Introduction
Immune thrombocytopenia (ITP) is a common
hematologic disorder characterized by isolated thrombocytopenia,
which causes bleeding in the skin and mucosa. According to the
disease duration, ITP can be classified into three types, including
newly diagnosed, persistent and chronic, and chronic ITP (cITP)
(1). In ~1/3 of ITP cases, the
duration of thrombocytopenia will extend over 12 months, which is
defined as chronic ITP (2). ITP
occurs in children and adults; however, the disease is typically
chronic in adults (3). The incidence
primary ITP accounts for 3.3/100,000 adults per year (3). Although recent progress on the
development of thrombopoietin receptor agonists have changed the
management of chronic disease (4),
the detailed underlying mechanisms involved in the pathophysiology
of cITP remain poorly understood.
A dysfunctional proliferation of autoreactive T
cells, including T helper (Th)2, Th17, Th22 and T follicular helper
(TFH) cells, has been suggested to be responsible for the loss of
tolerance to self-platelet antigens in ITP. TFH cells, a distinct
subset of CD4+ T cells, specialize in providing critical
assistance to germinal center (GC) B cells (5). TFH cells contribute to the generation
and maintenance of GCs, support Ig class switching and assist in
differentiation of GCs into memory B cells and long-lived plasma
cells (6). Immune-mediated disease
may occur due to the inability of TFH cells to maintain immune
homeostasis (7). Notably, the role
of TFH cells in autoimmune diseases has been studied in systemic
lupus erythematosus (SLE) mouse models (7). Furthermore, the frequency of TFH cells
has been indicated to be downregulated in the peripheral blood of
patients with SLE (8). Additionally,
the frequency of TFH is associated with the proportion of GC B
cells and plasma cells. Antiplatelet antibodies of IgG, IgA and IgM
produced by B cells facilitate phagocytosis by macrophages. Of
note, platelet destructions are more severe with IgG compared with
other isotypes (9).
Further research on the roles of TFH cells in the
development of adult cITP are still required. The limited studies
on pre- and post-treatment (intravenous immunoglobulin and
corticosteroids) comparisons of TFH cells and the small samples
sizes of patients assessed may bias the possible role of TFH cells
in ITP. In the present study, the functional change of circulating
TFH cells was investigated using cell counting, CD4+ TFH
cell-associated cytokine profiles and expression determination of
transcription factors, including Bcl-6, c-Maf, Blimp-1 and PD-1 in
a cohort of patients with cITP at pre- and post-treatment time
points. The present findings may provide useful insights on the
role of TFH cells in cITP.
Materials and methods
Patients and healthy volunteers
A total of 54 patients diagnosed with cITP who
received treatment with intravenous immunoglobulin, corticosteroids
or a combination, were enrolled in the study for blood analyses.
There were 32 female patients (59%) and 22 male patients (41%),
aged 28–61 years. Only patients who fit the international
guidelines for ITP (10) were
included in the present study. Cases that refused data collection
or had autoimmune disorders were excluded from the present study.
Based on the efficacy of the treatment, the patients were divided
into responders and non-responders. A total of 30 healthy controls
were recruited at the same time as patients, there were 16 female
(53.3%) and 14 male (46.7%), aged 24–60 years. A total of 5 ml
blood samples were collected from the patients at the Hematology
Outpatient Department in The First Affiliated Hospital of Soochow
University (Suzhou, China) between January 2016 and July 2017.
Ethylenediaminetetraacetic acid (EDTA)-stabilized venous blood (5
ml) was obtained from patients and healthy adults. Whole blood (2
ml) samples were stored at 4°C for flow cytometric analysis. Serum
isolated from venous blood samples was used to investigate the
change of CD4+ TFH cell-associated factors by comparing
responders (n=30) to non-responders (n=24). Peripheral blood
mononuclear cells (PBMCs) were isolated from blood samples using
gradient centrifugation (3,000 × g/min for 5 min at room
temperature) and stored at −80°C for further use. Patients who did
not achieve a platelet count >30 G/l were considered as
non-responders. Expression of serum platelet-associated
immunoglobulin G (PAIgG) in all patients was examined. All
participants provided their informed consent in accordance with the
declaration of Helsinki. The study was approved by the Ethics
Committee of The First Affiliated Hospital of Soochow University
(Suzhou, China).
Flow cytometry
The following antibodies were used for flow
cytometry analysis: Cluster of differentiation (CD)4-phycoerythrin
(PE)-CY5, CD3-PE-CY7 (Beckman Coulter, Inc., Miami, FL, USA), C-X-C
chemokine receptor type 5 (CXCR5)-fluorescein and inducible
costimulatory molecule (ICOS)-PE (BioLegend, San Diego, CA, USA).
Whole blood (50 µl) was incubated with specific antibodies for 30
min at room temperature. Following incubation, red blood cells were
lysed and washed twice by phosphate-buffered saline (PBS) solution.
Cells were acquired on Beckman Coulter FC500 and analyzed with CXP
analysis software 2.2 (Beckman Coulter, Inc.). For each tube, at
least 10,000 events were collected in a gate created around the
viable lymphocyte population. Data were presented as percentage of
cells in the lymphocyte population. Flow cytometric measurement of
PAIgG was performed using LEGENDplex Human Immunoglobulin Isotyping
Panel-IgGs (4-plex) with V-bottom Plate (cat. no. 740715;
BioLegend). The platelet-rich plasma (PRP) was obtained by
centrifugation at 180 × g for 15 min. PRP was washed twice with PBS
containing 10 mM EDTA and 0.5% bovine albumin. Notably, the
platelet concentration was adjusted to 1×108/ml. The
platelet population was gated with forward scatter light and side
scatter light. Quantification of four human immunoglobulins,
including IgG1, IgG2, IgG3 and IgG4 was performed according to the
manufacturer's protocol.
