Introduction
Renal cell carcinoma (RCC) is the most common type
of malignant kidney cancer and occurs primarily on the renal
tubular epithelial system, excluding metastatic neoplasms. In
addition, due to the aberrant expression of genes in renal tubular
epithelial cells, they gradually transform into RCC cells (1). In a study conducted in 2014, ~3.9% of
new cancer cases were RCC, with a median age of 64 years at
diagnosis in the USA alone (2). Due
to the lack of typical clinical symptoms and an early diagnostic
marker for RCC, metastasis has already developed in ~40% of
patients when RCC is diagnosed (3).
Currently, surgery remains to be the most effective treatment
strategy for RCC and an abundance of postoperative adjuvant
therapies have been approved for treatment of RCC; however, the
most effective treatment is unknown (2,4).
Therefore, it is necessary to identify a highly specific
RCC-associated maker for early diagnosis of RCC.
MicroRNA (miRNA) are a family of short non-coding
RNA, with a length of 19–25 nucleotides. miRNA guide the
RNA-induced silencing complex to miRNA target sites (3′
untranslated region of mRNA) and then regulate gene expression at
the post-translational level, leading to the inhibition of
translation or mRNA degradation (5).
With increasing research, more and more studies have demonstrated
that miRNA may serve an important role in the occurrence and
development of various cancer types (6–8), such as
RCC (9–11).
miR-211-5p was predicted using computational methods
using conservation with mouse and Fugu rubripes sequences (12). The sequence maps to human chromosome
15 and miR-211-5p has been demonstrated to serve an important role
in several cancer types, including colorectal cancer (CRC)
(13), gastric cancer (14), non-small cell lung cancer (15) and hepatocellular carcinoma (16). The aim of the present study was to
reveal the expression and function of miR-211-5p in RCC.
Materials and methods
Ethics statement
All patients signed the informed consent forms prior
to initiation of the present study. The present study was approved
by the Ethical Review Committee of the Peking University Shenzhen
Hospital (Shenzhen, China) and complied with the Declaration of
Helsinki.
Specimens and cell lines
RCC tissues and paired adjacent normal tissues were
obtained from patients undergoing surgery at the Peking University
Shenzhen Hospital from the 17th of March 2013 to the 30th of
December 2015. The patients had not received any anticancer
treatment prior to surgery. A total of 24 patients were enrolled in
the present study (18 males and 6 females), with a mean age of 51
years. Once RCC tissues and paired adjacent normal tissues were
completely removed, all tissues were immediately collected and
stored at −80°C until the RNA was later extracted. The clinical and
pathological characteristics of the patients are presented in
Table I.
 | Table I.Clinicopathological characteristics of
patients with renal cell carcinoma. |
Table I.
Clinicopathological characteristics of
patients with renal cell carcinoma.
| Characteristic | Number of
cases |
|---|
| Mean age, range
(years) | 51 (27–72) |
| Sex |
|
|
Male | 18 |
|
Female | 6 |
| Histological
type |
|
| Clear
cell | 20 |
|
Papillary | 4 |
| Fuhrman grade |
|
| I | 15 |
| II | 7 |
|
III | 1 |
| IV | 1 |
| American Joint
Committee on Cancer clinical stage |
|
| I | 15 |
| II | 8 |
|
III+IV | 1 |
RCC cell lines (ACHN, 786O, Caki-1 and 769P) and the
human embryo kidney cell line (293T) were acquired from the
Guangdong and Shenzhen Key Laboratory of Male Reproductive Medicine
and Genetics (Shenzhen, China). All cells were cultured in
Dulbecco's modified Eagle's medium (DMEM; Invitrogen; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) supplemented with 10% fetal
bovine serum (FBS; Hyclone; GE Healthcare Life Sciences, Logan, UT,
USA), 1% antibiotics (100 U/ml penicillin and 100 mg/ml
streptomycin; Gibco; Thermo Fisher Scientific, Inc.) and 1%
glutamine. The cells were incubated in a 5% CO2 humidified
incubator at 37°C.
RNA extraction and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
According to the manufacturer's protocol, total RNA
from tissues and cells was isolated using TRIzol reagent
(Invitrogen; Thermo Fisher Scientific, Inc.) and purified using an
RNeasy Maxi kit (Qiagen GmbH, Hilden, Germany). Following the
measurement of the RNA concentration using a NanoDrop 2000c (Thermo
Fisher Scientific, Inc.), an miScript II RT kit (Qiagen GmbH) was
used to synthetize cDNA with reverse transcriptase, according to
manufacturer's protocol. The temperature protocol was as follows:
37°C for 60 min, 95°C for 5 min and kept at 4°C. Subsequently, the
expression of miR-211-5p was detected with an miScript
SYBR®Green PCR kit (Qiagen GmbH) by qPCR on a Roche
light cycler 480 Real-Time PCR System (Roche Diagnostics, Basel,
Switzerland). The 10 µl reaction mixture consisted of 5 µl 2X
QuantiTect SYBR Green PCR Master mix, 3.7 µl RNase-free water, 1 µl
cDNA template, 0.4 µl specific miRNA primer and 10X miScript
Universal Primer. The forward primer of miR-211-5p was
5′-TTCCCTTTGUCATCCTTCGCCT-3′ and the reverse primer was a universal
primer, which was provided in the miScript SYBR®Green
PCR kit. U6 was chosen as an internal control. The U6 primer
sequences were as follows: Forward 5′-CTCGCTTCGGCAGCACA-3′ and
reverse 5′-ACGCTTCACGAATTTGCGT-3′. The thermocycling conditios were
as follows: 95°C for 2 min, 95°C for 10 sec, 55°C for 30 sec and
72°C for 30 sec, for 40 cycles. The expression levels of miR-211-5p
in tissues and cells were analyzed using the 2−ΔΔCq
method (17).
Cell transfection
The expression of miR-211-5p in 786O and ACHN cells
was upregulated/downregulated by transfection of the synthesized
miR-211-5p mimic/inhibitor (Shanghai GenePharma Co., Ltd.,
Shanghai, China) using Lipofectamine 2000® (Invitrogen;
Thermo Fisher Scientific, Inc.) and Opti-MEM® I Reduced
Serum Medium (Gibco; Thermo Fisher Scientific, Inc.), following the
manufacturer's protocol. The concentration of the miRNA mimic and
inhibitors used for transfection are presented in Table II. The efficiency of transfection
was measured by RT-qPCR as aforementioned. The sequences of the
miRNA and primers used are presented in Table III.
| Table II.Concentration of the miRNA
mimic/inhibitors used for the transfection. |
Table II.
Concentration of the miRNA
mimic/inhibitors used for the transfection.
| Plate | siRNA | Final volume | Lipo2000 |
|---|
| 96-well | 0.5 µl (5
pmol) | 100 µl | 0.25 µl |
| 24-well | 1 µl (20 pmol) | 500 µl | 1 µl |
| 12-well | 2 µl (40 pmol) | 1 ml | 2 µl |
|
6-well | 5 µl (100
pmol) | 2 ml | 5 µl |
| Table III.Sequences of primers and miRNA. |
Table III.
