Introduction
Colorectal cancer (CRC) is one of the most prevalent
and malignant cancers in the world, with more than one million new
diagnosed patients each year (1).
CRC contributes to a lot of cancer-related deaths. CRC progression
is accompanied with accumulation of mutations, which lead to
inactivation of tumor suppressors or activation of oncogenes
(2). Especially, activation of
Wnt/β-catenin signaling pathway is observed in almost 90% of
patients diagnosed with CRC (3,4).
Although much effort has been made, no effective therapeutic
approach has been developed to fully treat CRC. The five-year
survival rate of CRC patients remains quite poor (5). Therefore, there is an urgent need to
explore the underlying mechanism of CRC progression.
MicroRNAs (miRNAs/miRs) are a class of naturally
short noncoding RNAs with a length of 19–25 nucleotides and largely
expressed in almost all cell types (6). miRNAs have been demonstrated to
suppress gene expression by association with the specific sequence
in the 3′-untranslated region (3′-UTR) of target mRNAs (7,8). More
and more evidences show that miRNAs exert important functions in a
diversity of biological processes, and influence cell fate and
functions (9–11). So far, a large amount of miRNAs have
been shown to serve as tumor suppressors or oncogenes in almost all
types of cancers, including lung cancer, breast cancer, gastric
cancer and CRC (12–16). For example, microRNA-381 inhibits the
metastasis of gastric cancer by targeting TMEM16A expression
(17). Downregulation of miR-218
contributes to epithelial-mesenchymal transition and tumor
metastasis in lung cancer by targeting Slug/ZEB2 signaling
(18). However, the roles of most
miRNAs in CRC still remain largely unknown.
miR-6852 has been shown to induce cell cycle arrest
and necrosis in cervical cancer cells (19). Whether miR-6852 regulates CRC
progression requires to be investigated. In this study, we found
that miR-6852 was downregulated in CRC tissues and correlated with
tumor metastasis and patients' prognosis. Moreover, we showed that
overexpression of miR-6852 significantly inhibited the
proliferation and invasion of CRC cells. Mechanistically, we found
that TCF7 is a direct target of miR-6852. We also demonstrated that
TCF7 was upregulated in CRC tissues, and promoted CRC cell
proliferation and invasion. Taken together, our study demonstrated
that miR-6852 acts as a tumor suppressor in CRC through targeting
TCF7.
Materials and methods
Patient samples
The Ethics Committee of Harbin Medical University
Cancer Hospital (Harbin, China) approved this study, and all
patients gave their informed consent prior to surgery. Forty-four
CRC tissues and 44 adjacent normal tissues from human patients were
obtained from Harbin Medical University Cancer Hospital. The
clinicopathological data of the patients were listed in Table I. For mRNA extraction, samples were
frozen in liquid nitrogen immediately after surgical removal and
maintained at−80°C until use. Additional samples were fixed in 10%
neutral-buffered formalin overnight, processed, paraffin embedded,
and sectioned.
 | Table I.Associations between miR-6852
expression and clinical characteristics in CRC patients (n=44). |
Table I.
Associations between miR-6852
expression and clinical characteristics in CRC patients (n=44).
| Clinicopathologic
parameters | Low expression
(n=22) | High expression
(n=22) | P-value |
|---|
| Age (years) |
|
| 0.5365 |
| ≤65 | 10 | 7 | |
|
>65 | 12 | 15 |
|
| Sex |
|
| 0.5434 |
| Male | 14 | 11 |
|
|
Female | 8 | 11 |
|
| Tumor size
(cm) |
|
| 0.0268 |
| ≤5 | 4 | 12 |
|
|
>5 | 18 | 10 |
|
| Lymph node
metastasis |
|
| 0.0122 |
|
Yes | 13 | 4 |
|
| No | 9 | 18 |
|
| TNM stage |
|
| 0.0329 |
|
I–II | 8 | 16 |
|
|
III–IV | 14 | 6 |
|
Cell lines and cell culture
Human CRC cancer cell lines (SW480, HCT8, HT29, LOVO
and HCT116) and normal colorectal mucosa cells cell line (FHC) were
obtained from ATCC (Manassas, VA, USA) and maintained in DMEM
medium supplemented with 10% fetal bovine serum (FBS; Invitrogen;
Thermo Fisher Scientific, Inc., Waltham, MA, USA), 100 U/ml
penicillin, and 100 mg/ml streptomycin (Invitrogen; Thermo Fisher
Scientific, Inc.). Cells were incubated at 37°C in a humidified
atmosphere with 5% CO2.
Cell transfection
siRNA against TCF7 (5′-GGAAGAGAGGACAAGGAAT-3′) and
miR-6852 mimics (5′-CCCUGGGGUUCUGAGGACAUG-3′), inhibitors
(5′-CATGTCCTCAGAACCCCAGGG-3′) or controls
(5′-UCACAACCUCCUAGAAAGAGUAGA-3′) were synthetized by Invitrogen
(Thermo Fisher Scientific, Inc.). 50 nM miR-6852 mimics, inhibitors
or controls were transduced into HCT8 and SW480 cells with
Lipofectamine 3000 (Invitrogen; Thermo Fisher Scientific, Inc.) at
20°C. 48 h after transfection, the culture medium was replaced with
fresh medium, followed by analysis.
Cell proliferation
Each well of a 96-well plate contained approximately
5,000 transfected cells. Cell viability was assessed using a Cell
Counting Kit-8 (CCK-8) assay (7 Sea Biotech, Shanghai, China).
Following incubation for 72 h, 10 µl CCK-8 was added to each well
and incubated for 2 h in an incubator. The absorbance value was
determined using a multimode microplate reader (Berthold
Technologies GmbH & Co. KG, Bad Wildbad Germany) at 450 nm.
