Introduction
Alzheimer's disease (AD) is a neurodegenerative
disease clinically characterized by cognitive and intellectual
dysfunction; specific pathological hallmarks include the presence
of senile plaques, cerebral amyloid angiopathy and neurofibrillary
tangles (1). The number of people
living with dementia is currently estimated to be ~24 million and
its prevalence is forecast to double every 20 years until 2040
(2). The pathogenesis of AD is
complex and has not yet been elucidated, however the results of
numerous studies support the amyloid β protein (Aβ) hypothesis of
AD development (3–5).
Amyloid-β (Aβ) is hypothesized to trigger the onset
and development of AD. It also promotes oxidative stress,
neurotropic signaling, Tau phosphorylation and synaptic dysfunction
and activates inflammatory factors (6). The generation of Aβ is dependent on the
processing of amyloid precursor protein (APP), which involves two
signaling pathways via sequential limited proteolysis. In the
amyloidogenic pathway, APP is cleaved by β-secretase, generating a
C-terminal fragment (CTF)-β and soluble (s)APP-β (7). CTF-β is further cleaved by γ-secretase
to generate Aβ (8). Alternatively,
APP may be processed via the nonamyloidogenic pathway, which is
regulated by α- and γ-secretase. The cleavage of α-secretase
generates CTF-α and sAPP-α, and CTF-α is subsequently cleaved by
the γ-secretase complex to generate an APP intracellular domain and
p3 peptides, thus impeding Aβ production (7). It has been demonstrated that the
cleavage pathway of APP serves an important role in the formation
and accumulation of Aβ (9).
Aβ levels depend on APP processing, as well as on
its elimination via transport processes, cell-mediated clearance
and Aβ degradation (10). It has
been demonstrated that Aβ-degrading enzymes are required to degrade
Aβ (11). One such enzyme,
neprilysin (NEP), degrades monomeric Aβ and its oligomeric forms
Aβ40 and Aβ42 (12). Furthermore, insulin-degrading enzyme
(IDE) may be involved in Aβ degradation, as indicated by Kurochkin
and Goto (13). Therefore, NEP and
IDE are the most promising Aβ-degrading candidates.
Previous studies have focused on ways to decrease Aβ
production and reduce Aβ levels, which are the prime drug targets
for AD (14,15). Therefore, targeting Aβ metabolism may
be a potential method of treating AD. Currently, the development of
novel drugs for AD treatment has focused on ways of decreasing Aβ
levels; however the results of clinical trials have not been
promising (16,17). Western medicine is a single-target,
single-component and symptomatic treatment paradigm, while
traditional Chinese medicine (TCM) therapy is a multi-target,
multi-component and treatment strategy that has been applied to
treat various diseases. Tongluo Xingnao effervescent tablets (TXET)
are derived from Qionggui Tang [Ligusticum striatum DC. and
Angelica sinensis (Oliv.) Diels], which has been used in
Japan, China, South Korea and other East Asian countries for
>2,000 years, usually in combination with Scutellaria
baicalensis Georgi. In TCM, it is hypothesized that Qionggui
Tang may be used to treat patients with various conditions. The
addition of Scutellaria baicalensis Georgi may enhance the
effect of Qionggui Tang on the efflux of pathogenic toxins.
Therefore, the function of TXET involves promoting blood
circulation to remove toxins, as well as improving memory and
cognition (18–21). In a previous study by our group, TXET
improved cognition and synaptic function; it also improved cerebral
metabolism and mitochondrial function (18). Additionally, it was demonstrated that
TXET may downregulate levels of cyclin-dependent kinase (CDK)5 and
glycogen synthase kinase (GSK)-3, key proteins that regulate tau
protein phosphorylation, thus inhibiting tau phosphorylation in an
animal model of AD (19).
Furthermore, TXET may increase acetylcholine (ACh) and choline
acetyltransferase (ChAT) levels, thus enhancing the central
cholinergic neuronal system in the hippocampi of rats with AD
(20). Another study by our group
demonstrated that TXET increases the expression of IDE, indicating
that TXET may regulate the metabolism of Aβ to treat AD (21). In the present study, the anti-AD
effect of TXET on the production and elimination of Aβ was
investigated in AD transgenic mice.
Materials and methods
Reagents
TXET, based on the traditional Chinese formula
Qionggui Tang [Ligusticum striatum DC and Angelica
sinensis (Oliv.) Diels] plus Scutellaria baicalensis
Georgi, is a patented prescription (Patent no. ZL
201310132096.2) (22) owned by the
Affiliated Hospital of Chengdu University of Traditional Chinese
Medicine (Chengdu, China) and used in the long-term treatment of
dementia. Ligusticum striatum DC. (1,250 g) and Angelica
sinensis (Oliv.) Diels (250 g) were extracted via supercritical
carbon dioxide fluid extraction to collect residues and volatile
oils. Ethanol was added to dissolve the volatile oils and the
preparation was mixed with β-cyclodextrin dissolved in purified
water (1:10) at 60°C. The preparation contained these inclusion
complexes and when the preparation was fully mixed, it was dried
and the residue was crushed into a fine powder. Scutellaria
baicalensis Georgi (750 g) and residues were extracted with 60%
ethanol by heating under reflux 3 times, followed by filtration.
The filtrate was concentrated to extractum such that the relative
density was between 1.1 and 1.2; the extractum was dried using a
vacuum dryer and made into grain. The inclusion complexes and the
grain were blended with corresponding excipients and compressed
into tablets, with each tablet weighing 1.2 g (active ingredient,
0.6 g). TXET was provided by the Affiliated Hospital of Chengdu
University of Traditional Chinese Medicine (Chengdu, China) and its
quality analysis was performed by liquid chromatography (LC)
assays, as previously described (23). The LC assay was performed using an
LC-30A (Shimadzu Corporation, Kyoto, Japan) liquid chromatography
system, with a LabSolutions 5.41.20 chromatography workstation
(Shimadzu Corporation) and an Agilent Poroshell 120 EC-C 18 column
(2.1×100 mm, 2.7 µm; Agilent Technologies Inc., Santa Clara, CA,
USA). The results demonstrated that peaks of ferulic acid (18.5
min), baicalin (25.2 min), baicalein (33.4 min) and wogonin (48.9
min) appeared successively (data not shown), which indicated that
the quality of TXET was high.
Aricept (cat. no. 100526A) was purchased from Eisai
China Inc. (Eisai Co., Ltd., Tokyo, Japan) Human Aβ1-40 (cat. no.
KHB3481) and Aβ1-42 ELISA kits (cat. no. KHB3441) were purchased
from Invitrogen (Thermo Fisher Scientific, Inc., Waltham, MA, USA).
Aβ monoclonal antibodies (cat. no. 803015) were purchased from
BioLegend, Inc. (San Diego, CA, USA). The DAB Kit for
Immunohistochemistry was purchased from ZG ZSGB-Bio, Inc. (Beijing,
China). APP (cat. no. 11090), CTF-α (cat. no. 11088) and CTF-β
(C99; cat. no. 10321) antibodies were purchased from
Immuno-Biological Laboratories Co., Ltd, (Gunma, Japan). IDE (cat.
no. ab32216) and NEP (cat. no. ab81688) antibodies were purchased
from Abcam (Cambridge, UK). The presenilin-1 (PS1) antibody (cat.
no. 5643) was purchased from Cell Signaling Technology, Inc.