Sequences of primers and miRNA.
| Primer/miRNA | Direction | Sequence
(5′-3′) |
|---|
| miR-211-5p | F |
TTCCCTTTGUCATCCTTCGCCT |
|
| R | Universal primers
(miScript SYBR Green polymerase chain reaction kit) |
| U6 | F |
CTCGCTTCGGCAGCACA |
|
| R |
ACGCTTCACGAATTTGCGT |
| miR-211-5p
mimic | F |
UUCCCUUUGUCAUCCUUCGCCU |
|
| R |
GCGAAGGAUGACAAAGGGAAUU |
| miR-211-5p
inhibitor | – |
AGGCGAAGGAUGACAAAGGGAA |
| NC | F |
UUCUCCGAACGUGUCACGUTT |
|
| R |
ACGUGACACGUUCGGAGAATT |
| Inhibitor NC | – |
CAGUACUUUUGUGUAGUACAA |
MTT assay
The cell activity of the 786O and ACHN cells was
determined using an MTT assay. Cells (~5,000 cells/well) were
seeded into 96-well plates and transfected with miR-211-5p mimic,
inhibitor, negative control (NC) and inhibitor NC using
Lipofectamine® 2000. A total of 96 h later, 20 µl MTT (5
mg/ml; Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) was added
into each well of the 96-well plate. Following 4 h at 37°C, 100 µl
dimethyl sulfoxide (Sigma-Aldrich; Merck KGaA) was added to each
well. Subsequently, the 96-well plate was shocked in a
reciprocating decolorization shaking table (TSB-108; Qilinbeier,
Jiangsu, China) for 10 min in the dark at room temperature.
Finally, the optical density (OD) value of each well was measured
by an ELISA microplate reader (Bio-Rad Laboratories, Inc.,
Hercules, CA, USA) at a wavelength of 595 nm (with 620 nm as the
reference wavelength).
Cell Counting Kit-8 (CCK-8) assay
The proliferation ability of the 786O and ACHN cells
was analyzed using a CCK-8 assay (Beyotime Institute of
Biotechnology, Shanghai, China). Cells (~5,000 cells/well) were
seeded into a 96-well plate and transfected according to the
aforementioned protocol. Following transfection, 10 µl CCK-8
solution was added to each well and culture was continued for 30
min in the dark at room temperature. The OD value of each well was
then measured using an ELISA microplate reader at a wavelength of
450 nm (with 620 nm as the reference wavelength) at 0, 24, 48 and
72 h following transfection.
Transwell assay
The migration and invasion ability of the 786O and
ACHN cells in vitro was examined by Transwell assays.
Transwell chambers (BD Biosciences, Franklin Lakes, NJ, USA) with
Matrigel were used to assess invasion ability, while the Transwell
chambers without Matrigel were used to assess migration ability. A
total of 24 h after transfection, ~2×104 cells were added into each
upper chamber with DMEM and DMEM supplemented with 10% FBS was
added to the lower chambers. The chambers were incubated for 48 h
in a 5% CO2 incubator at 37°C. The cells in the lower chamber were
fixed and stained successively by 4% paraformaldehyde and crystal
violet each for 20 min at room temperature. Finally, a light
microscope was used to observe the cells in the bottom of chamber
at a magnification of ×100.
Wound scratch assay
The migration ability of the 786O and ACHN cells
in vitro was also examined by wound scratch assays. In
total, ~1×106 cells were cultured in each well of a 6-well plate.
After 24 h, cells were transfected with miR-211-5p mimic,
inhibitor, NC and inhibitor NC using Lipofectamine®
2000. A vertical line was scratched in the cells using a sterile
1-ml pipette tip. The images of the scratches were respectively
captured by a digital camera system at 0, 12 and 24 h.
Flow cytometry assay
The apoptosis rate of the 786O and ACHN cells in
vitro was analyzed by a flow cytometry assay. In total, ~1×106
cells were incubated in each well of the 6-well plate and were
transfected following the manufacturer's instructions. All cells
were harvested after 48 h and washed twice with cold PBS. Following
this, 100 µl 1X binding buffer (Invirtogen; Thermo Fisher
Scientific, Inc.) was used to resuspend the cells. Next, 5 µl
Annexin V-fluorescein isothiocyanate (Invitrogen; Thermo Fisher
Scientific, Inc.) and 5 µl propidium iodide (Invitrogen; Thermo
Fisher Scientific, Inc.) were added to the experimental group and
stained for 15 min in the dark at room temperature. Finally, the
apoptosis rate was analyzed by flow cytometry (EPICS Xl-4; Beckman
Coulter, Inc., Brea, CA, USA) after adding 400 µl 1X binding buffer
to each tube. All assays were repeated at least three times. FlowJo
7.6.1 software was utilized for the analysis of results (FlowJo
LLC, Ashland, OR, USA).
Statistical analyses
Data were presented as the mean ± standard error of
the mean. Significance of differential expression was analyzed by
using the Student's t-test, while paired t-tests were used to
compare the expression levels of miR-211-5p in matched tumor/normal
tissues. One-way analysis of variance followed by Bonferroni's post
hoc tests were used to compare the expression level of miR-211-5p
in RCC cell lines. SPSS 23.0 (IBM Corp., Armonk, NY, USA) was used
for all statistical analyses. P<0.05 was considered to indicate
a statistically significant difference.
Results
Expression level of miR-211-5p is
downregulated in RCC tissues and cell lines
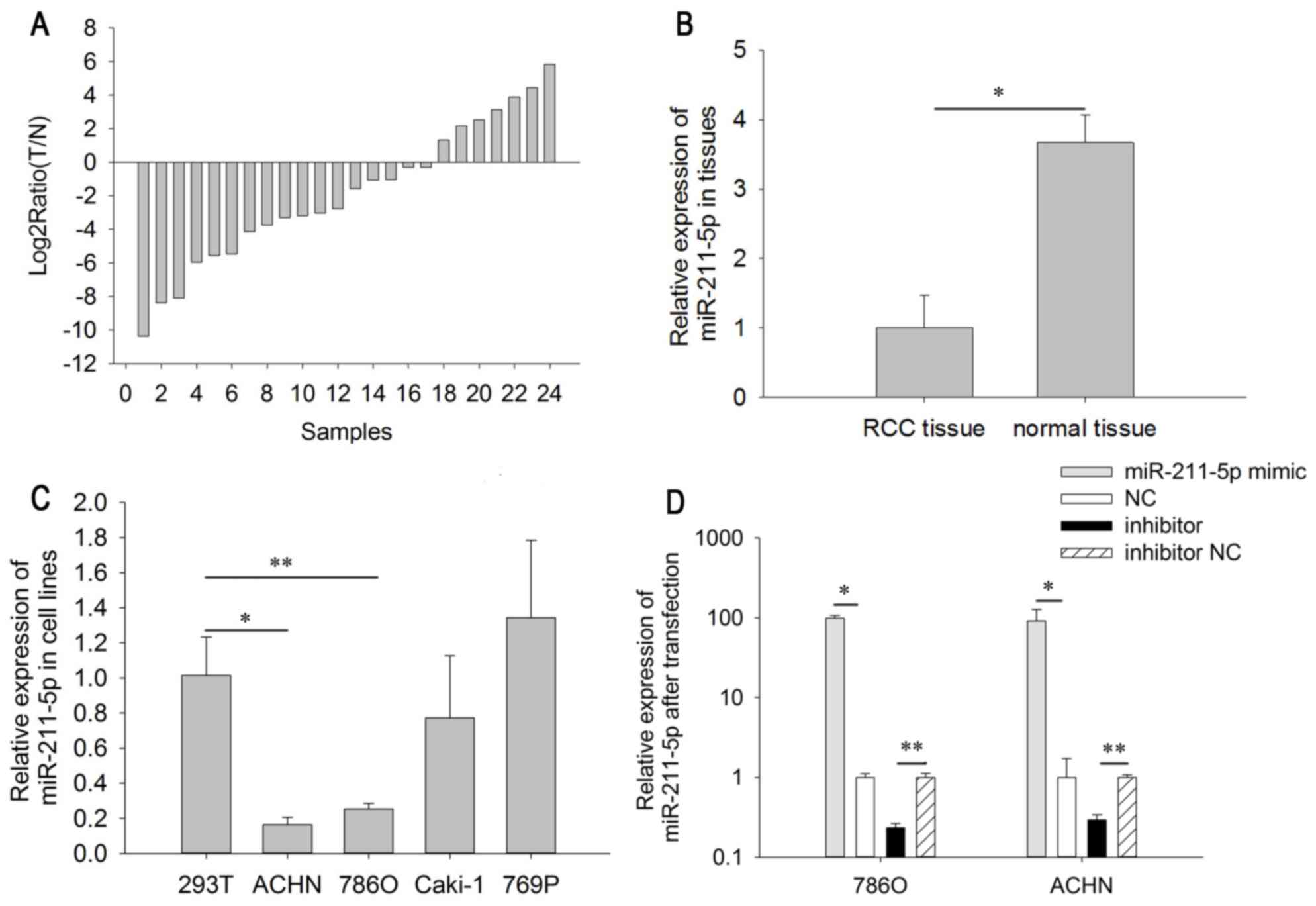
The relative expression level of miR-211-5p in RCC
tissues is demonstrated in the Fig.
