Introduction
Epilepsy is a chronic brain disease commonly found
in clinical practice, which can occur due to combined action of
various factors. Epidemiological studies have found that the
incidence of epilepsy shows an upward trend year by year, and China
is also a high-incidence area of epilepsy, where nearly half of
epileptic patients have refractory epilepsy (1,2). At
present, the main treatment for epilepsy is drug therapy, and more
than 10 kinds of first-line antiepileptic drugs are used in the
clinic. However, many epilepsy types cannot be effectively
controlled, leading to serious financial burden on both the
patients and society (3). The onset
of epilepsy generally includes three processes, namely, epileptic
brain injury, epileptic seizure latency and recurrence of epilepsy
(4). The occurrence of epilepsy
involves a variety of molecular, cellular and neuronal changes, and
the intervention in relevant molecular pathways involved in the
occurrence of epilepsy can effectively prevent the incidence of
epilepsy, reducing the exaltation of neurons (5).
Controlling and reducing the early onset of epilepsy
is able to effectively prevent the recurrence of epilepsy later and
reduce the damage caused by epilepsy to the cognitive and memory
functions in patients (6). A study
by Bowen et al (7) found that
patients with epilepsy have a certain degree of cognitive, learning
and memory impairment, which may be closely related to the damage
in hippocampal neurons, and a study by Bardai et al
(8) discovered that the excitability
of hippocampal neurons is correlated with synaptic plasticity, the
formation of a new synapsis is sure to build up an excited loop
that can significantly increase the sensitivity of epileptic
seizures, but increased synaptic plasticity will lead to neuronal
death. Mitochondrion plays an important role in the process of
apoptosis. A large number of studies have proven that mitochondrial
dysfunction often occurs in neurons of brain tissues with various
diseases such as cerebral ischemia, depression and Parkinson's
disease, and is closely associated with secondary damage occurring
after cerebral ischemia; mitochondrion is involved in the energy
metabolism of eukaryotes, and mitochondrial damage will lead to
significant reduction in its production capacity, resulting in
insufficient energy supply in cells (9,10). A
study by McFarland et al (11) found that epileptic seizures can lead
to a certain degree of damage to mitochondrial functions, but the
mechanism has not been studied. This study explored the
relationship between the occurrence of epilepsy and the
mitochondrial damage in hippocampal neurons and the possible
molecular mechanism by establishing rat models of epilepsy.
Materials and methods
Animals and grouping
A total of 30 male Sprague-Dawley (SD) rats weighing
250–280 g were purchased from Guangdong Medical Laboratory Animal
Center (experimental animal production license number SCXK
2016–0002). These rats were raised in a specific pathogen-free
(SPF) animal room at 22–27°C, with a humidity of 50–60% and a 12 h
light/12 h dark cycle. All animals had free access to food and
water. After adapting to SPF environment for one week, SD rats were
randomly divided into normal group (n=15) and model group (n=15),
and pretreated with lithium chloride (intraperitoneal injection) at
24 h before experiment. During the experiment, rats in model group
were given pilocarpine (30 mg/kg) to establish rat models of
epilepsy. After 60 min of epileptic seizures, rats in model group
were intraperitoneally injected with diazepam (10 mg/kg) to
terminate reaction, and rats in normal group were injected with the
same dose of saline.
The provisions for experimental animals in the Guide
for the Care and Use of Laboratory Animals by the National Research
Council were strictly abided in all operations on animals. The
experimental scheme was approved by the Experimental Animal Ethics
Committee in Daqing Longnan Hospital (Daqing, China).
Determination of epileptiform
behavior
After modeling, behavioral changes of rats in the
control and model groups were recorded. The onset of epilepsy was
determined via the Racine seizure scale, namely, no abnormalities
such as convulsion were observed: level 0; regular chewing and
blinking, and convulsions of mouth and face were found: level I;
spastic movement was found in head and neck: level II; unilateral
forelimb convulsion was found, without hind limb erection: level
III; bilateral forelimb convulsion combined with body erection was
observed: level IV; orthotonos, dorsiflexion, falling down and
general tetany-clonus were found: level V. Levels IV–V were
considered as epileptic seizures, and modeling was successful.
During epileptic seizures, electroencephalogram (EEG) was used to
record the brain waves of hippocampus of rats in two groups to
observe the presence of cluster spike discharge: rats in two groups
were anesthetized and placed on a brain solid positioner to expose
the skull and bregma, and then electrodes were inserted into the
hippocampi of rats (3.8 mm away from the bregma, 2 mm beside the
midline, and 5 mm under the dura mater), and the reference
electrode was placed subcutaneously on the counter mastoid.
Nissal staining
After behavioral detection, each rat in the two
groups was anesthetized, its thoracic cavity was opened to expose
the heart, and then cardiac perfusion was performed with 100 ml
normal saline. After the lungs on both sides and the liver of the
rat were obviously whitened, 100 ml paraformaldehyde was used for
fixation, and the brain tissue of the rat was taken and
continuously fixed in 4% paraformaldehyde for 24 h, followed by
gradient dehydration with 20 and 30% sucrose solutions,
respectively. The brain tissue of the rat was sectioned,
dehydrated, embedded in paraffin, sliced serially and coronally
using a paraffin slicing machine, attached to a treated glass
slide, and dried at 60°C for 2 h. Then, Nissl staining was
performed, that is, the section was dewaxed and hydrated, subjected
to hyalinization and dehydration, washed with distilled water 3
times (3 min/time), and stained with 1% toluidine blue for 6 min.