Flow cytometry
Cells transiently transfected with the indicated
plasmids were harvested 48 h after transfection by trypsinization,
washed with ice-cold phosphate-buffered saline, and fixed with 70%
ethanol overnight. The cells were then collected by centrifugation
and resuspended in PI solution [50 mg/ml in phosphate-buffered
saline (PBS)] containing 0.25 mg/ml of RNase A. After incubation
for 15 min in the dark at 4°C, the cells were analyzed by flow
cytometry (FACS Canto II; BD Biosciences, Franklin Lakes, NJ, USA)
using an instrument equipped with Cell Quest software (BD
Biosciences). The percentages of the cells in S phase were counted
and compared.
Transwell assay
Before experiment, the Matrigel was melted at 4°C
and diluted with serum-free culture medium (1:3). On the surface of
polycarbonate membrane, 40 µl diluted matrigel was spread and
placed in the incubator for 4 h of coagulation for later use. After
cells in logarithmic phase were starved in the serum-free DMEM
media for 24 h, cells were digested using 0.25% ethylene diamine
tetraacetic acid (EDTA) trypsin for preparation of single-cell
suspension using the serum-free DMEM culture medium, in which cell
density was adjusted to 4×105/ml. In the upper transwell
chamber, 200 µl serum-free cell suspension was added, and the wells
were grouped according to the experiment requirement with 3
replicative wells in each group. In the lower transwell chamber,
600 µl DMEM medium supplemented with 10% fetal bovine serum were
added into each well for 24 h of culture in an incubator. The
chambers were taken out and washed with PBS to remove the medium.
In the upper chamber, cells that failed to pass through the
membrane were scrubbed using a wet cotton swab followed by 20 min
of fixation in methanol drying at room temperature. Furthermore,
cells were stained using crystal violet for 20 min, and placed
under the inverted microscope to observe the quantity of cells that
passed through the membrane. Cell count was performed in 5 high
magnification vision (×400), and the average cell count was used as
the result.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNAs were extracted with TRIzol according to
the manufacturer's protocol. cDNA was synthesized with the M-MLV
reverse transcriptase (Promega Corporation, Madison, WI, USA).
Transcripts were analyzed with ABI 7300 qPCR system using specific
primer pairs. Relative expressions were calculated and normalized
to endogenous Actb or U6. Expressions were normalized to endogenous
controls and calculated using relative quantification method
(2−ΔΔCq) (20). The
primer sequences are as follows: Universal miRNA primer
(5′-AACGAGACGACGACAGAC-3′), U6 (5′-GCAAATTCGTGAAGCGTTCCATA-3′),
miR-6852 (5′-CCCTGGGGTTCTGAGGACATG-3′), Actb (forward,
5′-CGGCGCCCTATAAAACCCA-3′; and reverse, 5′-GAGGCGTACAGGGATAGCAC-3′)
and TCF7 (forward, 5′-CTGGGCAAGGAAGCCATAGG-3′; reverse,
5′-TGCTGTACCTGTGTGCTCTG-3′).
Western blot analysis
After transfection, CRCs total protein was extracted
by RIPA buffer (Beyotime, Beijing, China). All the protein lysates
were separated using 10% SDS-PAGE and transferred onto a
polyvinylidene fluoride membrane. The membrane was incubated with
specific primary anti-human antibodies including TCF7 (1:1,000,
cat. no. 2203; Cell Signaling Technology, Inc., Danvers, MA, USA)
and GAPDH (1:5,000; Abcam, Cambridge, UK), followed by their
respective appropriate secondary antibody (1:5,000; Biogot
Technology, Nanjing, China). GAPDH was used as a control.
Luciferase reporter assay
The fragment of TCF7 3′-UTR containing putative
miR-6852 binding sites or the mutant fragment was synthesized and
inserted into the pmirGLO vector (Promega Corporation). The cells
were co-transfected with these plasmids with miR-6852 mimic or
miR-6852 mimic control. The Dual-Luciferase Reporter Assay System
(Promega Corporation) was used to measure luciferase activity.
Renilla luciferase activity was used as the normalized control.
Statistical analysis
All statistical analyses were performed using the
Statistical Package for the Social Sciences version 20.0 software
(SPSS, Inc., Chicago, IL, USA). Survival curves were calculated
using the Kaplan-Meier method and were analyzed using the log-rank
test. For comparisons, one-way analysis of variance followed by
Tukey's post hoc test and two-tailed Student's t-tests were
performed, as appropriate. The correlations were analyzed using
Pearson's correlation coefficients. The Pearson's Chi-square test
was used to assess the association between clinicopathological
features and miR-6852. P<0.05 was considered to indicate a
statistically significant difference.