1A. As indicated in Fig. 1B, the
expression level of miR-211-5p in RCC tissues (1.000±0.466) was
significantly lower than that in adjacent normal tissues
(3.664±0.407) (P<0.05). The results of miR-211-5p expression in
the cell lines demonstrated that the relative expression of
miR-211-5p was significantly lower in 786O (0.252±0.032, P<0.01)
and ACHN cells (0.253±0.032, P<0.001) than in 293T cells
(1.000±0.217). However, there was no significant difference between
the expression level in Caki-1 (0.772±0.355, P=0.401) and 769P
cells (1.341±0.441, P=0.366) compared with that in 293T cells
(Fig. 1C). The above results suggest
that miR-211-5p may act as a tumor suppressor in RCC.
Cell transfection efficiency
validation
RT-qPCR was performed to detect the transfection
efficiency of miR-211-5p mimic or inhibitor, compared with NC or
inhibitor NC. The results demonstrated that the expression levels
of miR-211-5p were 0.234±0.032 times higher (786O cells, P=0.003)
and 0.293±0.048 times higher (ACHN cells, P=0.005) in cells
transfected with miR-211-5p inhibitor compared with the levels in
those transfected with inhibitor NC after 24 h. The expression
levels of miR-211-5p were 99.310±7.915 times higher (786O cells,
P=0.021) and 91.321±36.326 times higher (ACHN cells, P=0.018) in
cells transfected with miR-211-5p mimic compared with the levels in
those transfected with NC after 24 h (Fig. 1D).
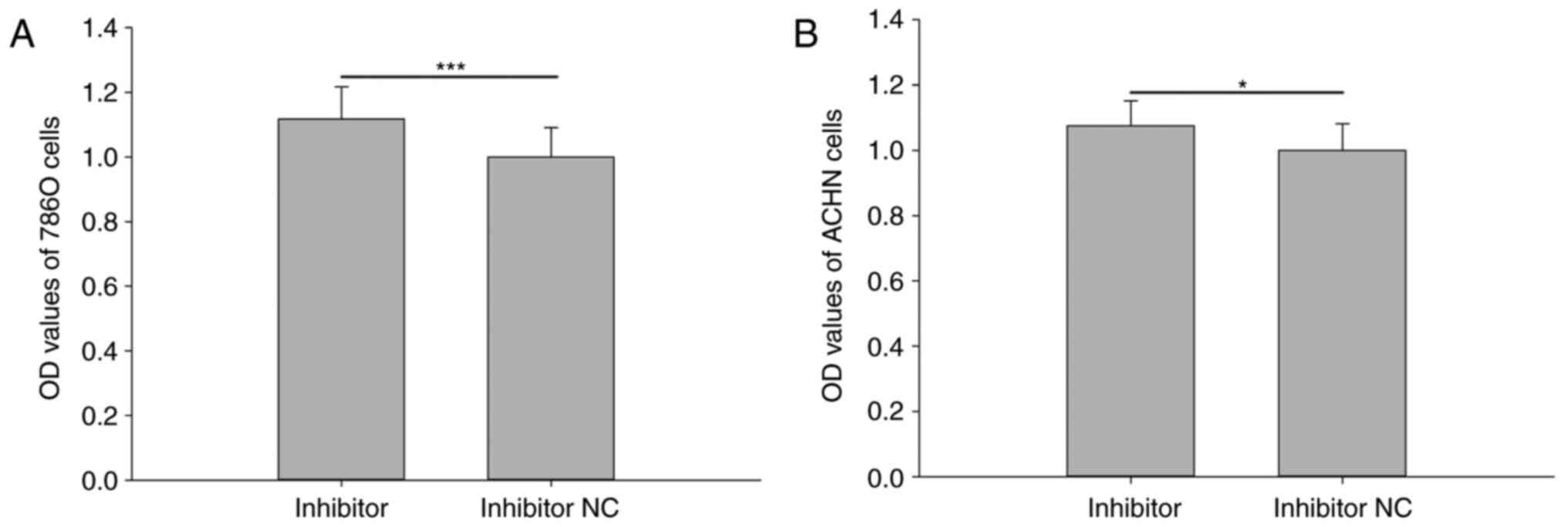
Downregulation of miR-211-5p promotes
786O and ACHN cell viability
Cell viability was analyzed using an MTT assay. The
results revealed that the relative viability of 786O cells
transfected with miR-211-5p inhibitor was significantly increased
compared with that observed in cells transfected with inhibitor NC
(1.117±0.100 vs. 1.000±0.090, respectively; P<0.001; Fig. 2A). The relative viability of ACHN
cells was also significantly increased in cells transfected with
miR-211-5p inhibitor compared with that observed in cells
transfected with inhibitor NC (1.075±0.077 vs. 1.000±0.082,
respectively; P<0.05; Fig. 2B).
However, there was no significant difference observed between the
mimic group and NC group for the viability of 786O and ACHN cells
(P>0.05; data not shown).
