Introduction
Diabetic nephropathy (DN) is a microvascular
complication of diabetes that is clinically characterized by
proteinuria (1). Previous studies
have demonstrated that podocytes have a vital role in the induction
of proteinuria, which contributes to DN (2–4).
Podocytes are located in the glomerular basement membrane outside
the glomerular capillaries (5).
Proteinuria was separated from proteinuria podocytes and
apoptosisoccursin glomerularbasement membrane (6,7).
However, previous studies have demonstrated that podocyte
detachment and apoptosis occur prior to proteinuria (6,8). It has
also previously been indicated that podocyte dysfunction may serve
as a pathway contributing to proteinuria via
epithelial-to-mesenchymal transition (EMT) (9,10).
Blocking EMT has also been indicated to reverse podocyte apoptosis
in its early phase (9,11). Thus, these findings may provide a
novel pathway to alleviate DN.
Podocytes are terminally differentiated visceral
epithelial cells that are located outside the glomerular
capillaries where they form the final filtration barrier to protein
loss (12–14). In diabetic nephropathy, podocytes
injury leads to the disruption of the filtration barrier and
protein leakage. The loss of podocytes has been indicated as an
important early pathologic marker (15–17).
Accumulating evidence suggests that EMT is a possible engagement
for podocyte depletion in diabetic nephropathy. During EMT,
epithelial cells are transformed to mesenchymal cells in response
to injurious stimuli (11,13,16).
Insulin-like growth factor binding protein 7
(IGFBP7) is a member of the IGF binding protein family, and is able
to regulate cell proliferation, differentiation, angiogenesis, cell
adhesion and cellular senescence in a variety of cells (18–23).
Furthermore, silence of IGFBP7 is associated with poor
post-operative survival in various types of cancer, such as glioma,
liver and colorectal cancer (24–27).
However, the effects of IGFBP7 on apoptosis and EMT of high glucose
induced-podocyteshave are yet to be elucidated.
In the present study, podocytes were identified and
the role of IGFBP7 on cell proliferation, apoptosis and EMT of
podocytes mediated by high glucose was analyzed. The expression
levels of transforming growth factor (TGF)-β1/mothers against
decapentaplegic homolog (Smad) pathway were also detected, and it
was observed that silencing IGFBP7 inhibited the TGF-β1/Smad
pathway in podocytes induced by high glucose. These findings
suggest that IGFBP7 may serve as a potential therapeutic target for
DN.
Materials and methods
Cell culture
Human podocytes (cat. no. BNCC340460) were obtained
from the Type Culture Collection of the Chinese Academy of Sciences
(Shanghai, China). Podocytes were cultured in an incubator (95%
humidity, 5% CO2) at 37°C for 48 h in Ham's F12 nutrient
mixture-Dulbecco's modified Eagle's medium (Thermo Fisher
Scientific, Inc., Waltham, MA, USA) with 10% fetal bovine serum
(Gibco; Thermo Fisher Scientific, Inc.) and 1%
penicillin-streptomycin G (Invitrogen; Thermo Fisher Scientific,
Inc.).
Cell treatment
Podocytes (3×105 cells/ml) were seeded in
6-wells plates, and treated with PBS and glucose at 5, 10, 20, 40,
60, 80 or 100 mM at room temperature. The podocytes were then
transferred to the incubator and held for 6, 12, 24 and 48 h at
37°C. Negative control small interfering (si)RNA (siNC;
5′-ACGUGACACGUUCGGAGAATT-3′) and siRNA against IGFBP7 (siIGFBP7;
5′-CATCCAATTCCCAAGGACAG-3′) vectors were chemically synthesized by
Shanghai GenePharma Co., Ltd. (Shanghai, China) and prepared for
transfection. Podocytes were plated in 6-wells plates at a density
of 1×106 cells per well. The vectors (50 nM) were
transfected into podocytes using Lipofectamine® 2000
(Invitrogen; Thermo Fisher Scientific, Inc.) for 48 h according to
the manufacturer's instructions. following 48 h transfection,
experiments were performed.
RNA extraction and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
Total RNA was isolated using TRIzol (Invitrogen;
Thermo Fisher Scientific, Inc.) following the manufacturer's
protocol and reverse transcribed to cDNA assay using an miScript II
RT kit (Qiagen GmbH, Hilden, Germany). The mRNA expression levels
were measured using the SYBR-Green PCR Master Mix kit (Takara
Biotechnology Co., Ltd., Dalian, China). The target genes and GAPDH
were analyzed by RT-qPCR using an ABI 7500 system (Applied
Biosystems; Thermo Fisher Scientific, Inc.). The thermocycling
conditions were as follows: Pre-degeneration, 90°C for 10 min;
(denaturation, 90°C for 40 sec; annealing, 62°C for 35 sec, for 30
cycles) extension, 72°C for 30 sec. Primers were designed as
follows: IGFBP7 forward, 5′-GTGGCCCAGAAAAGCATGAA-3′ and reverse,
5′-AAGTTGGGTATAGCTCGGCA-3′; apoptosis inducing factor (AIF)
forward, 5′-CCCAATGTTGAGTTGGCCAA-3′ and reverse,
5′-AGCGTGATCATGGTGCTCTA-3′; caspase-3 forward,
5′-TTGCCACCTGTCCAGTTTTG-3′ and reverse, 5′-AGGAGTGAGTGGTCTTGCTC-3′;
caspase-9 forward, 5′-GCCCCATATGATCGAGGACA-3′ and reverse,
5′-CAGAAACGAAGCCAGCATGT-3′; nephrin forward,
5′-ACCGTGAGCTCCTTCTATCG-3′ and reverse, 5′-CATTCCACAGATGCAGAGCC-3′;
phosphorylated (p)-cadherin forward, 5′-GAGAAGGAGACAGGCTGGTT-3′ and
reverse, 5′-GGGTAAACTTGGGCTTGTGG-3′; fibroblast-specific protein
(FSP)-1 forward, 5′-GAGAAGGAGACAGGCTGGTT-3′ and reverse,
5′-GGGTAAACTTGGGCTTGTGG-3′; α-smooth muscle actin (SMA) forward,
5′-CTGGGGAGTTGGAAGCAGTA-3′ and reverse, 5′-CTGTTTCACAAGCCCCTCAC-3′;
Snail forward, 5′-AGTGGTTCTTCTGCGCTACT-3′ and
reverse,5′-GTAGGGCTGCTGGAAGGTAA-3′; TGF-β1 forward,
5′-GTGGTGATCATGGAAGACGC-3′ and reverse, 5′-TTGTGGTCAGGATTCTCGCT-3′;
and Smad7 forward, 5′-TAGCCGACTCTGCGAACTAG-3′ and reverse,
5′-CACTCTCGTCTTCTCCTCCC-3′; GAPDH forward,
5′-GTGGAGTCTACTGGCGTCTT-3′ and reverse, 5′-CCTTCCACGATGCCAAAGTT-3′.
Data were quantified applying the 2−ΔΔCq method
(28). The relative mRNA expression
levels were normalized to GAPDH. 3 independent experiments were
performed, and the mean data collected was calculated.