Results
Expression of miR-6852 in CRC tissues
and cell lines
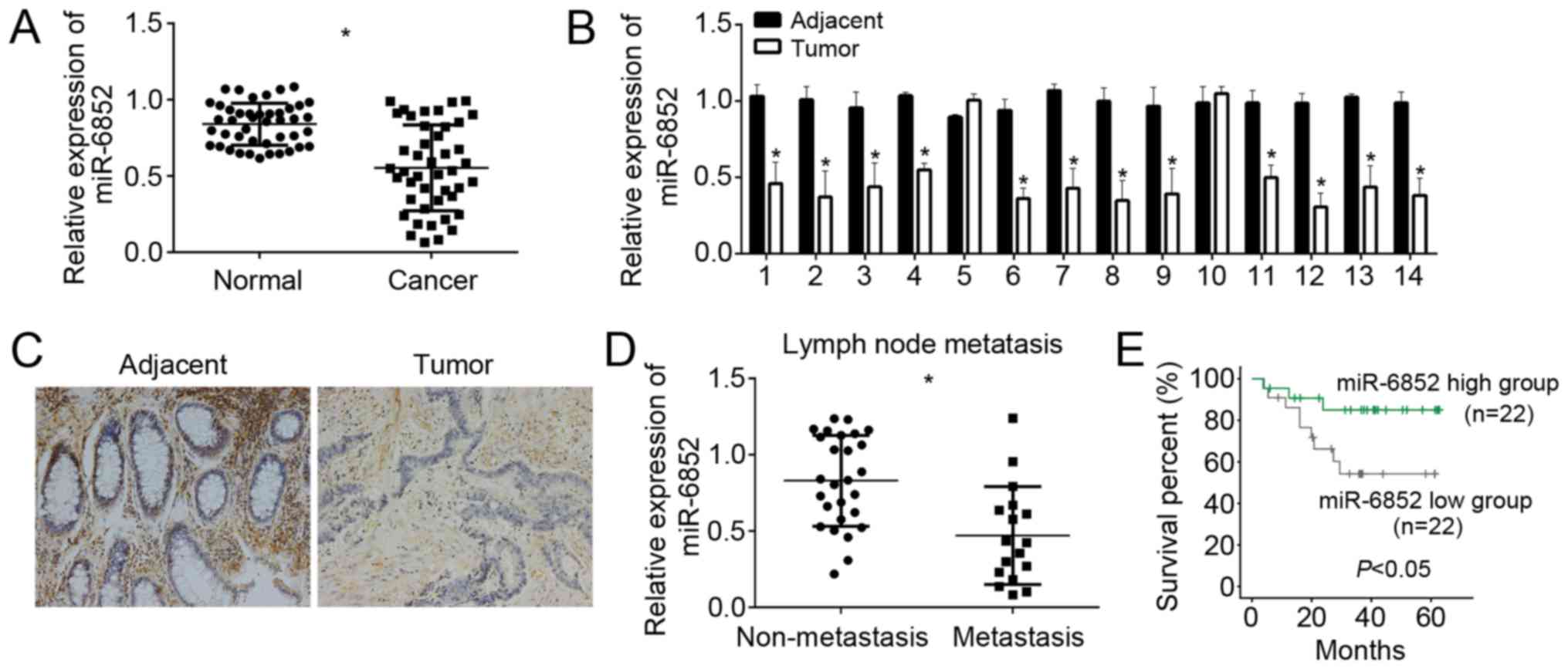
To explore the role of miR-6852 in CRC, we analyzed
the expression patterns of miR-6852 in CRC tissues and adjacent
normal tissues by RT-qPCR. The results showed that miR-6852 was
downregulated in CRC tissues (n=44) compared with adjacent normal
tissues (n=44) (Fig. 1A). We also
checked the expression of miR-6852 in 14 pairs of CRC tissues and
normal tissues and found that miR-6852 was significantly
downregulated in most CRC tissues compared to matched adjacent
normal tissues (Fig. 1B).
Furthermore, we verified miR-6852 expression by RNA in situ
hybridization. We found that miR-6852 was remarkably downregulated
in CRC tissues compared with matched normal tissues (Fig. 1C). We determined the correlation
between miR-6852 expression and CRC metastasis. Through RT-qPCR
analysis, we found that miR-6852 displayed lower expression in CRC
samples with lymph node metastasis (n=17) than that without lymph
node metastasis (n=27) (Fig. 1D). To
determine whether miR-6852 could serve as a prognostic biomarker
for CRC patients, we performed Kaplan-Meier survival curse analysis
according to miR-6852 expression (mean value was chosen for
cut-off) in collected 44 CRC samples. We divided these samples into
miR-6852 low expression group (n=22) and high expression group
(n=22). The results indicated that CRC patients with lower
expression of miR-6852 showed shorter survival rate (Fig. 1E), which indicated that miR-6852 was
a good prognostic biomarker.
miR-6852 overexpression suppresses CRC
cell proliferation and invasion
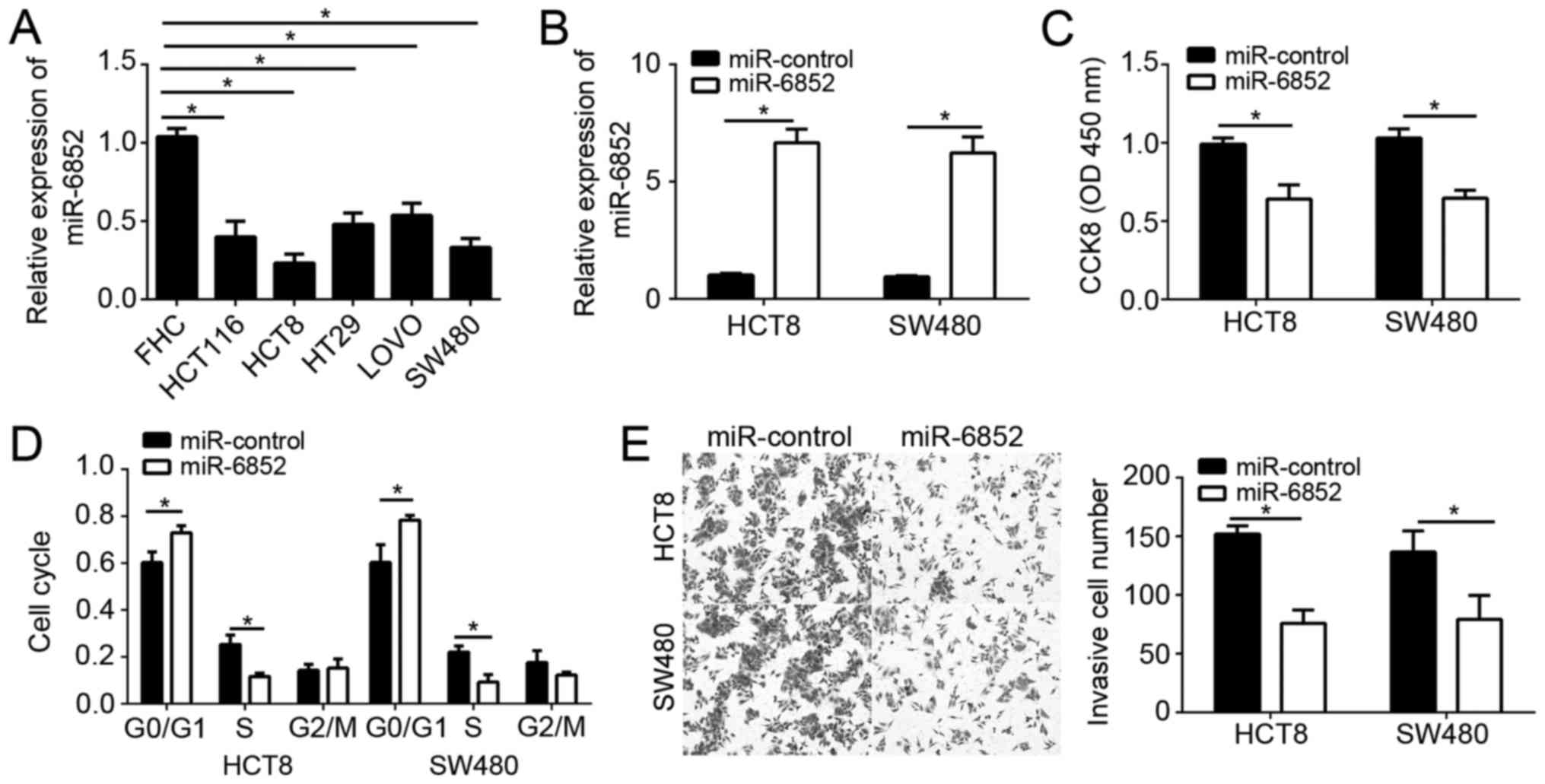
To further investigate the effects of miR-6852 on
CRC cells, we first analyzed its expression patterns in CRC cell