Subsequently, the section was washed with distilled water 3 times
(3 min/time), placed in 70% ethanol for 2 min and then 95% ethanol
for 5 min, and placed in absolute ethanol for color separation
until the nucleus was light blue, and the background was colorless.
The section was then immersed in xylene for 5 min, mounted, and
placed under a microscope (Olympus, Tokyo, Japan) for
observation.
Measurement of adenosine triphosphate
(ATP) level in hippocampus tissue
The level of ATP in the hippocampus tissue of each
rat in two groups was measured by a rat ATP assay kit. The tissue
was lysed by adding lysis buffer (ratio of tissue to lysis buffer:
100 mg:1 ml) and homogenized with an ultrasonic homogenizer under
an ice bath until the tissue was completely lysed without
macroscopic tissue fragments. Then, the tissue was centrifuged at
9,800 × g for 10 min at 4°C, and the supernatant was collected and
stored at −20°C for subsequent experiments. ATP test solution (100
µl) was added into a standard tube and a detection tube in the
dark, and then the two tubes were placed at room temperature for 5
min of reaction to degrade ATP. After that, ATP standard substance
with different concentrations was added into the standard tube,
quickly mixed and placed on a microplate reader (Bio-Rad
Laboratories, Inc., Hercules, CA, USA) to determine the relative
light unit (RLU) and counted photons per minute (CPM) values of the
sample and a standard curve was drawn; different sets of samples
were added to the detection tube, and ATP concentration in the
sample was measured using a microplate reader.
Ribonucleic acid (RNA) extraction and
quantitative polymerase chain reaction (qPCR)
After the rat hippocampus tissue was obtained, the
tissue (100 mg) was added with TRIzol lysis solution (1 ml) for
homogenization in the ice bath, added with 300 µl chloroform, and
thoroughly mixed. Then, the tissue was placed on ice for 10 min and
centrifuged at 9,800 × g and 4°C for 15 min. After separation of
the supernatant, an equal amount of isopropanol was added, placed
at room temperature for 5 min, and centrifuged at 4°C and 9,800 × g
for 10 min. The supernatant was discarded, and 1 ml 75% ethanol was
added for washing, followed by centrifugation at 4°C and 9,800 × g.
After that, the supernatant was discarded, 50 µl
diethylpyrocarbonate (DEPC) water was added for dissolution, and
then RNA was obtained. The optical density (OD) value and RNA
integrity were then tested, and RNA was stored at −20°C until use.
The RNA was reverse transcribed into complementary deoxyribonucleic
acid (cDNA) using a reverse transcription kit, acting as a
template. After adding qPCR reaction system, the reaction was
carried out under the following conditions: 95°C for 10 sec, 95°C
for 5 sec, 60°C for 15 sec and 72°C for 15 sec, with 39 cycles in
total, and then 72°C for 5 sec and 95°C for 2 min to stop the
reaction. Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) was used
as internal reference. The expression level of the corresponding
gene was calculated by the 2−ΔΔCq method (12). The primers were synthesized by
Tiangen Biotech Co., Ltd. The sequences are shown in Table I.
 | Table I.PCR primers. |
Table I.
PCR primers.
| Gene | Sequence |
|---|
| Fatty acid | F:
5′-AGGCTAACCCCACTCTATGAATC-3′ |
| synthetase (Fas) | R:
5′-TCTTGCCTTTGGTGGACTA-3′ |
| Fas ligand
(FasL) | F:
5′-AGAGGAACCACAGCACA-3′ |
|
| R:
5′-TCACTCCAGAAAGCAGGACA-3′ |
| Caspase-3 | F: 5′-
ATGGACAACAACGAAACCTC-3′ |
|
| R:
5′-TTAGTGATAAAAGTACAGTTCTT-3′ |
| D-loop | F:
5′-AATCTACCATCCTCCGTG-3′ |
|
| R:
5′-GACTAATGATTCTTCACCGT-3′ |
| GAPDH | F:
5′-CAGTGCCAGCCTCGTCTCAT-3′ |
|
| R:
5′-AGGGCCATCCACAGTCTTC-3′ |
Western blot analysis
The rat hippocampus tissue was collected and added
with lysis solution for homogenization until no macroscopic tissue
was observed and then, centrifuged at 9,800 × g at 4°C for 10 min.
The supernatant was removed and the total protein concentration in
the hippocampus tissue was measured using a bicinchoninic acid
(BCA) protein assay kit. Sample loading system with the same
concentration was prepared, and the sample was loaded. Sodium
dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) was
applied to separate protein, and then protein was transferred onto
a polyvinylidene fluoride (PVDF) membrane under a constant pressure
of 90 V and blocked with newly prepared 5% non-fat dry milk for 2
h. After that, target band was cut down based on the size of
protein band, and incubated with rabbit anti-rat fatty acid
synthetase (Fas), Fas ligand (FasL), caspase-3 and GAPDH
polyclonalantibodies (1:1000; cat. nos.: ab82419, ab15285,
ab13847,ab9485; Abcam; Cambridge, MA, USA) at 4°C overnight. Then,
the band was washed with Tris-buffered saline with
Tween®−20 (TBST) 5 times (5 min/time), and incubated
with goat anti-rabbit secondary polyclonal antibody (1:1200; cat.
no.: ab6721; Abcam) at room temperature for 2 h, and washed with
TBST 3 times (10 min/time). Electrochemiluminescence (ECL) mixture
was prepared, followed by tableting in the dark. Then, luminescence
solution was added for color development, and film was dried, and
subjected to software processing. Fas/GAPDH, FasL/GAPDH and
caspase-3/GAPDH indicated the relative expression levels of
proteins in each group.