Western blot analysis
An RIPA buffer (Beyotime Institute of Biotechnology,
Haimen, China) was used to lyse cells, according to the
manufacturer's protocol, and a BCA Protein Assay kit (Thermo Fisher
Scientific, Inc.) was applied to detect the protein concentrations.
Equivalent proteins (25 µg) were separated by 10% SDS-PAGE and
transferred to a polyvinyl difluoride (PVDF) membrane (PerkinElmer,
Inc., Waltham, MA, USA). PVDF membranes were subsequently blocked
for 2 h at room temperature with 5% non-fat dry milk and incubated
with the following primary antibodies overnight at 4°C: Anti-GAPDH
(1:2,000; cat. no. ab8245), anti-IGFBP7 (1:1,000; cat. no.
ab74169), anti-AIF (1:1,000; cat. no. ab1998), anti-cleaved
(c)-Caspase-3 (1:1,500; cat. no. ab13586), anti-c-Caspase-9
(1:1,500; cat. no. ab25758), anti-nephrin (1:1,000; cat. no.
ab58968), anti-p-cadherin (1:1,000; cat. no. ab19350), anti-FSP-1
(1:2,000; cat. no. ab40722), anti-α-SMA (1:1,000; cat. no. ab5694),
anti-Snail (1:1,000; cat. no. ab53519), anti-TGF-β1 (1:1,000; cat.
no. ab92486), anti-Smad7 (1:1,000; cat. no. ab55493), anti-p-Smad3
(1:1,000; cat. no. ab52903) and anti-smad3 (1:1,000; cat. no.
ab40854; all Abcam, Cambridge, UK). Subsequently, the PVDF
membranes were incubated with secondary antibody [horseradish
peroxidase conjugated (HRP) mouse anti-rabbit immunoglobulin G
(IgG); 1:5,000; cat. no. sc-2357; Goat anti-mouse IgG-HRP; 1:6,000;
cat. no. sc-2005; Donkey anti-goat IgG-HRP; 1:6,000; cat. no.
sc-2020; each Santa Cruz Biotechnology, Inc., Dallas, TX, USA] for
1 h at room temperature. The results were analyzed using the
enhanced chemiluminescence substrate kit (GE Healthcare, Chicago,
IL, USA) and the automatic chemiluminescence image analysis system
(Tanon 4200; YPH-Bio, Beijing, China).
Cell proliferation assay
An MTT assay was performed to determine the
proliferation of podocytes exposed to various treatments. The
treated cells (3×103 cells/well) were seeded in a
96-well plate and cultured for 6, 12, 24 and 48 h at 37°C. Next,
cells were incubated with 20 µl MTT (5 mg/ml) solution for 4 h at
37°C and 10 µl dimethyl sulfoxide (cat. no. 6-68-5; MOLBASE
Biotechnology Co., Ltd., Shanghai, China) was added to dissolve the
formazan. The absorbance was measured at 490 nm using a microplate
reader.
Cell apoptosis analysis
Podocytes (5×104 cells/well) were seeded
in 6-well plates and cultured at 37°C for 24 h. Then, cells were
treated with PBS (Control), siNC, high glucose (HG; 60 mM), siNC
and HG, or siIGFBP7 and HG for 12 h. Cells were subsequently
centrifuged (1,000×g, 5 min, 4°C) and fixed for 2 min in 70% (v/v)
ethanol on ice. According to the manufacturer's protocol, the cells
were double-stained using an Annexin V-fluorescein
isothiocyanate/propidium iodide kit (BD Biosciences, Franklin
Lakes, NJ, USA) for 20 min at room temperature in the dark. Samples
were immediately analyzed for apoptosis using a FACSCalibur flow
cytometer (BD Biosciences). The data was analyzed by the ModFit LT
2.0 software (Verity Software House, Inc., Tophsham, ME, USA).
Immunofluorescence staining
Podocytes (5×104 cells/well) were seeded
in 6-well plates and cultured at 37°C for 12, 24 and 48 h. Then,
cells were washed with PBS and fixed with 4% paraformaldehyde at
4°C for 20 min. The fixed cells were washed with PBS for 3 times,
and permeabilized with 0.2% Triton X-100 for 3 min. Cells were
subsequently washed with PBS, and blocked with 10% goat serum
(Shanghai Haoran Biological Technology Co., Ltd., Shanghai, China)
for 30 min at 37°C. The anti-synaptopodin antibody (1:600; cat. no.
ab220345; Abcam) was used to incubate with the cells overnight at
4°C. Following washing with PBS in triplicate, cells were incubated
with an Alexa-Fluor 633-conjugated secondary antibody (1:5,000;
cat. no. A20005; ThermoFisher Scientific, Inc.) at room temperature
for 1 h. DAPI (Thermo Fisher Scientific, Inc.) was then added to
stain the nuclei at room temperature for 15 min. Finally, samples
were evaluated under fluorescence microscopy (magnification, ×200;
Olympus Corporation, Tokyo, Japan).
ELISA
TGF-β1 concentration was analyzed using an ELISA kit
(cat. no. SX01158; Shanghai Senxiong Biotech Industry Co., Ltd.,
Shanghai, China), according to the manufacturer's protocol.
Podocytes (5×104 cells/well) were seeded in 6-well
plates and cultured in at 37°C for 24 h. cells were then treated
with PBS (Control), siNC, high glucose (HG; 60 mM), siNC and HG, or
siIGFBP7 and HG for 12 h at 37°C. Diluted standard product (20 µl)
was prepared. Enzyme reagent (100 µl) was added for 30 min at 37°C,
and subsequently color agent (100 µl) was added to plates for 15
min at 37°C. Finally, the reaction was terminated using termination
liquid (50 µl). Subsequently, the absorbance (OD value) was
measured at 450 nm using a microplate reader. A standard curve was
produced using GraphPad prism 7 software (GraphPad Software, Inc.,
La Jolla, CA, USA) and the different concentrations of samples were
obtained by comparing the OD value of the samples to the standard
curve.
Statistical analysis
Data were analyzed using SPSS 19.0 software (IBM
Corp., Armonk, NY, USA). Significant differences between groups
were assessed using one-way analysis, and the post hoc LSD test was
performed. Data are presented as the mean ± standard deviation.
P<0.05 was considered to indicate a statistically significant
difference.
Results
Glucose promotes podocyte
proliferation in a time-dependent and dose-dependent manner
To explore whether glucose is associated with the
progression and development of podocytes, the expression level of
synaptopodin in podocytes was measured via immunofluorescence
assay. Blue and red staining represented the nucleus and
synaptopodin, respectively. Results demonstrated that synaptopodin
staining at 48 h was deeper than that of 12 and 24 h. This
indicated that synaptopodin was highly expressed in podocytes
(Fig. 1A). In addition, podocytes
were treated with PBS and glucose at 0, 5, 10, 20, 40, 60, 80 and
100 mM for 6, 12, 24 and 48 h. The results indicated that the
proliferation ability of podocytes increased with increasing dosage
and time (Fig. 1B).
