Introduction
Glaucoma is a leading cause of irreversible
blindness and it has been predicted that79.6 million people
worldwide will suffer with glaucoma by 2020 (1). Primary open-angle glaucoma (POAG) is
the most prevalent type of glaucoma worldwide (1). The onset of POAG is associated with a
number of factors, including age, sex, race, myopia and diabetes
mellitus (2). However, the
underlying pathogenic mechanism remains to be elucidated. Elevated
intraocular pressure (IOP) is a major risk factor of POAG. The
elevation of IOP is caused by a low aqueous humor flow rate
resulting from lesions in the aqueous humor outflow pathways
(3). The trabecular meshwork, which
comprises endothelial-like cells, is the major location of aqueous
humor outflow (3). These trabecular
meshwork cells are critical for the maintenance of the aqueous
humor outflow pathway (4). A
previous study reported that patients with POAG have pathological
changes in the trabecular meshwork, including a reduced number of
human trabecular meshwork cells (HTMCs), an accumulation of
extracellular matrix and cytoskeletal changes (5). It has recently been demonstrated that
oxidative damage to the trabecular meshwork may serve an important
role in the development of POAG (6).
It has been reported that the expression of antioxidants, including
superoxide dismutase-1 (SOD1) and glutathione (GSH), is increased
in trabecular meshwork of patients with POAG (7).
Homocysteine (Hcy) is an intermediate product of
methionine metabolism, produced by methionine transmethylation in
the human body (8). Hcy causes
vascular injury related lesions in vascular endothelial cells
during oxidative stress and has been confirmed to be an independent
risk factor in the development of cardiovascular diseases (9). The aqueous humor is the flow of blood
from the uvea into the ciliary processes; after filtration through
reticular capillaries, this fluid is secreted by ciliary epithelial
cells (2). As such, the tissue
components of aqueous humor are similar to those of plasma, albeit
the concentrations are somewhat different. It has been reported
that, at high concentrations, Hcy may cause glaucoma due to its
oxidative stress function (10).
However, the association between Hcy and glaucoma remains
controversial (11,12). The aim of the present study was to
compare Hcy concentrations in the plasma and aqueous humor of
patients with senile cataracts and patients with POAG to
investigate the effects of Hcy on oxidative stress damage in
HTMCs.
Materials and methods
Clinical information and grouping
All patients were treated at the Second Affiliated
Hospital of Nanchang University (Nanchang, China) between January
2014 and December 2015 and provided written informed consent.
Patients were divided into the POAG group (patients with POAG) and
the control group (patients with senile cataracts). Patients with
comorbidities, including hypertension, diabetes mellitus, liver and
kidney dysfunction, cerebral infarction or thyroid dysfunction,
were excluded from the present study. In the control group,
patients with a history of cataracts, eye trauma or eye diseases
were also excluded. The POAG group comprised 41 patients (22 men,
19 women; mean age 59.20±12.8 years). The control group comprised
53 patients (30 men, 23 women; mean age 62.51±8.49 years). The
present study was performed in accordance with the Declaration of
Helsinki and was approved by the Ethics Committee of the Second
Affiliated Hospital of Nanchang University.
POAG diagnostic criteria (2)
The diagnostic criteria for POAG were as follows: i)
IOP >21 mmHg at any time; ii) fundus examination or optical
coherence tomography (OCT) examination revealing glaucomatous optic
nerve damage; iii) visual field examination revealing
characteristic visual field loss; iv) gonioscope and ultrasound
biomicroscope (UBM) examinations revealing an open anterior chamber
angle; and v) no signs of secondary glaucoma or a non-glaucomatous
cause for the optic neuropathy.
Detection of Hcy in aqueous humor and
plasma
A total of 3 ml of fasting blood was collected from
study participants and centrifuged (1,000 × g) for 15 min at room
temperature to separate the serum. Serum Hcy levels were measured
using an AU-500 automatic biochemical analyzer (Beckman Coulter,
Inc., Brea, CA, USA) according to the manufacturer's protocol. The
reference range of normal Hcy values was ≤10 mmol/l. During
trabeculectomy (POAG group) or phacoemulsification cataract surgery
(control group), the anterior chamber was punctured to aspirate
0.2–0.3 ml of the aqueous humor and Hcy levels were detected using
the AU-500 automatic biochemical analyzer.
Cell culture
Primary HTMCs (cat. no. HUM-iCell-n012) were
purchased from iCell Bioscience, Inc. (Shanghai, China). Primary
HTMCs were cultured at 37°C in Dulbecco's modified Eagle's medium
(DMEM; Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) containing
15% fetal bovine serum (FBS; Gibco; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA) in an incubator containing 5% CO2 with
100% humidity. The culture medium was replaced every 3–4 days.
HTMCs were monitored under optical microscope at a magnification of
×100 and ×400. When cell growth reached ~80% confluence at the
bottom of the culture flasks, cells were dissociated using 1–2 ml
0.25% EDTA-trypsin, passaged and continuously cultured. Cells at
the third and fourth generations were used in the experiments.