lines by RT-qPCR. Results indicated that miR-6852 was also
downregulated in CRC cell lines compared to FHC normal cells
(Fig. 2A). We chose HCT8 and SW480
cells for following experiments. We overexpressed miR-6852 by
transfection with miR-6852 mimics in HCT8 and SW480 cells. RT-qPCR
results showed that miR-6852 was significantly upregulated in HCT8
and SW480 cells transfected with miR-6852 mimics compared to
controls (Fig. 2B). CCK-8 assay were
used to analyze cell proliferation. As shown, overexpression of
miR-6852 significantly inhibited the proliferation of HCT8 and
SW480 cells (Fig. 2C). What's more,
FACS assay demonstrated that overexpression of miR-6852
significantly reduced the cells in S phase (Fig. 2D), indicating a decreased cell-cycle.
Metastasis leads to CRC recurrence and malignance. Therefore, we
analyzed the effect of miR-6852 on cell invasion. Transwell assays
indicated that overexpression of miR-6852 significantly suppressed
the invasion in HCT8 and SW480 cells (Fig. 2E). Taken together, these data
demonstrated that miR-6852 inhibited CRC cell proliferation and
invasion.
TCF7 is a target of miR-6852
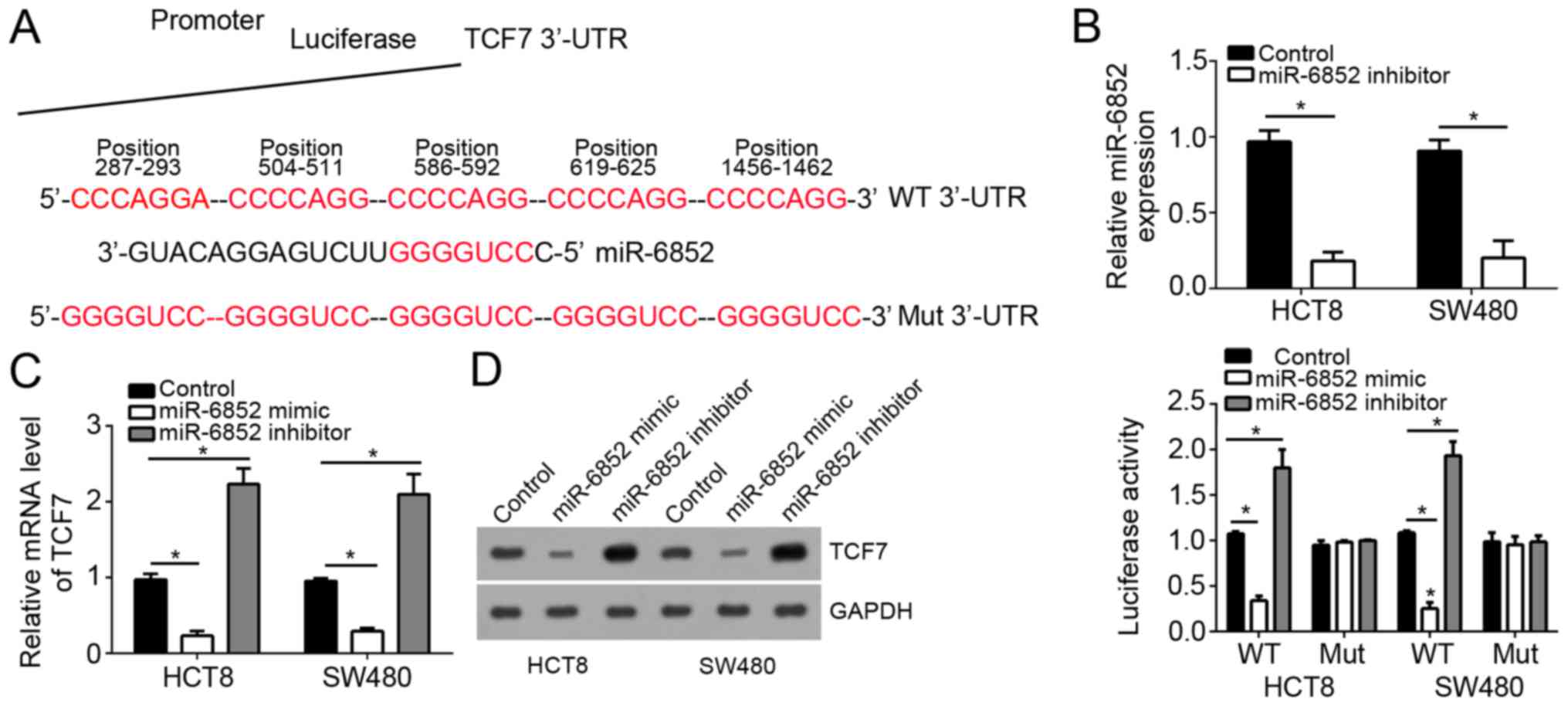
To further explore the downstream mechanism, we
searched for the target genes of miR-6852. By the targetscan tool,
we found that TCF7 was a potential target of miR-6852. TCF7 is a
key regulator of Wnt/β-catenin signaling pathway, which is
abnormally activated in most CRC patients. Therefore, we chose TCF7
for next analysis. We found that there were five potential binding
sites of miR-6852 in the 3′-UTR of TCF7 mRNA (Fig. 3A). To verify it, we constructed
luciferase reporter vectors. We mutated the potential binding sites
in the 3′-UTR of TCF7 mRNA to construct a mutant reporter plasmid.
