Introduction
Dental pulp cells (DPCs) include unique mesenchymal
stem cells called dental pulp stem cells (DPSCs). Previous studies
have demonstrated that DPSCs have the potential for
multi-directional differentiation into osteoblasts, odontoblasts,
adipocytes, chondrocytes and neural-like cells in vitro
(1,2)
and may be useful seed cells for tissue engineering or stem cell
therapy in a number of disease conditions (3–5). DPSCs
may be obtained easily from deciduous or permanent teeth that are
discarded as medical waste. Previous studies have compared the
difference between DPSCs from deciduous teeth and permanent teeth
(6); however, differences between
DPSCs from permanent teeth of immature and mature patients have not
yet been described. A previous study established a number of DPC
lines isolated from extracted wisdom teeth as a cell source for
induced pluripotent stem (iPS) cells and the donor teeth of these
DPCs lines were divided into three developmental stages: Crown
completed (CC), root forming (RF) and root completed (RC) (7). Teeth from 14–16 year old patients were
defined as immature teeth (CC and RF), whereas teeth from 23–60
year old patients were defined as mature teeth (RC). The DPCs
obtained from immature permanent teeth exhibited a much higher
reprogramming potential compared with DPCs from mature permanent
teeth (7). Furthermore, distal-less
homeobox 4, a transcription factor that is highly expressed in DPCs
from immature permanent teeth, significantly promoted human iPS
cell generation as well as octamer-binding protein 3/4,
sex-determining region Y box 2 and Kruppel-like factor 4 (7). These findings suggest that the
biological characteristics of DPSCs differ in immature and mature
permanent teeth. In the present study, the molecular
characteristics of DPCs from immature and mature permanent teeth
were compared based on bioinformatics analysis. The results of the
present study may offer a basis for selecting a suitable
preservation time for DPCs, as well as developing a suitable
molecular intervention strategy for these cells to optimize
features for tissue engineering or stem cell therapy.
Materials and methods
Use of the Gene Expression Omnibus
(GEO) database to obtain differentially expressed genes in DPCs
from mature and immature permanent teeth
Tamaoki et al (7) previously used an array to analyze gene
expression profiles and investigate the effect of the developmental
stage of teeth on global gene expression in human DPCs. The
microarray data were deposited in the GEO online database
(https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE52853),
the detailed information for which was described in a previous
article and the online webpage (https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE52853)
(7). In the present study,
differentially expressed genes in DPCs from mature and immature
permanent teeth were acquired from the GEO database. The protocol
was described as follows: i) Search ‘dental pulp cells’ in the GEO
database (http://www.ncbi.nlm.nih.gov/gds/) to obtain the
GSE52853 datasets (Title: Human Dental Pulp Cells Mature vs.
Immature stages); ii) GEO2R analysis: Use GEO2R, one of GEO's own
data analysis tools, to identify differentially expressed genes in
DPCs from mature and immature permanent teeth. The default GEO2R
statistical tests were applied. The expression value was stated
with reference to that of mature permanent teeth and a moderated
t-statistic was used to compare the expression of genes in DPCs
from mature permanent teeth with those from immature permanent
teeth. Genes that differed most significantly (P<0.001;
|logFC|>1) were identified and were selected for further
analysis (8). Some genes with
P<0.001 and |logFC|<1 that serve a crucial role in tooth
development were also included in the present study. In total, 500
differentially expressed genes were selected for further
analysis.
Use of the Database for Annotation,
Visualization and Integrated Discovery (DAVID) to evaluate 500
differentially expressed genes in DPCs from mature and immature
permanent teeth
The DAVID bioinformatics database is an integrated
online database that includes a number of powerful annotation
resources, including Gene Ontology (GO), Biocarta and Kyoto
Encyclopedia of Genes and Genomes (KEGG) databases (9). In the present study, the DAVID
bioinformatics database was used to analyze the protein
associations, biological characteristics and functional
characteristics of differentially expressed genes in DPCs from
mature and immature permanent teeth. The protocol was as follows:
i) Access DAVID (https://david.ncifcrf.gov/) and copy the 500
differentially expressed gene names into box A; ii) select the
‘official gene symbol’ as the gene identifier type and click the
‘Submit List’ button; iii) choose ‘Homo sapiens’ as
annotations; and iv) select ‘Start Analysis’. The modified Fisher's
Exact test was used in the present study, and P<0.05 was
considered to indicate a statistically significant difference.