Statistical analysis
Data in this study are expressed as mean ± standard
deviation. Statistical Product and Service Solutions (SPSS) 19.0
software (SPSS Inc., Chicago, IL, USA) was employed for data
analysis. Student's t-test was used for comparisons between two
groups. Pearsons analysis was adopted for inter-factor
correlations. P<0.05 suggested that the difference was
statistically significant.
Results
Evaluations of epileptiform symptoms
in rats
After rats in both groups received inducing
treatment, symptoms such as perspiration, piloerection and eye
congestion were observed at approximately 10 min after modeling in
rats of the model group, and epileptiform behavior of rats was
evaluated via Racine seizure scale. The results revealed that
modeling was successful in 15 rats, with typical epileptiform
symptoms, and all rats in the control group had typical
epileptiform symptoms (Table II).
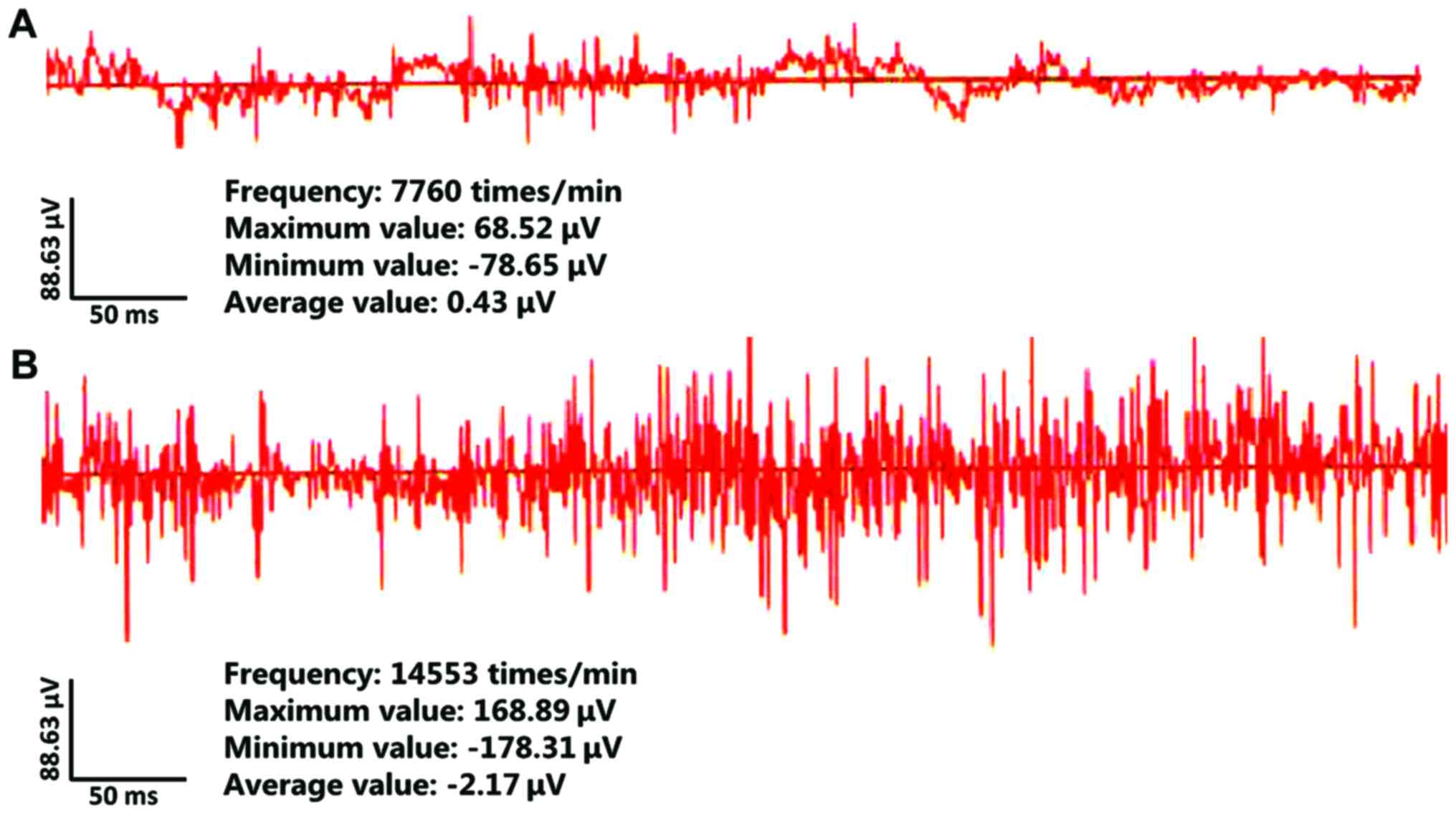
EEG was applied to detect epileptic rat cluster spike discharge.
The results indicated that typical cluster spike discharge was
found in the hippocampus of rats in the model group, and no
abnormal discharge was observed in the rats in control group
(Fig. 1).
| Table II.Scores for epileptic seizures. |
Table II.