ELISA detection of cytokines
Serum IL-2, IL-4, IL-10 and IL-21 levels were
detected using ELISA with a xMark Microplate Absorbance
Spectrophotometer (Bio-Rad Laboratories, Inc., Hercules, CA, USA)
according to manufacturer's protocol. All samples were analyzed in
duplicate. The ELISA kits used in the experiment were Human IL-2
ELISA MAX™ Deluxe (cat. no. 431804), Human IL-4 ELISA MAX™ Deluxe
(cat. no. 430304), Human IL-10 ELISA MAX™ Standard (cat. no.
430601) and Human IL-21 ELISA MAX™ Deluxe (cat. no. 433804), all
purchased from BioLegend.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA from PBMCs was extracted using an AllPrep
RNA/DNA/miRNA Universal kit (Qiagen AB, Sollentuna, Sweden)
according to the manufacturer's instructions. cDNA was synthesized
using an RT-qPCR kit (Thermo Fisher Scientific, Inc., Waltham, MA,
USA). qPCR was performed on an Applied Biosystems 7500 Real-Time
PCR System (Applied Biosystems; Thermo Fisher Scientific, Inc.).
The sequences of the primers used were as follows: Bcl-6, forward
5′-AGTTTATTAAGGCCAGTGA-3′ and reverse 5′-GATAGGCCATGATGTCTT-3′;
c-Maf, forward 5′-ACTGGCAATGAGCAACTCCG-3′ and reverse
5′-GCTGATGATGCGGTCGGTCT-3′; B lymphocyte-induced maturation
protein-1 (Blimp-1), forward 5′-TCCAGCACTGTGAGGTTTCA-3′ and reverse
5′-TCAAACTCAGCCTCTGTCCA-3′; and PD-1, forward
5′-GACAACGCCACCTTCACCT-3′ and reverse 5′-GCTTGTCCGTCTGGTTGCT-3′.
GAPDH, forward 5′-AATCCCATCACCATCTTCCA-3′ and reverse
5′-TGGACTCCACGACGTACTCA-3′. qPCR was performed with SYBR Select
Master Mix (Thermo Fisher Scientific Inc.) and 500 nM forward and
reverse primers. Thermal cycle conditions used were as follows:
50°C for 2 min and 95°C for 10 min, followed by 40 cycles at 95°C
for 1 sec and 60°C for 30 sec Gene expression values were
calculated by the comparative threshold cycle method (11). Stable expressed GAPDH served as
endogenous control.
Response criteria
Respondents were classified as responders who were
responding to intravenous immunoglobulin and corticosteroids
therapy, and non-responders. Responders were defined as a normal
platelet count of >30×109/l, a doubling of baseline
platelet count and no bleeding symptoms. Non-responders were
defined as a platelet count of <30×109/l or below
twice as much as the pre-treatment platelet count.