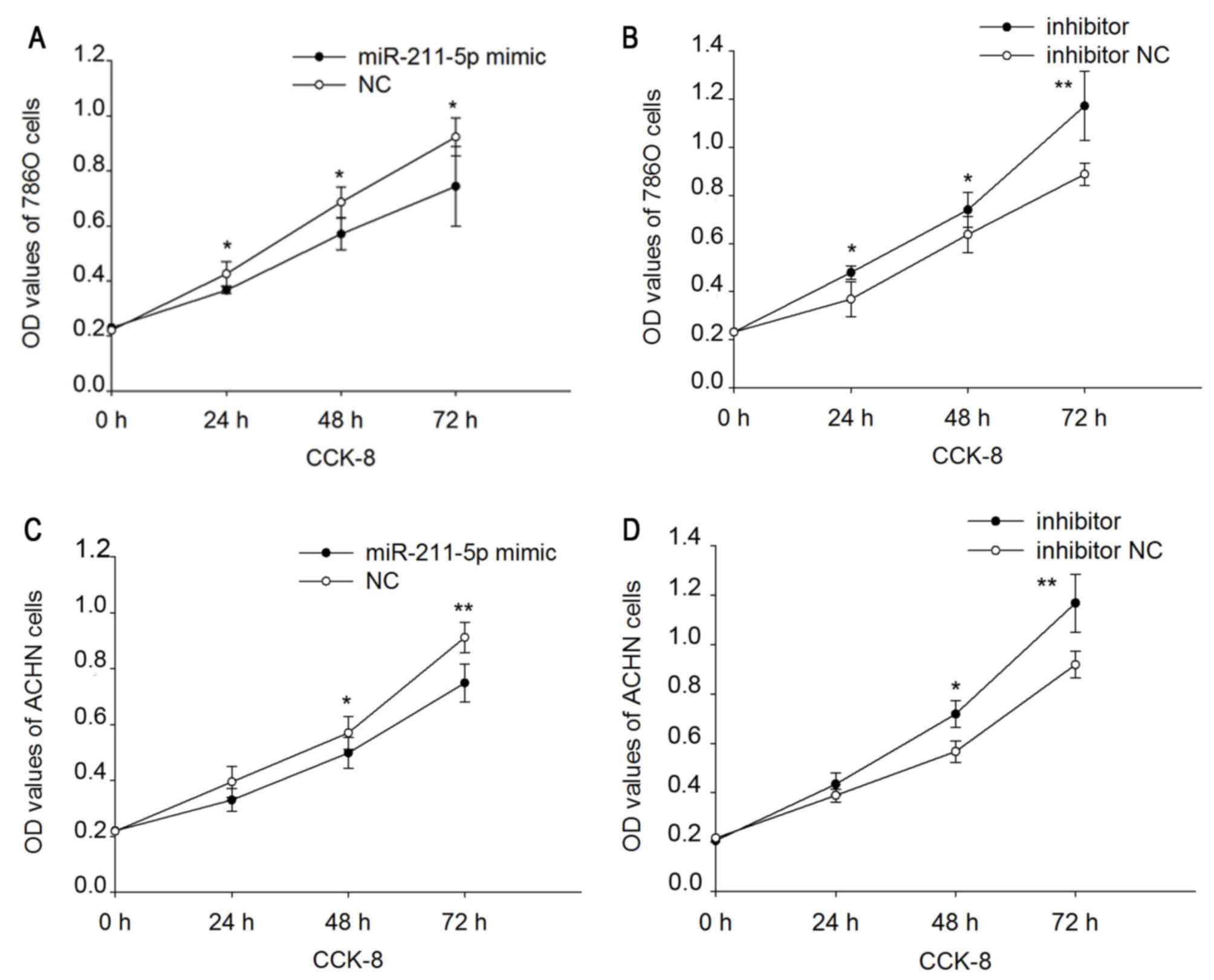
Upregulation/downregulation of
miR-211-5p inhibits/promotes 786O and ACHN cell proliferation
A CCK-8 assay was performed to assess the
proliferation ability of 786O and ACHN cells (Fig. 3). The results demonstrated that the
upregulation/downregulation of miR-211-5p inhibited/promoted the
proliferation of the 786O and ACHN cells. Proliferation was
downregulated by 14.030 (P<0.05), 16.802 (P<0.05) and 19.416%
(P<0.05) (Fig. 3A) in 786O cells,
and 16.385 (P=0.150), 12.627 (P<0.05) and 17.886% (P<0.01)
(Fig. 3C) in ACHN cells following
transfection with miR-211-5p mimic at 24, 48 and 72 h,
respectively, compared with that observed in cells transfected with
NC. Proliferation was upregulated by 29.957 (P<0.05), 16.118
(P<0.05) and 31.945% (P<0.01) (Fig. 3B) in 786O cells, and 11.691
(P=0.095), 26.840 (P<0.05) and 27.187% (P<0.01) (Fig. 3D) in ACHN cells following
transfection with miR-211-5p inhibitor at 24, 48 and 72 h,
respectively, compared with that observed in cells transfected with
inhibitor NC.
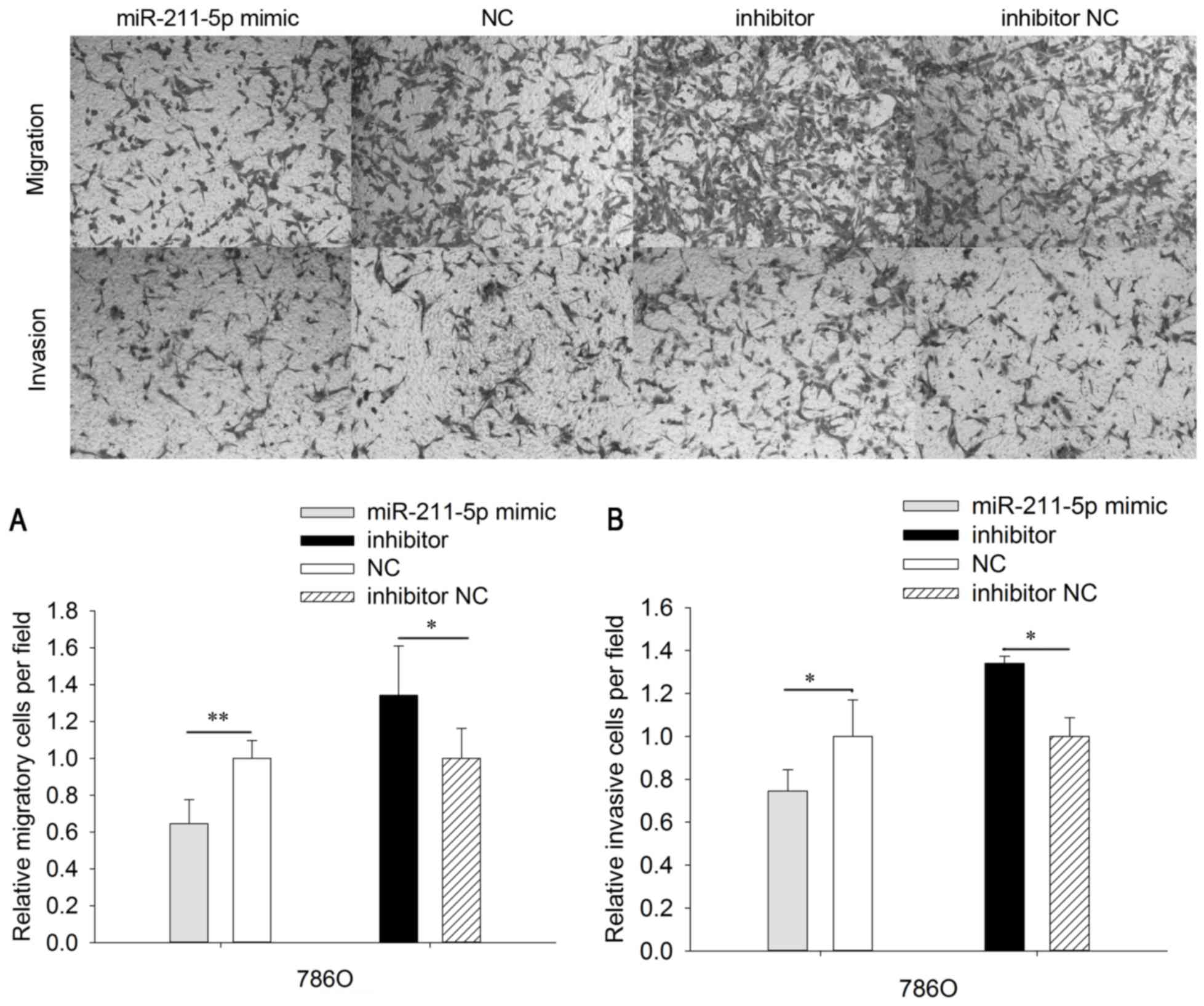
Upregulation/downregulation of
miR-211-5p inhibits/promotes 786O and ACHN cell mobility
To analyze the mobility of 786O and ACHN cells,
wound scratch and Transwell assays were designed (Figs. 4 and 5). The results of Transwell assays
demonstrated that the migratory ability of 786O cells was
significantly downregulated by 35.351% (P<0.01) in the
miR-211-5p mimic group compared with that observed in the NC group
(Fig. 4). Furthermore, the migratory
ability of 786O cells was significantly upregulated by 34.207%
(P<0.05) in the miR-211-5p inhibitor group compared with the
level observed in the inhibitor NC group. In ACHN cells, the
migratory ability was significantly downregulated by 24.582%
(P<0.01) in the miR-211-5p mimic group compared with that
observed in the NC group. The migratory ability was significantly
upregulated by 38.620% (P<0.05) in ACHN cells of the miR-211-5p
inhibitor group compared with the level observed in the inhibitor
NC group (Fig. 5).