(Danvers, MA, USA). The β-actin antibody (cat. no. sc-47778), goat
anti-rabbit IgG-HRP (cat. no. sc-2004) and goat anti-mouse IgG-HRP
(cat. no. sc-2005) were provided by Santa Cruz Biotechnology, Inc.
(Dallas, TX, USA). The γ-secretase activity kit (cat. no. FP003)
was purchased from R&D Systems, Inc. (Minneapolis, MN, USA) The
BCA protein assay kit and radioimmunoprecipitation assay (RIPA)
lysis buffer were purchased from Applygen Technologies, Inc.
(Beijing, China).
Animals and administration
procedure
A total of 60 3-month-old male APPswe/PS1dE9 mice
and 15 age-matched C57BL/6J mice (20±2 g) were purchased from
Beijing HFK Bioscience Co., Ltd. (Beijing, China; Certificate SCXK
Jing 2009–0004;). In the animal observation room with a 12 h
light/dark cycle, animals had ad libitum access to food and
water and were housed under a suitable temperature (22±2°C) and
humidity (65±5%). The treatment and care of animals adhered to the
international laws and regulations indicated in the National
Institutes of Health Guide for the Care and Use of Laboratory
Animals (24). All animal protocols
were approved by the Institutional Animal Care and Use Committee of
Dongfang Hospital affiliated with the Beijing University of Chinese
Medicine (Beijing, China).
The APPswe/PS1dE9 mice were randomly divided into
four groups (each n=5): A model group, an Aricept (1 mg/kg) group
(positive control), a TXET low-dose (0.45 g/kg) group and a TXET
high-dose (1.8 g/kg) group. C57BL/6J mice served as the normal
control group, which alongside the model group was treated with an
equal volume of sterile saline. A total of 10 ml/kg/day solution
was administered to all mice intragastrically for 180 days.
Morris water maze (MWM) test
All mice underwent the MWM test, which evaluates
spatial learning and memory performance, on day 176 of
administration. For the place navigation task, mice trained for 5
days consecutively, while the escape latency and total swimming
distance were recorded in one 120 sec session. The platform was
then removed and the probe trial was conducted to record the
distance each mouse crossed in the target area (where the platform
was placed formerly) and the opposite area. All tests were
performed at night.
Brain tissue preparation
For immunochemistry, three mice in each group were
selected randomly and sacrificed under ether anesthesia by
decapitation. Brains were harvested and fixed with 4%
paraformaldehyde at 4°C overnight using perfusion fixation,
followed by paraffin-embedding. Other mice (34±4 g) were sacrificed
under ether anesthesia by decapitation; the brain tissue was
immediately dissected, snap-frozen in liquid nitrogen and stored at
−80°C until processing.
Immunohistochemistry
Paraffin-embedded tissue was serially cut into
4-µm-thick sections. Sections were rinsed with 0.01 mol/l PBS
following dewaxing in xylene, rehydrated through decreasing
concentrations of ethanol and washed in 0.01 mol/l PBS. For
Aβ-containing plaque immunostaining, antigens were unmasked by
microwaving sections in 10 mmol/l citrate buffer, (pH 6.0) for 15
min and allowed to cool. Sections were then blocked with 5% goat
serum (cat. no. ZLI-9022; OriGene Technologies, Inc., Beijing,
China) for 1 h at room temperature, sections were treated with 3%
H2O2 for 10 min to eliminate endogenous
peroxidase activity. Following rinsing in PBS, sections were
incubated with anti-Aβ monoclonal antibodies (1:200) overnight at
4°C. Following rinsing with PBS and the addition of Polymer Helper
(polink-2 plus polymer HRP detection system; OriGene Technologies,
Inc.; cat. no. PV-9001) incubated at 37°C for 10 min), the sections
were incubated with horseradish peroxidase conjugated Goat
anti-Mouse IgG-Fragment crystallizable (1:500, cat. no. ab20043;
Abcam) at 37°C for 1 h and stained with 3,3-diaminobenzidine of DAB
kit (cat. no. ZLI-9018; OriGene Technologies, Inc.) for 6 min and
hematoxylin for 10 min at room temperature. Following dehydration,
sections were covered with neutral balsam. Using an inverted
fluorescence microscope (IX71; Olympus Corporation, Tokyo, Japan;
magnification ×200), images of all sections of each group were
captured and the number of Aβ plaques in the hippocampi were
determined using Image-Pro Plus software (version 6.0; Media
Cybernetics, Inc., Rockville, MD, USA).
ELISA
Human Aβ1–40 and Aβ1–42 ELISA
kits were used to detect the concentration of Aβ in the mouse
brain. The brain tissue was homogenized and centrifuged at 5,000 ×
g at 4°C for 15 min and the supernatant was used to detect Aβ
according to the manufacturer's protocol of the kits.
Western blot analysis
The expression of APP, CTF-α, CTF-β, PS1, NEP and
IDE proteins were measured by western blot analysis. Brain tissues
were homogenized in RIPA lysis buffer and the homogenate was
centrifuged at 4°C and 15,000 × g for 20 min. The protein
concentration in the supernatant was determined using the BCA kit,
following the manufacturer's protocol. A total of 40 µg protein was
separated using 10% SDS-PAGE and transferred to a polyvinylidene
difluoride membrane (EMD Millipore, Billerica, MA, USA). The
membrane was blocked using 5% skimmed milk (cat. no. 1172GR100;
BioFroxx GmbH, Einhausen, Germany) in TBS-T at room temperature for
1 h and incubated with primary antibodies (APP, PS1, NEP and IDE
were diluted by 1:1,000; CTF-α and CTF-β were diluted by 1:500) at
4°C overnight, and β-actin (1:1,000) was used as loading control.
The membrane was then incubated with goat anti-mouse horseradish
peroxidase conjugated (HRP) IgG (1:2,000) and goat anti-rabbit
IgG-HRP (1:5,000) at room temperature for 2 h and visualized using
Super ECL Plus enhanced chemiluminescent kit (cat. no. P1010;
Applygen Technologies Inc.). Membranes were then exposed to the
GeneGnomeXRQ bio imaging system (Syngene, Frederick, MD, USA) and
the images were quantified using Image J software version 1.46r
(National Institutes of Health, Bethesda, MD, USA).
Secretase activity
The γ-secretase activity kit was used to detect the
activity of γ-secretase in mouse brain tissues. The brain tissue
was weighed, homogenized in a RIPA buffer and then centrifuged at
15,000 × g and 4°C for 20 min. The supernatant was assessed
following the manufacturer's protocol.
Statistical analysis
All data were analyzed by SPSS software (version
13.0; SPSS, Inc., Chicago, IL, USA) and the results are presented
as the mean ± standard deviation. For comparisons among groups,
one-way analysis of variance (ANOVA) was used. P<0.05 indicated
that the difference between groups was statistically
significant.