Use of online mendelian inheritance in
man (OMIM) to analyze the genetic characteristics of sonic hedgehog
(SHH)
The OMIM database is a comprehensive, authoritative
compendium of human genes and genetic phenotypes that is freely
available online. OMIM focuses on the association between genotype
and phenotype and the entries contain links to other genetics
resources. SHH was differentially expressed in DPCs from mature and
immature permanent teeth and exhibited complex effects in DPCs. In
the present study, the OMIM database was used to analyze the
genetic characteristics of SHH. The protocol was as follows: i)
Access the OMIM website (http://www.omim.org/), and enter ‘SHH’ in the search
box; ii) select ‘600725. SONIC HEDGEHOG; SHH’; iii) select ‘MGI
Mouse Gene’ from the ‘Animal Models’ menu; and iv) review the
genetic information about SHH, including ‘GO Classifications’,
‘Mutations, Alleles, and Phenotypes’ and ‘Expression’.
Use of the search Tool for the
Retrieval of Interacting Genes/Proteins (STRING) online database to
analyze functional protein association networks of SHH
STRING is a database of known and predicted
protein-protein interactions that includes direct (physical) and
indirect (functional) associations stemming from computational
prediction, interactions aggregated from other primary databases
and information transfer between organisms (10). In the present study, the STRING
online database was used to analyze functional protein association
networks of SHH. The protocol was as follows: i) Access the STRING
online database (http://www.string-db.org/); ii) enter the protein name
‘SHH’ and choose organism as ‘Homo sapiens’ or ‘Mus
musculus’ and search; and iii) obtain the network chart of
functional protein association networks of SHH.
Results
A total of 500 genes in DPCs were
demonstrated to be differentially expressed in mature and immature
permanent teeth
A number of differentially expressed genes in were
identified in DPCs from mature and immature permanent teeth via
analysis of GEO2R in the GEO database. The 500 genes that differed
most significantly (P<0.001) were selected for further analysis
using the DAVID online database.
Differentially expressed genes in DPCs
from mature and immature permanent teeth may be divided into
immunity, inflammation and cell signaling-associated biological
process subgroups
Following DAVID analysis, the 500 differentially
expressed genes in DPCs were divided into subgroups based on the GO
term ‘biological process’. The majority of these subgroups were
separated into three groups, associated with immunity, inflammation
and cell signaling. Genes in the immune-associated groups (Table I) were demonstrated to regulate
lymphocyte activation and proliferation, antigen processing and