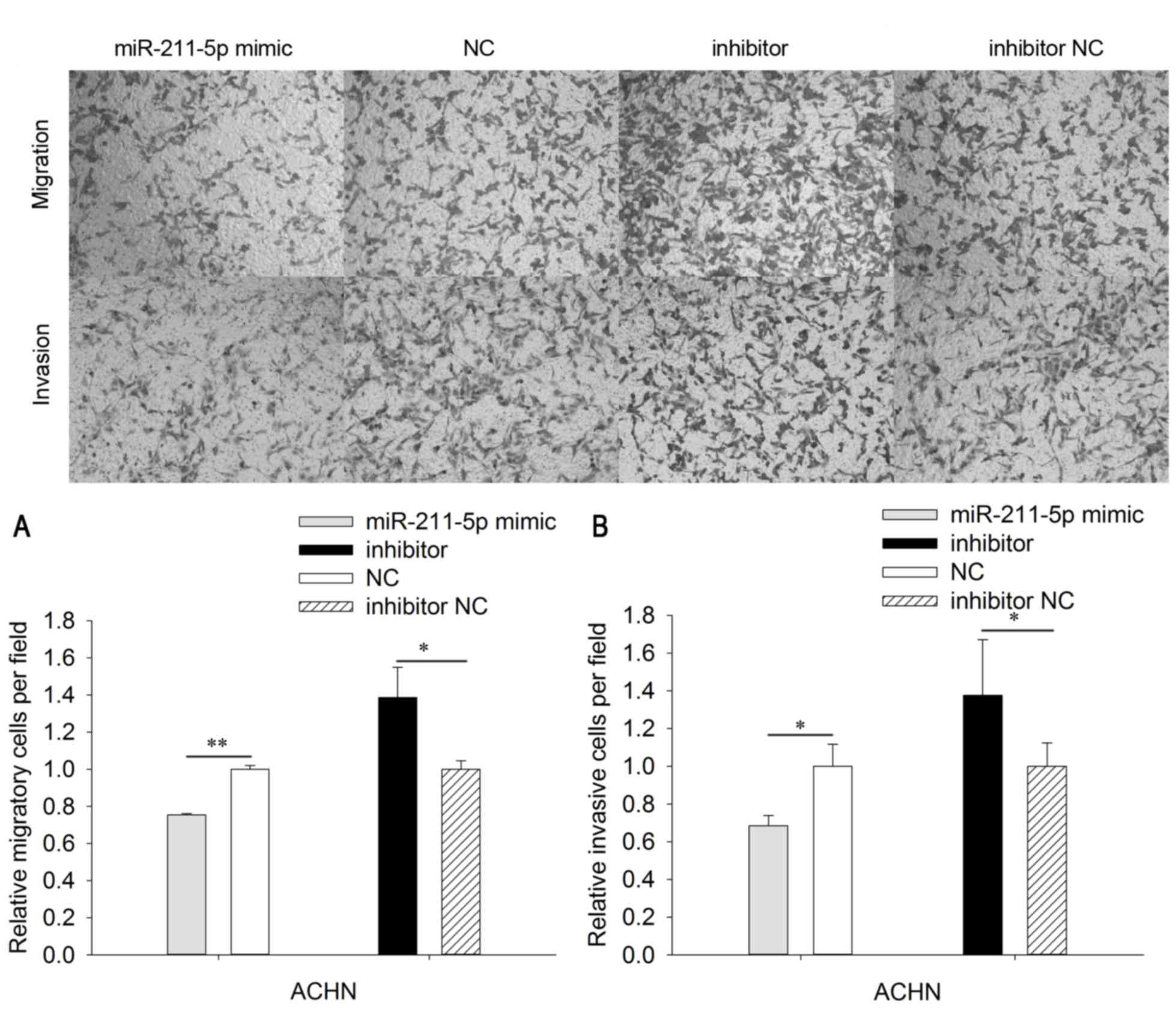
In order to determine the invasive ability of cells,
Transwell chambers with Matrigel were utilized. The invasive
ability of 786O cells was significantly downregulated by 35.351%
(P<0.05) in the miR-211-5p mimic group compared with that
observed in the NC group. The invasive ability of 786O cells was
significantly upregulated by 34.207% (P<0.05) in the miR-211-5p
inhibitor group compared with that observed in the inhibitor NC
group (Fig. 4). In addition, the
invasive ability of ACHN cells was significantly downregulated by
31.625% (P<0.05) in the miR-211-5p mimic group compared with
that observed in the NC group, while the invasive ability of ACHN
cells was significantly upregulated by 37.628% (P<0.05) in the
miR-211-5p inhibitor group compared with that observed in the
inhibitor NC group (Fig. 5).