Through luciferase reporter assays, we found that overexpression of
miR-6852 significantly inhibited the luciferase activity in HCT8
and SW480 cells transduced with WT reporter vector and vice versa,
whereas transfection with mutant reporter vector had no this effect
(Fig. 3B). Moreover, by RT-qPCR and
western blot, we found that overexpression of miR-6852
significantly inhibited the mRNA and protein levels of TCF7 in HCT8
and SW480 cells, and vice versa (Fig. 3C
and D).
TCF7 is upregulated in CRC
tissues
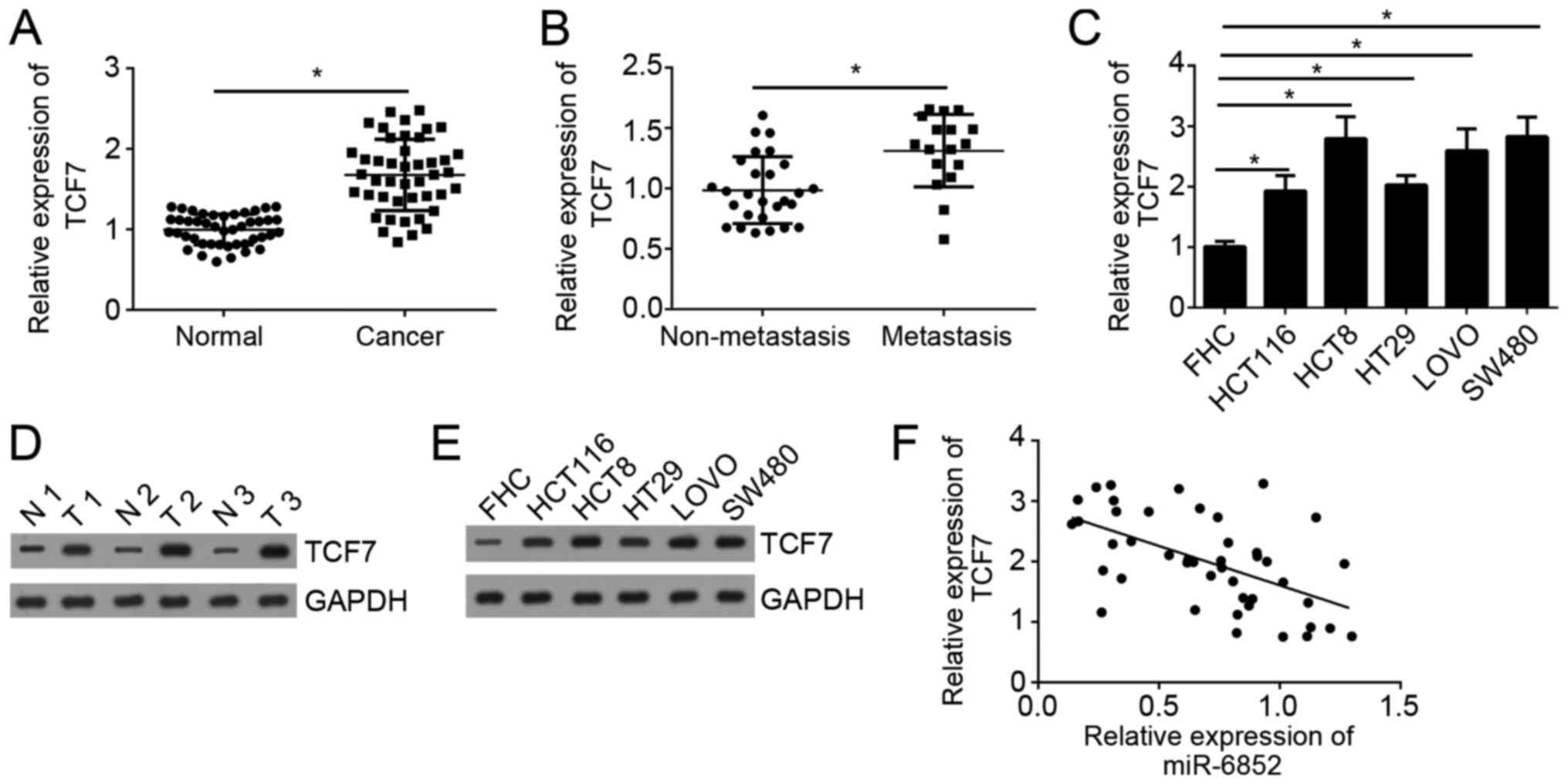
We analyzed the expression patterns of TCF7 in CRC
tissues by western blot. As shown in Fig. 4A, we found that TCF7 expression was
significantly upregulated in CRC tissues compared with adjacent
normal tissues. Moreover, the expression of TCF7 was significantly
upregulated in metastatic CRC tissues compared with non-metastatic
tissues (Fig. 4B). Consistently, the
expression of TCF7 was also upregulated in CRC cell lines compared
with FHC cells (Fig. 4C). Moreover,
the protein levels of TCF7 were also upregulated in CRC tissues and
cell lines compared to normal controls (Fig. 4D and E). Through RT-qPCR, we also
demonstrated that the expression of miR-6852 was negatively
correlated with that of TCF7 in CRC tissues (Fig. 4F).