Cell proliferation/cytotoxicity assay
using Cell Counting Kit-8 (CCK-8)
HTMCs at the logarithmic growth phase were
dissociated using 0.25% EDTA-trypsin to prepare the cell
suspension. Cells were diluted using DMEM containing 15% FBS to a
density of 1×104 cell/ml, inoculated into 96-well plates
and cultured in an incubator containing 5% CO2 with 100%
humidity at 37°C. When the cell monolayer at the bottom of the
wells reached confluence, the culture medium was aspirated and
cells were washed 3 times with PBS. The culture medium was replaced
and Hcy was added at a concentration of 30, 100, 300 or 1,000
µmol/l. Vehicle wells contained cells in DMEM supplemented with 15%
FBS, while mock wells only contained DMEM supplemented with 15%
FBS. Each treatment was performed in triplicate. Cells were
continuously cultured for 48 h and washed with PBS, following which
100 µl DMEM containing 15% FBS was added to each well. A total of
10 µl CCK-8 solution (Sigma-Aldrich; Merck KGaA) was added to each
well, samples were thoroughly mixed and the cells were incubated
for 4 h at 37°C. The absorbance of each well at 450 nm was measured
using a microplate reader.
Detection of intracellular
mitochondrial membrane potential (MMP) using the JC-1 method
Following treatment, MMP detection was performed
using the JC-1 method (12). Cells
were treated with Hcy for 48 h at 37°C, washed with PBS,
dissociated with trypsin, centrifuged (1,000 × g) for 15 min at
room temperature and collected. Cells were resuspended in DMEM
supplemented with 15% FBS, mixed thoroughly with 0.5 ml JC-1
staining working solution (Abcam, Cambridge, UK) and incubated at
37°C for 20 min. Cells were then centrifuged (600 × g) at 4°C for 4
min, washed twice with ice cold JC-1 staining buffer (1X) and
resuspended in 1 ml JC-1 staining buffer (1X). The cells were
centrifuged (600 × g) at 4°C again for 4 min, resuspended in JC-1
staining buffer (1X) and loaded onto a flow cytometer for
qualitative analysis. More than 10,000 cells were observed.
Detection of intracellular reactive
oxygen species (ROS)
Cells were treated with Hcy for 48 h, dissociated
and centrifuged as described above. Cells were mixed thoroughly
with 1 ml DMEM and 1 µl DCFH-DA (Abcam) and incubated in a 37°C
incubator for 20 min. The cells were washed 3 times with serum-free
DMEM culture medium and loaded onto a flow cytometer for analysis.
More than 10,000 cells were observed.
Detection of SOD1 expression in cells
using western blotting
Cells were treated with Hcy for 48 h, dissociated
and centrifuged as described above. The cell pellet containing
5×106 cells was mixed with 0.5 ml lysis buffer (Bio-Rad
Laboratories, Inc., Hercules, CA, USA), vortexed and suspended.
Each 0.5 ml lysis buffer was mixed with 1 ml protein extraction
reagent (Bio-Rad Laboratories, Inc.) and centrifuged (10,000 × g)
at 4°C for 10 min. The protein layer was extracted, mixed with
anhydrous alcohol and centrifuged (10,000 × g) at 4°C for 3 min.
The protein pellet and protein concentration was determined using
the BCA method. Equal amounts (40 µg/lane) of protein were
separated by SDS-PAGE (10% stacking gel and 5% separating gel).
Proteins were transferred onto a polyvinylidene difluoride membrane
and incubated with the anti-SOD1 primary antibodies (cat. no.
sc-271014; Santa Cruz Biotechnology, Inc., Dallas, TX, USA) and
anti-β-actin antibodies (cat. no. ab8226; Abcam) at a dilution of
1:2,000 overnight at 4°C. β-actin expression was used to normalize
SOD1 expression. The membrane was subsequently washed and incubated
with a horseradish peroxidase-conjugated rabbit anti-mouse
Immunoglobulin G secondary antibody (cat. no. ab97046; Abcam) with
dilution of 1:2,000 for 1 h. Pierce ECL Western Blotting Substrate
(Thermo Fisher Scientific, Inc.) was then added in a dark room and
subsequent steps performed in accordance with manufacturer's
protocol. Gel image analysis was performed using UVP GelDoc-It
Imager (UVP LLC, Upland, CA, USA) and the relative densities of the
blots were measured by VisionWorks® Image Acquisition
and Analysis Software (version 7.0; UVP LLC).
Statistical analysis
Data are presented as the mean ± standard deviation
and were analyzed using SPSS 19.0 (IBM Corp., Armonk, NY, USA).
Comparisons between groups were performed using the Mann-Whitney U
test. The results of in vitro experiments were compared
using the Kruskal-Wallis test followed by a Mann-Whitney post hoc
test with Bonferroni correction. P<0.05 was considered to
indicate a statistically significant difference.