Scores for epileptic seizures.
| Groups | I | II | III | IV | V |
|---|
| Control group
(n) | 13 | 2 | 0 | 0 | 0 |
| Model group (n) | 0 | 0 | 1 | 5 | 9 |
| P-value |
|
|
| <0.05 | <0.05 |
| t-value |
|
|
| 0.586 | 0.678 |
Injuries in hippocampal neurons
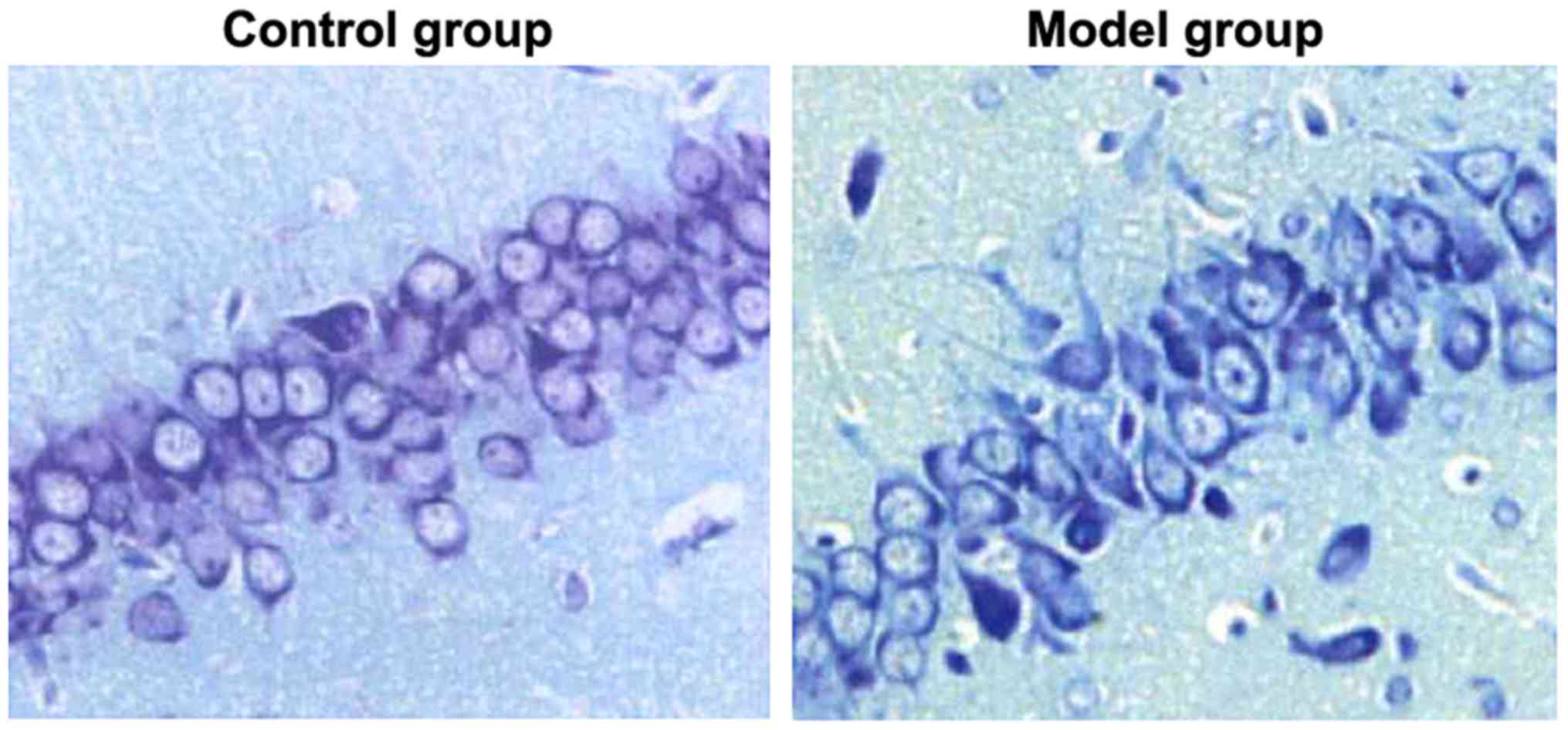
Nissl staining was used to detect the injuries in
hippocampal neurons of rats in the two groups. As shown in Fig. 2, the hippocampal neurons of rats in
the control group had compact arrangement, complete cell
morphology, clear cell structure and abundant Nissl bodies in the
cytoplasm, and the hippocampal neurons of rats in the model group
were loosely arranged, with fuzzy cell outline, shrunken cell body,
pyknotic nuclei, and significantly reduced Nissl bodies in
cytoplasm. It meant that hippocampal neurons in rat models of
epilepsy had severe damage.