Results
TXET improves the learning and memory
dysfunction of APPswe/PS1dE9 mice
APPswe/PS1dE9 mice exhibit cognitive damage and
behavioral changes at 12 weeks old (25). The learning and memory abilities of
mice were examined by the MWM. The results of the place navigation
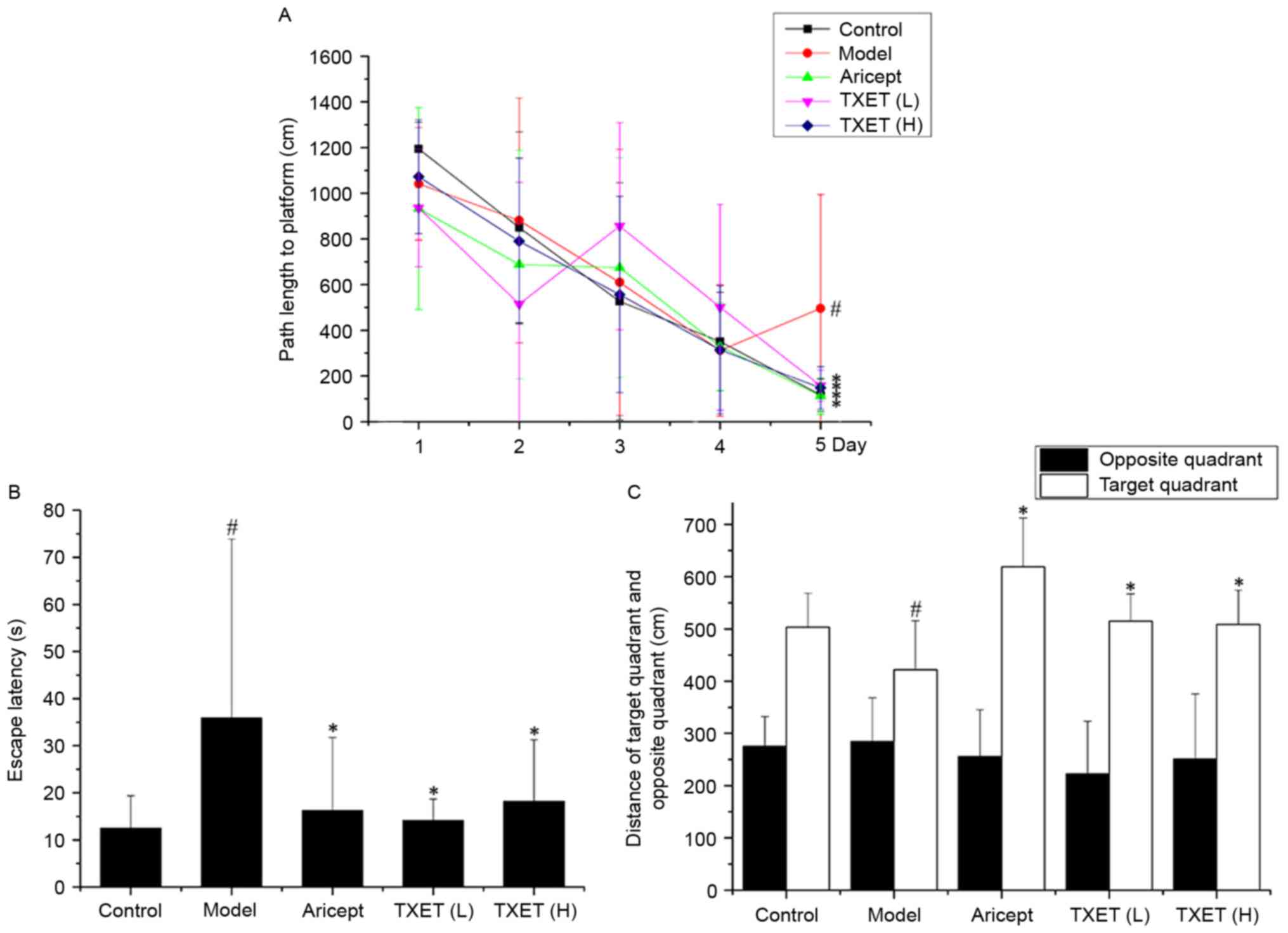
task are presented in Fig. 1. The
average path length during the training days gradually declined.
Compared with the control group, the path length and escape latency
were significantly lengthened in the model group on day 5
(P<0.05; Fig. 1A and B). By
contrast, the TXET high-dose, TXET low-dose and Aricept groups all
exhibited significantly decreased escape latency and path length on
day 5 compared with the model group (all P<0.05). The results of
the spatial probe test identified a significant reduction in the
distance of the target quadrant between the model and control
groups (Fig. 1C; P<0.05).
Compared with the model group, the TXET high-dose, TXET low-dose
and Aricept groups significantly increased the distance of the
target area (Fig. 1C; P<0.05).
These results indicate that TXET may improve learning and attenuate
memory dysfunction.
TXET reduces the level and deposition
of Aβ in the hippocampi of APPswe/PS1dE9 mice
It has been demonstrated that APPswe/PS1dE9 mice
exhibit plaque formation in the brain at the age of 17 weeks
(25), making them a suitable model
for studying Aβ metabolism in AD. To evaluate the anti-AD effect of
TXET in Aβ metabolism, the effect of TXET on Aβ levels and
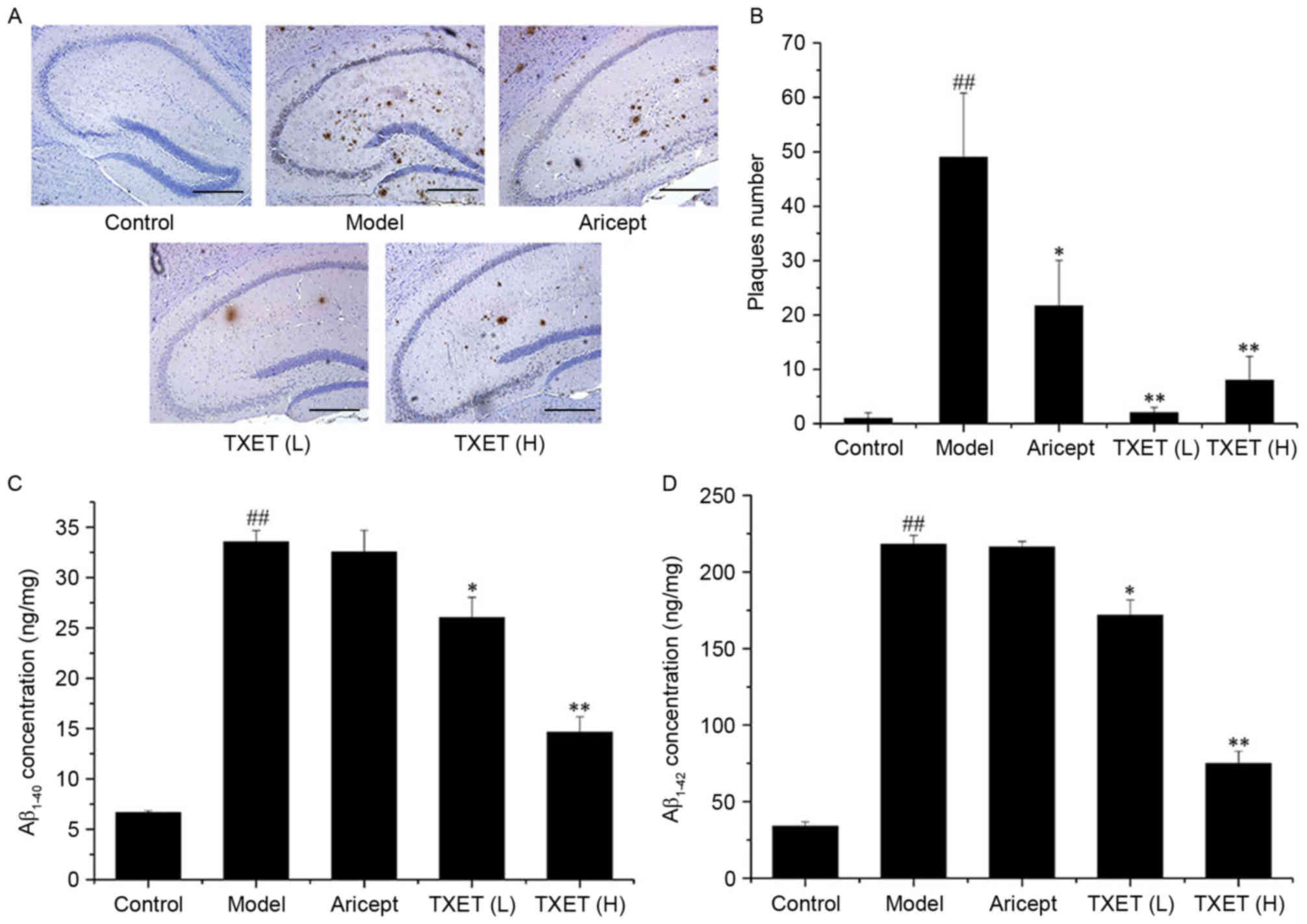
deposition was investigated. As presented in Fig. 2A, the number and size of the
Aβ-positive plaques decreased in the hippocampi of AD mice
following TXET administration. Statistical analysis revealed that
low- and high-dose TXET (P<0.01) and Aricept (P<0.05)
significantly reduced the number of Aβ-positive plaques compared
with the model group (Fig. 2B). TXET
treatment also reduced Aβ levels. Aβ levels in the hippocampus were