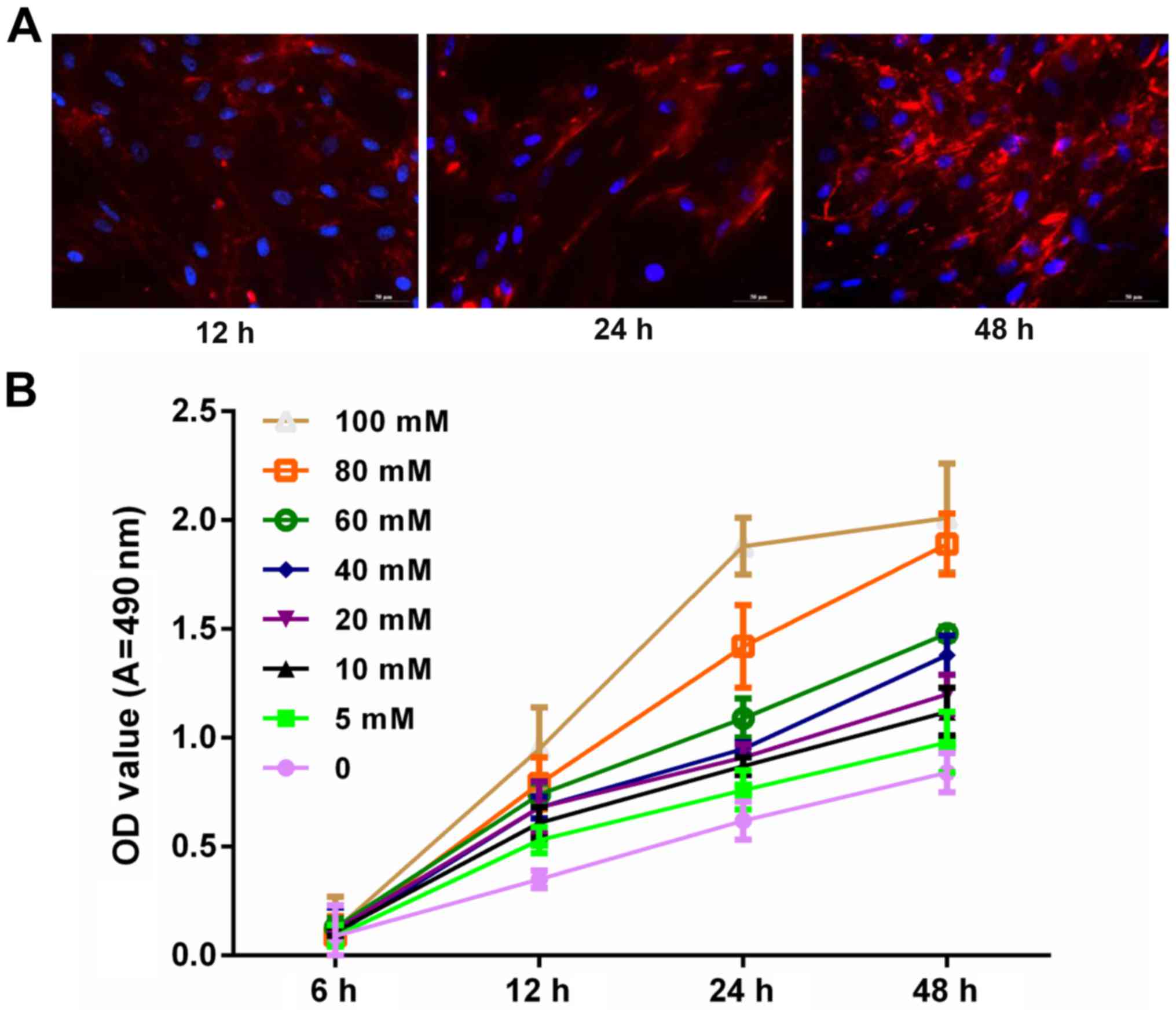 | Figure 1.Glucose promoted podocyte
proliferation in a marked time- and dose-dependent manner. (A)
Podocytes were cultured at 37°C for 12, 24 and 48 h. The
synaptopodin expression was detected using immunofluorescence
assay. The cells were visualized under fluorescence microscopy.
Blue and red fluorescence represents the nucleus and synaptopodin,
respectively. (B) MTT assay was performed to measure the
proliferation of podocytes treated with PBS and glucose at 0, 5,
10, 20, 40, 60, 80 and100 mM for 6, 12, 24 and 48 h. OD, optical
density; A, absorbance. |
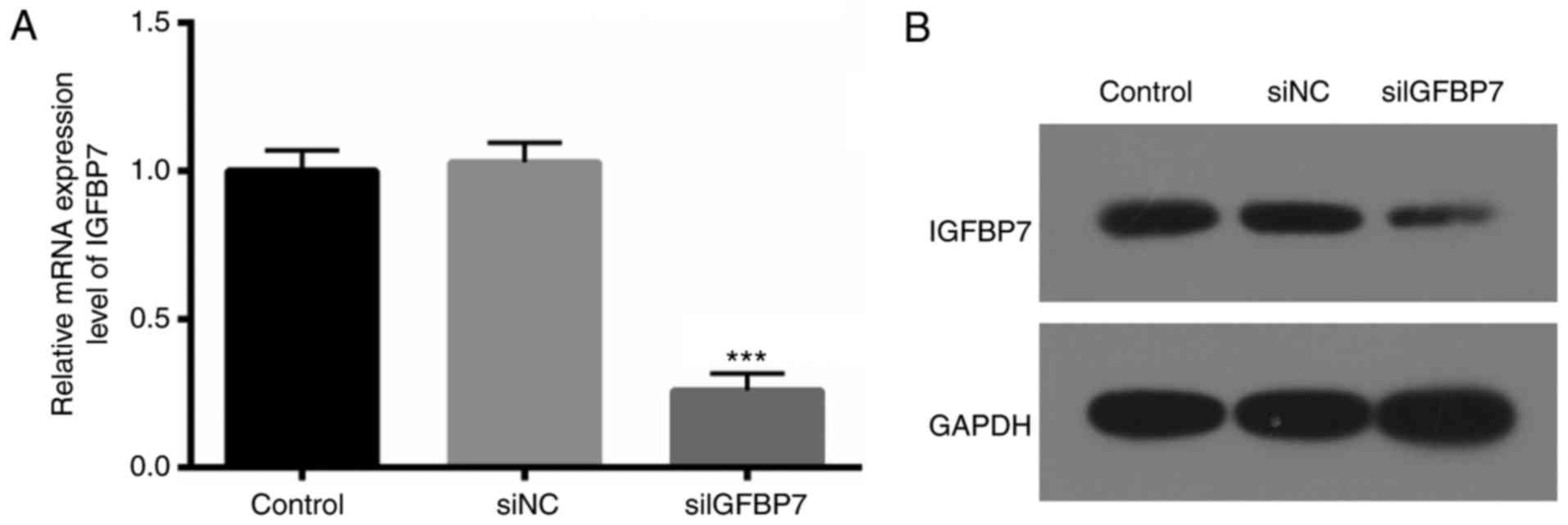
IGFBP7 silencing in podocytes via
siRNA
To further determine the exact roles of IGFBP7 in
podocytes, IGFBP7 expression was blocked via transfection with
siIGFBP7. RT-qPCR and western blotting were then used to verify the
IGFBP7 knockdown efficiency, and it was. demonstrated that IGFBP7
mRNA (Fig. 2A) and protein (Fig. 2B) expression was significantly
(P<0.001) and markedly downregulated, respectively, in podocytes
transfected with siIGFBP7, compared with siNC. These results
indicated that the efficiency and specificity of siIGFBP7 was
high.