Statistical analysis
All statistical analyses were performed using SPSS
for Microsoft Windows (version 13.0; SPSS, Inc., Chicago, IL, USA)
and GraphPad Prism 5 (GraphPad Software, Inc., La Jolla, CA, USA).
Quantitative data are presented as mean ± standard deviation. The
unpaired Student's t-test or one-way analysis of variance with the
Tukey multiple-comparison post hoc test, χ2 test and the
Pearson correlation coefficient were employed. P<0.05 was
considered to indicate a statistically significant difference.
Results
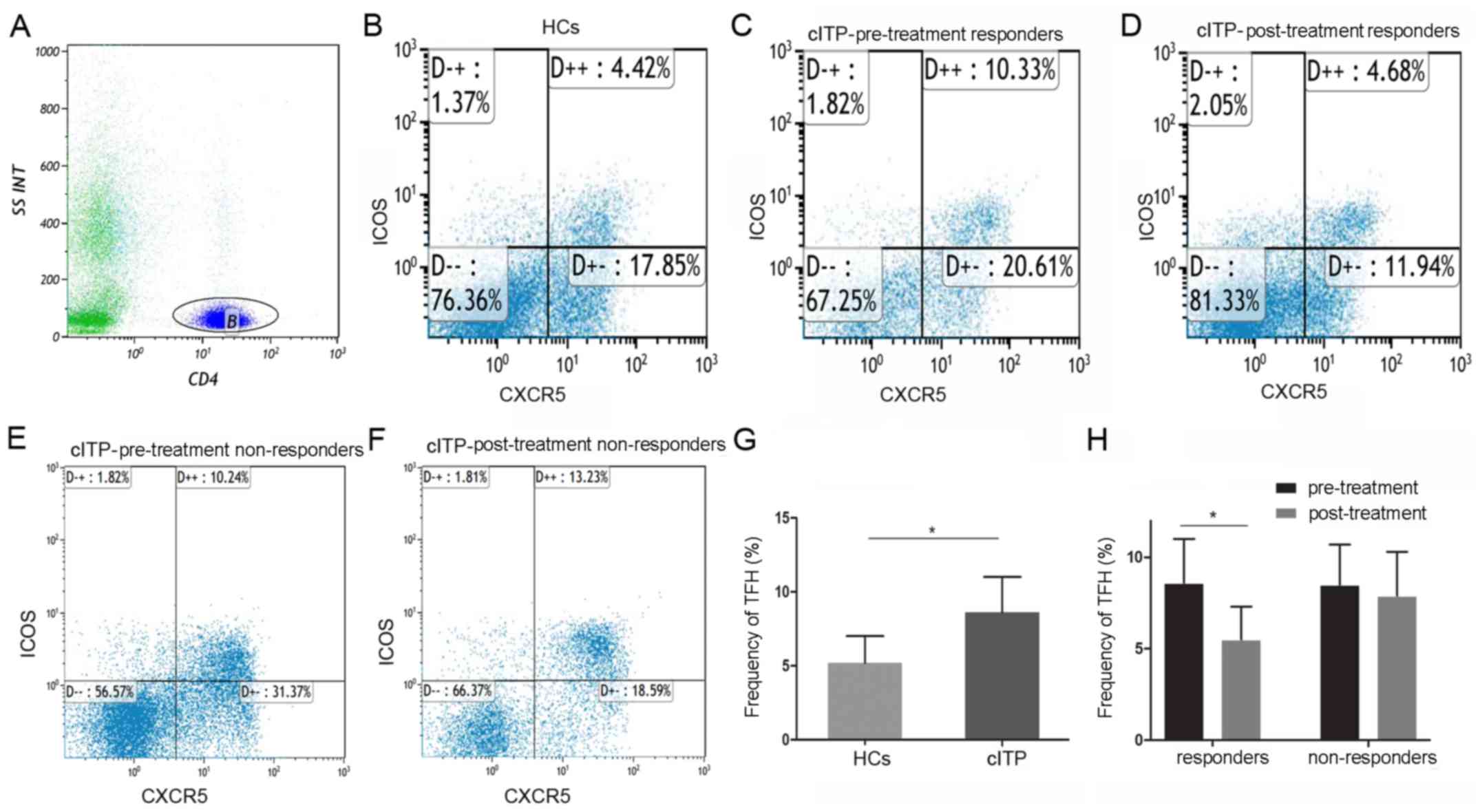
Circulating TFH cells in the
peripheral blood of patients with cITP and healthy controls
The baseline characteristics of patients with cITP
and healthy controls are indicated in Table I. There was no significant difference
in the distribution of age, sex and platelet count between the cITP
group and healthy controls. At least 10,000 events were collected
in a gate around the lymphocytes. In order to identify TFH cells,
CD4+ lymphocytes were gated on. Subsequently, the
percentages of CD4+ CXCR5+ICOS+
cells were gated and determined by flow cytometry.
 | Table I.Baseline characteristics of patients
with chronic immune thrombocytopenia and healthy controls. |
Table I.
Baseline characteristics of patients
with chronic immune thrombocytopenia and healthy controls.
| Variables | Healthy controls,
n=30 (%) | Responders, n=30
(%) | Non-responders,
n=24 (%) | P-value |
|---|
| Sex (n, %) |
|
Female | 16 (53.3) | 18 (60.0) | 14 (58.3) | NS |
|
Male | 14 (46.7) | 12 (40.0) | 10 (41.7) | NS |
| Age (years,
range) | 41
(24–60) | 45
(25–61) | 43
(28–60) | NS |
| Disease duration
(months, range) | – | 28
(13–124) | 27
(15–120) | NS |
| Platelet count
(109/l, mean ± SD) | 112.5±10.9 | 34.9±14.1 | 35.3±15.7 | 0.05 |
As indicated in Fig.
1, the proportion of circulating TFH cells in all pretreated
patients with cITP was significantly higher compared with healthy
controls (8.6±2.4 and 5.2±1.8%, P<0.05). Compared with
pre-treatment, the frequencies of circulating TFH cells in the
responders were significantly reduced post-treatment (8.8±2.5 and
5.5±1.8%, P<0.05), whereas no significant difference was
identified in the non-responders (8.5±2.2 and 7.9±2.4%,
P>0.05).
