, that could be mediated by the MyD88 signaling
(34). Hence, from all that
mentioned above, the pretreatment with anti-TLR4 mAb inhibited the
expressions of TLR4, decreased the secretion of MyD88 thereby
suppressing the activation of NF-κΒ, which reduced the inflammatory
response to sepsis-associated ARDS.
Consequently, our present study provided evidence
that the inhibition of TLR4/MyD88 signaling pathway might relieve
sepsis-associated ARDS in rats through regulating macrophage
activation. It is believed that the pretreatment of anti-TLR4 mAb
inhibited the TLR4/MyD88 signaling pathway and alleviated
inflammatory response. Further studies are needed to understand the
molecular mechanism underlying the role of TLR4/MyD88 signaling
pathway in sepsis-associated ARDS and thus to support its potential
clinical application.
References
|
1
|
Han S and Mallampalli RK: The acute
respiratory distress syndrome: From mechanism to translation. J
Immunol. 194:855–860. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lin WC, Chen CW, Huang YW, Chao L, Chao J,
Lin YS and Lin CF: Kallistatin protects against sepsis-related
acute lung injury via inhibiting inflammation and apoptosis. Sci
Rep. 5:124632015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Wang M, Yan J, He X, Zhong Q, Zhan C and
Li S: Candidate genes and pathogenesis investigation for
sepsis-related acute respiratory distress syndrome based on gene
expression profile. Biol Res. 49:252016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Xu Z, Huang Y, Mao P, Zhang J and Li Y:
Sepsis and ARDS: The dark side of histones. Mediators Inflamm.
2015:2050542015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Boyle AJ, Mac Sweeney R and McAuley DF:
Pharmacological treatments in ARDS; a state-of-the-art update. BMC
Med. 11:1662013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Dizier S, Forel JM, Ayzac L, Richard JC,
Hraiech S, Lehingue S, Loundou A, Roch A, Guerin C and Papazian L;
ACURASYS study investigators, ; PROSEVA Study Group, : Early
hepatic dysfunction is associated with a worse outcome in patients
presenting with acute respiratory distress syndrome: A post-Hoc
analysis of the ACURASYS and PROSEVA studies. PLoS One.
10:e01442782015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Mansur A, Steinau M, Popov AF, Ghadimi M,
Beissbarth T, Bauer M and Hinz J: Impact of statin therapy on
mortality in patients with sepsis-associated acute respiratory
distress syndrome (ARDS) depends on ARDS severity: A prospective
observational cohort study. BMC Med. 13:1282015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
de Pablo R, Monserrat J, Reyes E, Díaz D,
Rodríguez-Zapata Mla Hera Ad, Prieto A and Alvarez-Mon M:
Sepsis-induced acute respiratory distress syndrome with fatal
outcome is associated to increased serum transforming growth factor
beta-1 levels. Eur J Intern Med. 23:358–362. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Herwig MC, Tsokos M, Hermanns MI,
Kirkpatrick CJ and Müller AM: Vascular endothelial cadherin
expression in lung specimens of patients with sepsis-induced acute
respiratory distress syndrome and endothelial cell cultures.
Pathobiology. 80:245–251. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Avlas O, Fallach R, Shainberg A, Porat E
and Hochhauser E: Toll-like receptor 4 stimulation initiates an
inflammatory response that decreases cardiomyocyte contractility.
Antioxid Redox Signal. 15:1895–1909. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Liu L, Gu H, Liu H, Jiao Y, Li K, Zhao Y,
An L and Yang J: Protective effect of resveratrol against
IL-1β-induced inflammatory response on human osteoarthritic
chondrocytes partly via the TLR4/MyD88/NF-κB signaling pathway: An
‘in vitro study’. Int J Mol Sci. 15:6925–6940. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kim KH, Jo MS, Suh DS, Yoon MS, Shin DH,
Lee JH and Choi KU: Expression and significance of the TLR4/MyD88
signaling pathway in ovarian epithelial cancers. World J Surg
Oncol. 10:1932012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhu HT, Bian C, Yuan JC, Chu WH, Xiang X,
Chen F, Wang CS, Feng H and Lin JK: Curcumin attenuates acute
inflammatory injury by inhibiting the TLR4/MyD88/NF-κB signaling
pathway in experimental traumatic brain injury. J
Neuroinflammation. 11:592014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Castoldi A, Braga TT, Correa-Costa M,
Aguiar CF, Bassi ÊJ, Correa-Silva R, Elias RM, Salvador F,
Moraes-Vieira PM, Cenedeze MA, et al: TLR2, TLR4 and the MYD88
signaling pathway are crucial for neutrophil migration in acute
kidney injury induced by sepsis. PLoS One. 7:e375842012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Huang C, Pan L, Lin F, Dai H and Fu R:
Monoclonal antibody against Toll-like receptor 4 attenuates
ventilator-induced lung injury in rats by inhibiting MyD88- and
NF-κB-dependent signaling. Int J Mol Med. 39:693–700. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Han LP, Li CJ, Sun B, Xie Y, Guan Y, Ma ZJ
and Chen LM: Protective effects of celastrol on diabetic liver
injury via TLR4/MyD88/NF-κB signaling pathway in type 2 diabetic
rats. J Diabetes Res. 2016:26412482016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Barberà-Cremades M, Baroja-Mazo A and
Pelegrín P: Purinergic signaling during macrophage differentiation
results in M2 alternative activated macrophages. J Leukoc Biol.