measured by ELISA, which demonstrated that levels of
Aβ1–40 were significantly lowered by 56.02% (P<0.01)
and 21.29% (P<0.05) following treatment with high- and low-dose
TXET, respectively, compared with the model group (Fig. 2C). Levels of Aβ1–42 were
also significantly reduced in the hippocampus by 66.14% (P<0.01)
and 21.62% (P<0.05) following treatment with high- and low-dose
TXET, respectively, compared with the model group (Fig. 2D). However, Aricept exhibited no
significant effect on Aβ concentration in the hippocampi of
APPswe/PS1dE9 mice. These results suggest that TXET reduces Aβ
levels and deposition.
TXET reduces Aβ by inhibiting
β-secretase in APPswe/PS1dE9 mice
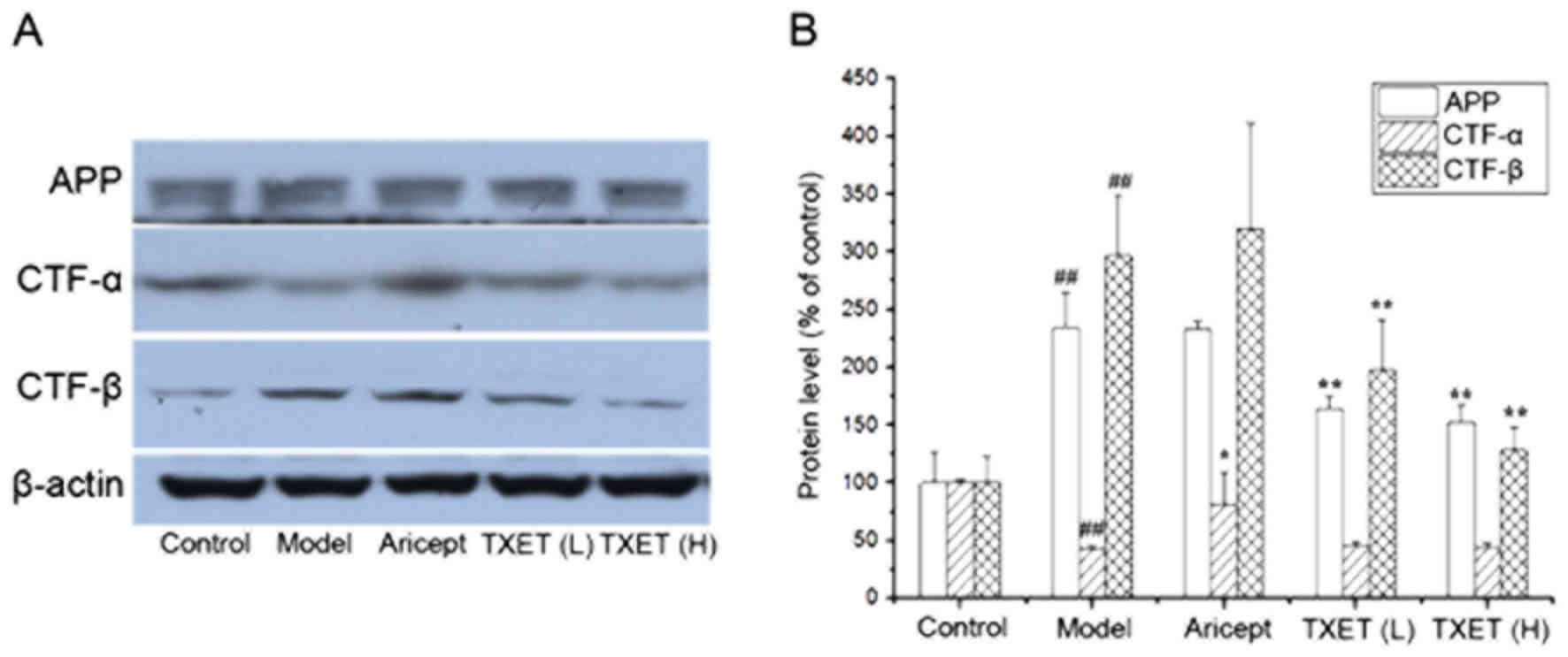
The cleavage pathway of APP serves a major role in
Aβ generation (9). The reduction of
Aβ production may be associated with the regulation of APP
processing; therefore, the effects of TXET on the expression of APP
were examined. It was demonstrated that the expression of APP was
significantly decreased in mice administered with low- and
high-dose TEXT, compared with the model group (P<0.01),
indicating that TXET reduces Aβ levels via the regulation of APP
processing. APP is cleaved by α- or β-secretase via two cleavage
pathways. Therefore, the protein levels of CTF-α and CTF-β cleaved,
respectively, by α- or β-secretase, were examined (Fig. 3). The data indicated that the
expression of CTF-α significantly increased following treatment
with Aricept compared with the model group (P<0.05), but no
significant differences were identified following TXET
administration. A significant reduction in the expression of CTF-β
was observed in the TXET-treated groups (P<0.01) compared with
the model group. There were no significant differences between the
expression of CTF-β in the model and Aricept groups. These data
suggest that TXET decreases Aβ production by regulating APP
processing, which inhibits β-secretase.