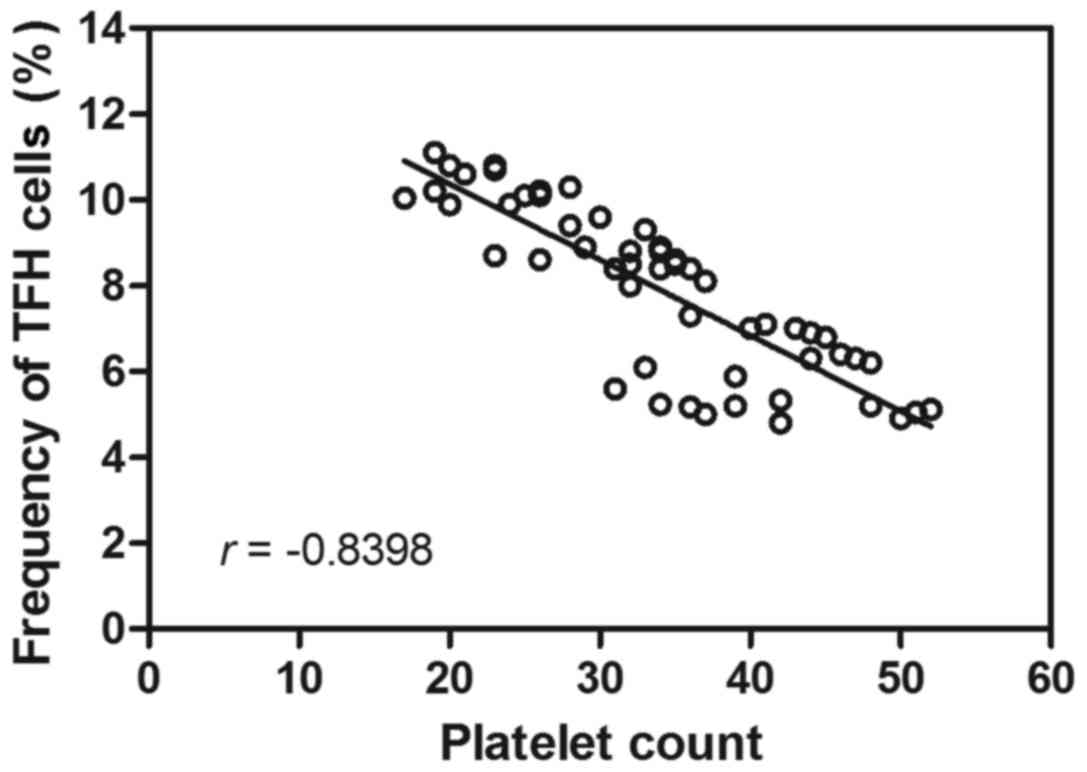
Correlation analysis between
circulating TFH cell proportions, PAIgG levels and platelet counts
in patients with cITP
Following the observational changes indicated with
regards to the circulating TFH cell ratio in patients with cITP and
respondents, the possible correlation between the proportion of
circulating TFH cells and platelet counts was assessed. Correlation
analysis revealed no correlation between the circulating TFH cell
ratio and PAIgG (r=0.21, P>0.05; data not shown). Furthermore,
the TFH cell percentage was negatively correlated with the platelet
count in peripheral blood (r=−0.84, P<0.05; Fig. 2). These results suggested that the
TFH cell ratio may be involved in decreasing the platelet count in
patients with cITP.
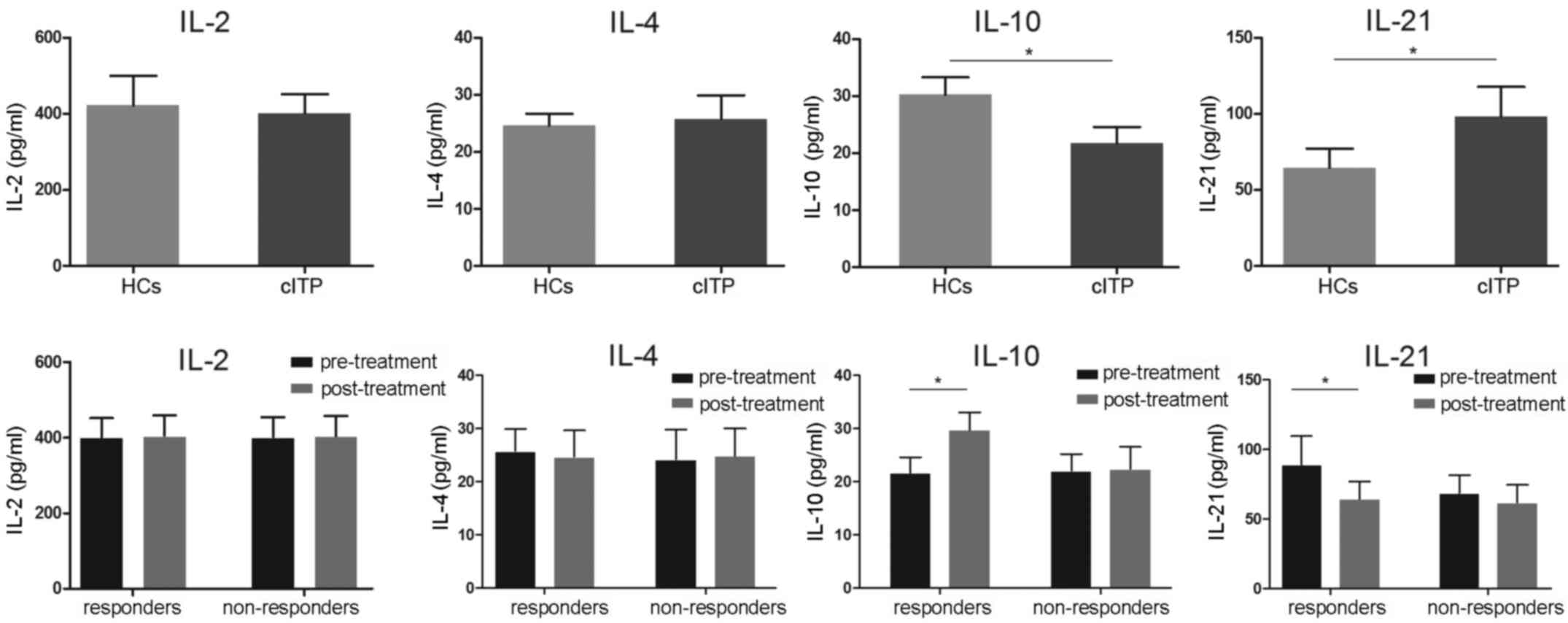
Serum IL-2, IL-4, IL-10 and IL-21
levels of cITP-associated cytokines
Following the results regarding the circulating TFH
cell ratio, the level of associated serum cytokines IL-2, IL-4,
IL-10 and IL-21 were evaluated using ELISA. As indicated in
Fig. 3, serum levels of IL-21 in
patients with cITP that were pretreated were significantly higher
compared with healthy controls, whereas the levels were
significantly lower in the pretreated responders compared with
responders post-treatment (P<0.05). Serum levels of IL-10 in
pretreated patients with cITP were significantly reduced compared
with the healthy controls, while the levels of IL-10 were
significantly elevated following treatment in the responders
(P<0.05). No significance differences in IL-10 and IL-21
expression levels were observed in non-responders (P>0.05).