Results
Detection of Hcy concentrations in
plasma and aqueous humor of patients with POAG
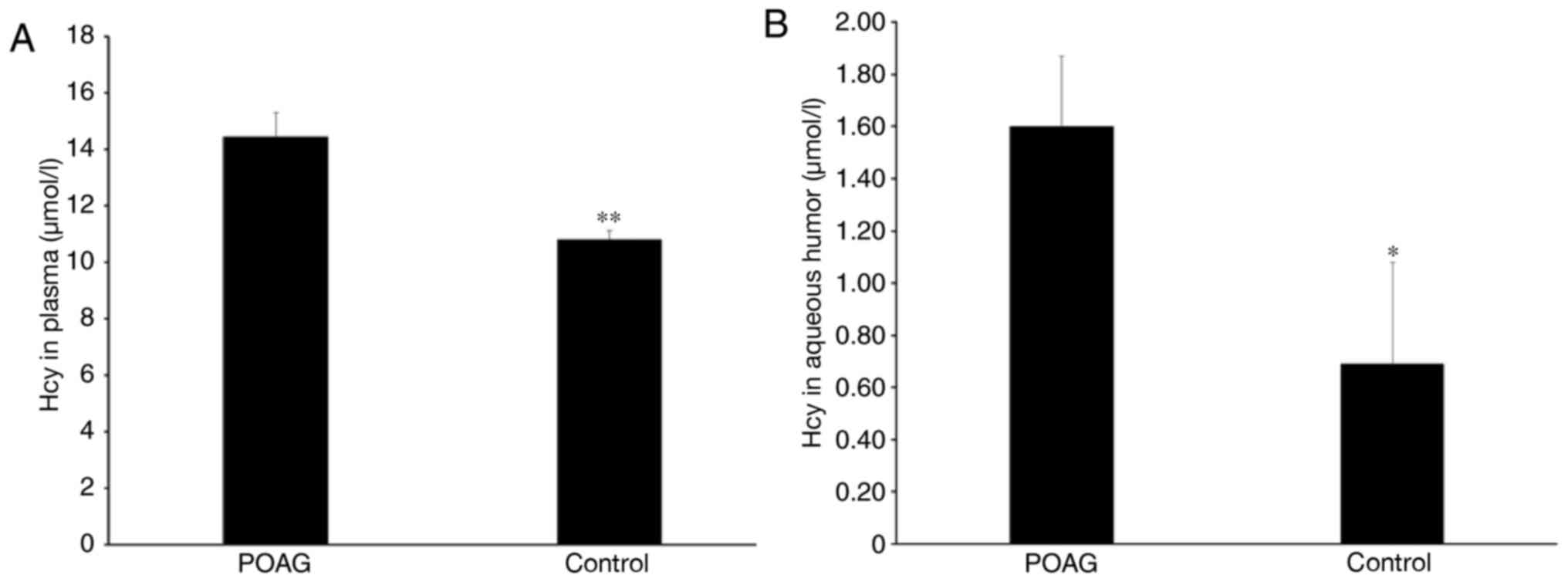
The results demonstrated that plasma Hcy
concentrations in the POAG and control groups were 14.44±0.86 and
10.82±0.29 µmol/l, respectively. Plasma Hcy was significantly
increased in the POAG group compared with patients with senile
cataracts (P<0.01; Fig. 1A). In
the aqueous humor, plasma concentrations were 1.60±0.27 and
0.69±0.39 µmol/l in the POAG and control groups, respectively.
Compared with the control group, Hcy was significantly upregulated
in the aqueous humor of patients with POAG (P<0.05; Fig. 1B).
Growth characteristics of HTMCs and
cell proliferation/cytotoxicity assay
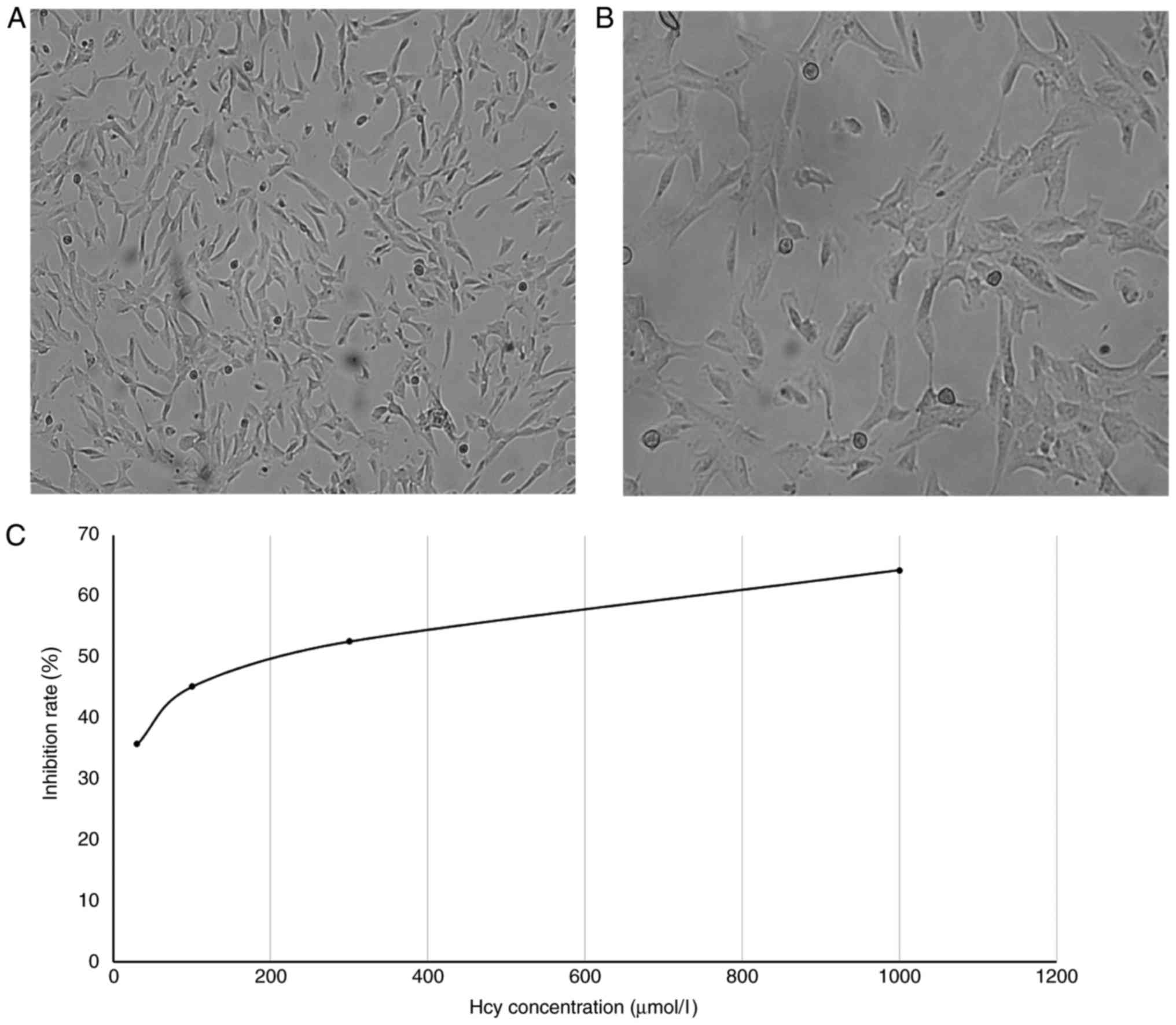
Normal HTMCs were generally wide, large and flat,
with many processes. A number of morphologies, including
star-shaped, spindle-shaped and polygonal-shaped, were observed.
Cells overlapped, the cytoplasm was clear and bright and nuclei
were large and round or oval-shaped (Fig. 2A and B). During passaging, the cells
grew faster during the third and fourth generations and began to
become adherent at 10–12 h. Cells were completely attached within
24 h. Generally, cells completely fused to form a closely connected
cell monolayer within 1 week. After the 6th generation, the cell
growth rate decreased and the cells were spindle-shaped.
Furthermore, the cytoplasm-to-nucleus ratio decreased, pigment
granule deposition increased, growth speed decreased and the cells
were not readily adherent. Therefore, the third to fifth generation
of HTMCs were used in the present study.
The cell viability in control wells was considered
to be 100% and cell inhibition rates in the other groups were
calculated using the following formula: [optical density (OD) of
vehicle wells-OD of experimental wells)/(OD of vehicle wells-OD of
mock wells) × 100. The results revealed that cell viability was