The Aβ reduction of TXET is irrelevant
to the cleavage of γ-secretase in APPswe/PS1dE9 mice
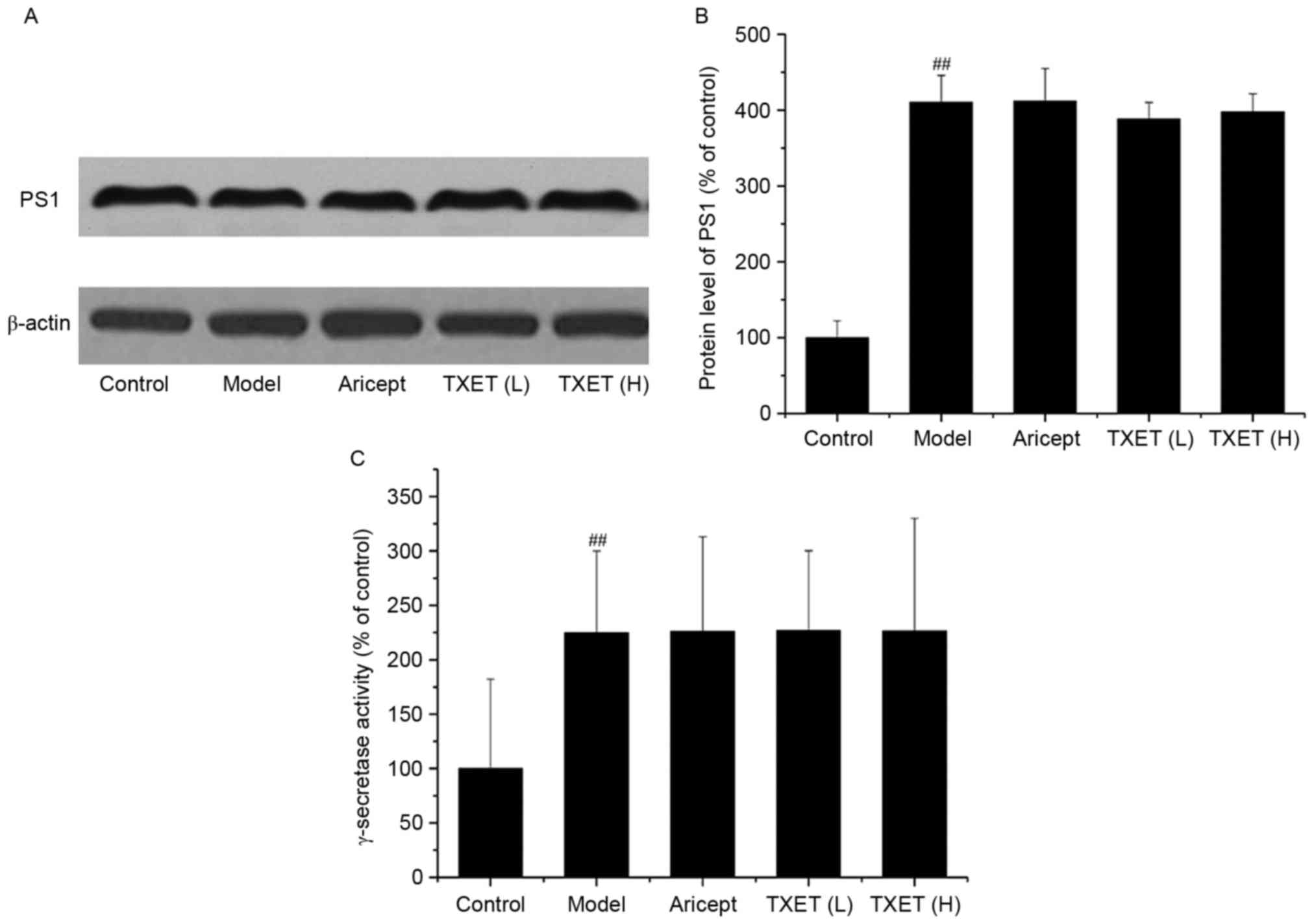
The involvement of γ-secretase is the final step in
APP processing and PS1 is a component of the γ-secretase complex
(26). Thus, the effect of TXET
treatment on the expression of PS1 and the activity of γ-secretase
was examined (Fig. 4). The results
demonstrated that TXET treatment did not significantly affect PS1
expression and γ-secretase activity. These data suggest that TXET
does not reduce Aβ levels in APPswe/PS1dE9 mice via the regulation
of γ-secretase.
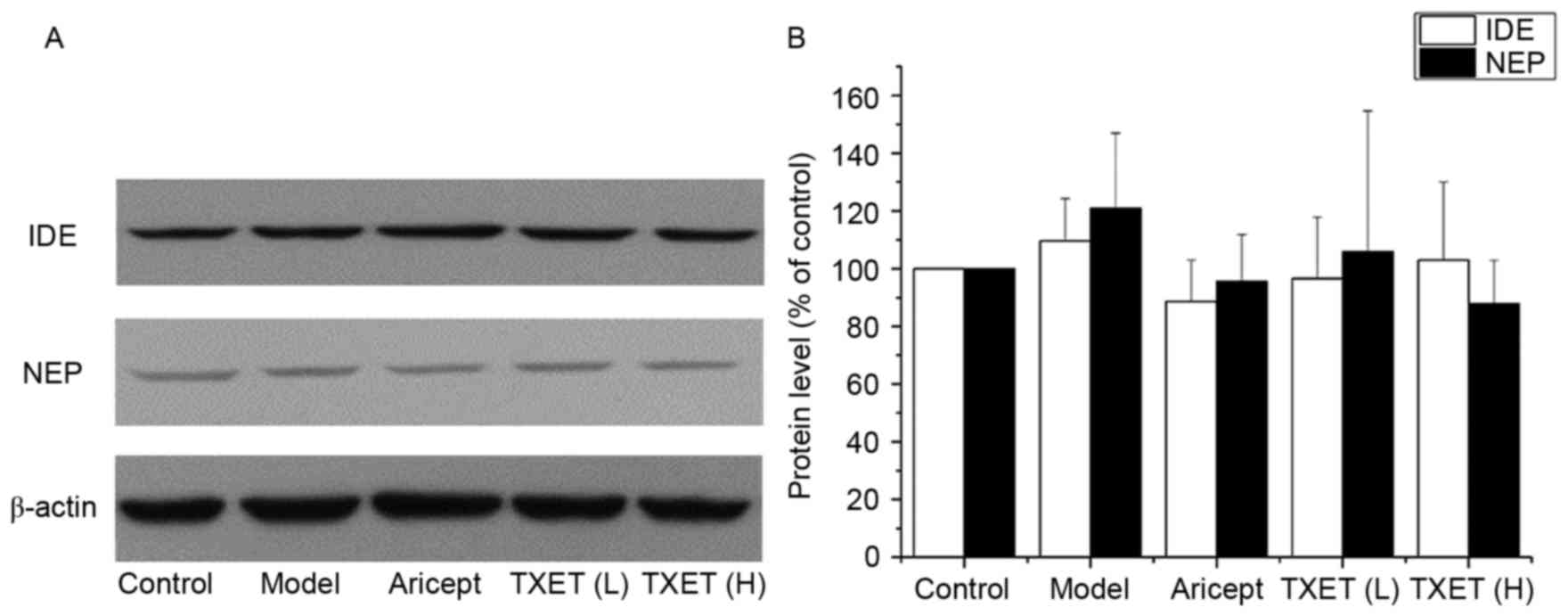
The reduced effect on Aβ of TXET is
not associated with NEP or IDE in APPswe/PS1dE9 mice
Aβ levels in the brain are dependent on APP
processing and Aβ degradation. NEP and IDE are the most important
Aβ-degrading enzymes (12,13). TXET decreased Aβ production by
inhibiting β-secretase; therefore, the association between
β-secretase and the Aβ degradation enzymes was investigated
(Fig. 5). The results demonstrated
that TXET treatment did not significantly affect the expression of
NEP and IDE compared with the model group.