negative regulation of immune system processes. A total of two
genes, including cluster of differentiation 24 and SHH, existed in
almost all subgroups. The 500 differentially expressed genes were
enriched into different KEGG pathway subgroups based on DAVID
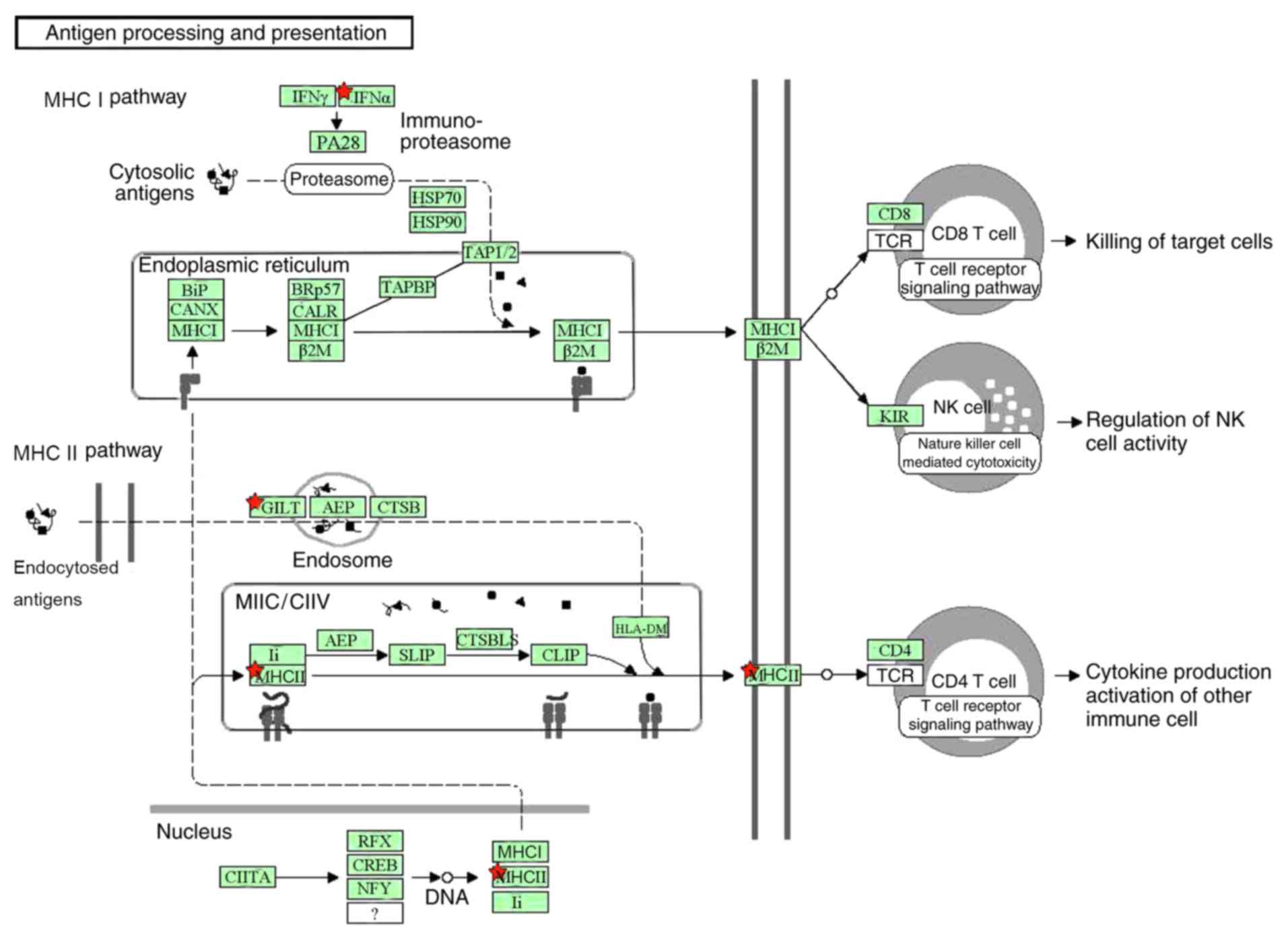
analysis. A total of four genes, including γ-interferon-inducible
lysosomal thiol reductase, human leukocyte antigen class II
histocompatibility antigen (HLA)-DR β 5 chain, HLA DP β 1 chain and
leukotriene A-4 hydrolase, were enriched into the ‘Antigen
processing and presentation’ pathway (Fig. 1). Genes in the
inflammation-associated groups (Table
II) may act to regulate leukocyte migration, proliferation,
activation and chemotaxis, mononuclear cell proliferation and the
defense response. Genes in the cell signaling-associated groups
(Table III) regulate chemotaxis,
cell-cell signaling transduction, cell surface receptor linked
signal transduction and G-protein coupled receptor protein
signaling pathways. These findings suggest that immune features,
response to infection and signaling may be different in DPCs from
mature and immature permanent teeth.
 | Table I.Differentially expressed genes in
dental pulp cells from mature and immature permanent teeth were
enriched into immunity associated subgroups based on ‘Biological
Process’ terms analyzed using the Database for Annotation,
Visualization and Integrated Discovery. |
Table I.
Differentially expressed genes in
dental pulp cells from mature and immature permanent teeth were
enriched into immunity associated subgroups based on ‘Biological
Process’ terms analyzed using the Database for Annotation,
Visualization and Integrated Discovery.
|
| Differentially
expressed genes |
|
|---|
|
|
|
|
|---|
| Term | Upregulated in
IPT | Downregulated in
IPT | Fold enrichment |
|---|
| Regulation of
lymphocyte activation | CD24, LAG3 | INHBA, NCK2, CDKN1A,
TNFRSF4, SHH | 3.677229 |
| Regulation of
lymphocyte proliferation | CD24 | NCK2, CDKN1A,
TNFRSF4, SHH | 4.683562 |
| Regulation of B cell
activation | CD24 | INHBA, CDKN1A,
TNFRSF4 | 6.097814 |
| Negative regulation
of lymphocyte activation | CD24, LAG3 | INHBA, SHH | 5.759046 |
| Regulation of B cell
proliferation | CD24 | CDKN1A, TNFRSF4 | 8.330049 |
| Antigen processing
and presentation of peptide or polysaccharide antigen via MHC class
II | HLA-DPB1 | IFI30, HLA-DRB5 | 7.067921 |
| Negative regulation
of immune system process | CD24, LAG3 | INHBA, SHH, | 3.746849 |
| Table II.Differentially expressed genes in DPCs
from mature and immature permanent teeth were enriched into
inflammation-associated subgroups based on ‘Biological Process’
terms analyzed using the Database for Annotation, Visualization and
Integrated Discovery. |
Table II.