Furthermore, no significant differences in IL-2 and IL-4 levels
were identified between pretreated patients and healthy controls or
responders and non-responders (P>0.05; Fig. 3).
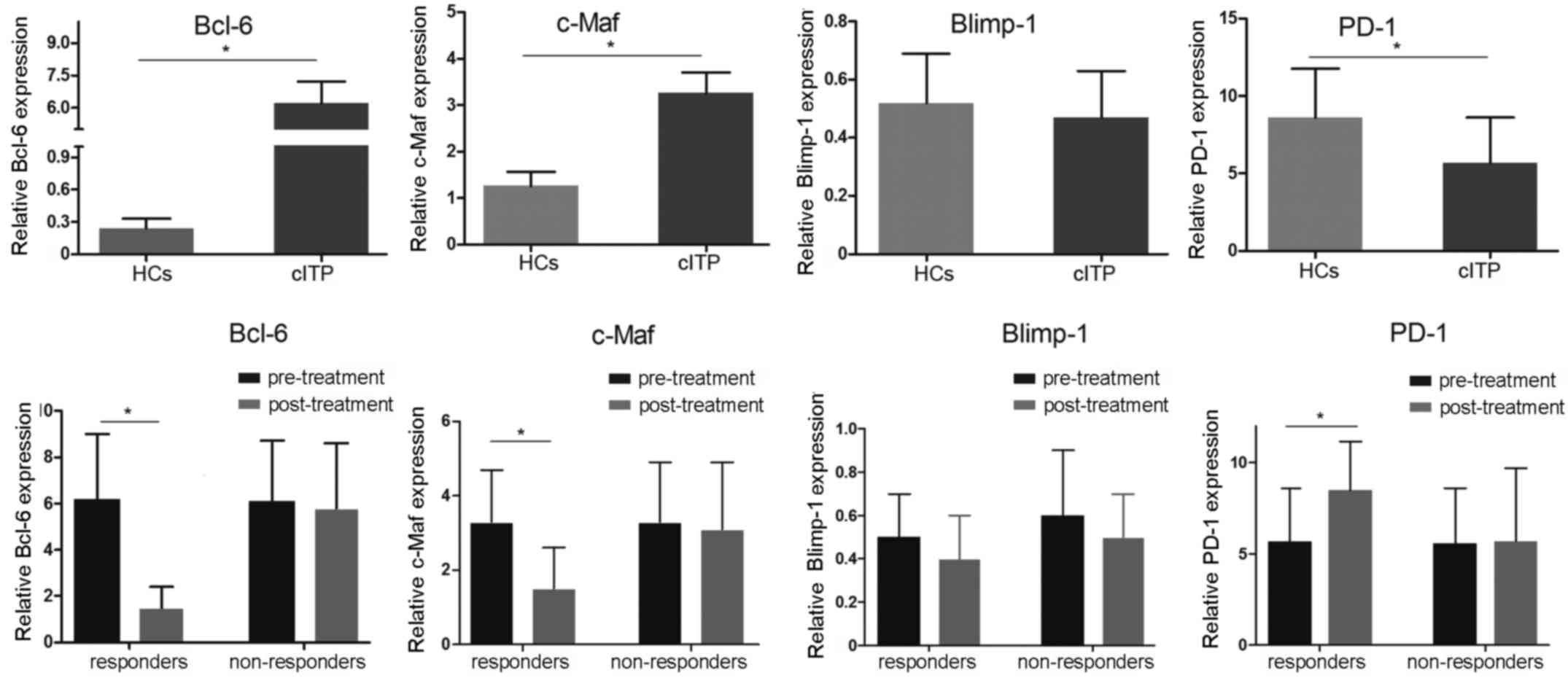
Transcription factor expression levels
in patients with cITP
To investigate the mRNA expression levels of the
possible involvement of Bcl-6, c-Maf, Blimp-1 and PD-1
transcription factors, RT-qPCR was applied. Compared with healthy
controls, the mRNA expression levels of Bcl-6 and c-Maf in
pretreated patients with cITP were significantly elevated, whereas
the expression levels of PD-1 were significantly decreased
(P<0.05; Fig. 4). There was no
significant difference in Blimp-1 mRNA expression between
pretreated patients with cITP and healthy controls (P>0.05).
Furthermore, the mRNA expression levels of Bcl-6 and c-Maf in cITP
responders post-treatment were significantly downregulated while
PD-1 in cITP responders post-treatment was significantly increased
compared with pretreated patients with cITP (P<0.05; Fig. 4), whereas no significant difference
was observed in expression level of Blimp-1 (P>0.05).
Discussion
Increasing evidence has revealed the association of
TFH cells and autoimmune diseases. In the present study, the
proportions of circulating TFH cells in patients with cITP were
significantly increased in the peripheral blood; however, this was
decreased to control levels following treatment. Additionally, a
negative correlation was indicated between the percentage of
circulating TFH cells and platelet count, demonstrating circulating
TFH cells may serve a role in the pathogenesis of adult cITP.