Discussion
The primary pathological change that occurs during
AD is the excessive deposition of Aβ, caused by the decrease in
metabolism that occurs during the aging process (27). Aβ deposition is closely associated
with Aβ homeostasis (28). Under
normal physiological conditions, the generation and elimination of
Aβ maintains a dynamic balance. However, if Aβ homeostasis is
disrupted, Aβ may aggregate, resulting in the formation of
neurofibrillary tangles, cell death and promoting the onset and
development of AD. Therefore, decreasing Aβ production and
inhibiting Aβ aggregation may be developed as novel therapeutic
strategies to treat patients with AD.
Previous studies have indicated that reducing the
formation of Aβ plaques in the brain is the key to AD therapy, thus
verifying the Aβ hypothesis of AD (29,30).
Reducing Aβ levels depends on APP processing by β- and γ-secretase,
as well as Aβ elimination via transport processes, cell-mediated
clearance and Aβ degradation (31).
It has been reported that upregulating α-secretase and
downregulating β-secretase may decrease the generation of Aβ
(32). Furthermore, inhibition of
γ-secretase may reduce Aβ concentrations in the plasma and
cerebrospinal fluid of patients with AD (33). Previous in vitro and in
vivo studies have reported that altering catabolism may
decrease Aβ levels, including via NEP (34–36) and
IDE (37,38). However, there are currently no data
regarding the long-term safety and efficacy of Aβ inhibitors or
associated drugs. Thus, it is important to identify reliable,
effective and safe novel therapeutic strategies that may be used to
treat patients with AD.
Different TCM treatments have been widely used to
treat various diseases and are safe and effective. It has been
hypothesized that herbal medicine may improve cognitive dysfunction
to treat AD (39–41). TCM defines dementia as a disease of
abnormal consciousness, which is caused by mental dysfunction and
deterioration of the brain tissue. TXET, based on the traditional
Chinese formula Qionggui Tang plus Scutellaria
baicalensis Georgi, is a patented prescription that has been
used as a long-term treatment of patients with dementia at the
Affiliated Hospital of Chengdu University of Traditional Chinese
Medicine. It may be a reliable and effective method of treating AD
(18–22,41–44).
The results of previous studies have revealed that
TXET may improve dysfunction in learning and memory, improve
cerebral metabolism and mitochondrial function (18), downregulate levels of CKD5 and GSK-3
to inhibit tau phosphorylation and promote hippocampal
synaptophysin remodeling in AD rats (19,42).
Furthermore, TXET reduces ACh and ChAT to enhance the central
cholinergic neuronal system in the AD rat hippocampus (20). It may also regulate the p38 and the
mitogen-activated protein kinase signaling pathways in vascular
dementia (VD), the second most common cause of dementia following
AD (23). Additionally, it may
increase levels of cytochrome c oxidase in the hippocampus
in VD (43) and in an AD model
induced by chronic cerebral ischemia (44). Furthermore, TXET may increase the
expression of IDE in the hippocampus of AD rats (20), suggesting that TXET may act to treat
AD by altering the metabolism of Aβ.
Transgenic APP695sw/PS1dE9 mice overexpressing
APP695sw and PSEN1dE9 represent an ideal animal model to use to
study the pathogenesis of AD and evaluate novel experimental
therapeutics for AD (45).
APP695sw/PS1dE9 mice overproduce Aβ and exhibit cognitive damage
and behavioral changes at 12 weeks of age, In addition, plaque
formation occurs in their brains at the age of 17 weeks, which are
beneficial for identifying the association between TXET and Aβ
metabolism. A previous study indicated that TXET may be used to
treat AD by altering the metabolism of Aβ (20). The present study demonstrated that
APP695sw/PS1dE9 mice in the model group exhibited learning and
memory dysfunction, as well as the increased production of
Aβ1–40, Aβ1–42 and plaques in the brain.
However, TXET administration prolonged the escape latency, reduced
the average path length in training days and the distance of the
target area in mice undergoing MWM. Plaque formation, associated
with Aβ metabolism, was inhibited following administration in the
mice at 3-months-old (12 weeks) and TXET reduced the content of
Aβ1–40, Aβ1–42 and the expression of plaques
in the hippocampi of APP695sw/PS1dE9 mice. The result suggested
that TXET may attenuate learning and memory dysfunction in AD by
regulating Aβ metabolism in the brain.
Aricept is widely used to treat AD. It has been
reported that Aricept may reverse attention deficits in the 3xTgAD
mouse model, which exhibits Aβ plaques and neurofibrillary tangles
in the brain (46). In the present
study, it was indicated that Aricept improved learning and memory
deficits, but had little effect on the content of Aβ plaques or the
expression of CTF-β, NEP and IDE. However, it was also demonstrated
that Aricept reduced APP and increased CTF-α levels, indicating
that Aricept may regulate APP processing by activating α-secretase.
Aricept, an AChE inhibitor, is known to increase cholinergic
function in the cerebral cortex (47). However, the mechanism by which
α-secretase is regulated by Aricept requires further study.
It is hypothesized that Aβ accumulation serves the
most important role in the pathogenesis of AD; activating
inflammatory factors, as well as promoting oxidative stress, tau
phosphorylation and synaptic dysfunction (48). The results of the present study
demonstrated that TXET may reduce the content of Aβ1–40,
Aβ1–42 and the expression of plaques in the hippocampi
of APP695sw/PS1dE9 mice. However, it remains unclear if Aβ
generation or elimination mediates the Aβ-reducing effect of TXET.
The results of the current study indicate that TXET reduces the
expression of APP, suggesting that TXET may decrease Aβ levels via
APP processing. The data also demonstrated that TXET reduces CTF-β
but has no effect on CTF-α. This suggests that TXET may decrease Aβ
generation by inhibiting β-secretase but not α-secretase. PS1 is a
component of the γ-secretase complex, which is involved in the
final step of Aβ generation (26).
The current study demonstrated that the expression of PS1 and the
activity of γ-secretase were not significantly altered following
TXET administration, suggesting that Aβ reduction does not regulate
γ-secretase. Furthermore, there was no difference in the expression
of NEP and IDE following TXET administration in APP695sw/PS1dE9
mice, suggesting that the Aβ-reducing effect of TXET is not
associated with NEP and IDE. However, a previous study by our group
indicated that TXET increases the expression of IDE in the
hippocampus of AD rats (21), which
is inconsistent with the results of the present study. The two
models of AD used in the different studies were assessed and it was
identified that the methods of analysis differed between them. This
may have caused the results of these two studies to be
inconsistent; however, further analysis is required for
clarification. Overall, these results suggest that TXET may reduce
Aβ accumulation by downregulating β-secretase.