Differentially expressed genes in DPCs
from mature and immature permanent teeth were enriched into
inflammation-associated subgroups based on ‘Biological Process’
terms analyzed using the Database for Annotation, Visualization and
Integrated Discovery.
|
| Differentially
expressed genes |
|
|---|
|
|
|
|
|---|
| Term | Upregulated in
IPT | Downregulated in
IPT | Fold enrichment |
|---|
| Leukocyte
migration | CD24 | AZU1, PODXL,
ITGAM | 6.819923 |
| Regulation of
leukocyte activation | CD24, LAG3 | INHBA, TNFRSF4, SHH,
NCK2, CDKN1A | 3.278493 |
| Regulation of
mononuclear cell proliferation | CD24 | NCK2, CDKN1A,
TNFRSF4, SHH | 4.627805 |
| Regulation of
leukocyte proliferation | CD24 | NCK2, CDKN1A,
TNFRSF4, SHH | 4.627805 |
| Positive regulation
of leukocyte proliferation | CD24 | NCK2, CDKN1A,
TNFRSF4 | 5.553366 |
| Negative regulation
of leukocyte activation | CD24, LAG3 | INHBA, SHH, AZU1,
ITGAM | 5.455939 |
| Leukocyte
chemotaxis |
|
| 6.303821 |
| Defense response | F11R, TRPV1, CD24,
AOC3 | C3AR1, DEFB4A,
TNFRSF4, AZU1, INHBA, APOL1 | 1.643435 |
| Table III.Differentially expressed genes in
dental pulp cells from mature and immature permanent teeth were
enriched into cell signaling-associated subgroups based on
‘Biological Process’ terms analyzed by the Database for Annotation,
Visualization and Integrated Discovery. |
Table III.
Differentially expressed genes in
dental pulp cells from mature and immature permanent teeth were
enriched into cell signaling-associated subgroups based on
‘Biological Process’ terms analyzed by the Database for Annotation,
Visualization and Integrated Discovery.
|
| Differentially
expressed genes |
|
|---|
|
|
|
|
|---|
| Term | Upregulated in
IPT | Downregulated in
IPT | Fold enrichment |
|---|
| Cell-cell
signaling | CD24 | WNT5A, GRM5, SHH,
GRM4, INHBA, NMUR2, SLC1A1, CLN8, LTA, CACNA1B | 2.202835 |
| Taxis |
| AZU1, C3AR1, DEFB4A,
ITGAM | 3.887356 |
| Chemotaxis |
| AZU1, C3AR1, CMKLR1,
DEFB4A, ITGAM, SLIT2 | 3.887356 |
| Cell surface recept
or linked signal transduction | GLI1, ADAM32, KLRF1,
CD24, AXIN2, LAG3 | WNT5A, C3AR1, OR5H1,
TAS2R5, TACR2, OR11A1, DEFB4A, OR8U1, SHH, ITGAM, AZU1, NMUR2,
CMKLR1, GPR78, KCNK2, GRM5, OR6M1, INHBA, NCK2, GPR32, GRM4,
OR52K2, KNG1 | 1.549916 |
| G-protein coupled
receptor protein signaling pathway | GPRC5C | KNG1, C3AR1, OR5H1,
NXPH4, TAS2R5, CMKLR1, TACR2, GPR78, OR11A1, NPR3, DEFB4A, OR8U1,
KCNK2, OR6M1, AZU1, GRM5, GRM4, GPR32, PTGIR, OR52K2, NMUR2,
OR1F2P | 1.592328 |
| Metal ion
transport | TRPV1, KCNK12 | NMUR2, KCNK2, CALHM1,
SLC17A3, SLC17A4, CAMK2D, SLC39A5, KCNQ2, CACNA1B | 2.006377 |
Genetic characteristics of SHH
The OMIM database analysis revealed that SHH is
expressed in cellular projections, extracellular regions and plasma
membranes and is associated with biological processes, including
cell proliferation, cell differentiation and immune system
processes. It was also demonstrated that, in the craniofacial
region, SHH mutation could lead to a number of deformities in mice
(Table IV) or humans (Table V), including abnormal maxillary
morphology, abnormal incisor morphology, tooth development arrest
and single median maxillary incisor. These findings further confirm
that SHH serves a crucial role in the development of teeth and may
also cause DPCs from mature and immature permanent teeth to exhibit
different biological characteristics.