significantly decreased in all wells treated with Hcy compared with
the vehicle (P<0.01; Table I and
Fig. 2C). Hcy appeared to inhibit
cell proliferation, with higher Hcy concentrations inducing higher
inhibition rates.
 | Table I.Effects of Hcy on human trabecular
meshwork cells proliferation as assessed using Cell Counting
Kit-8. |
Table I.
Effects of Hcy on human trabecular
meshwork cells proliferation as assessed using Cell Counting
Kit-8.
| Group | Absorbance | Inhibition rate
(%) |
|---|
| Mock | 0 | 0 |
| Vehicle | 1.02±0.0226 | 0 |
| Hcy (µmol/l) |
|
|
| 30 |
0.6988±0.0078a | 35.74 |
| 100 |
0.6137±0.0431a | 45.20 |
| 300 |
0.5472±0.0282a | 52.60 |
|
1,000 |
0.442±0.0516a | 64.31 |
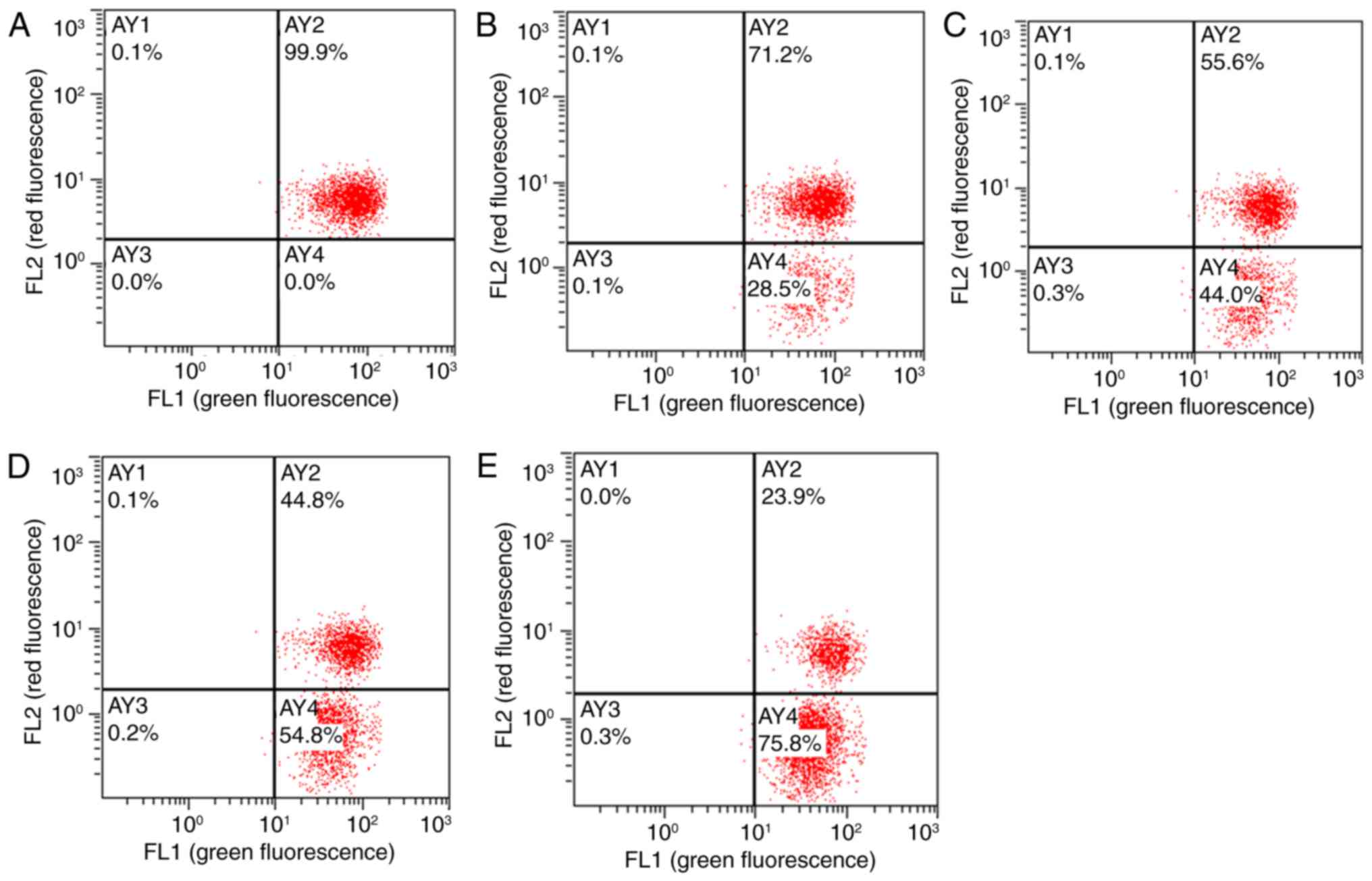
Detection of MMP in HTMCs
The JC-1 fluorescent probe, which has an excitation
wavelength of 488 nm and a monomer emission wavelength of 525 nm,
is able to enter cells and localize to the mitochondrial membrane
(13). Depending on changes in the
MMP, the probe emits different colors of fluorescence. In cells
with higher MMPs, JC-1 forms an aggregate in the mitochondrial
matrix and produces red fluorescence. At lower MMPs, the probe
produces green fluorescence. The results of flow cytometry revealed
good HTMC activity and normal MMPs in vehicle wells (Fig. 3); in these cells, JC-1 accumulated in
the AY2 region and exhibited strong red fluorescence. Compared with
the vehicle group, JC-1 accumulation in the AY2 area decreased in
HTMCs following treatment with Hcy for 48 h; these changes occurred
in 71.2, 55.6, 44.8 and 23.9% of total cells in the 30, 100, 300
and 1,000 µmol/l Hcy group, respectively. These results indicated
that the concentration of JC-1 aggregates decreased and the MMPs
decreased.
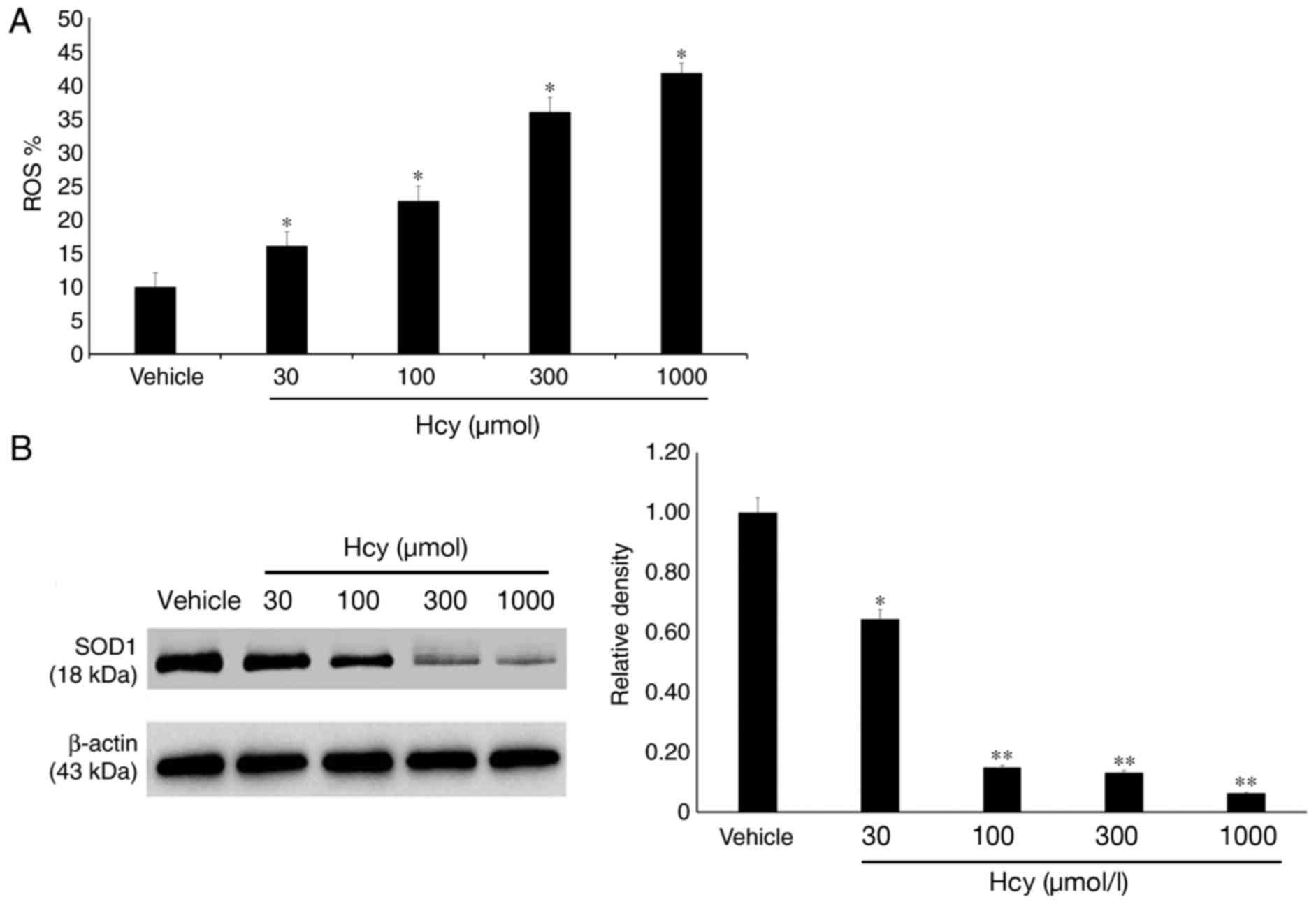
Detection of oxidative stress in
HTMCs
ROS were detected using flow cytometry and the