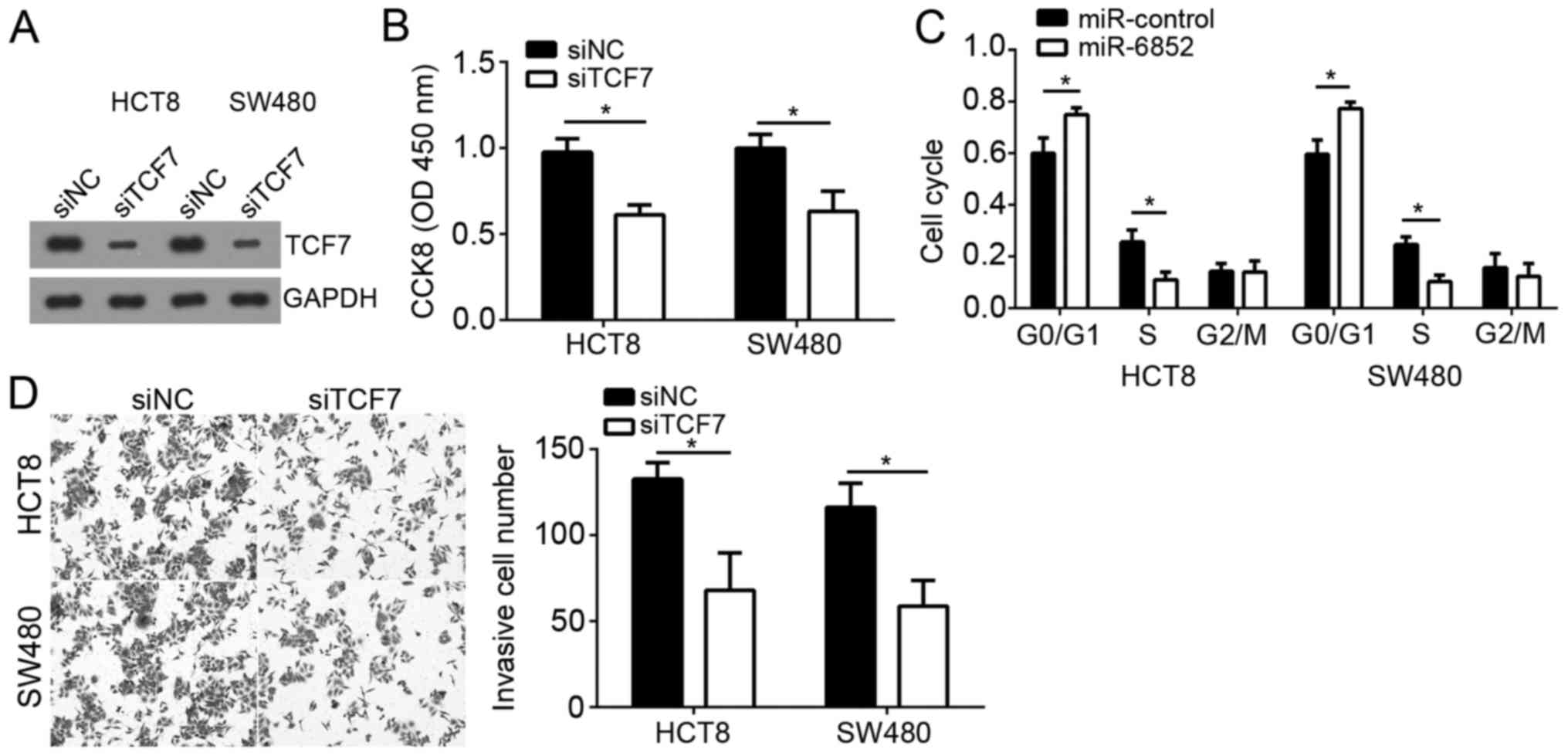
Knockdown of TCF7 inhibits the
proliferation and invasion of CRC cells
To further determine the role of TCF7 in CRC cells,
we knocked down TCF7 in HCT8 and SW480 with specific siRNA. Western
blot analysis indicated that TCF7 was significantly downregulated
in HCT8 and SW480 cells transduced with siTCF7 compared to control
group (Fig. 5A). CCK-8 assay
indicated that knockdown of TCF7 significantly suppressed the
proliferation of HCT8 and SW480 cells (Fig. 5B). Moreover, less cells transduced
with siTCF7 entered into S phase (Fig.
5C). Finally, we showed that knockdown of TCF7 also inhibited
the invasion of HCT8 and SW480 cells (Fig. 5D). Taken together, our study
demonstrated that miR-6852 suppressed the proliferation and
invasion of CRC cells by targeting TCF7, which acted as an oncogene
in CRC cells.
Discussion
CRC is one of the most common malignances and its
incidence is increasing year by year (1). However, no effective therapy has been
developed for treatment of CRC patients especially with metastasis.