| Table IV.Craniofacial phenotypes in an SHH
mutant mouse model. |
Table IV.
Craniofacial phenotypes in an SHH
mutant mouse model.
| Mouse genotype | Mouse phenotypes |
|---|
|
ShhDsh/ShhDsh | AM |
|
Shhtm1Chg/Shhtm1Chg | ACM, ACBM,
ANSM |
|
Shhtm3(Cre)Chg/Shhtm3(Cre)Chg | ACM |
|
ShhDsh/Shh+ | ACMM, ANM, AFM,
AIM, OBM, AMM |
|
ShhHx/Shh+ | CHP |
|
Shhtm1Amc/Shhtm2Amc |
|
| Tg(KRT14-Cre)1Amc/0
(conditional) | ACMM, AAP, AICM,
AMCM, SM, AABM, ATD, GRI, GRM, ADM, AEM, ANBM, APSF, PSH, CSP |
|
Shhtm1Amc/Shhtm2Amc |
|
|
Tg(Thy1-Cre)703Vaw/? (conditional) | ACM |
|
Shhtm2Chg/Shh+ |
|
| Tg(Sox2-Cre)1Amc/0
(conditional) | ANM, AMSM, PSFMM,
CSP |
| Table V.Craniofacial phenotypes in patients
with SHH-associated diseases. |
Table V.
Craniofacial phenotypes in patients
with SHH-associated diseases.
| Diseases associated
with SHH | Human
phenotypes |
|---|
| Holoprosencephaly
3 | MF, MR, AN, PB, CC,
HL |
| Microphthalmia,
isolated, with coloboma 5 | OC, AP, MP, BMP,
IFC |
| Solitary median
maxillary central incisor | CC, HL, AP, MP, SP,
TULV, CUL, CP, TP, PMPP, SMMI, |
|
| ARN, SN, CA, NNM,
MS, ANB, AS, CB, ICB, MP |
Functional protein association
networks of SHH
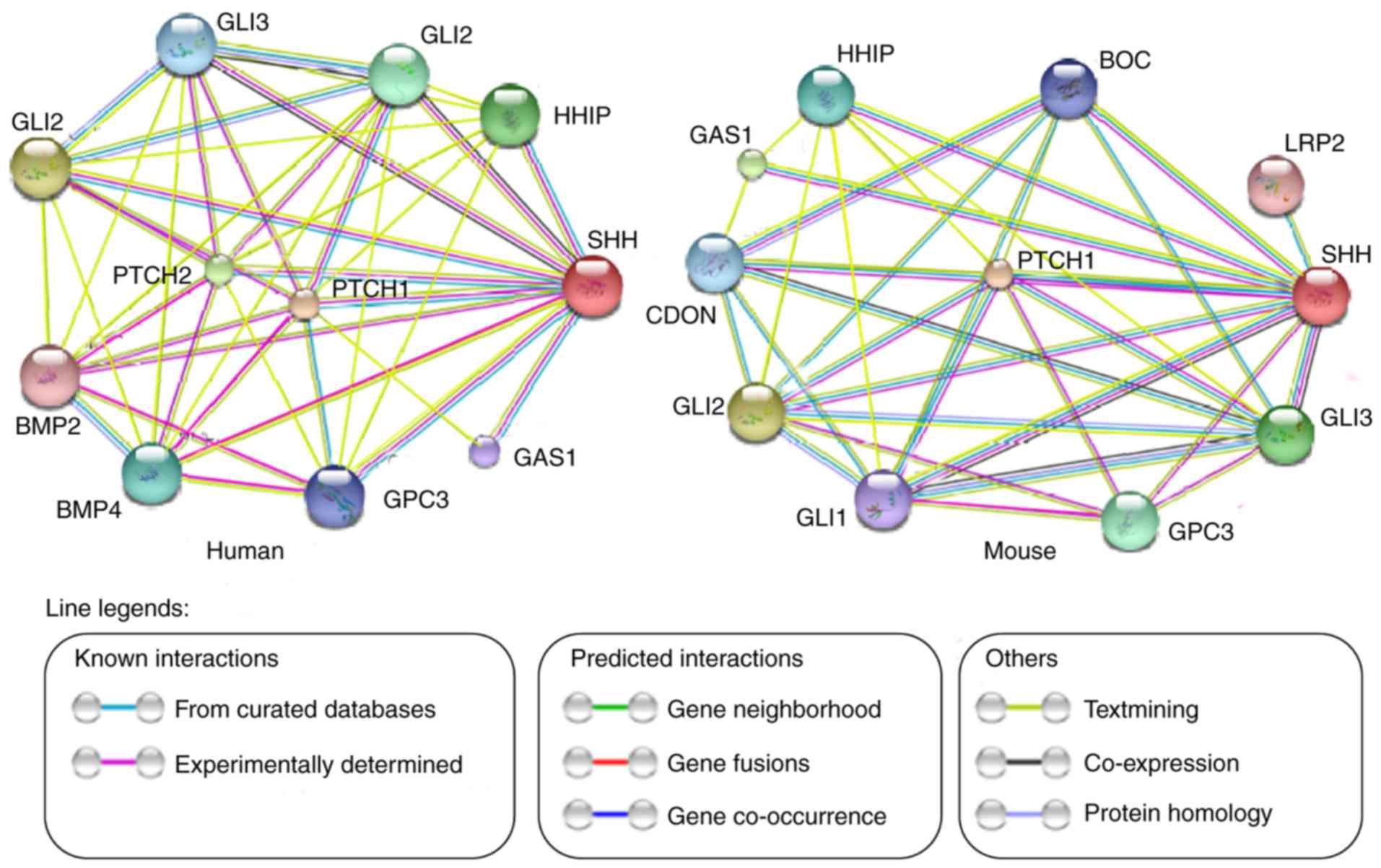
Based on STRING analysis, it was demonstrated that
SHH has functional protein associations with a number of other
proteins in mice and humans, including glioma-associated oncogene
(GLI)1, GLI2, growth arrest-specific protein 1 (GAS1), bone
morphogenetic protein (BMP)2 and BMP4 (Fig. 2). These findings indicate that SHH
may interact with other genes to regulate the biological
characteristics of DPCs.
Discussion
Dental pulp tissues may be derived from migrating
cranial neural crest cells during development; these tissues harbor
various populations of multipotential stem cells and are useful
cell resource for dental tissues, bone and neuron regeneration
(1,11,12).
DPSCs may be obtained from deciduous, inflamed deciduous, immature
permanent or mature permanent teeth and the cells from each source