results demonstrated that the ROS levels in groups treated with
30–1,000 µmol/l Hcy significantly increased compared with the
vehicle group (P<0.05; Fig.
4A).
Furthermore, the expression of the oxidative stress
marker SOD1 in HTMCs was detected following treatment with
different concentrations of Hcy. The results revealed that SOD1
expression in HTMCS decreased following Hcy treatment in a
dose-dependent manner (Fig. 4B;
P<0.05).
Discussion
A major risk factor of glaucoma is pathological
elevation of IOP (2). Lesions in
aqueous humor outflow pathways may obstruct the outflow of the
aqueous humor, thereby elevating IOP (3). The trabecular meshwork is a major
location of aqueous humor outflow (3). Oxidative degeneration of the trabecular
meshwork damages the aqueous humor outflow pathways, resulting in
insufficient ocular local blood transfusion and, in particular,
promoting the degeneration of endothelial cells (6). Early glaucoma is characterized by
endogenous cell loss, a reduction of antioxidant levels in the
aqueous humor, HTMC apoptosis caused by mitochondrial damage and
endothelial cell dysfunction in the anterior chamber (14). A previous in vitro study
reported that ROS are able to induce structural changes in the
cytoskeleton of trabecular meshwork cells and the extracellular
matrix or increase the expression of adhesion molecules, thereby
affecting outflow of the aqueous humor and further increasing IOP
(15). In addition, it has been
reported that all indicators in the aqueous humor, including the
total antioxidant capacity, total oxidation value and oxidative
stress index, are significantly increased in patients with POAG
compared with those with senile cataracts (16).
Hcy is an intermediate product of the methionine
cycle pathway and contains a thiol group. When plasma Hcy
concentration >10 µmol/l, Hcy may be self-oxidized to produce
oxygen free radicals and peroxides, which in turn damage the