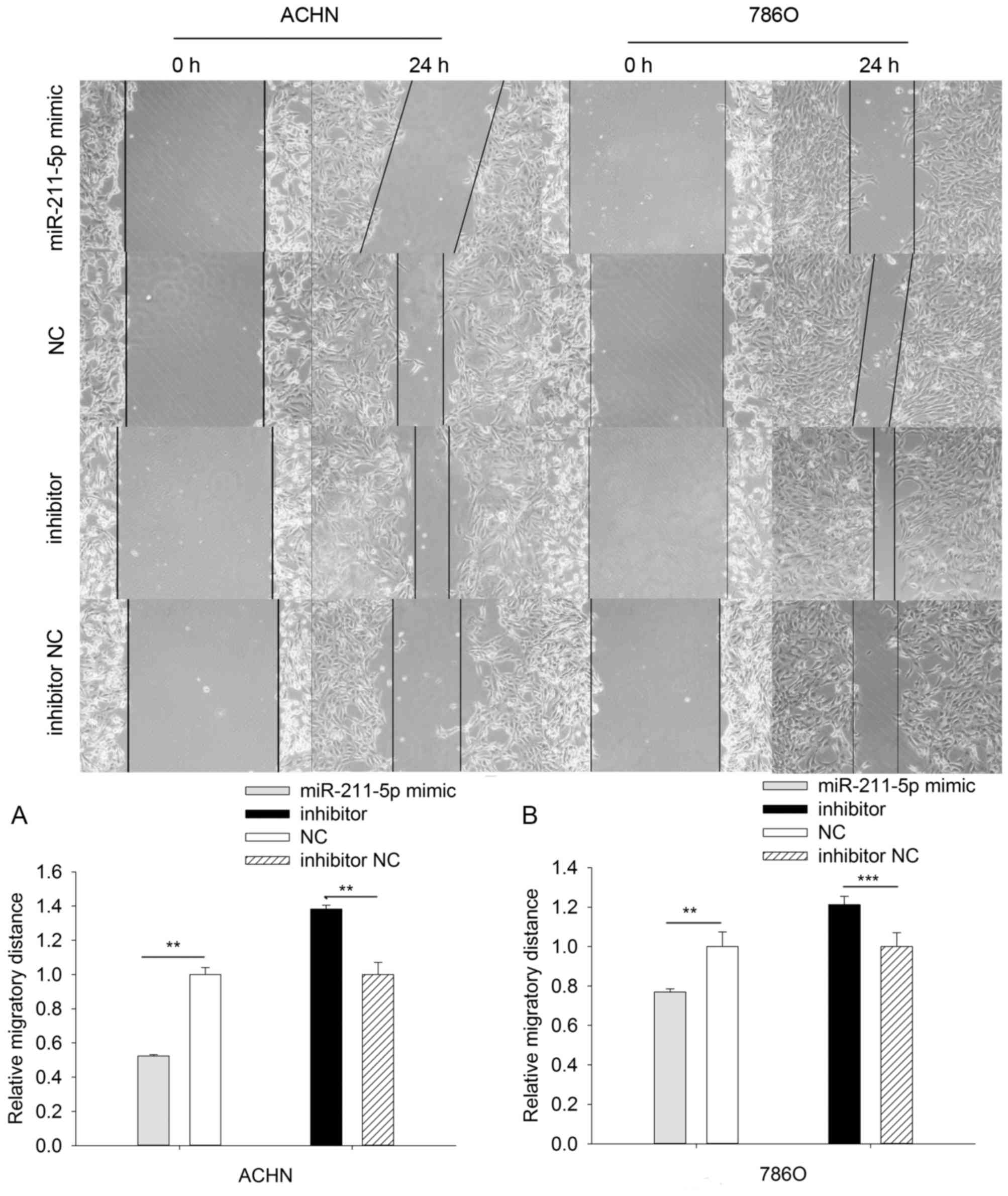
The results of the scratch wound assay demonstrated
that the migratory ability of 786O cells was significantly
downregulated by 23.131% (P<0.01) in the miR-211-5p mimic group
compared with that observed in the NC group, while the migratory
ability was significantly upregulated by 21.287% (P<0.001) in
the miR-211-5p inhibitor group compared with the observed in the
inhibitor NC group. The results in ACHN cells were similar to those
in the 786O cells, which demonstrated that the migratory ability
was significantly downregulated by 47.671% (P<0.01) in the
miR-211-5p mimic group compared with that observed in the NC group,
while the migratory ability was significantly upregulated by
38.300% (P<0.01) in the miR-211-5p inhibitor group compared with
the observed in the inhibitor NC group (Fig. 6).
Upregulation/downregulation of
miR-211-5p induces/inhibits 786O and ACHN cell apoptosis
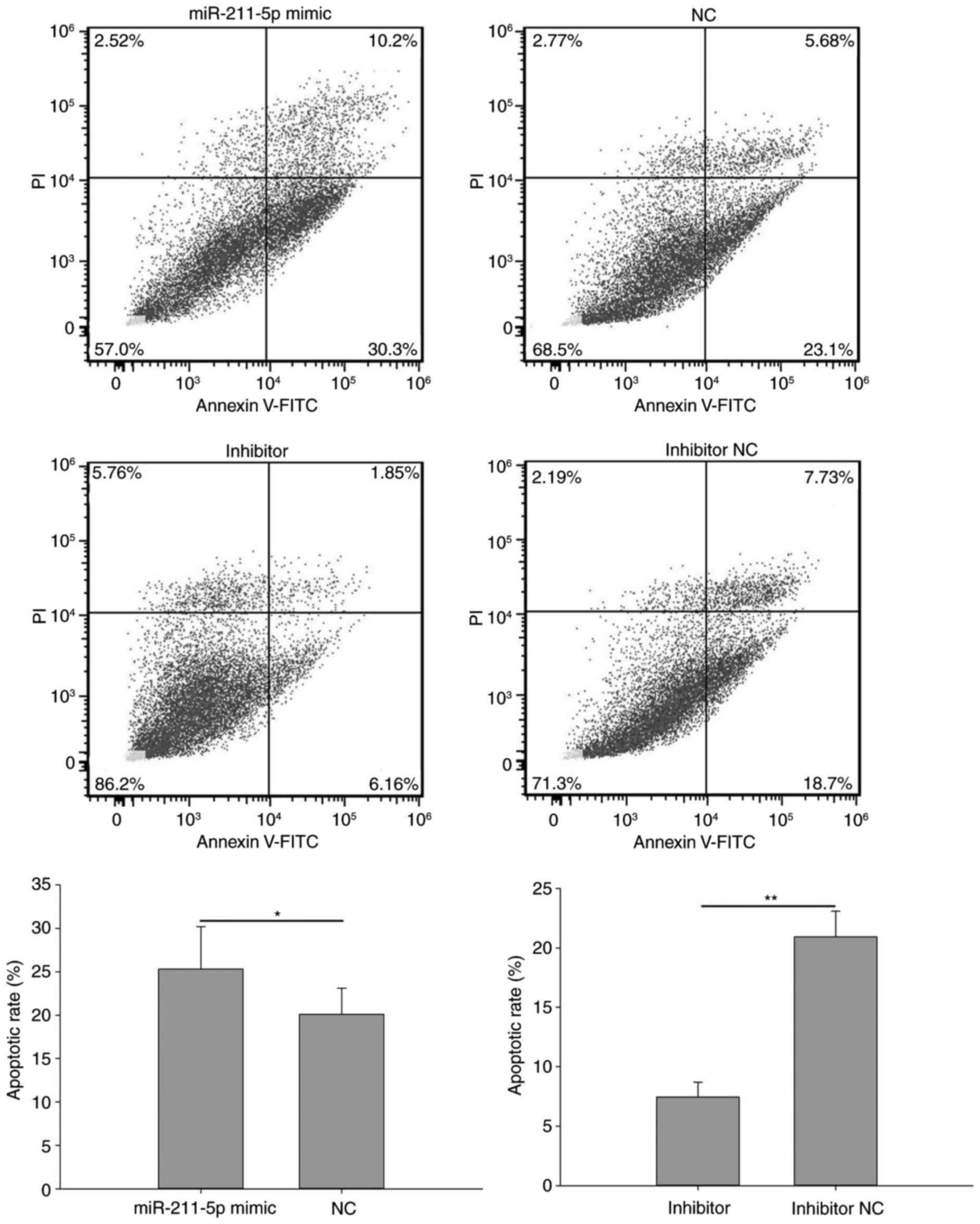
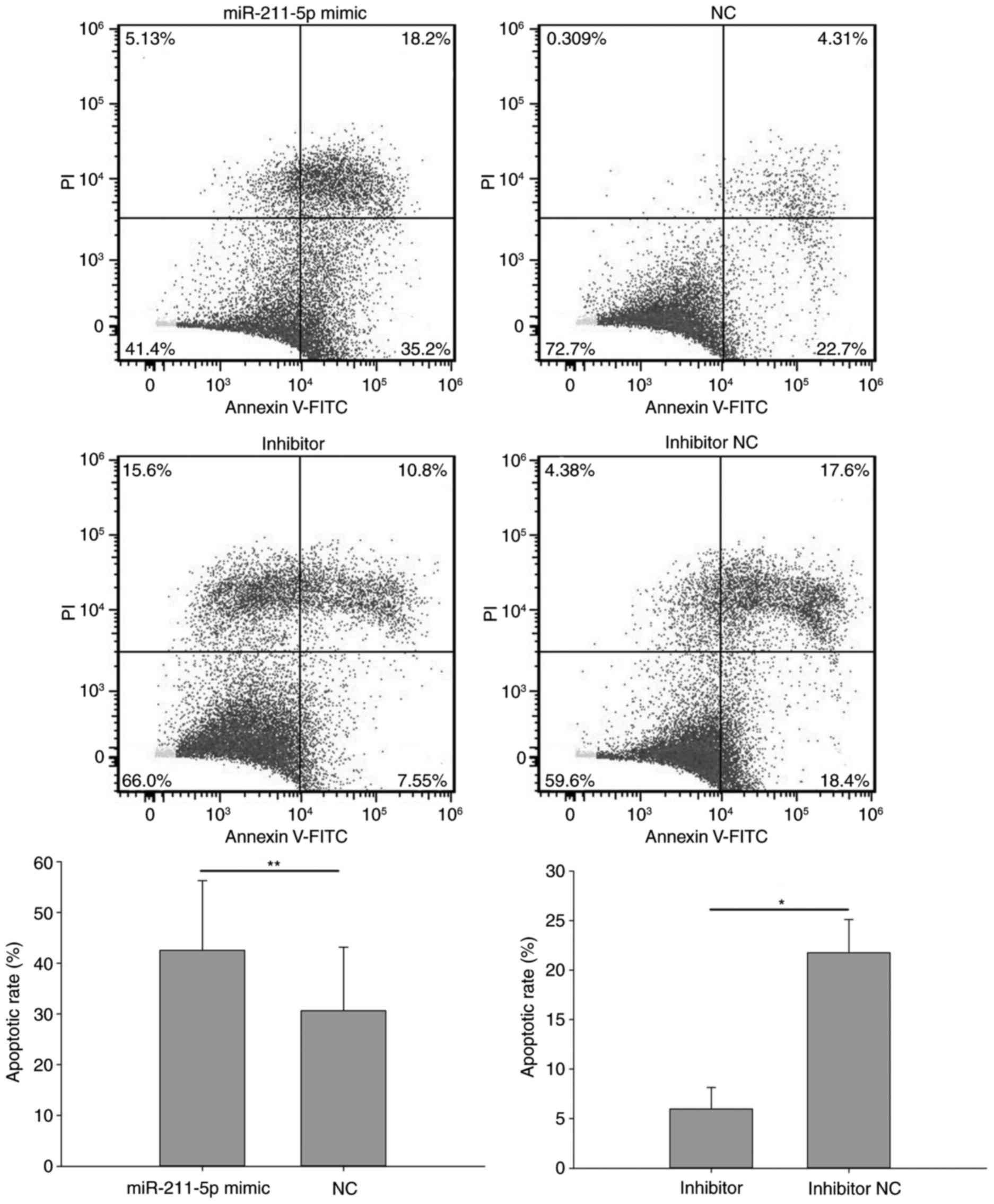
Flow cytometry was performed to analyze the
apoptotic rate of cells (Figs. 7 and
8). The results demonstrated that
the apoptotic rate of 786O cells transfected with miR-211-5p mimic
was significantly increased compared with that observed in the NC
group (25.333±4.854 vs. 20.100±3.000%, respectively; P<0.05;
Fig. 7). A similar outcome was also
observed in ACHN cells, with an apoptotic rate of 42.567±13.723 vs.
30.633±12.548% (P<0.01) in the miR-211-5p mimic and NC groups,
respectively (Fig. 8). Additionally,
the apoptotic rate of 786O cells transfected with miR-211-5p
inhibitor was significantly decreased compared with that observed
in the inhibitor NC group (7.453±1.243 vs. 20.933±2.155%,
respectively; P<0.01; Fig. 7).
Similarly, the apoptotic rate of ACHN cells was 5.963±2.170 and
21.767±3.350% (P<0.05) in the inhibitor and inhibitor NC groups,
respectively (Fig. 8). These results
suggest that the upregulation/downregulation of miR-211-5p
induced/inhibited 786O and ACHN cell apoptosis.
Discussion
It is well known that miRNA are important
post-transcriptional regulators and that they are involved in
various physiological and pathological processes, including cell
differentiation, cell proliferation and tumorigenesis (18). miRNA have been demonstrated to serve
as oncogenes when they are overexpressed or tumor suppressor genes
when they downregulated (19).
Previous studies have indicated that miR-211-5p was downregulated
in hepatocellular carcinoma, gastric cancer and ovarian cancer
(14,16,20).