Detection of mitochondrial function in
hippocampal neurons
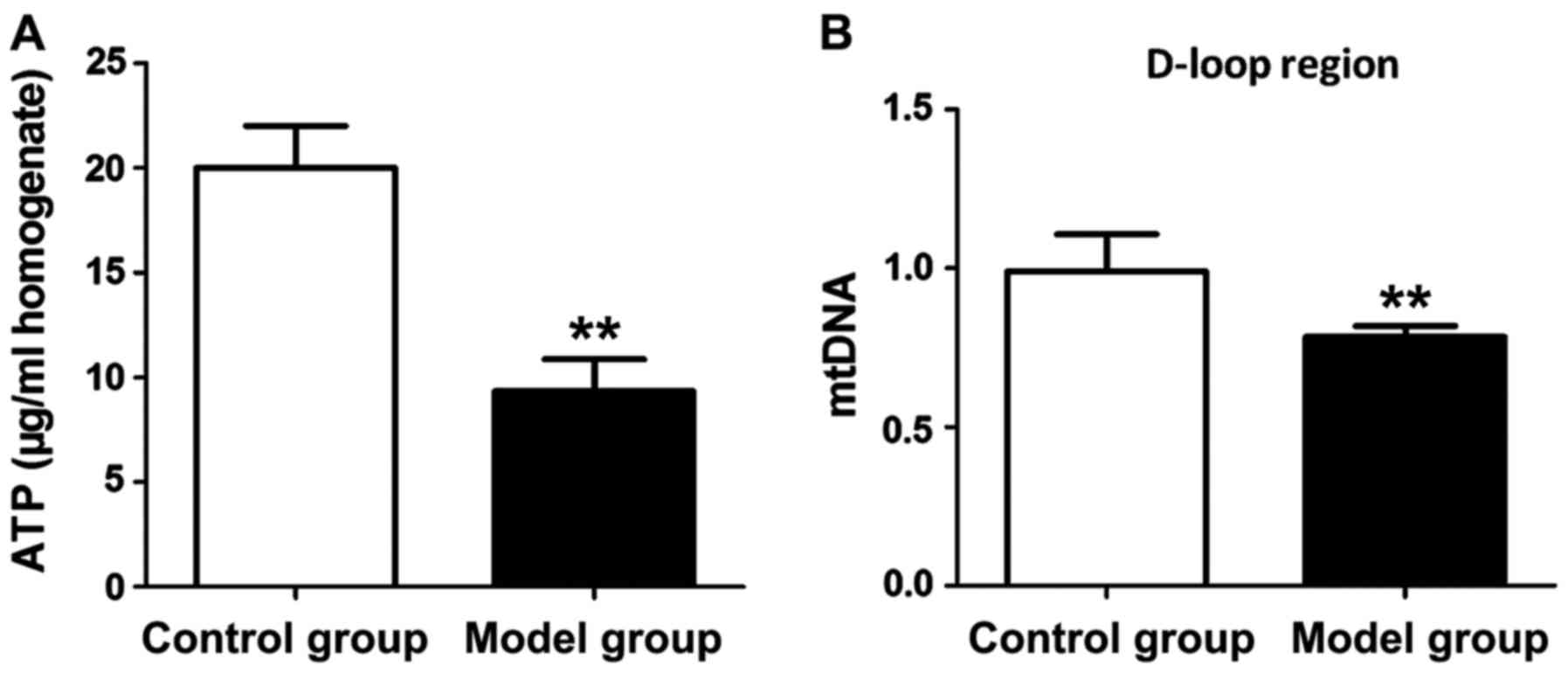
ATP levels and mitochondrial DNA levels in
hippocampus tissues of rats in both groups were measured to
evaluate the mitochondrial function of hippocampal neurons.
According to Fig. 3, both ATP level
and mitochondrial DNA level in hippocampus tissue of the rat in the
model group were significantly lower than those in the control
group (P<0.01).
Measurements of mRNA and protein
expression levels
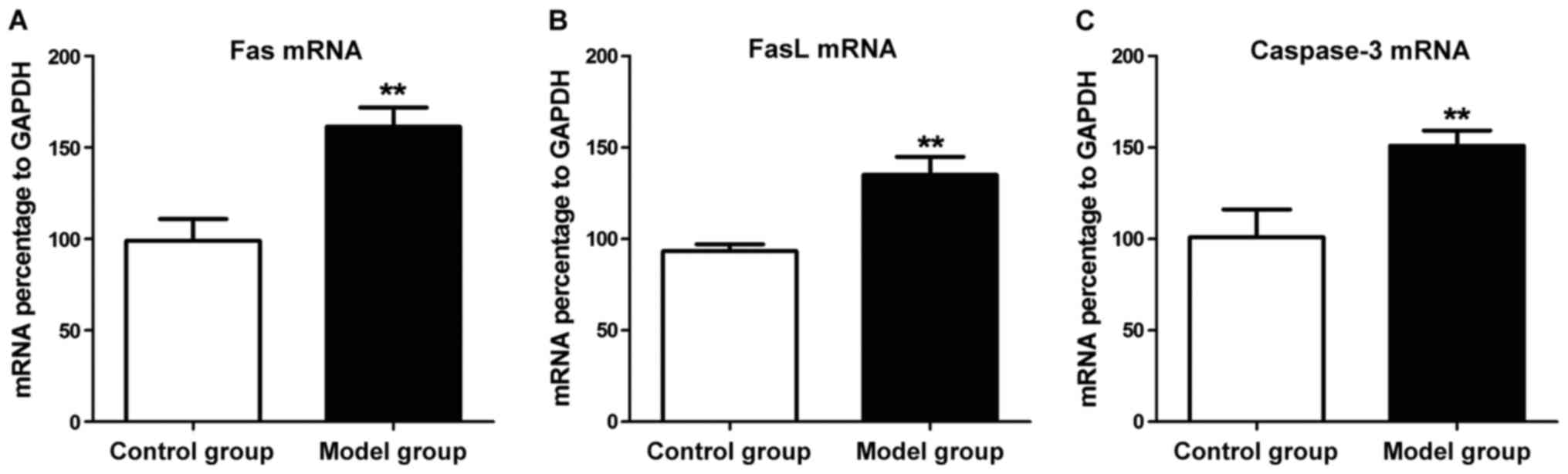
The expression levels of related genes in
hippocampus tissues of rats in two groups were detected by qPCR.
The results revealed that mRNA expression levels in hippocampus
tissue of the rat in the model group were overtly higher than those
in the control group (P<0.01) (Fig.