The results of the present study suggest that TXET
may ameliorate cognitive dysfunction and decrease Aβ levels, while
regulating Aβ metabolism by downregulating β-secretase. However,
further studies are required to elucidate its underlying mechanism
of action. The results have laid the foundation for the development
of TCM with an inhibitor of β-secretase as a target.
Acknowledgements
The authors would like to thank Dr Wang Ping and Dr
Fang Zeng for the assistance on manuscript revision. Thanks also to
Ms Qian Wu, Ms Ting Pan, Mr Hang Zheng and Ms Lixia Qing for their
technical support with experimental study and data collection.
Funding
The present study was supported by the National
Natural Science Foundation of China (Key Program; grant no.
81430100), the Major National Science and Technology Projects
Creation of Major New Drugs (grant no. 2013ZX09103002-008) and the
Sichuan Province Outstanding Youth Academic Technology Leaders
Subsidy Scheme (grant no. 2011JQ0014).
Availability of data and materials
The analyzed datasets generated during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
SX provided experimental concepts and design,
offered scientific direction and reviewed the manuscript. WF
performed the research, the data collection and analysis and
drafted the manuscript. TM and YD performed the research and aided
manuscript revision. JW and HC performed the assessment of murine
learning and memory abilities examined by MWM and participated in
data collection. All authors participated in the manuscript
preparation and read and approved the final manuscript.
Ethics approval and consent to
participate
The treatment and care of animals adhered to the
international laws and regulations indicated in the National
Institutes of Health Guide for the Care and Use of Laboratory
Animals. All animal protocols were approved by the Institutional
Animal Care and Use Committee of Dongfang Hospital affiliated with
the Beijing University of Chinese Medicine (Beijing, China).
Consent for publication
Not applicable.
Competing interests
All authors declare that they have no competing
interests.
References
|
1
|
Wilhelmus MM, Otte-Höller I, Wesseling P,
de Waal RM, Boelens WC and Verbeek MM: Specific association of
small heat shock proteins with the pathological hallmarks of
Alzheimer's disease brains. Neuropathol Appl Neurobiol. 32:119–130.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Reitz C, Brayne C and Mayeux R:
Epidemiology of Alzheimer disease. Nat Rev Neurol. 7:137–152. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Golde TE: The Abeta hypothesis: Leading us
to rationally-designed therapeutic strategies for the treatment or
prevention of Alzheimer disease. Brain Pathol. 15:84–87. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Schenk D, Basi GS and Pangalos MN:
Treatment strategies targeting amyloid β-protein. Cold Spring Harb
Perspect Med. 2:a0063872012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Selkoe DJ and Hardy J: The amyloid
hypothesis of Alzheimer's disease at 25 years. EMBO Mol Med.
8:595–608. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Lane RF, Shineman DW, Steele JW, Lee LB
and Fillit HM: Beyond amyloid: The future of therapeutics for
Alzheimer's disease. Adv Pharmacol. 64:213–271. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Obregon D, Hou H, Deng J, Giunta B, Tian
J, Darlington D, Shahaduzzaman M, Zhu Y, Mori T, Mattson MP and Tan
J: Soluble amyloid precursor protein-α modulates β-secretase
activity and amyloid-β generation. Nat Commun. 3:7772012.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
George-Hyslop S and Fraser PE: Assembly of
the presenilin γ-/ε-secretase complex. J Neurochem. 120 Suppl
1:S84–S88. 2012. View Article : Google Scholar
|
|
9
|
Zhang HM, Zhang X and Li Y: The regulation
of curcumin on the amyloidogentic pathaway of APP in Alzheimer's
disease. Chin Pharmacol Bull. 25:361–366. 2009.(In Chinese).
|
|
10
|
Grimm MO, Mett J, Stahlmann CP, Haupenthal
VJ, Zimmer VC and Hartmann T: Neprilysin and Aβ clearance: Impact
of the APP intracellular domain in NEP regulation and implications
in Alzheimer's disease. Front Aging Neurosci. 5:982013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wang DS, Dickson DW and Malter JS:
Beta-Amyloid degradation and Alzheimer's disease. J Biomed
Biotechnol. 2006:584062006. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kanemitsu H, Tomiyama T and Mori H: Human
neprilysin is capable of degrading amyloid beta peptide not only in
the monomeric form but also the pathological oligomeric form.
Neurosci Lett. 350:113–116. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kurochkin IV and Goto S: Alzheimer's
beta-amyloid peptide specifically interacts with and is degraded by
insulin degrading enzyme. FEBS Lett. 345:33–37. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Hayes CD, Dey D, Palavicini JP, Wang H,
Patkar KA, Minond D, Nefzi A and Lakshmana MK: Striking reduction
of amyloid plaque burden in an Alzheimer's mouse model after
chronic administration of carmustine. BMC Med. 11:812013.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Eckman EA and Eckman CB: Abeta-degrading
enzymes: Modulators of Alzheimer's disease pathogenesis and targets
for therapeutic intervention. Biochem Soc Trans. 33:1101–1105.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Siemers ER, Sundell KL, Carlson C, Case M,
Sethuraman G, Liu-Seifert H, Dowsett SA, Pontecorvo MJ, Dean RA and
Demattos R: Phase 3 solanezumab trials: Secondary outcomes in mild
Alzheimer's disease patients. Alzheimers Dement. 12:110–120. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Turtle CJ, Hanafi LA, Berger C, Hudecek M,
Pender B, Robinson E, Hawkins R, Chaney C, Cherian S, Chen X, et
al: Immunotherapy of non-Hodgkin's lymphoma with a defined ratio of
CD8+ and CD4+ CD19-specific chimeric antigen receptor-modified T
cells. Sci Transl Med. 8:355ra1162016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Dai Y, Ma T, Ren X, Wei J, Fu W, Ma Y, Xu
S and Zhang Z: Tongluo Xingnao Effervescent Tablet preserves
mitochondrial energy metabolism and attenuates cognition deficits
in APPswe/PS1De9 mice. Neurosci Lett. 630:101–108. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Xu SJ, Dai Y, Zhang YJ, Xiong M, Ma YT and
Zhong ZD: Effects of tongluo xingnao effervescent tablet on
expression of CKD5 and GSK-3 in rat model of hippocampus with Aβ
injection. West China J Pharm Sci. 28:140. 2013.(In Chinese).
|
|
20
|
Zhang YJ, Ju SH, Hu Y, Ren XY and Xu SJ:
Effect of Tongluo Xingnao effervescent tablets on cerebral
cholinergic function of mice dementia model induced by scopolamine.