However, miR-211-5p was overexpressed in non-small cell lung cancer
(15). Previous microarray analyses
indicated that miR-211-5p was significantly downregulated in RCC
tissue compared with adjacent normal tissues (21). The results of the present study also
indicated that the expression of miR-211-5p was downregulated in
human RCC tissues and cell lines, according to RT-qPCR.
Furthermore, the present study also demonstrated
that miR-211-5p has a negative impact in RCC cell lines,
particularly on migration and invasion. This result was similar to
that demonstrated in a study by Wang et al (22). In addition, the present study
indicated that upregulation/downregulation of miR-211-5p
induced/inhibited 786O and ACHN cell apoptosis. This phenomenon was
also demonstrated in ovarian cancer (20). It has also been suggested that
miR-211 acted as a tumor suppressor in epithelial ovarian cancer
(EOC) and inhibited cell proliferation by regulating
cyclin-dependent kinase 6 in EOC cells (20). In addition, long non-coding (lnc)RNA
ucoo2kmd.1 was highly expressed in CRC tissues compared with
adjacent normal tissues and regulated cluster of differentiation 44
as a molecular decoy for miR-211-3p (13). However, a study by Xu et al
(23) revealed that miR-211-3p
served as an oncogene in CRC and promoted cell proliferation by
targeting lncRNA tumor suppressor candidate 7 in CRC cell lines.
Furthermore, a case-control study discovered that miR-211 was
associated with poor prognosis in CRC, while no statistically
significant differences between clinicopathological factors and
miR-211 expression level were identified in the CRC group (24).
In addition to cancer, miR-211 also serves an
important role in the initiation and development of other diseases.
A study by Sun et al (25)
indicated that miR-133, miR-135, miR-204 and miR-211 acted as
negative regulators and inhibited differentiation of
osteoprogenitors by attenuating the essential transcription factor
Runt-related transcription factor 2 (RUNX2). Furthermore, Atlasi
et al (26) demonstrated that
miR-211 acted as an endogenous attenuator of this transcription
factor and the effect in calcium deposition did not correlate with
the effect in RUNX2, which demonstrated that miR-211 could exert
its effects on calcification through the Wnt signaling pathway. In
addition, a study by Panizo et al (27) concluded that miR-29b, miR-133b and
miR-211 have direct roles in vascular smooth muscle calcification
induced by high phosphorus, and may be novel therapeutic targets in
the management of vascular calcification.