may exhibit different biological characteristics (13). A previous study demonstrated that
inflamed human dental pulp retains tissue regeneration potential
(14). However, it has been
suggested that inflamed DPSCs have reduced stemness and
immunomodulatory properties and the expression of a number of
molecules is different compared with non-inflamed DPSCs (15). This implies that inflamed DPSCs may
not be a viable therapeutic cell source in cases of graft versus
host disease and organ rejection (15). Previous findings have also
demonstrated that DPSCs from deciduous teeth exhibit an
upregulation in genes that participate in pathways associated with
cell proliferation and the extracellular matrix including,
fibroblast growth factor and tumor growth factor β (16–18).
Although it has previously been reported that DPSCs from mature and
immature teeth have the potential for multi-directional
differentiation, the molecular and biological differences between
DPSCs from mature and immature permanent teeth has not been fully
investigated. The results of the present study demonstrate that a
number of differentially expressed genes in DPCs from mature and
immature permanent teeth are associated with immunity, inflammation
and cell signaling; furthermore, the findings herein suggest that
immune features, response to infection and signaling may be
different in DPCs from mature and immature permanent teeth. The
differential expression patterns of genes also indicate that DPCs
from immature permanent teeth may be more suitable for tissue
engineering or stem cell therapy. In the present study, only one
gene exhibited a statistically significant difference for the
adjusted P (adj.P) value (adj.P<0.05), suggesting that there may
be some level of error. Therefore, the differentially expressed
genes with a high |logFC| value should be focused on, and
quantitative polymerase chain reaction (qPCR) should be performed
to confirm differential expression prior to starting a functional
study.