The underlying mechanism of CRC progression remains elusive. There
is a very urgent requirement to search novel biomarkers for CRC
diagnosis or prognosis and therapeutic targets for CRC
intervention. Several miRNAs have been demonstrated to be promising
indicators of cancer characteristics, including in CRC. Thus,
understanding the roles and functional mechanism of miRNAs in CRC
progression will contribute to the discovery of biomarkers and
therapeutic targets. In this study, we studied a functionally
undefined miRNA, miR-6852, in CRC. We also demonstrated its
functional mechanism by several experiments.
miRNAs participate in all kinds of biological
processes because of their regulatory roles on gene expression
(21). Several miRNAs are
demonstrated dysregulated in cancers, including gastric cancer
(22), colon cancer (23), breast cancer (24), non-small cell lung cancer (25), bladder cancer (26), ovarian cancer (27), prostate cancer (28) and pancreatic cancer (29). In CRC, many miRNAs has been shown to
regulate tumor progression. For example, miR-21
post-transcriptionally downregulates tumor suppressor Pdcd4 and
stimulates invasion, intravasation and metastasis in CRC (30). miRNA-27b targets vascular endothelial
growth factor C to inhibit tumor progression and angiogenesis in
CRC (31). MicroRNA-497 targets
insulin-like growth factor 1 receptor and has a tumor suppressive
role in human CRC (32). Previous
study indicated miR-6852 induces necrosis in cervical cancer cells
(19). However, the function of
miR-6852 in CRC remains largely unknown. In this study, we found
that miR-6852 was downregulated in CRC tissues and cell lines.
Moreover, we found that miR-6852 expression level was negatively
correlated with tumor metastasis and positively correlated with
patients' survival rate, which suggested that miR-6852 might serve
as a prognostic biomarker in CRC. We also demonstrated that
overexpression of miR-6852 significantly inhibited the
proliferation and invasion of CRC cells. These data suggested that
miR-6852 functions as a tumor suppressor in CRC.
TCF7 is well known for its function in T lymphocyte
development (33). Previous study
indicated that TCF7 can initiate the Wnt/β-catenin signaling
cascade (34), which is abnormally
activated in most CRC cases. Increasing evidences demonstrate TCF7
is widely involved in tumorigenesis, including prostate cancer
(35), osteosarcoma (36) and so on. How TCF7 expression is
regulated requires to be further investigated. In our study, we
found that TCF7 was a target gene of miR-6852 in CRC cells.
Overexpression of miR-6852 significantly reduced the mRNA and
protein levels of TCF7 in CRC cells. We showed that there was an
inverse correlation between the expression of miR-6852 and TCF7 in
CRC tissues. Moreover, we found that TCF7 was significantly
upregulated in CRC tissues compared with normal tissues. And the
expression of TCF7 was correlated with CRC metastasis. Through
functional experiments, we demonstrated that TCF7 knockdown
significantly suppressed the proliferation and invasion of CRC
cells, which indicated that miR-6852-mediated inhibition on cell
proliferation and invasion relies on downregulation of TCF7 at
least in part.
In conclusion, our study demonstrated that miR-6852
suppressed the proliferation and invasion by targeting TCF7 in CRC
cells. And we found that miR-6852 might serve as a prognostic
biomarker for CRC patients.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
BC and XH initiated, designed this work, analyzed,
interpreted the results and wrote this manuscript. All authors read
and approved the final manuscript.
Ethics approval and consent to
participate
For the use of human samples, the protocol for this
study was approved by the Institutional Ethics Committee of Harbin
Medical University Cancer Hospital and all enrolled patients signed
a written informed consent document.
Patient consent for publication
All patients within this study provide consent for
the publication of their data.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Harrison S and Benziger H: The molecular
biology of colorectal carcinoma and its implications: A review.
Surgeon. 9:200–210. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Song M and Chan AT: Diet, gut microbiota,
and colorectal cancer prevention: A review of potential mechanisms
and promising targets for future research. Curr Colorectal Cancer
Rep. 13:429–439. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Powell SM, Zilz N, Beazer-Barclay Y, Bryan
TM, Hamilton SR, Thibodeau SN, Vogelstein B and Kinzler KW: APC
mutations occur early during colorectal tumorigenesis. Nature.
359:235–237. 1992. View
Article : Google Scholar : PubMed/NCBI
|
|
4
|
Coppedè F, Lopomo A, Spisni R and Migliore
L: Genetic and epigenetic biomarkers for diagnosis, prognosis and
treatment of colorectal cancer. World J Gastroenterol. 20:943–956.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Lee YC, Lee YL, Chuang JP and Lee JC:
Differences in survival between colon and rectal cancer from SEER
data. PLoS One. 8:e787092013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ambros V: The functions of animal
microRNAs. Nature. 431:350–355. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Valencia-Sanchez MA, Liu J, Hannon GJ and
Parker R: Control of translation and mRNA degradation by miRNAs and
siRNAs. Genes Dev. 20:515–524. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Miranda KC, Huynh T, Tay Y, Ang YS, Tam
WL, Thomson AM, Lim B and Rigoutsos I: A pattern-based method for
the identification of MicroRNA binding sites and their
corresponding heteroduplexes. Cell. 126:1203–1217. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
He L and Hannon GJ: MicroRNAs: Small RNAs
with a big role in gene regulation. Nat Rev Genet. 5:522–531. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Agrawal L, Sahu S, Ghosh S, Shiga T,
Fujita D and Bandyopadhyay A: Inventing atomic resolution scanning
dielectric microscopy to see a single protein complex operation
live at resonance in a neuron without touching or adulterating the
cell. J Integr Neurosci. 15:435–462. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhang B, Pan X, Cobb GP and Anderson TA:
microRNAs as oncogenes and tumor suppressors. Dev Biol. 302:1–12.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Lu G, Fu D, Jia C, Chai L, Han Y, Liu J,
Wu T, Xie R, Chang Z, Yang H, et al: Reduced miR-105-1 levels are
associated with poor survival of patients with non-small cell lung
cancer. Oncol Lett. 14:7842–7848. 2017.PubMed/NCBI
|
|
14
|
Tahiri A, Aure MR and Kristensen VN:
MicroRNA networks in breast cancer cells. Methods Mol Biol.
1711:55–81. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Du X, Liu B, Luan X, Cui Q and Li L:
miR-30 decreases multidrug resistance in human gastric cancer cells
by modulating cell autophagy. Exp Ther Med. 15:599–605.