IL-2 is an essential factor in preventing autoimmune
disease development and inhibiting TFH differentiation (12). Previous data has demonstrated TFH
cells can express cytokines, such as TH2-associated cytokine IL-4
(13). However, changes in IL-4
levels were not observed in cITP and ITP responders in the present
study. In light of these findings, it was suggested that these two
cytokines may be involved in the early stage of antibody
generation, however, their role in patients with cITP may be
minor.
Notably, IL-10 is abundantly produced in healthy
persons (14). Xin et al
(15) identified that
IL-10-producing TFH cells have an increased capacity to form stable
TFH-B cell conjugates compared with their IL-10-TFH counterparts,
suggesting that IL-10+TFH cells may specialize in
providing distress signals to B cells during chronic infection.
Importantly, depletion of
IL-10+/IL-21+-coproducing CD4+ T
cells or deletion of IL-10, specifically from TFH cells, resulted
in impaired GC B cell responses, lymphocytic choriomeningitis
virus-specific antibody production and viral control (16). Subsequently, a heterogeneous
population of TFH cells was determined and a critical role for
TFH-derived IL-10 in promoting humoral immunity during persistent
viral infection was elucidated (17). In agreement with these findings, the
present study revealed a significant change of IL-10 in cITP and
ITP responders. However, further flow cytometry analysis of
IL-10+ TFH cells may be required to confirm its
role.
IL-21 is a type I cytokine that signals via a
specific receptor protein, IL-21 receptor (18,19), and
the common cytokine receptor γ-chain, γc, which is shared by the
receptors for IL-2, IL-4, IL-7, IL-9, IL-15 and IL-21 (20). In addition, it has been recognized
that IL-21 may be a master cytokine that promotes the expression of
Bcl-6 in CD4+ T cells (21–23). TFH
cells produce high levels of IL-21, a cytokine that is critical for
GC formation and also for the generation of TFH cells (24). In the present study, IL-21 was
significantly increased in cITP and significantly decreased in ITP
responders post-treatment. These results further confirmed the
possible regulatory role of IL-21 in ITP.
Bcl-6 is a selectively expressed transcription
factor in murine and human TFH cells (25,26).
Notably, Bcl-6 was previously demonstrated to be inhibitory in Th2
responses by blocking STAT6 from binding to DNA (27). Furthermore, a previous study revealed
that Bcl-6-deficient mice developed multi-organ inflammatory
diseases, exhibited enhanced IgE production and defective GC
reaction (28). Previous results
have suggested that Bcl-6 deficiency in T cells resulted in
impaired TFH cell development in vitro and in vivo
and that Bcl6 expression in B and T cells is required for GC
reactions (29–31). Notably, transcriptional repressor
Bcl-6 was considered as the critical gene involved with TFH cell
differentiation (29). It has been
suggested that overexpression of Bcl-6 promotes the mRNA expression
of several TFH cell-associated genes in the absence of exogenous
cytokines. Furthermore, Bcl-6 has been indicated to suppress the
expression of various microRNAs that are considered to control TFH
cell generation, including miR-17-92 (29). These findings suggest that Bcl-6
regulates TFH cell development through repression of microRNAs and
Th1, Th2 and Th17 lineage-specific transcription factors (32). The present study revealed
significantly decreased levels of Bcl-6 and IL-21 and increased
levels of IL-10 in responders, indicating the Th1 and Th2
modulating effect of the Bcl-6 gene.
c-Maf is a transcription factor in the AP-1 family
with a basic region/leucine zipper that is highly expressed by
mature TFH cells and is thought to primarily function as a
regulator of cytokines that can promote B cell proliferation and
differentiation (33–35). The present study revealed c-Maf was
significantly elevated in cITP and decreased in ITP responders
post-treatment, suggesting a possible association of TFH cells in
cITP and ITP responders. Furthermore, Sahoo et al (35) recently reported that c-Maf promotes
IL-4 secretion in TFH cells through direct binding to the CNS2
region in the IL-4 locus and via induction of interferon regulatory
factor 4, thus revealing a distinct role of c-Maf in IL-4 secretion
between Th2 and TFH cell subsets. However, significant changes in
IL-4 levels in cITP and ITP responders were not indicated in the
present study, suggesting an alternative influence of Th2 and TFH
cell subsets on IL-4 secretion.
PD-1, an immunoreceptor that belongs to the
CD28/CTLA-4 family, has been demonstrated as an important molecule
expressed on TFH cells (36).
Typically, PD-1 is a negative regulatory signaling molecule that
results in the inhibition of effector T cells via their specific
ligands (PD-L1 and PD-L2), which are expressed on target cells
(37). Furthermore, PD-1 signaling
contributes to induce memory B cell differentiation and promotes
the generation of high-affinity, long-lived plasma cells (38). Previous studies have indicated an
increased frequency of PD-1+ TFH cells in several
autoimmune renal diseases, including Henoch-Schönlein purpura
nephritis (39), IgA nephropathy
(40) and diabetic nephropathy
(41). A significant downregulation
of PD-1 expression levels was observed in patients with cITP in the
present study compared with healthy controls; however, PD-1 levels
were significantly increased in responders post-treatment compared
with pre-treatment. Taken together, these results indicate possible
involvement of TFH cells in ITP.
Bcl-6 and Blimp-1 present vital but opposing
influences in the development of TFH cells, mutual antagonism
between Bcl-6 and Blimp-1 is a primary mechanism for commitment to
the T effector and TFH cell fates (42). Although Blimp-1 was previously
indicated to be involved in Th cell differentiation (43), the present results indicate a
remarkable change in the Bcl-6 gene levels and did not demonstrate
a significant change of Blimp-1 levels at transcription level in
cITP patients.