function and structure of vascular endothelial cells. The results
of the present study demonstrated that Hcy concentrations in the
plasma and aqueous humor of patients with POAG were significantly
higher compared with the control group, suggesting that Hcy is
upregulated in patients with POAG. These results were similar to
those of Ghanem et al (17).
However, other studies reported that there were no statistical
differences in Hcy levels of patients with glaucoma and healthy
subjects (18,19). The authors hypothesized that this
discrepancy may be attributed to ethnic factor since large
differences existed in the prevalence and phenotype of POAG among
different ethnic groups (20,21).
Plasma Hcy levels in patients with senile cataracts in the present
study were also slightly higher than 10 µmol/l. This result may be
associated with a number of factors, including age, a vegetarian
diet, Helicobacter pylori infection or increased incidence
of atrophic gastritis, all of which may cause malabsorption of
coenzymes, including folic acid, VB12 and VB6, which are required
for the metabolism of Hcy (22).
In the present study it was also investigated
whether treating the trabecular meshwork with high concentrations
of Hcy was able to cause structural and functional damage to HTMCs.
For healthy adults, Hcy concentrations >15 µmol/l are considered
hyperhomocysteinemia (23).
Therefore, based on a study by Perła-Kaján et al (23), an initial Hcy concentration of 30
µmol/l was used in this study. The results demonstrated that cell
proliferation was significantly inhibited following Hcy treatment.
The inhibition of HTMC proliferation increased in a dose-dependent
manner. It has previously been reported that HTMCs treated with 40%
O2 exhibited decreased activity after 10 days (24). This suggests that treating HTMCs with
Hcy as an oxidizing agent may induce apoptosis and inhibit cell
proliferation.