2018.PubMed/NCBI
|
|
16
|
Dopeso H, Rodrigues P, Bilic J, Bazzocco
S, Cartón-García F, Macaya I, de Marcondes PG, Anguita E, Masanas
M, Jiménez-Flores LM, et al: Mechanisms of inactivation of the
tumour suppressor gene RHOA in colorectal cancer. Br J Cancer.
118:106–116. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Cao Q, Liu F, Ji K, Liu N, He Y, Zhang W
and Wang L: MicroRNA-381 inhibits the metastasis of gastric cancer
by targeting TMEM16A expression. J Exp Clin Cancer Res. 36:292017.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Shi ZM, Wang L, Shen H, Jiang CF, Ge X, Li
DM, Wen YY, Sun HR, Pan MH, Li W, et al: Downregulation of miR-218
contributes to epithelial-mesenchymal transition and tumor
metastasis in lung cancer by targeting Slug/ZEB2 signaling.
Oncogene. 36:2577–2588. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Poudyal D, Herman A, Adelsberger JW, Yang
J, Hu X, Chen Q, Bosche M, Sherman BT and Imamichi T: A novel
microRNA, hsa-miR-6852 differentially regulated by Interleukin-27
induces necrosis in cervical cancer cells by downregulating the
FoxM1 expression. Sci Rep. 8:9002018. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Garzon R, Fabbri M, Cimmino A, Calin GA
and Croce CM: MicroRNA expression and function in cancer. Trends
Mol Med. 12:580–587. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Guo J, Miao Y, Xiao B, Huan R, Jiang Z,
Meng D and Wang Y: Differential expression of microRNA species in
human gastric cancer versus non-tumorous tissues. J Gastroenterol
Hepatol. 24:652–657. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Díaz R, Silva J, García JM, Lorenzo Y,
García V, Peña C, Rodríguez R, Muñoz C, García F, Bonilla F and
Domínguez G: Deregulated expression of miR-106a predicts survival
in human colon cancer patients. Genes Chromosomes Cancer.
47:794–802. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wang Y, Yu Y, Tsuyada A, Ren X, Wu X,
Stubblefield K, Rankin-Gee EK and Wang SE: Transforming growth
factor-β regulates the sphere-initiating stem cell-like feature in
breast cancer through miRNA-181 and ATM. Oncogene. 30:1470–1480.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Li J, Yang H, Li Y, Liu Y, Chen S, Qi C,
Zhang Q, Lan T, He X, Guan XY and Wang L: microRNA-146
up-regulation predicts the prognosis of non-small cell lung cancer
by miRNA in situ hybridization. Exp Mol Pathol. 96:195–199. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Canturk KM, Ozdemir M, Can C, Öner S, Emre
R, Aslan H, Cilingir O, Ciftci E, Celayir FM, Aldemir O, et al:
Investigation of key miRNAs and target genes in bladder cancer
using miRNA profiling and bioinformatic tools. Mol Biol Rep.
41:8127–8135. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Yang H, Kong W, He L, Zhao JJ, O'Donnell
JD, Wang J, Wenham RM, Coppola D, Kruk PA, Nicosia SV and Cheng JQ:
MicroRNA expression profiling in human ovarian cancer: miR-214
induces cell survival and cisplatin resistance by targeting PTEN.
Cancer Res. 68:425–433. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Beltran H, Yelensky R, Frampton GM, Park
K, Downing SR, MacDonald TY, Jarosz M, Lipson D, Tagawa ST, Nanus
DM, et al: Targeted next-generation sequencing of advanced prostate
cancer identifies potential therapeutic targets and disease
heterogeneity. Eur Urol. 63:920–926. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Dillhoff M, Liu J, Frankel W, Croce C and
Bloomston M: MicroRNA-21 is overexpressed in pancreatic cancer and
a potential predictor of survival. J Gastrointest Surg.
12:2171–2176. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Asangani IA, Rasheed SAK, Nikolova DA,
Leupold JH, Colburn NH, Post S and Allgayer H: MicroRNA-21 (miR-21)
post-transcriptionally downregulates tumor suppressor Pdcd4 and
stimulates invasion, intravasation and metastasis in colorectal
cancer. Oncogene. 27:2128–2136. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Ye J, Wu X, Wu D, Wu P, Ni C, Zhang Z,
Chen Z, Qiu F, Xu J and Huang J: miRNA-27b targets vascular
endothelial growth factor C to inhibit tumor progression and
angiogenesis in colorectal cancer. PLoS One. 8:e606872013.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Guo ST, Jiang CC, Wang GP, Li YP, Wang CY,
Guo XY, Yang RH, Feng Y, Wang FH, Tseng HY, et al: MicroRNA-497
targets insulin-like growth factor 1 receptor and has a tumour
suppressive role in human colorectal cancer. Oncogene.
32:1910–1920. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Weber BN, Chi AW, Chavez A, Yashiro-Ohtani
Y, Yang Q, Shestova O and Bhandoola A: A critical role for TCF-1 in
T-lineage specification and differentiation. Nature. 476:63–68.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Cui L, Guan Y, Qu Z, Zhang J, Liao B, Ma
B, Qian J, Li D, Li W, Xu GT and Jin Y: WNT signaling determines
tumorigenicity and function of ESC-derived retinal progenitors. J
Clin Invest. 123:1647–1661. 2013. View
Article : Google Scholar : PubMed/NCBI
|
|
35
|
Chen WY, Liu SY, Chang YS, Yin JJ, Yeh HL,
Mouhieddine TH, Hadadeh O, Abou-Kheir W and Liu YN: MicroRNA-34a
regulates WNT/TCF7 signaling and inhibits bone metastasis in
Ras-activated prostate cancer. Oncotarget. 6:441–457.
2015.PubMed/NCBI
|
|
36
|
Wang Y, Zhang S, Xu Y, Zhang Y, Guan H, Li
X, Li Y and Wang Y: Upregulation of miR-192 inhibits cell growth
and invasion and induces cell apoptosis by targeting TCF7 in human
osteosarcoma. Tumor Biol. 37:15211–15220. 2016. View Article : Google Scholar
|