IGFBP7 silencing inhibits apoptosis of
podocytes mediated by HG
It wasthen detected whether IGFBP7 serves a role in
the apoptosis of podocytes mediated by HG via apoptosis assay. The
results demonstrated that the apoptotic ability was significantly
increased in the HG group compared with control (P<0.001), and
that IGFBP7 silencing significantly inhibited the apoptotic ability
of podocytes induced by HG (P<0.001; Fig. 3A and B). In addition, the expression
levels of three apoptosis-related genes (AIF, caspase-3 and
caspase-9) were detected using RT-qPCR and western blotting. It was
demonstrated that AIF, c-caspase-3 and c-caspase-9 expressions were
significantly increased in the HG group compared with the control
group, and that silencing IGFBP7 significantly downregulated
expression of the three genes in podocytes induced by HG
(P<0.001; Fig. 3C and D). These
data indicated that silencing IGFBP7suppresses cell apoptosis of
podocytes mediated by HG.
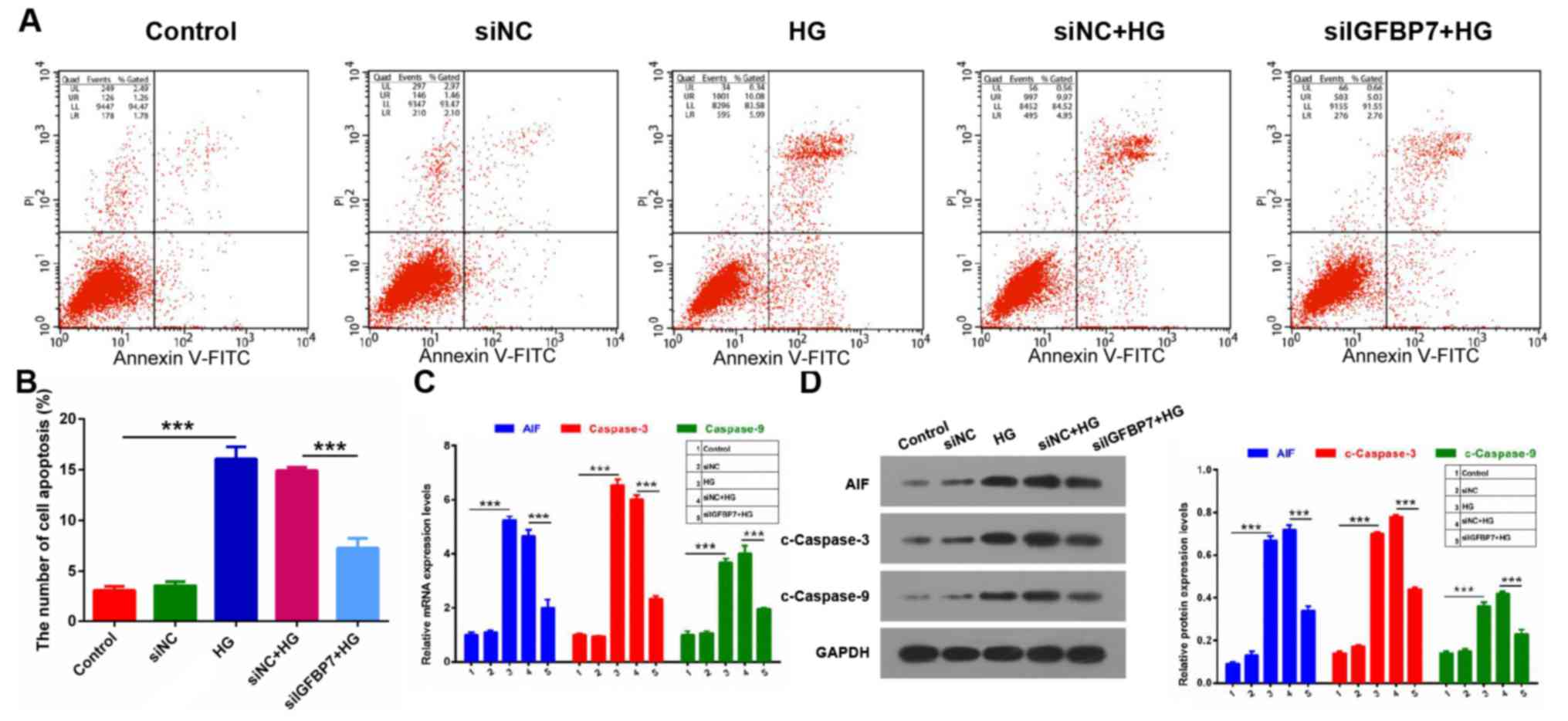 | Figure 3.Silencing IGFBP7 inhibited apoptosis
of podocytes mediated by HG. Podocytes were treated with PBS
(control), siNC, 60 mM glucose (HG), siNC and HG, and siIGFBP7 and
HG for 12 h. (A) Total apoptotic cells were measured using flow
cytometry in treated podocytes. (B) Quantitative analysis of number
of apoptotic cells. (C) AIF, caspase-3 and caspase-9 expression was
detected via reverse transcription-quantitative polymerase chain
reaction. (D) Western blotting of AIF, c-caspase-3 and c-caspase-9.
Protein levels were quantified using densitometry. ***P<0.001.
IGFBP7, insulin-like growth factor-binding protein 7; HG, high
glucose; siNC, normal control small interfering RNA; siIGFBP7,
IGFBP7 small interfering RNA; AIF, apoptosis inducing factor; c,
cleaved; PI, propidium iodide; FITC, fluorescein
isothiocyanate. |
Silencing IGFBP7 inhibits EMT of
podocytes induced by HG
To further investigate whether IGFBP7 participates
in the process of cell EMT of podocytes, RT-qPCR and western
blotting were used to analyze nephrin, p-cadherin, FSP-1, α-SMA and
Snail expression. It was demonstrated that HG inhibited the
expression of epithelial markers (nephrin and p-cadherin) and
promoted the expression of mesenchymal markers (FSP-1, α-SMA and
Snail), which were reversed by silencing IGFBP7 (Fig. 4).