Tradit Chin Drug Res Pharmacol. 25:272–276. 2014.(In Chinese).
|
|
21
|
Zhang YJ, Dai Y, Hu Y, Ma YT, Xu SJ and
Wang YY: Effect of tongluo xingnao effervescent tablet on learning
and memory of AD rats and expression of insulin-degrading enzyme in
hippocampus. Zhongguo Zhong Yao Za Zhi. 38:2863–2867. 2013.(In
Chinese). PubMed/NCBI
|
|
22
|
Xu SJ and Dai Y: A pharmaceutical
composition for treating neurodegenerative diseases and its
preparation method and application CN Patent CN103181954A. Filed
April 12, 2013; issued July 3. 2013, (In Chinese).
|
|
23
|
Ren X, Wei J, Gong D, Hu Y, Chen H and Xu
S: Tongluoxingnao effervescent tablets ameliorate learning and
memory impairment in a rat model of vascular dementia via the
regulation of the p38 and ERK MAPK signaling pathways. Int J Clin
Exp Med. 9:5400–5412. 2016.
|
|
24
|
Institute of Laboratory Animal Resources
(US), Committee on Care and Use of Laboratory Animals, National
Institutes of Health (US), and Division of Research Resources, .
Guide for the care and use of laboratory animals. The National
Academies Press; Washington, DC: 1985
|
|
25
|
Buttini M, Masliah E, Barbour R, Grajeda
H, Motter R, Johnson-Wood K, Khan K, Seubert P, Freedman S, Schenk
D and Games D: Beta-amyloid immunotherapy prevents synaptic
degeneration in a mouse model of Alzheimer's disease. J Neurosci.
25:9096–9101. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Li T, Li YM, Ahn K, Price DL, Sisodia SS
and Wong PC: Increased expression of PS1 is sufficient to elevate
the level and activity of γ-secretase in vivo. PLoS One.
6:e281792011. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Grimm A, Friedland K and Eckert A:
Mitochondrial dysfunction: The missing link between aging and
sporadic Alzheimer's disease. Biogerontology. 17:281–296. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Selkoe DJ and Hardy J: The amyloid
hypothesis of Alzheimer's disease at 25 years. EMBO Mol Med.
8:595–608. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Izzo NJ, Staniszewski A, To L, Fa M, Teich
AF, Saeed F, Wostein H, Walko T III, Vaswani A, Wardius M, et al:
Alzheimer's therapeutics targeting Amyloid beta 1–42 oligomers I:
Abeta 42 oligomer binding to specific neuronal receptors is
displaced by drug candidates that improve cognitive deficits. PLoS
One. 9:e1118982014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Sevigny J, Chiao P, Bussière T, Weinreb
PH, Williams L, Maier M, Dunstan R, Salloway S, Chen T, Ling Y, et
al: The antibody aducanumab reduces Aβ plaques in Alzheimer's
disease. Nature. 537:50–56. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wang YJ, Zhou HD and Zhou XF: Clearance of
amyloid-beta in Alzheimer's disease: Progress, problems and
perspectives. Drug Discov Today. 11:931–938. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Liu XP, Qian X, Xie Y, Qi Y, Peng MF, Zhan
BC and Lou ZQ: Betaine suppressed Aβ generation by altering amyloid
precursor protein processing. Neurol Sci. 35:1009–1013. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Siemers ER, Quinn JF, Kaye J, Farlow MR,
Porsteinsson A, Tariot P, Zoulnouni P, Galvin JE, Holtzman DM,
Knopman DS, et al: Effects of a gamma-secretase inhibitor in a
randomized study of patients with Alzheimer disease. Neurology.
66:602–604. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Iwata N, Tsubuki S, Takaki Y, Watanabe K,
Sekiguchi M, Hosoki E, Kawashima-Morishima M, Lee HJ, Hama E,
Sekine-Aizawa Y and Saido TC: Identification of the major
Abeta1-42-degrading catabolic pathway in brain parenchyma:
Suppression leads to biochemical and pathological deposition. Nat
Med. 6:143–150. 2000. View
Article : Google Scholar : PubMed/NCBI
|
|
35
|
El-Amouri SS, Zhu H, Yu J, Gage FH, Verma
IM and Kindy MS: Neprilysin protects neurons against Abeta peptide
toxicity. Brain Res. 1152:191–200. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Marr RA, Rockenstein E, Mukherjee A, Kindy
MS, Hersh LB, Gage FH, Verma IM and Masliah E: Neprilysin gene
transfer reduces human amyloid pathology in transgenic mice. J
Neurosci. 23:1992–1996. 2003.PubMed/NCBI
|
|
37
|
Bulloj A, Leal MC, Surace EI, Zhang X, Xu
H, Ledesma MD, Castaño EM and Morelli L: Detergent resistant
membrane-associated IDE in brain tissue and cultured cells:
Relevance to Abeta and insulin degradation. Mol Neurodegener.
3:222008. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Farris W, Mansourian S, Chang Y, Lindsley
L, Eckman EA, Frosch MP, Eckman CB, Tanzi RE, Selkoe DJ and
Guenette S: Insulin-degrading enzyme regulates the levels of
insulin, amyloid beta-protein, and the beta-amyloid precursor
protein intracellular domain in vivo. Proc Natl Acad Sci USA.
100:pp. 4162–4167. 2003; View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Wu TY, Chen CP and Jinn TR: Traditional
Chinese medicines and Alzheimer's disease. Taiwan J Obstet Gynecol.
50:131–135. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Howes MJR and Houghton PJ: Plants used in
Chinese and Indian traditional medicine for improvement of memory
and cognitive function. Pharmacol Biochem Behav. 75:513–527. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Dos S, Antos-Neto LL, de Vilhena Toledo
MA, Medeiros-Souza P and de Souza GA: The use of herbal medicine in
Alzheimer's disease: a systematic review. Evid Based Complement
Alternat Med. 3:441–445. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Zhang YJ, Xu SJ, Dai Y, Xiong M and Ma YT:
Effects of Tongluoxingnao Effervescent Tablet on synaptophysin of
injecting Aβ in the hippocampus rat model. Pharm Clin Chin Mat Med.
28:84–87. 2012.(In Chinese).
|
|
43
|
Ren XY, Hu Y, Wei JP, Ma YT and Xu SJ:
Effects of Tongluo Xingnao effervescent tablets on expression of
cytochrome C oxidase in cells in hippocampus of VD rat model. Chin
J Basic Med Tradit Chin Med. 2:159–161. 2015.(In Chinese).
|
|
44
|
Ren XY, Hu Y, Wei JP, Fu WJ, Xu SJ and
Wang YY: Effects of Tongluo Xingnao effervescent tablets on blood
rheology, iNOS, VEGF and LDH-5 in MID rats. Zhongguo Zhong Yao Za
Zhi. 41:1119–1123. 2016.(In Chinese). PubMed/NCBI
|
|
45
|
Huang H, Nie S, Cao M, Marshall C, Gao J,
Xiao N, Hu G and Xiao M: Characterization of AD-like phenotype in
aged APPSwe/PS1dE9 mice. Age (Dordr). 38:303–322. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Romberg C, Mattson MP, Mughal MR, Bussey
TJ and Saksida LM: Impaired attention in the 3xTgAD mouse model of
Alzheimer's disease: Rescue by donepezil (Aricept). J Neurosci.
31:3500–3507. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Stahl SM: The new cholinesterase
inhibitors for Alzheimer's disease, Part 1: Their similarities are
different. J Clin Psychiatry. 61:710–711. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Gandy S: The role of cerebral amyloid beta
accumulation in common forms of Alzheimer disease. J Clin Invest.
115:1121–1129. 2005. View Article : Google Scholar : PubMed/NCBI
|