The JC-1 method was used to evaluate the reduction
of MMP, which is used as an early marker of apoptosis (25). The JC-1 fluorescent probe enters
cells and accumulates in the mitochondrial matrix to form JC-1
aggregates, which exhibit red fluorescence (26). If the MMP decreases, JC-1 is unable
to aggregate in the mitochondrial matrix and remains a monomer that
exhibits green fluorescence (26).
In the present study, untreated HTMCs exhibited red fluorescent
JC-1, whereas cells treated with different concentrations of Hcy
exhibited green fluorescent JC-1 that increased in intensity at
higher Hcy concentrations. These results indicate that the MMP
decreased. It was therefore speculated that Hcy induced the
reduction of MMP, thereby damaging cell metabolism and inhibiting
HTMC proliferation.
Flow cytometry and immunoblotting were performed to
measure ROS levels and the expression of SOD1, which is primarily
responsible for O2-clearance (27). As mentioned previously, Hcy treatment
increased ROS production in HTMCs. Further investigated revealed
that SOD1 expression in HTMCs was downregulated and was
significantly negatively correlated with Hcy concentration; these
results indicate that Hcy inhibits SOD1 expression. Koh et
al (28) used mouse bone marrow
cells to demonstrate that the production of ROS increases following
Hcy stimulation, while Kang et al (29) reported that increased plasma Hcy
concentrations and reduced SOD1 levels were observed in young
patients with central retinal vein occlusion. Based on these
findings, it was speculated that Hcy may directly induce oxidative
stress in HTMCs to cause the excessive production of ROS and
promote cell apoptosis and also aggravate the oxidative damage by
inhibiting the expression of intracellular antioxidant enzymes.
In summary, the results of the present study
demonstrate that plasma and aqueous humor concentrations of Hcy
increase in patients with POAG and that elevated Hcy concentrations
may be a risk factor of POAG. Excessively elevated Hcy
concentrations induce oxidative stress in HTMCs, increase the
production of ROS, downregulate the expression of antioxidant
enzymes (including SOD1) and reduce MMPs to further inhibit HTMC
proliferation. These deleterious effects were also dose-dependent.
Hcy may therefore be associated with the onset of POAG. The
concentration of SOD1 was used as the oxidative stress marker; as
the concentration of SOD1 was not high and the underlying mechanism
was not elucidated, there were some limitations in the present
study. Future studies should investigate the molecular mechanism,
ion channels expression and signaling pathways associated with
Hcy-induced oxidative stress in HTMCs.
Acknowledgements
Not applicable.
Funding
The present study was supported by the National
Natural Science Foundation of China (grant nos. 81460088 and
81760176), the Jiangxi Provincial Training Program for
Distinguished Young Scholars (grant no. 20171BCB23092), the Jiangxi
Provincial Key R&D Program (grant no. 20171BBG70099), the
Jiangxi Provincial Natural Science Foundation for Youth Scientific
Research (grant no. 20171BAB215032) and the Scientific Research
Program of Jiangxi Health and Family Planning Commission (grant no.
20171076).
Availability of data and materials
The datasets used or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
ZPY and KS designed the study and performed the
experiments. YZZ and LS performed the experiments. YLZ analyzed the
data and KS wrote the manuscript.
Ethics approval and consent to
participate
All patients provided written informed consent. The
present study was performed in accordance with the Declaration of
Helsinki and was approved by the Ethics Committee of the Second
Affiliated Hospital of Nanchang University.
Consent for publication
The patients provided written informed consent for
publication.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
Hcy
|
homocysteine
|
|
HTMCs
|
human trabecular meshwork cells
|
|
POAG
|
primary open-angle glaucoma
|
|
IOP
|
intraocular pressure
|
|
SOD1
|
superoxide dismutase-1
|
|
GSH
|
glutathione
|
|
MMP
|
mitochondrial membrane potential
|
References
|
1
|
Quigley HA and Broman AT: The number of
people with glaucoma worldwide in 2010 and 2020. Br J Ophthalmol.
90:262–267. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kanski J and Bowling B: Clinical
Ophthalmology: A systematic approach. 7th edition. Elsevier;
London: pp. 3311–3331. 2011
|
|
3
|
Xin C, Tian N, Li M, Wang H and Wang N:
Mechanism of the reconstruction of aqueous outflow drainage. Sci
China Life Sci. Dec 22–2017.(Epub Ahead of Print). View Article : Google Scholar
|
|
4
|
Saccà SC, Gandolfi S, Bagnis A, Manni G,
Damonte G, Traverso CE and Izzotti A: The outflow pathway: A tissue
with morphological and functional unity. J Cell Physiol.
231:1876–1893. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Chrysostomou V, Rezania F, Trounce IA and
Crowston JG: Oxidative stress and mitochondrial dysfunction in
glaucoma. Curr Opin Pharmacol. 13:12–15. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhao J, Wang S, Zhong W, Yang B, Sun L and
Zheng Y: Oxidative stress in the trabecular meshwork (Review). Int
J Mol Med. 38:995–1002. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ghanem AA, Arafa LF and El-Baz A:
Oxidative stress markers in patients with primary open-angle
glaucoma. Curr Eye Res. 35:295–301. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Long Y and Nie J: Homocysteine in Renal
Injury. Kidney Dis (Basel). 2:80–87. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Sanz J, Moreno PR and Fuster V: The year
in atherothrombosis. J Am Coll Cardiol. 62:1131–1143. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Bleich S, Jünemann A, von Ahsen N, Lausen
B, Ritter K, Beck G, Naumann GO and Kornhuber J: Homocysteine and
risk of open-angle glaucoma. J Neural Transm (Vienna).