In conclusion, the present study demonstrated that
miR-211-5p was downregulated in RCC tissues and cell lines. The
present study also indicated that miR-211-5p serves important roles
in cellular functions, including proliferation, migration, invasion
and apoptosis. Thus, this study suggests that miR-211-5p acts as a
tumor suppressor in RCC. Further research should focus on the
underlying mechanism of miR-211-5p in RCC and on investigating the
possible use of miR-211-5p as a biomarker for RCC.
Acknowledgements
The authors thank reviewers for helpful comments on
the manuscript and all the patients for consenting to provide
tissue samples.
Funding
This study was supported by grants from the National
Natural Science Foundation of China (grant no. 81101922), Science
and Technology Development Fund Project of Shenzhen (grant nos.
JCYJ20150403091443329 and JCYJ20170307111334308), the fund of
‘San-ming’ Project of Medicine in Shenzhen (grant no.
SZSM201612066) and the fund of Guangdong Key Medical Subject.
Availability of data and materials
The analyzed data sets generated during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
YL and TW conceived and designed the experiments.
JQ, XP, TH, CL and PC performed the experiments, analyzed the data
and drafted the paper. ZZ and SY conceived the experiments. All
authors have read and approved this manuscript.
Ethics approval and consent to
participate
All patients signed the informed consent forms prior
to initiation of the present study. The present study was approved
by the Ethical Review Committee of the Peking University Shenzhen
Hospital (Shenzhen, China) and complied with the Declaration of
Helsinki.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Chow WH, Dong LM and Devesa SS:
Epidemiology and risk factors for kidney cancer. Nat Rev Urol.
7:245–257. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Motzer RJ, Jonasch E, Agarwal N, Beard C,
Bhayani S, Bolger GB, Chang SS, Choueiri TK, Costello BA, Derweesh
IH, et al: Kidney cancer, version 3.2015. J Natl Compr Canc Netw.
13:151–159. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Flanigan RC, Campbell SC, Clark JI and
Picken MM: Metastatic renal cell carcinoma. Curr Treat Options
Oncol. 4:385–390. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Afriansyah A, Hamid AR, Mochtar CA and
Umbas R: Targeted therapy for metastatic renal cell carcinoma. Acta
Med Indones. 48:335–347. 2016.PubMed/NCBI
|
|
5
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Shui Y, Yu X, Duan R, Bao Q, Wu J, Yuan H
and Ma C: miR-130b-3p inhibits cell invasion and migration by
targeting the Notch ligand Delta-like 1 in breast carcinoma. Gene.
609:80–87. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Chang C, Liu T, Huang Y, Qin W, Yang H and
Chen J: MicroRNA-134-3p is a novel potential inhibitor of human
ovarian cancer stem cells by targeting RAB27A. Gene. 605:99–107.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Chen T, Ren H, Thakur A, Yang T, Li Y,
Zhang S, Wang T and Chen M: miR-382 inhibits tumor progression by
targeting SETD8 in non-small cell lung cancer. Biomed Pharmacother.
86:248–253. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhou W, Bi X, Gao G and Sun L: miRNA-133b
and miRNA-135a induce apoptosis via the JAK2/STAT3 signaling
pathway in human renal carcinoma cells. Biomed Pharmacother.
84:722–729. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Jin L, Li Y, Liu J, Yang S, Gui Y, Mao X,
Nie G and Lai Y: Tumor suppressor miR-149-5p is associated with
cellular migration, proliferation and apoptosis in renal cell
carcinoma. Mol Med Rep. 13:5386–5392. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kurozumi A, Kato M, Goto Y, Matsushita R,
Nishikawa R, Okato A, Fukumoto I, Ichikawa T and Seki N: Regulation
of the collagen cross-linking enzymes LOXL2 and PLOD2 by
tumor-suppressive microRNA-26a/b in renal cell carcinoma. Int J
Oncol. 48:1837–1846. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lim LP, Glasner ME, Yekta S, Burge CB and
Bartel DP: Vertebrate microRNA genes. Science. 299:15402003.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wu X, He X, Li S, Xu X, Chen X and Zhu H:
Long Non-Coding RNA ucoo2kmd.1 regulates CD44-dependent cell growth
by competing for miR-211-3p in colorectal cancer. PLoS One.
11:e01512872016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wang CY, Hua L, Sun J, Yao KH, Chen JT,
Zhang JJ and Hu JH: MiR-211 inhibits cell proliferation and
invasion of gastric cancer by down-regulating SOX4. Int J Clin Exp
Pathol. 8:14013–14020. 2015.PubMed/NCBI
|
|
15
|
Ye L, Wang H and Liu B: miR-211 promotes
non-small cell lung cancer proliferation by targeting SRCIN1.
Tumour Biol. 37:1151–1157. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Jiang G, Cui Y, Yu X, Wu Z, Ding G and Cao
L: miR-211 suppresses hepatocellular carcinoma by downregulating
SATB2. Oncotarget. 6:9457–9466. 2015.PubMed/NCBI
|
|
17
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Lee RC, Feinbaum RL and Ambros V: The C.
elegans heterochronic gene lin-4 encodes small RNAs with antisense
complementarity to lin-14. Cell. 75:843–854. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Shenouda SK and Alahari SK: MicroRNA
function in cancer: Oncogene or a tumor suppressor? Cancer
Metastasis Rev. 28:369–378. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Xia B, Yang S, Liu T and Lou G: miR-211
suppresses epithelial ovarian cancer proliferation and cell-cycle
progression by targeting Cyclin D1 and CDK6. Mol Cancer. 14:572015.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Cheng T, Wang L, Li Y, Huang C, Zeng L and
Yang J: Differential microRNA expression in renal cell carcinoma.
Oncol Lett. 6:769–776. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wang K, Jin W, Jin P, Fei X, Wang X and
Chen X: miR-211-5p suppresses metastatic behavior by targeting
SNAI1 in renal cancer. Mol Cancer Res. 15:448–456. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Xu J, Zhang R and Zhao J: The novel long
noncoding RNA TUSC7 inhibits proliferation by sponging MiR-211 in
colorectal cancer. Cell Physiol Biochem. 41:635–644. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Sümbül AT, Göğebakan B, Bayram S, Batmacı
CY and Öztuzcu S: MicroRNA 211 expression is upregulated and
associated with poor prognosis in colorectal cancer: A case-control
study. Tumour Biol. 36:9703–9709. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Sun Y, Byon CH, Yuan K, Chen J, Mao X,
Heath JM, Javed A, Zhang K, Anderson PG and Chen Y: Smooth muscle
cell-specific runx2 deficiency inhibits vascular calcification.
Circ Res. 111:543–552. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Atlasi Y, Noori R, Gaspar C, Franken P,
Sacchetti A, Rafati H, Mahmoudi T, Decraene C, Calin GA, Merrill BJ
and Fodde R: Wnt signaling regulates the lineage differentiation
potential of mouse embryonic stem cells through Tcf3
down-regulation. PLoS Genet. 9:e10034242013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Panizo S, Naves-Díaz M, Carrillo-López N,
Martínez-Arias L, Fernández-Martín JL, Ruiz-Torres MP,
Cannata-Andía JB and Rodríguez I: MicroRNAs 29b, 133b and 211
regulate vascular smooth muscle calcification mediated by high
phosphorus. J Am Soc Nephrol. 27:824–834. 2016. View Article : Google Scholar : PubMed/NCBI
|