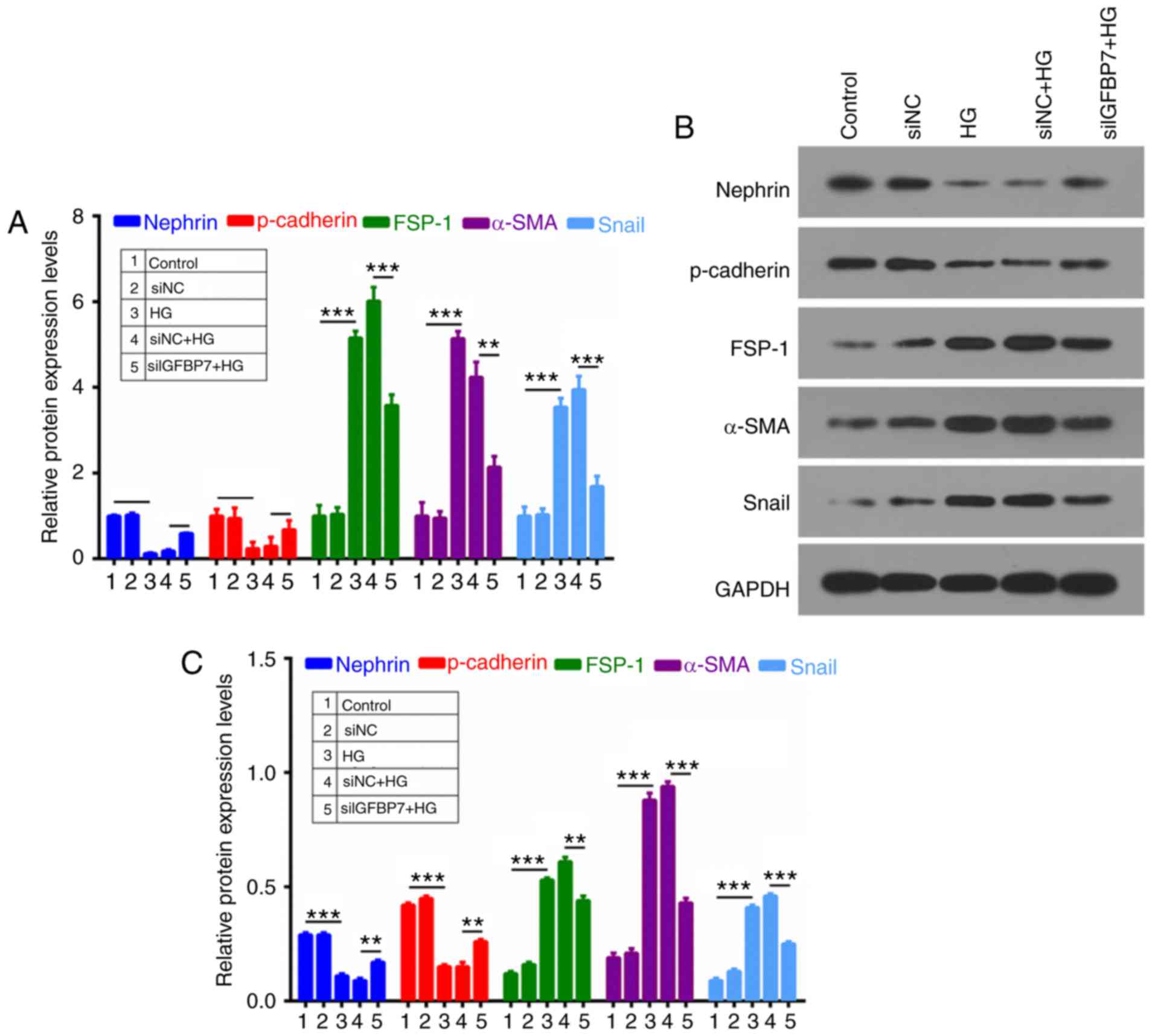 | Figure 4.Silencing IGFBP7 inhibited epithelial
mesenchymal transformation of podocytes induced by HG. Podocytes
were treated with PBS (control), siNC, 60 mM glucose (HG), siNC and
HG, and siIGFBP7 and HG for 12 h. (A) Reverse
transcription-quantitative polymerase chain reaction was used to
detect nephrin, p-cadherin, FSP-1, α-SMA and Snail expression in
treated podocytes. Nephrin, p-cadherin, FSP-1, α-SMA, and Snail
expressions were analyzed via (B) western blotting and (C)
quantified. **P<0.01; ***P<0.001. IGFBP7, insulin-like growth
factor-binding protein 7; HG, high glucose; siNC, normal control
small interfering RNA; siIGFBP7, IGFBP7 small interfering RNA; p,
phosphorylated; FSP-1, fibroblast-specific protein-1; α-SMA,
α-smooth muscle actin. |
IGFBP7 silencing suppresses the
TGF-β1/Smad pathway in podocytes induced by HG
TGF-β1/Smad signaling is associated with antimitotic
activities in epithelial tissues (29). An ELISA assay was used to detect
TGF-β1 concentration, which indicated that HG significantly
increased TGF-β1 concentration, compared with controls
(P<0.001), and IGFBP7 silencing significantly decreased TGF-β1
concentration (P<0.05; Fig. 5A).
RT-qPCR and western blotting were then performed to measure the
expression levels of TGF-β1 and Smad7, which indicated that HG
significantly upregulated TGF-β1 expression, and significantly
downregulated Smad7 expression, and that IGFBP7 silencing
significantly reversed this HG-mediated regulation (P<0.001;
Fig. 5B and C). In addition, protein
expression levels of p-Smad3 and Smad3 were detected, and it was
demonstrated that HG significantly promoted the phosphorylation of
Smad3, and that IGFBP7 silencing further inhibited the
phosphorylation of Smad3 induced by HG (P<0.001; Fig. 5D).
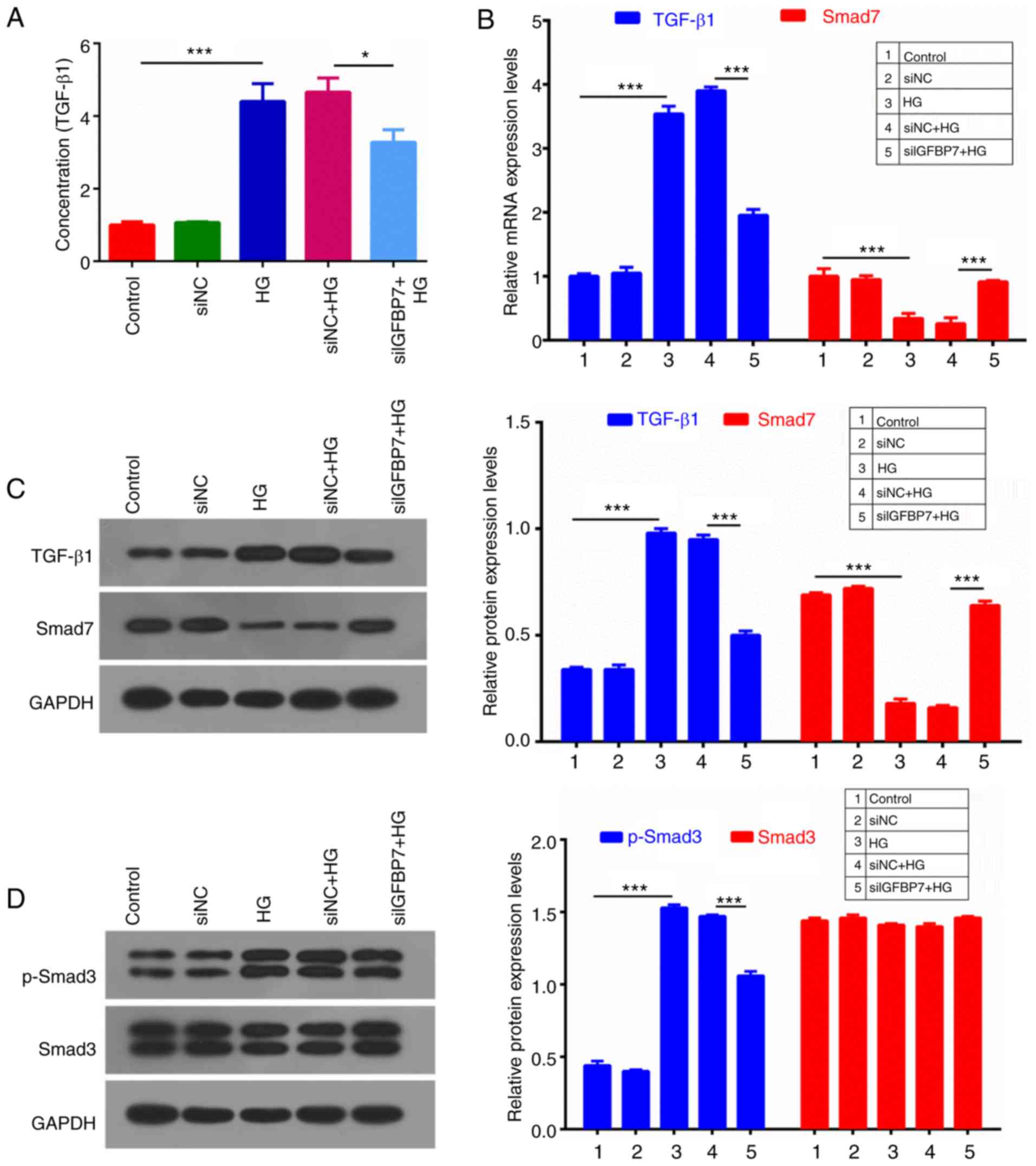 | Figure 5.Silencing IGFBP7 suppressed the
TGF-β1/Smad pathway in podocytes induced by HG. Podocytes were
treated with PBS (control), siNC, 60 mM glucose (HG), siNC and HG,
and siIGFBP7 and HG for 12 h. (A) TGF-β1 concentration was detected
using ELISA. (B) The mRNA expression levels of TGF-β1 and Smad7
were analyzed using reverse transcription-quantitative polymerase
chain reaction. (C) The protein expression levels of TGF-β1 and
Smad7 were analyzed via western blotting. (D) The protein
expression levels of p-Smad3 and Smad3 were analyzed via western
blotting. *P<0.05; ***P<0.001. IGFBP7, insulin-like growth
factor-binding protein 7; TGF-Smad7 were analyzed via western
bloSmad, mothers against decapentaplegic homolog; HG, high glucose;
siNC, normal control small interfering RNA; siIGFBP7, IGFBP7 small
interfering RNA; p, phosphorylated. |
Discussion
DN is one of the main microvascular complications of
diabetes mellitus, and it has been identified as a leading cause of
end stage renal disease in the United States (30,31).
Meanwhile, DN also is a serious threat to people's health in China
(32,33). The elucidation of the pathogenesis of
DN and the development of novel therapeutic measures are of
importance for patients diagnosed with DN. The pathogenesis of DN
is complex, and is associated with many factors, including
hyperglycemia, lipid metabolism disorders, abnormal hemodynamics,
inflammatory cytokines, oxidative stress and cell apoptosis
(34–36). Following the construction of podocyte
cell strains and the rising development of biochemical technology,
podocytes have been increasingly used in research (37,38).
Podocyte injury has been identified to serve an important role in
the pathogenesis of DN and is considered as a pivotal factor for