4). Western blotting was used to detect related protein
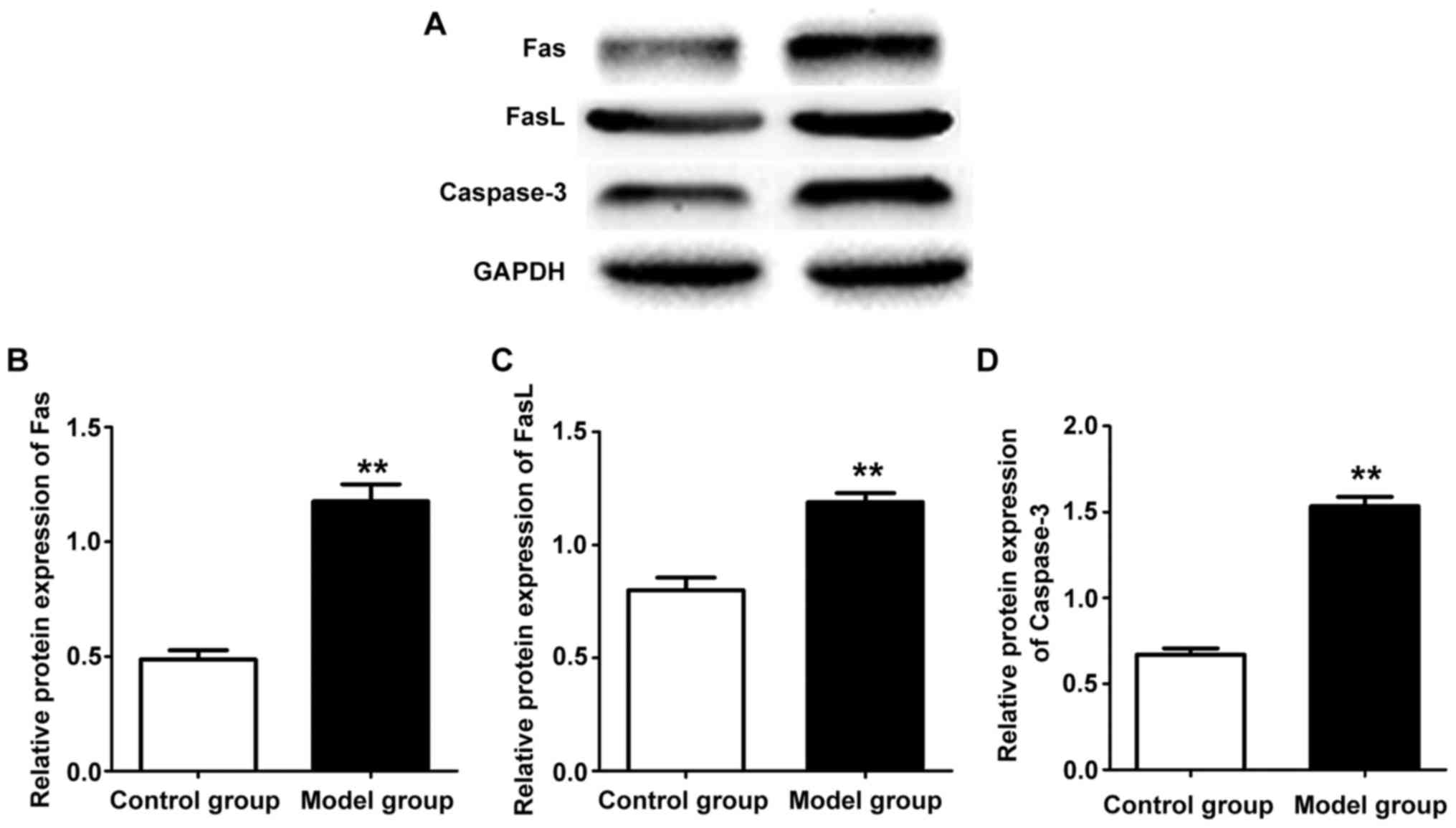
expression levels in hippocampus tissue of rats in the two groups.
The results showed that protein expression levels in hippocampus
tissue of the rat in the model group were increased evidently
compared with those in the control group (P<0.01) (Fig. 5). Western blot results show the
expression of Fas, FasL and caspase-3 protein is consistent with
mRNA expression.
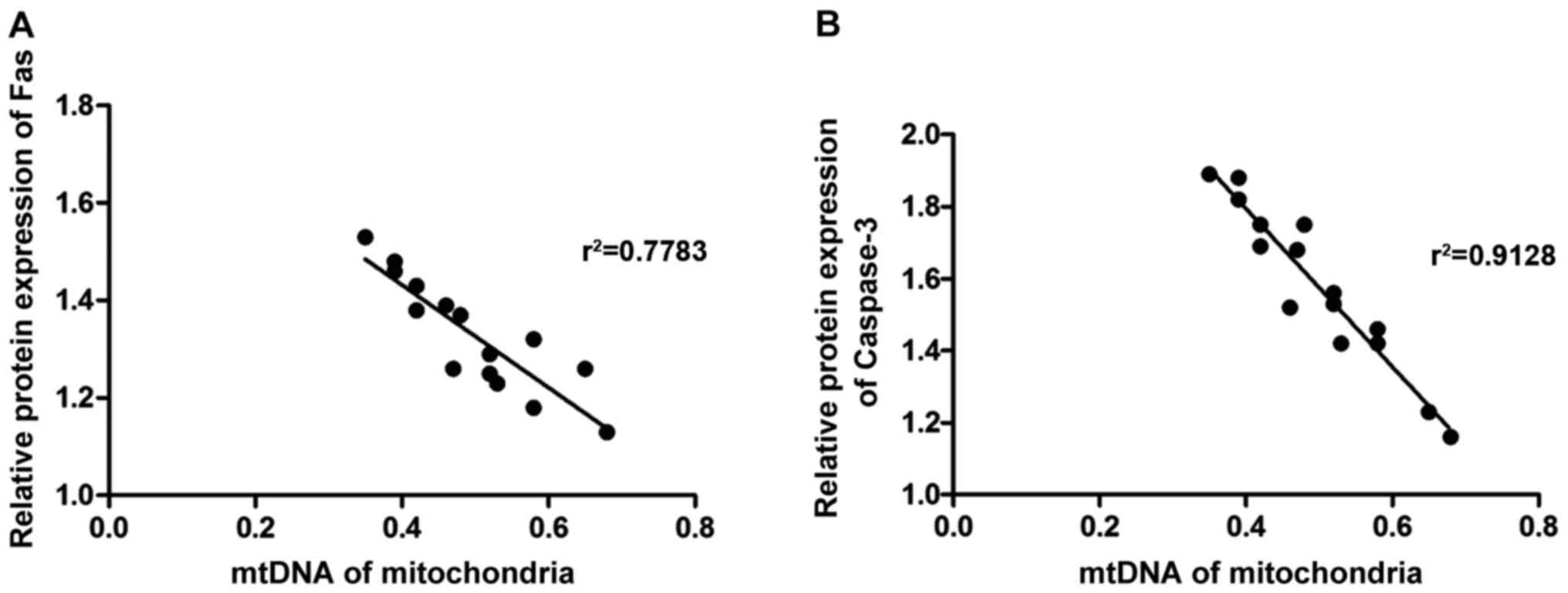
Correlation analysis
Pearsons analysis was used to detect the
correlations of mitochondrial dysfunction with the expression of
Fas and caspase-3. As shown in Fig.