In conclusion, the present study demonstrated that
the expansion of circulating TFH cells may serve a role in the
immunopathogenesis of cITP. These data aided to elucidate that
transcription factors such as Bcl-6, c-Maf and PD-1 and specific
cytokine signaling of IL-10 and IL-21 may correlate with the
abnormal activation of TFH cells in cITP. The present data revealed
a novel mechanism in patients with cITP and provided an
understanding of the role of TFH cells in cITP in offering
diagnostic values and potential novel therapeutic strategies.
Further research is required to fully clarify the clinical outcome
of respondent and non-respondent cITP patients with expanded TFH
cells.
Acknowledgements
Not applicable.
Funding
The present work is supported by the National
Natural Science Foundation of China (grant no. 81700132) and the
Natural Science Foundation of Jiangsu Province (grant no.
BK20170324).
Availability of data and materials
The data and materials used to support the findings
of this study are included within the published article.
Authors' contributions
LD and CR conceived and designed the study. LD, LH,
LC, ZW and MZ performed the experiments. LH reviewed and edited the
manuscript. XB and YH analyzed the data. CR gave final approval of
the version to be published. All authors read and approved the
manuscript.
Ethics approval and consent to
participate
Approval for the present study was obtained by the
Ethics Committee of The First Affiliated Hospital of Soochow
University.
Patient consent for publication
All patients admitted to the study provided informed
consent for their participation of the present study and
publication of the data.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Nomura S: Advances in diagnosis and
treatments for immune thrombocytopenia. Clin Med Insights Blood
Disord. 9:15–22. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Audia S, Mahévas M, Samson M, Godeau B and
Bonnotte B: Pathogenesis of immune thrombocytopenia. Autoimmun Rev.
16:620–632. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Lambert MP and Gernsheimer TB: Clinical
updates in adult immune thrombocytopenia. Blood. 129:2829–2835.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Cooper N: State of the art-how i manage
immune thrombocytopenia. Br J Haematol. 177:39–54. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Audia S, Rossato M, Trad M, Samson M,
Santegoets K, Gautheron A, Bekker C, Facy O, Cheynel N,
Ortega-Deballon P, et al: B cell depleting therapy regulates
splenic and circulating T follicular helper cells in immune
thrombocytopenia. J Autoimmun. 77:89–95. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Eivazi S, Bagheri S, Hashemzadeh MS,
Ghalavand M, Qamsari ES, Dorostkar R and Yasemi M: Development of T
follicular helper cells and their role in disease and immune
system. Biomed Pharmacother. 84:1668–1678. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Shekhar S and Yang X: The darker side of
follicular helper T cells: From autoimmunity to immunodeficiency.
Cell Mol Immunol. 9:380–385. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Yuan X, Cheng G and Malek TR: The
importance of regulatory T-cell heterogeneity in maintaining
self-tolerance. Immunol Rev. 259:103–114. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Chan H, Moore JC, Finch CN, Warkentin TE
and Kelton JG: The IgG subclasses of platelet-associated
autoantibodies directed against platelet glycoproteins IIb/IIIa in
patients with idiopathic thrombocytopenic purpura. Br J Haematol.
122:818–824. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Sanz MÁ, García Vicente V, Fernández A,
López MF, Grande C, Jarque I, Martínez R, Mingot ME, Monteagudo E,
Ribera JM, et al: Guidelines for diagnosis, treatment and
monitoring of primary immune thrombocytopenia. Med Clin (Barc).
138:261.e1–261.e17. 2012.(In Spanish).
|
|
11
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ballesteros-Tato A, León B, Graf BA,
Moquin A, Adams PS, Lund FE and Randall TD: Interleukin-2 inhibits
germinal center formation by limiting T follicular helper cell
differentiation. Immunity. 36:847–856. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Fairfax KC, Everts B, Amiel E, Smith AM,
Schramm G, Haas H, Randolph GJ, Taylor JJ and Pearce EJ:
IL-4-secreting secondary T follicular helper (Tfh) cells arise from
memory T cells, not persisting Tfh cells, through a B
cell-dependent mechanism. J Immunol. 194:2999–3010. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
den Hartog G, van Osch TLJ, Vos M, Meijer
B, Savelkoul HFJ, van Neerven RJJ and Brugman S: BAFF augments IgA2
and IL-10 production by TLR7/8 stimulated total peripheral blood B
cells. Eur J Immunol. 48:283–292. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Xin G, Schauder DM, Zander R and Cui W:
Two is better than one: Advances in pathogen-boosted immunotherapy
and adoptive T-cell therapy. Immunotherapy. 9:837–849. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Laidlaw BJ, Cui W, Amezquita RA, Gray SM,
Guan T, Lu Y, Kobayashi Y, Flavell RA, Kleinstein SH, Craft J and
Kaech SM: Production of IL-10 by CD4(+) regulatory T cells during
the resolution of infection promotes the maturation of memory
CD8(+) T cells. Nat Immunol. 16:871–879. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Chowdhury A, Del Rio Estrada PM, Tharp GK,
Trible RP, Amara RR, Chahroudi A, Reyes-Teran G, Bosinger SE and
Silvestri G: Decreased T follicular regulatory cell/T follicular
helper cell (TFH) in Simian Immunodeficiency virus-infected rhesus
macaques may contribute to accumulation of TFH in chronic
infection. J Immunol. 195:3237–3247. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Jeong J, Kim WH, Yoo J, Lee C, Kim S, Cho
JH, Jang HK, Kim DW, Lillehoj HS and Min W: Identification and
comparative expression analysis of interleukin 2/15 receptor β
chain in chickens infected with E. tenella. PLoS One. 7:e377042012.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Lewis KE, Selby MJ, Masters G, Valle J,
Dito G, Curtis WR, Garcia R, Mink KA, Waggie KS, Holdren MS, et al:
Interleukin-21 combined with PD-1 or CTLA-4 blockade enhances
antitumor immunity in mouse tumor models. Oncoimmunology.