inducing proteinuria and glomerular sclerosis (5). This provides a novel approach for
preventing and treating DN.
Podocytes are also known as glomerular visceral
epithelial cells, which comprise the glomerular filtration barrier
with a fenestrated endothelial layer and the glomerular basement
membrane (5). Podocytes are highly
differentiated cells and have a limited proliferative ability
(39). When the number of
exfoliative cells exceeds that of proliferative cells, the
glomerular filtration barrier is destroyed, which leads to massive
proteinuria, excretion of collagen IV by podocytes, and ultimately
glomerular sclerosis (40).
Hyperglycemia, angiotensin II, lipid metabolism disorder, oxidative
stress and inflammatory cytokines stimulate podocytes to be
dysfunctional and impaired (41).
Podocytes also have a variety of specific markers, such as
synaptopodin, Wilms tumor protein, glomerular epithelial protein-1,
nephrin, podocalyxin, and C3b (42).
Among them, synaptopodin is a protein coupling with actin
microfilament, which is located in glomerular podocytes and
hindbrain synapses (43,44). The present study demonstrated that
synaptopodin was highly expressed in podocytes, and that HG
promoted podocyte proliferation in a time-dependent and
dose-dependent manner.
IGFBP7 has binding affinities to IGFs and has been
demonstrated to either positively or negatively regulate the IGF
signaling pathway, and serve a crucial role in cell growth,
differentiation and development in an IGF-independent manner
(45). It has also been demonstrated
that IGFBP7 serves an important role in DN, for example, IGFBP7 is
associated with the process of human renal proximal tubular
epithelial cells via Smad2/4 (46).
However, the role of IGFBP7 in DN is yet to be elucidated. In the
present study, IGFBP7 expression in podocytes was blocked via
transfection with siRNA, which reduced the HG-induced increase of
podocyte apoptosis. HG also increased AIF, c-caspase-3 and
c-caspase-9 expression, which was ameliorated by IGFBP7
silencing.
A number of previous studies have suggested that EMT
serves a role podocyte depletion in DN, which is responsible for
the onset of proteinuria (47–49). It
has been demonstrated that TGF-β significantly promotes EMT by
reducing levels of epithelial markers (nephrin and p-cadherin) and
increasing mesenchymal marker (FSP-1) levels (50,51).
Furthermore, α-SMA is also a mesenchymal marker (52). Snail is a pivotal transcriptional
factor that participates in initiating EMT (11,16,17,11). In
the present study, it was demonstrated that HG significantly
inhibited nephrin and p-cadherin expressions, and promoted the
expressions of FSP-1, α-SMA and Snail, which was ameliorated by
IGFBP7 silencing. This suggested that silencing IGFBP7 inhibited
EMT of podocytes induced by HG.
The cytokines of the TGF-β family have a number of
biological effects through different signal transduction pathways,
among which, the Smad protein family serves an important role
(55). The process of transmembrane
signaling depends on the Smad protein family, which regulates
expression of a number of genes (56). In the present study, the expression
of TGF-β1 and its signal transduction elements (Smad2/3 and Smad7)
was examined, and renal extracellular matrix accumulation in the
renal tissues of experimental diabetes was assessed to further
clarify the effects of Smads on the development of DN. It was
demonstrated that HG significantly upregulated TGF-β1 expression,
promoted phosphorylation of Smad3 and downregulated Smad7
expression, which was ameliorated by IGFBP7 silencing.