SHH, a mammalian homologue of the Drosophila
secreted morphogen hedgehog, serves crucial role in the development
of many tissues (19). Previous
studies have demonstrated that SHH is expressed and serves key
roles during all stages of tooth development, including tooth
initiation, patterning and eruption (20–22). SHH
mutation results in retarded grown, teeth malformation and
mispatterned teeth in mice. The results of the present study
demonstrate that SHH, which is differentially expressed in DPCs
from mature and immature permanent teeth, is associated with many
subgroups based on biological process GO terms. OMIM database
analysis indicated that SHH serves a crucial role in the
development of craniofacial tissues, suggesting that the
differential expression of SHH may be responsible for the different
biological characteristics of DPCs from mature and immature
permanent teeth. Previous studies have also reported that SHH
protein could bind to Patched to release Smoothened to activate the
downstream signaling cascade (23).
GLI1-3, Huntington-interacting protein and GAS1 serve crucial roles
in regulating the SHH pathway (24–26). In
the present study, STRING database analysis revealed that SHH has
functional protein associations with a number of other proteins,
including GLI1, GLI2, GAS1, BMP2 and BMP4 in mice and humans. This
suggests that SHH may interact with other genes to regulate the
biological characteristics of DPCs.
In conclusion, 500 differentially expressed genes
were identified in DPCs from mature and immature permanent teeth
using the GEO online database. Based on bioinformatics analysis
using the DAVID database, these genes were enriched into a number
of subgroups associated with immune features, inflammation and cell
signaling. These findings suggest that immune features, response to
infection and signaling pathways may differ between DPCs from
mature and immature permanent teeth and that DPCs from immature
permanent teeth may be more suitable for tissue engineering or stem
cell therapy. OMIM database analysis demonstrated that SHH serves
crucial role in the development of craniofacial tissues, including
teeth, and may be responsible for the biological differences in
DPCs from mature and immature permanent teeth. The results of the
present study may provide a basis for future functional
experiments, including selecting suitable DPSCs for use in tissue
engineering and stem cell therapy, as well as selecting
intervention molecules to optimize the features of these tissues.
qPCR may be required to confirm the differential expression prior
to the launch of a functional study.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Gronthos S, Brahim J, Li W, Fisher LW,
Cherman N, Boyde A, DenBesten P, Robey PG and Shi S: Stem cell
properties of human dental pulp stem cells. J Dent Res. 81:531–535.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Waddington RJ, Youde SJ, Lee CP and Sloan
AJ: Isolation of distinct progenitor stem cell populations from
dental pulp. Cells Tissues Organs. 189:268–274. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Gandia C, Armiñan A, García-Verdugo JM,
Lledó E, Ruiz A, Miñana MD, Sanchez-Torrijos J, Payá R, Mirabet V,
Carbonell-Uberos F, et al: Human dental pulp stem cells improve
left ventricular function, induce angiogenesis, and reduce infarct
size in rats with acute myocardial infarction. Stem Cells.
26:638–645. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Mead B, Logan A, Berry M, Leadbeater W and
Scheven BA: Dental pulp stem cells, a paracrine-mediated therapy
for the retina. Neural Regen Res. 9:577–578. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Nicola FC, Rodrigues LP, Crestani T,
Quintiliano K, Sanches EF, Willborn S, Aristimunha D, Boisserand L,
Pranke P and Netto CA: Human dental pulp stem cells transplantation
combined with treadmill training in rats after traumatic spinal
cord injury. Braz J Med Biol Res. 49:e53192016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Majumdar D, Kanafi M, Bhonde R, Gupta P
and Datta I: differential neuronal plasticity of dental pulp stem
cells from exfoliated deciduous and permanent teeth towards
dopaminergic neurons. J Cell Physiol. 231:2048–2063. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Tamaoki N, Takahashi K, Aoki H, Iida K,
Kawaguchi T, Hatakeyama D, Inden M, Chosa N, Ishisaki A, Kunisada
T, et al: The homeobox gene DLX4 promotes generation of human
induced pluripotent stem cells. Sci Rep. 4:72832014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wu S, Wu F and Jiang Z: Identification of
hub genes, key miRNAs and potential molecular mechanisms of
colorectal cancer. Oncol Rep. 38:2043–2050. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Huang da W, Sherman BT and Lempicki RA:
Systematic and integrative analysis of large gene lists using DAVID
bioinformatics resources. Nat Protoc. 4:44–57. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Szklarczyk D, Franceschini A, Wyder S,
Forslund K, Heller D, Huerta-Cepas J, Simonovic M, Roth A, Santos
A, Tsafou KP, et al: STRING v10: Protein-protein interaction
networks, integrated over the tree of life. Nucleic Acids Res.