99:289–299. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Chávez-Sánchez L, Garza-Reyes MG,
Espinosa-Luna JE, Chávez-Rueda K, Legorreta-Haquet MV and
Blanco-Favela F: The role of TLR2, TLR4 and CD36 in macrophage
activation and foam cell formation in response to oxLDL in humans.
Hum Immunol. 75:322–329. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Rittirsch D, Huber-Lang MS, Flierl MA and
Ward PA: Immunodesign of experimental sepsis by cecal ligation and
puncture. Nat Protoc. 4:31–36. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhang Z, Watt NJ, Hopkins J, Harkiss G and
Woodall CJ: Quantitative analysis of maedi-visna virus DNA load in
peripheral blood monocytes and alveolar macrophages. J Virol
Methods. 86:13–20. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Thompson JH and Richter WR:
Hemotoxylin-eosin staining adapted to automatic tissue processing.
Stain Technol. 35:145–148. 1960. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Tuo YL, Li XM and Luo J: Long noncoding
RNA UCA1 modulates breast cancer cell growth and apoptosis through
decreasing tumor suppressive miR-143. Eur Rev Med Pharmacol Sci.
19:3403–3411. 2015.PubMed/NCBI
|
|
23
|
Wang Y, Chen G, Yu X, Li Y, Zhang L, He Z,
Zhang N, Yang X, Zhao Y, Li N and Qiu H: Salvianolic acid B
ameliorates cerebral ischemia/reperfusion injury through inhibiting
TLR4/MyD88 signaling pathway. Inflammation. 39:1503–1513. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Huang C, Pan L, Lin F, Qian W and Li W:
Alveolar macrophage TLR4/MyD88 signaling pathway contributes to
ventilator-induced lung injury in rats. Xi Bao Yu Fen Zi Mian Yi
Xue Za Zhi. 31:182–189. 2015.(In Chinese). PubMed/NCBI
|
|
25
|
Loyau J, Malinge P, Daubeuf B, Shang L,
Elson G, Kosco-Vilbois M, Fischer N and Rousseau F: Maximizing the
potency of an anti-TLR4 monoclonal antibody by exploiting proximity
to Fcγ receptors. MAbs. 6:1621–1630. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Odegaard JI and Chawla A: Alternative
macrophage activation and metabolism. Annu Rev Pathol. 6:275–297.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Gong D, Shi W, Yi SJ, Chen H, Groffen J
and Heisterkamp N: TGFβ signaling plays a critical role in
promoting alternative macrophage activation. BMC Immunol.
13:312012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Jing X, Chen SS, Jing W, Tan Q, Yu MX and
Tu JC: Diagnostic potential of differentially expressed Homer1,
IL-1β and TNF-α in coronary artery disease. Int J Mol Sci.
16:535–546. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Lin X, Kong J, Wu Q, Yang Y and Ji P:
Effect of TLR4/MyD88 signaling pathway on expression of IL-1β and
TNF-α in synovial fibroblasts from temporomandibular joint exposed
to lipopolysaccharide. Mediators Inflamm. 2015:3294052015.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Ware LB, Koyama T, Zhao Z, Janz DR,
Wickersham N, Bernard GR, May AK, Calfee CS and Matthay MA:
Biomarkers of lung epithelial injury and inflammation distinguish
severe sepsis patients with acute respiratory distress syndrome.
Crit Care. 17:R2532013. View
Article : Google Scholar : PubMed/NCBI
|
|
31
|
Terpstra ML, Aman J, van Nieuw Amerongen
GP and Groeneveld AB: Plasma biomarkers for acute respiratory
distress syndrome: A systematic review and
meta-analysis'. Crit Care Med. 42:691–700. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Escoubet-Lozach L, Benner C, Kaikkonen MU,
Lozach J, Heinz S, Spann NJ, Crotti A, Stender J, Ghisletti S,
Reichart D, et al: Mechanisms establishing TLR4-responsive
activation states of inflammatory response genes. PLoS Genet.
7:e10024012011. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Dai H, Pan L, Lin F, Ge W, Li W and He S:
Mechanical ventilation modulates Toll-like receptors 2, 4, and 9 on
alveolar macrophages in a ventilator-induced lung injury model. J
Thorac Dis. 7:616–624. 2015.PubMed/NCBI
|
|
34
|
Chuffa LG, Fioruci-Fontanelli BA, Mendes
LO, Ferreira Seiva FR, Martinez M, Fávaro WJ, Domeniconi RF,
Pinheiro PF, Delazari Dos Santos L and Martinez FE: Melatonin
attenuates the TLR4-mediated inflammatory response through MyD88-
and TRIF-dependent signaling pathways in an in vivo model of
ovarian cancer. BMC Cancer. 15:342015. View Article : Google Scholar : PubMed/NCBI
|