In conclusion, the present study indicated that
glucose facilitated podocyte proliferation in a time-dependent and
dose-dependent manner, and that IGFBP7 silencing inhibited
apoptosis and EMT of podocytes mediated by HG via suppressing the
TGF-β1/Smad pathway. However, it is not yet clear how GFBP7
regulates caspases and EMT-related factors, and the specific
molecular mechanism of IGFBP7 in the apoptosis and EMT of podocytes
should be explored in further studies. The present study provides
further elucidation and a theoretical basis for novel diagnostic
and treatment options for DN.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
XJC, LW and XLW were responsible for the design of
the experimental scheme. XLW and FYH were responsible for analyzing
and interpreting data. XJC was involved in drafting the manuscript.
All authors were responsible for giving final approval of the
version to be published
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Zheng S, Powell DW, Zheng F, Kantharidis P
and Gnudi L: Diabetic nephropathy: Proteinuria, inflammation, and
fibrosis. J diabetes Res. 2016:52415492016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Wang S, Li Y, Zhao J, Zhang J and Huang Y:
Mesenchymal stem cells ameliorate podocyte injury and proteinuria
in a type 1 diabetic nephropathy rat model. Biol Blood Marrow
Transplant. 19:538–546. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Yasuda-Yamahara M, Kume S, Tagawa A,
Maegawa H and Uzu T: Emerging role of podocyte autophagy in the
progression of diabetic nephropathy. Autophagy. 11:2385–2386. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Zhao L, Wang X, Sun L, Nie H, Liu X, Chen
Z and Guan G: Critical role of serum response factor in podocyte
epithelial-mesenchymal transition of diabetic nephropathy. Diab
Vasc Dis Res. 13:81–92. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Yasuno K, Kamiie J and Shirota K: Analysis
of ultrastructural glomerular basement membrane lesions and
podocytes associated with proteinuria and sclerosis in
osborne-mendel rats with progressive glomerulonephropathy. J Vet
Sci. 14:223–226. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Huang L, You YS and Wu W: Role of
CD2-associated protein in podocyte apoptosis and proteinuria
induced by angiotensin II. Ren Fail. 36:1328–1332. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Tagawa A, Yasuda M, Kume S, Yamahara K,
Nakazawa J, Chin-Kanasaki M, Araki H, Araki S, Koya D, Asanuma K,
et al: Impaired podocyte autophagy exacerbates proteinuria in
diabetic nephropathy. Diabetes. 65:755–767. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Liao R, Liu Q, Zheng Z, Fan J, Peng W,
Kong Q, He H, Yang S, Chen W, Tang X and Yu X: Tacrolimus protects
podocytes from injury in lupus nephritis partly by stabilizing the
cytoskeleton and inhibiting podocyte apoptosis. PloS One.
10:e01327242015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Chitra PS, Swathi T, Sahay R, Reddy GB,
Menon RK and Kumar PA: Growth hormone induces transforming growth
factor-beta-induced protein in podocytes: Implications for podocyte
depletion and proteinuria. J Cell Biochem. 116:1947–1956. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ziyadeh FN and Wolf G: Pathogenesis of the
podocytopathy and proteinuria in diabetic glomerulopathy. Curr
Diabetes Rev. 4:39–45. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Yamaguchi Y, Iwano M, Suzuki D, Nakatani
K, Kimura K, Harada K, Kubo A, Akai Y, Toyoda M, Kanauchi M, et al:
Epithelial-mesenchymal transition as a potential explanation for
podocyte depletion in diabetic nephropathy. Am J Kidney Dis.
54:653–664. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Faul C, Asanuma K, Yanagida-Asanuma E, Kim
K and Mundel P: Actin up: Regulation of podocyte structure and
function by components of the actin cytoskeleton. Trends Cell Biol.
17:428–437. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Li Y, Kang YS, Dai C, Kiss LP, Wen X and
Liu Y: Epithelial-to-mesenchymal transition is a potential pathway
leading to podocyte dysfunction and proteinuria. Am J Pathol.
172:299–308. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Pavenstadt H, Kriz W and Kretzler M: Cell
biology of the glomerular podocyte. Physiol Rev. 83:253–307. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Durvasula RV and Shankland SJ: Podocyte
injury and targeting therapy: An update. Curr Opin Nephrol
Hypertens. 15:1–7. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kang YS, Li Y, Dai C, Kiss LP, Wu C and
Liu Y: Inhibition of integrin-linked kinase blocks podocyte
epithelial-mesenchymal transition and ameliorates proteinuria.
Kidney Int. 78:363–373. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Susztak K, Raff AC, Schiffer M and
Bottinger EP: Glucose-induced reactive oxygen species cause
apoptosis of podocytes and podocyte depletion at the onset of
diabetic nephropathy. Diabetes. 55:225–233. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Chen LH, Liu DW, Chang JL, Chen PR, Hsu
LP, Lin HY, Chou YF, Lee CF, Yang MC, Wen YH, et al: Methylation
status of insulin-like growth factor-binding protein 7 concurs with
the malignance of oral tongue cancer. J Exp Clin Cancer Res.
34:202015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Honore PM, Nguyen HB, Gong M, Chawla LS,
Bagshaw SM, Artigas A, Shi J, Joannes-Boyau O, Vincent JL, Kellum
JA and Sapphire: Topaz Investigators: Urinary tissue inhibitor of
metalloproteinase-2 and insulin-like growth factor-binding protein
7 for risk stratification of acute kidney injury in patients with
sepsis. Crit Care Med. 44:1851–1860. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Jiang J, Chen Z, Liang B, Yan J, Zhang Y,
Xu H, Huang Y and Jiang H: The change of circulating insulin like
growth factor binding protein 7 levels may correlate with
postoperative cognitive dysfunction. Neurosci Lett. 588:125–130.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Li N, Zhang Z, Zhang L, Wang S, Zou Z,
Wang G and Wang Y: Insulin-like growth factor binding protein 7, a
member of insulin-like growth factor signal pathway, involved in
immune response of small abalone haliotis diversicolor. Fish
Shellfish Immunol. 33:229–242. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Tamura K, Yoshie M, Hashimoto K and
Tachikawa E: Inhibitory effect of insulin-like growth
factor-binding protein-7 (IGFBP7) on in vitro angiogenesis of
vascular endothelial cells in the rat corpus luteum. J Reprod Dev.
60:447–453. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Yuan L, Fan WJ, Yang XG, Rao SM, Song JL
and Song GH: Inhibitory effect of exogenous insulin-like growth
factor binding protein 7 on proliferation of human breast cancer
cell line MDA-MB-453 and its mechanism. Sheng Li Xue Bao.
65:519–524. 2013.(In Chinese). PubMed/NCBI
|
|
24
|
Bolomsky A, Hose D, Schreder M, Seckinger
A, Lipp S, Klein B, Heintel D, Ludwig H and Zojer N: Insulin like
growth factor binding protein 7 (IGFBP7) expression is linked to
poor prognosis but may protect from bone disease in multiple
myeloma. J Hematol Oncol. 8:102015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Tian X, Zhang L, Sun L, Xue Y and Xie S:
Low expression of insulin-like growth factor binding protein 7
associated with poor prognosis in human glioma. J Int Med Res.