43(Database issue): D447–D452. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Chang CC, Chang KC, Tsai SJ, Chang HH and
Lin CP: Neurogenic differentiation of dental pulp stem cells to
neuron-like cells in dopaminergic and motor neuronal inductive
media. J Formos Med Assoc. 113:956–965. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Bray AF, Cevallos RR, Gazarian K and Lamas
M: Human dental pulp stem cells respond to cues from the rat retina
and differentiate to express the retinal neuronal marker rhodopsin.
Neuroscience. 280:142–155. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Rosa V, Dubey N, Islam I, Min KS and Nor
JE: Pluripotency of stem cells from human exfoliated deciduous
teeth for tissue engineering. Stem Cells Int. 2016:59578062016.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Alongi DJ, Yamaza T, Song Y, Fouad AF,
Romberg EE, Shi S, Tuan RS and Huang GT: Stem/progenitor cells from
inflamed human dental pulp retain tissue regeneration potential.
Regen Med. 5:617–631. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yazid FB, Gnanasegaran N, Kunasekaran W,
Govindasamy V and Musa S: Comparison of immunodulatory properties
of dental pulp stem cells derived from healthy and inflamed teeth.
Clin Oral Investig. 18:2103–2112. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Nakamura S, Yamada Y, Katagiri W, Sugito
T, Ito K and Ueda M: Stem cell proliferation pathways comparison
between human exfoliated deciduous teeth and dental pulp stem cells
by gene expression profile from promising dental pulp. J of Endod.
35:1536–1542. 2009. View Article : Google Scholar
|
|
17
|
Wang X, Sha XJ, Li GH, Yang FS, Ji K, Wen
LY, Liu SY, Chen L, Ding Y and Xuan K: Comparative characterization
of stem cells from human exfoliated deciduous teeth and dental pulp
stem cells. Arch Oral Biol. 57:1231–1240. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Volponi AA, Pang Y and Sharpe PT: Stem
cell-based biological tooth repair and regeneration. Trends Cell
Biol. 20:715–722. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Varjosalo M and Taipale J: Hedgehog:
Functions and mechanisms. Genes Dev. 22:2454–2472. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Li J, Feng J, Liu Y, Ho TV, Grimes W, Ho
HA, Park S, Wang S and Chai Y: BMP-SHH signaling network controls
epithelial stem cell fate via regulation of its niche in the
developing tooth. Dev Cell. 33:125–135. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Hu X, Zhang S, Chen G, Lin C, Huang Z,
Chen Y and Zhang Y: Expression of SHH signaling molecules in the
developing human primary dentition. BMC Dev Biol. 13:112013.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kumamoto H, Ohki K and Ooya K: Expression
of Sonic hedgehog (SHH) signaling molecules in ameloblastomas. J
Oral Pathol Med. 33:185–190. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Villavicencio EH, Walterhouse DO and
Iannaccone PM: The sonic hedgehog-patched-gli pathway in human
development and disease. Am J Hum Genet. 67:1047–1054. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Bai CB, Auerbach W, Lee JS, Stephen D and
Joyner AL: Gli2, but not Gli1, is required for initial Shh
signaling and ectopic activation of the Shh pathway. Development.
129:4753–4761. 2002.PubMed/NCBI
|
|
25
|
Gan H, Liu H, Zhang H, Li Y, Xu X, Xu X
and Xu J: SHh-Gli1 signaling pathway promotes cell survival by
mediating baculoviral IAP repeat-containing 3 (BIRC3) gene in
pancreatic cancer cells. Tumour Biol. 37:9943–9950. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Allen BL, Tenzen T and McMahon AP: The
Hedgehog-binding proteins Gas1 and Cdo cooperate to positively
regulate Shh signaling during mouse development. Genes Dev.
21:1244–1257. 2007. View Article : Google Scholar : PubMed/NCBI
|