109:1499–1504. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Xu F, Zhao X, Zeng SM, Li L, Zhong HB and
Li M: Homocysteine, B vitamins, methylenetetrahydrofolate reductase
gene, and risk of primary open-angle glaucoma: A meta-analysis.
Ophthalmology. 119:2493–2499. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Goyal A, Srivastava A, Sihota R and Kaur
J: Evaluation of oxidative stress markers in aqueous humor of
primary open angle glaucoma and primary angle closure glaucoma
patients. Curr Eye Res. 39:823–829. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Perelman A, Wachtel C, Cohen M, Haupt S,
Shapiro H and Tzur A: JC-1: Alternative excitation wavelengths
facilitate mitochondrial membrane potential cytometry. Cell Death
Dis. 3:e4302012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kruk J, Kubasik-Kladna K and Aboul-Enein
HY: The role oxidative stress in the pathogenesis of eye diseases:
Current status and a dual role of physical activity. Mini Rev Med
Chem. 16:241–257. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Babizhayev MA: Biomarkers and special
features of oxidative stress in the anterior segment of the eye
linked to lens cataract and the trabecular meshwork injury in
primary open-angle glaucoma: Challenges of dual combination therapy
with N-acetylcarnosine lubricant eye drops and oral formulation of
nonhydrolyzed carnosine. Fundam Clin Pharmacol. 26:86–117. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ergan E, Ozturk F, Beyazyildiz E, Elgin U,
Sen E, Cankaya AB and Celik T: Oxidant/antioxidant balance in the
aqueous humor of patients with glaucoma. Int J Ophthalmol.
9:249–252. 2016.PubMed/NCBI
|
|
17
|
Ghanem AA, Mady SM, El awady HE and Arafa
LF: Homocysteine and hydroxyproline levels in patients with primary
open-angle glaucoma. Curr Eye Res. 37:712–718. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Leibovitzh H, Cohen E, Levi A, Kramer M,
Shochat T, Goldberg E and Krause I: Relationship between
homocysteine and intraocular pressure in men and women: A
population-based study. Medicine (Baltimore). 95:e48582016.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zacharaki F, Hadjigeorgiou GM, Koliakos
GG, Morrison MA, Tsezou A, Chatzoulis DZ, Almpanidou P, Topouridou
K, Karabatsas CH, Pefkianaki M, et al: Plasma homocysteine and
genetic variants of homocysteine metabolism enzymes in patients
from central Greece with primary open-angle glaucoma and
pseudoexfoliation glaucoma. Clin Ophthalmol. 8:1819–1825. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kim E and Varma R: Glaucoma in
latinos/hispanics. Curr Opin Ophthalmol. 21:100–105. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Racette L, Wilson MR, Zangwill LM, Weinreb
RN and Sample PA: Primary open-angle glaucoma in blacks: A review.
Surv Ophthalmol. 48:295–313. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wells JL and Dumbrell AC: Nutrition and
aging: Assessment and treatment of compromised nutritional status
in frail elderly patients. Clin Interv Aging. 1:67–79. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Perła-Kaján J, Twardowski T and Jakubowski
H: Mechanisms of homocysteine toxicity in humans. Amino Acids.
32:561–572. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Caballero M, Liton PB, Epstein DL and
Gonzalez P: Proteasome inhibition by chronic oxidative stress in
human trabecular meshwork cells. Biochem Biophys Res Commun.
308:346–352. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
He Y, Ge J and Tombran-Tink J:
Mitochondrial defects and dysfunction in calcium regulation in
glaucomatous trabecular meshwork cells. Invest Ophthalmol Vis Sci.
49:4912–4922. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Sung DK, Chang YS, Kang S, Song HY, Park
WS and Lee BH: Comparative evaluation of hypoxic-ischemic brain
injury by flow cytometric analysis of mitochondrial membrane
potential with JC-1 in neonatal rats. J Neurosci Methods.
193:232–238. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Izzotti A, Longobardi M, Cartiglia C and
Saccà SC: Mitochondrial damage in the trabecular meshwork occurs
only in primary open-angle glaucoma and in pseudoexfoliative
glaucoma. PLoS One. 6:e145672011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Koh JM, Lee YS, Kim YS, Kim DJ, Kim HH,
Park JY, Lee KU and Kim GS: Homocysteine enhances bone resorption
by stimulation of osteoclast formation and activity through
increased intracellular ROS generation. J Bone Miner Res.
21:1003–1011. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Kang SW, Chang TS, Lee TH, Kim ES, Yu DY
and Rhee SG: Cytosolic peroxiredoxin attenuates the activation of
Jnk and p38 but potentiates that of Erk in Hela cells stimulated
with tumor necrosis factor-alpha. J Biol Chem. 279:2535–2543. 2004.
View Article : Google Scholar : PubMed/NCBI
|