42:651–658. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Chen D, Siddiq A, Emdad L, Rajasekaran D,
Gredler R, Shen XN, Santhekadur PK, Srivastava J, Robertson CL,
Dmitriev I, et al: Insulin-like growth factor-binding protein-7
(IGFBP7): A promising gene therapeutic for hepatocellular carcinoma
(HCC). Mol Ther. 21:758–766. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Georges RB, Adwan H, Hamdi H, Hielscher T,
Linnemann U and Berger MR: The insulin-like growth factor binding
proteins 3 and 7 are associated with colorectal cancer and liver
metastasis. Cancer Biol Ther. 12:69–79. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Yu Z, Tang Y, Hu D and Li J: Inhibitory
effect of genistein on mouse colon cancer MC-26 cells involved
TGF-beta1/Smad pathway. Biochem Biophys Res Commun. 333:827–832.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Satirapoj B and Adler SG: Prevalence and
management of diabetic nephropathy in western countries. Kidney Dis
(Basel). 1:61–70. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Tomino Y and Gohda T: The Prevalence and
management of diabetic nephropathy in asia. Kidney Dis (Basel).
1:52–60. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Chen L, Su W, Chen H, Chen DQ, Wang M, Guo
Y and Zhao YY: Proteomics for biomarker identification and clinical
application in kidney disease. Adv Clin Chem. 85:91–113. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Han Q, Zhang J, Wang Y, Li H, Zhang R, Guo
R, Li L, Teng G, Wang J, Wang T and Liu F: Thyroid hormones and
diabetic nephropathy: An essential relationship to recognize.
Nephrology (Carlton). 2018. View Article : Google Scholar
|
|
34
|
Varga ZV, Giricz Z, Liaudet L, Hasko G,
Ferdinandy P and Pacher P: Interplay of oxidative,
nitrosative/nitrative stress, inflammation, cell death and
autophagy in diabetic cardiomyopathy. Biochim Biophys Acta.
1852:232–242. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Flyvbjerg A: The role of the complement
system in diabetic nephropathy. Nat Rev Nephrol. 13:311–318. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Wada J and Makino H: Inflammation and the
pathogenesis of diabetic nephropathy. Clin Sci (Lond). 124:139–152.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Imasawa T, Obre E, Bellance N, Lavie J,
Imasawa T, Rigothier C, Delmas Y, Combe C, Lacombe D, Benard G, et
al: High glucose repatterns human podocyte energy metabolism during
differentiation and diabetic nephropathy. FASEB J. 31:294–307.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Li JJ, Kwak SJ, Jung DS, Kim JJ, Yoo TH,
Ryu DR, Han SH, Choi HY, Lee JE, Moon SJ, et al: Podocyte biology
in diabetic nephropathy. Kidney Int. Suppl:S36–S42. 2007.
View Article : Google Scholar
|
|
39
|
Zhang Y, Chen Y, Yang F and Zhou J: HBx
transfection limits proliferative capacity of podocytes through
cell cycle regulation. Acta Biochim Biophys Sin (Shanghai).
46:1016–1023. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Anders HJ, Vielhauer V and Schlondorff D:
Chemokines and chemokine receptors are involved in the resolution
or progression of renal disease. Kidney Int. 63:401–415. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Ferder L, Inserra F and Martinez-Maldonado
M: Inflammation and the metabolic syndrome: Role of angiotensin II
and oxidative stress. Curr Hypertens Rep. 8:191–198. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Yang Y, Gubler MC and Beaufils H:
Dysregulation of podocyte phenotype in idiopathic collapsing
glomerulopathy and HIV-associated nephropathy. Nephron. 91:416–423.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Madne TH and Dockrell MEC: Human podocytes
responses to alternatively spliced Extra domain A Fibronectin in
culture. Cell Mol Biol (Noisy-le-Grand). 64:45–52. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Rochman M, Travers J, Abonia JP, Caldwell
JM and Rothenberg ME: Synaptopodin is upregulated by IL-13 in
eosinophilic esophagitis and regulates esophageal epithelial cell
motility and barrier integrity. JCI Insight. 2:967892017.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Chen D, Yoo BK, Santhekadur PK, Gredler R,
Bhutia SK, Das SK, Fuller C, Su ZZ, Fisher PB and Sarkar D:
Insulin-like growth factor-binding protein-7 functions as a
potential tumor suppressor in hepatocellular carcinoma. Clin Cancer
Res. 17:6693–6701. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Watanabe J, Takiyama Y, Honjyo J, Makino
Y, Fujita Y, Tateno M and Haneda M: Role of IGFBP7 in diabetic
nephropathy: TGF-β1 induces IGFBP7 via smad2/4 in human renal
proximal tubular epithelial cells. PloS One. 11:e01508972016.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Chang YP, Sun B, Han Z, Han F, Hu SL, Li
XY, Xue M, Yang Y, Chen L, Li CJ and Chen LM: Saxagliptin
attenuates albuminuria by inhibiting podocyte
epithelial-to-mesenchymal transition via sdf-1α in diabetic
nephropathy. Front Pharmacol. 8:7802017. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Chen T, Zheng LY, Xiao W, Gui D, Wang X
and Wang N: Emodin ameliorates high glucose induced-podocyte
epithelial-mesenchymal transition in-vitro and in-vivo. Cell
Physiol Biochem. 35:1425–1436. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Du M, Wang Q, Li W, Ma X, Wu L, Guo F,
Zhao S, Huang F, Wang H and Qin G: Overexpression of FOXO1
ameliorates the podocyte epithelial-mesenchymal transition induced
by high glucose in vitro and in vivo. Biochem Biophys Res Commun.
471:416–422. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Chen Q, Yang W, Wang X, Li X, Qi S, Zhang
Y and Gao MQ: TGF-β1 induces emt in bovine mammary epithelial cells
through the TGFβ1/smad signaling pathway. Cell Physiol Biochem.
43:82–93. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Ibrini J, Fadel S, Chana RS, Brunskill N,
Wagner B, Johnson TS and El Nahas AM: Albumin-induced epithelial
mesenchymal transformation. Nephron Exp Nephrol. 120:e91–102. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Igietseme JU, Omosun Y, Nagy T, Stuchlik
O, Reed MS, He Q, Partin J, Joseph K, Ellerson D, George Z, et al:
Molecular pathogenesis of chlamydia disease complications:
Epithelial-mesenchymal transition and fibrosis. Infect Immun.
86:e005852017. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Wiggins RC: The spectrum of
podocytopathies: A unifying view of glomerular diseases. Kidney
Int. 71:1205–1214. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Yu D, Petermann A, Kunter U, Rong S,
Shankland SJ and Floege J: Urinary podocyte loss is a more specific
marker of ongoing glomerular damage than proteinuria. J Am Soc
Nephrol. 16:1733–1741. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Kamato D, Burch ML, Piva TJ, Rezaei HB,
Rostam MA, Xu S, Zheng W, Little PJ and Osman N: Transforming
growth factor-beta signalling: role and consequences of Smad linker
region phosphorylation. Cell Signal. 25:2017–2024. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Ning W, Tao L, Liu C, Sun J, Xiao Y, Hu J,
Chen J, Zheng X and Wang W: Effect of enalapril on the expression
of TGF-beta1, p-Smad2/3 and Smad7 in renal interstitial fibrosis in
rats. Zhong Nan Da Xue Xue Bao Yi Xue Ban. 34:27–34. 2009.(In
Chinese). PubMed/NCBI
|