6, mitochondrial DNA level in rat hippocampal neurons was
negatively correlated with the protein expression of Fas and
caspase-3 (r2=222, P<0.05).
Discussion
Numerous studies have suggested that phases of
epilepsy induction and epilepticus are able to lead to changes in
neuronal ultramicro results, resulting in mitochondrial
dysfunction. A study by Chang et al (13) found that induction of epilepticus in
rats can significantly reduce the activities of reduced form of
nicotinamide-adenine dinucleotid (NADH) cytochrome c
reductase and aconitase in hippocampus, and with the alleviation of
symptoms of epilepsy, enzyme activities in vivo will
gradually recover. A study by McDonald et al (14) discovered that obvious structural
changes are observed in mitochondria of hippocampal neurons during
epileptic seizures, which may be manifested as rupture of membrane
and reduction of transmembrane potential. Epileptic seizures are
often accompanied by anomalies in oxygenic respiratory chain
complex I, resulting in massive electron leakage, thereby producing
reactive oxide species and damaging nerve cells. In addition,
seizures can overtly increase the oxidative stress response and
further aggravate mitochondrial damage. The above changes are
likely to result in decreased mitochondrial activity, affect
mitochondrial transcription and translation processes, and lead to
neuron necrosis at epilepsy sites due to a serious shortage of ATP
(15). In this study,
intraperitoneal injection of lithium chloride-pilocarpine was
carried out to induce epilepticus in rats, a large number of
necrotic hippocampal neurons were found in epileptic rats through
Nissal staining, and the simultaneous detections of ATP level and
mitochondrial DNA level in hippocampus tissues revealed that
mitochondria in hippocampus tissue had obvious damage. The results
are consistent with the corresponding reports. As a cholinergic
receptor agonist, pilocarpine can excite M-type cholinergic
receptors to cause excited paradoxical discharge of neurons,
thereby effectively simulating the cause of seizures and
establishing rat models of epilepsy. The modeling method is widely
used because it has high safety, clear epileptogenic process,
obvious symptoms of induced epilepsy, and human-like behavior and
changes of EEG (16).
Mitochondrion can mediate the occurrence of
apoptosis, Fas is a membrane protein involved in the process of
apoptosis, whose ligand is FasL, and the combination of the two can
form a trimer with apoptosis-inducing activity (17–19). A
study of Hu et al (20)
showed that Fas/FasL is able to regulate apoptosis through
Fas-associated protein with death domain (FADD), and the activation
of FADD will lead to obviously increased expression and activation
of caspase-8 in cells. Activated caspase-8 can further activate
caspase-3, leading to caspase cascade reaction and thus resulting
in apoptosis (21). This study found
that both mRNA and protein expression levels of Fas and FasL in
hippocampus tissue of epileptic rats were significantly increased,
indicating that the occurrence of epilepsy can activate Fas/FasL
signaling pathway, and further study found that caspase-3
expression level was also evidently elevated. The results suggest
that epilepsy-induced neuronal apoptosis is caused by Fas/FasL
pathway via FADD. Correlation analysis was used to evaluate the
relationships between the expression levels of Fas and caspase-3
and mitochondrial dysfunction. The results showed that there were
negative correlations between the expressions of Fas and caspase-3
and the level of mitochondrial DNA, that is, mitochondrial
dysfunction was closely related to the signaling pathway.
In conclusion, the occurrence of epilepsy in rats
may lead to mitochondrial dysfunction in rat hippocampal neurons,
thus resulting in apoptosis of hippocampal neurons, which may be
realized through activating Fas/FasL signaling pathway and
increasing caspase-3 expression.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
JF determined epileptiform behavior. LF performed
western blot analysis. JF and GZ helped with Nissal staining. All
authors have read and approved the final manuscript.
Ethics approval and consent to
participate
The provisions for experimental animals in the Guide
for the Care and Use of Laboratory Animals by the National Research
Council were strictly abided in all operations on animals. The
experimental scheme was approved by the Experimental Animal Ethics
Committee in Daqing Longnan Hospital (Daqing, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Sahoo SS, Lhatoo SD, Gupta DK, Cui L, Zhao
M, Jayapandian C, Bozorgi A and Zhang GQ: Epilepsy and seizure
ontology: Towards an epilepsy informatics infrastructure for
clinical research and patient care. J Am Med Inform Assoc.
24:82–89. 2014. View Article : Google Scholar
|
|
2
|
Liu J, Reeves C, Michalak Z, Coppola A,
Diehl B, Sisodiya SM and Thom M: Evidence for mTOR pathway
activation in a spectrum of epilepsy-associated pathologies. Acta
Neuropathol Commun. 2:712014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Speed D, OBrien TJ, Palotie A, Shkura K,
Marson AG, Balding DJ and Johnson MR: Describing the genetic
architecture of epilepsy through heritability analysis. Brain.
137:55–69. 2014. View Article : Google Scholar
|
|
4
|
Kaiboriboon K, Bakaki PM, Lhatoo SD and
Koroukian S: Incidence and prevalence of treated epilepsy among
poor health and low-income Americans. Neurology. 80:1942–1949.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ying Z, Najm I, Nemes A, Pinheiro-Martins
AP, Alexopoulos A, Gonzalez-Martinez J and Bingaman W:
Growth-associated protein 43 and progressive epilepsy in cortical
dysplasia. Ann Clin Transl Neurol. 1:453–461. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wirrell EC, Grossardt BR, Wong-Kisiel LC
and Nickels KC: Incidence and classification of new-onset epilepsy
and epilepsy syndromes in children in Olmsted County, Minnesota
from 1980 to 2004: A population-based study. Epilepsy Res.
95:110–118. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Bowen JM, Snead OC, Chandra K, Blackhouse
G and Goeree R: Epilepsy care in Ontario: An economic analysis of
increasing access to epilepsy surgery. Ont Health Technol Assess
Ser. 12:1–41. 2012.PubMed/NCBI
|
|
8
|
Bardai RJ, Lamberts RJ, Blom MT, Spanjaart
AM, Berdowski J, van der Staal SR, Brouwer HJ, Koster RW, Sander
JW, Thijs RD, et al: Lamberts, marieke T. epilepsy is a risk factor
for sudden cardiac arrest in the general population. PLoS One.
7:14–23. 2012. View Article : Google Scholar
|
|
9
|
Kovac S, Kostova Dinkova AT, Herrmann AM,
Melzer N, Meuth SG and Gorji A: Metabolic and homeostatic changes
in seizures and acquired epilepsy - mitochondria, calcium dynamics
and reactive oxygen species. Int J Mol Sci. 18:E19352017.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Fahrner JA, Liu R, Perry MS, Klein J and
Chan DC: A novel de novo dominant negative mutation in DNM1L
impairs mitochondrial fission and presents as childhood epileptic
encephalopathy. Am J Med Genet A. 170:2002–2011. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
McFarland KN, Liu J, Landrian I, Zeng D,
Raskin S, Moscovich M, Gatto EM, Ochoa A, Teive HA, Rasmussen A, et
al: Repeat interruptions in spinocerebellar ataxia type 10
expansions are strongly associated with epileptic seizures.
Neurogenetics. 15:59–64. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2−ΔΔCT method. Methods. 25:402–408. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Chang HJ, Liao CC, Hu CJ, Shen WW and Chen
TL: Psychiatric disorders after epilepsy diagnosis: A
population-based retrospective cohort study. PLoS One.
8:e599992013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
McDonald TS, Carrasco-Pozo C, Hodson MP
and Borges K: Alterations in cytosolic and mitochondrial
(U-13C)glucose metabolism in a chronic epilepsy mouse model.
eNeuro. Mar 9–2017.(Epub ahead of print). doi:
10.1523/ENEURO.0341-16.2017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Rowley S, Liang LP, Fulton R, Shimizu T,
Day B and Patel M: Mitochondrial respiration deficits driven by
reactive oxygen species in experimental temporal lobe epilepsy.
Neurobiol Dis. 75:151–158. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Meng DW, Liu HG, Yang AC, Zhang K and
Zhang JG: Stimulation of anterior thalamic nuclei protects against
seizures and neuronal apoptosis in hippocampal CA3 region of kainic
acid-induced epileptic rats. Chin Med J (Engl). 129:960–966. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yamada A, Arakaki R, Saito M, Kudo Y and
Ishimaru N: Dual role of Fas/FasL-mediated signal in peripheral
immune tolerance. Front Immunol. 8:4032017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Liu G, Yuan Y, Long M, Luo T and Bian J:
Beclin-1-mediated autophagy protects against cadmium-activated
apoptosis via the Fas/FasL pathway in primary rat proximal tubular
cell culture. Sci Rep. 8:3941–3948. 2017.
|
|
19
|
Zhou X, Hong T, Yu Q, Nie S, Gong D, Xiong
T and Xie M: Exopolysaccharides from Lactobacillus plantarum
NCU116 induce c-Jun dependent Fas/Fasl-mediated apoptosis via TLR2
in mouse intestinal epithelial cancer cells. Sci Rep. 7:142472017.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Hu L, Chen L, Yang G, Li L, Sun H, Chang
Y, Tu Q, Wu M and Wang H: HBx sensitizes cells to oxidative
stress-induced apoptosis by accelerating the loss of Mcl-1 protein
via caspase-3 cascade. Mol Cancer. 10:432011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhou X and Hong T: Exopolysaccharides from
Lactobacillus plantarum NCU116 induce c-Jun dependent
Fas/Fasl-mediated apoptosis via TLR2 in mouse intestinal epithelial
cancer cells. Retrovirology. 5:14–35. 2008.PubMed/NCBI
|