7:e13778732017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Gharibi T, Majidi J, Kazemi T,
Dehghanzadeh R, Motallebnezhad M and Babaloo Z: Biological effects
of IL-21 on different immune cells and its role in autoimmune
diseases. Immunobiology. 221:357–367. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Bryant VL, Ma CS, Avery DT, Li Y, Good KL,
Corcoran LM, de Waal Malefyt R and Tangye SG: Cytokine-mediated
regulation of human B cell differentiation into Ig-secreting cells:
Predominant role of IL-21 produced by CXCR5+ T follicular helper
cells. J Immunol. 179:8180–8190. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Vyas AK and Trehanpati N: Commentary:
IL-21 receptor antagonist inhibits differentiation of B cells
toward plasmablasts upon alloantigen stimulation. Front Immunol.
8:9342017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Audia S, Rossato M, Santegoets K, Spijkers
S, Wichers C, Bekker C, Bloem A, Boon L, Flinsenberg T, Compeer E,
et al: Splenic TFH expansion participates in B-cell differentiation
and antiplatelet-antibody production during immune
thrombocytopenia. Blood. 124:2858–2866. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Spolski R and Leonard WJ: IL-21 and T
follicular helper cells. Int Immunol. 22:7–12. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Fazilleau N, McHeyzer-Williams LJ, Rosen H
and McHeyzer-Williams MG: The function of follicular helper T cells
is regulated by the strength of T cell antigen receptor binding.
Nat Immunol. 10:375–384. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
26
|
King C, Tangye SG and Mackay CR: T
follicular helper (TFH) cells in normal and dysregulated immune
responses. Annu Rev Immunol. 26:741–766. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Harris MB, Chang CC, Berton MT, Danial NN,
Zhang J, Kuehner D, Ye BH, Kvatyuk M, Pandolfi PP, Cattoretti G, et
al: Transcriptional repression of Stat6-dependent
interleukin-4-induced genes by BCL-6: Specific regulation of
iepsilon transcription and immunoglobulin E switching. Mol Cell
Biol. 19:7264–7275. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Ye BH, Cattoretti G, Shen Q, Zhang J, Hawe
N, de Waard R, Leung C, Nouri-Shirazi M, Orazi A, Chaganti RS, et
al: The BCL-6 proto-oncogene controls germinal-centre formation and
Th2-type inflammation. Nat Genet. 16:161–170. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Yu D, Rao S, Tsai LM, Lee SK, He Y,
Sutcliffe EL, Srivastava M, Linterman M, Zheng L, Simpson N, et al:
The transcriptional repressor Bcl-6 directs T follicular helper
cell lineage commitment. Immunity. 31:457–468. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Nurieva RI, Chung Y, Martinez GJ, Yang XO,
Tanaka S, Matskevitch TD, Wang YH and Dong C: Bcl6 mediates the
development of T follicular helper cells. Science. 325:1001–1005.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wei Y and Hou M: T cells in the
pathogenesis of immune thrombocytopenia. Semin Hematol. 53 Suppl
1:S13–S15. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Xin N, Fu L, Shao Z, Guo M, Zhang X, Zhang
Y, Dou C, Zheng S, Shen X, Yao Y, et al: RNA interference targeting
Bcl-6 ameliorates experimental autoimmune myasthenia gravis in
mice. Mol Cell Neurosci. 58:85–94. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Kroenke MA, Eto D, Locci M, Cho M,
Davidson T, Haddad EK and Crotty S: Bcl6 and Maf cooperate to
instruct human follicular helper CD4 T cell differentiation. J
Immunol. 188:3734–3744. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Hiramatsu Y, Suto A, Kashiwakuma D, Kanari
H, Kagami S, Ikeda K, Hirose K, Watanabe N, Grusby MJ, Iwamoto I
and Nakajima H: c-Maf activates the promoter and enhancer of the
IL-21 gene, and TGF-beta inhibits c-Maf-induced IL-21 production in
CD4+ T cells. J Leukoc Biol. 87:703–712. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Sahoo A, Alekseev A, Tanaka K, Obertas L,
Lerman B, Haymaker C, Clise-Dwyer K, McMurray JS and Nurieva R:
Batf is important for il-4 expression in T follicular helper cells.
Nat Commun. 6:79972015. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Berger KN and Pu JJ: PD-1 pathway and its
clinical application: A 20 year journey after discovery of the
complete human PD-1 gene. Gene. 638:20–25. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Guo Y, Walsh AM, Canavan M, Wechalekar MD,
Cole S, Yin X, Scott B, Loza M, Orr C, McGarry T, et al: Immune
checkpoint inhibitor PD-1 pathway is down-regulated in synovium at
various stages of rheumatoid arthritis disease progression. PLoS
One. 13:e01927042018. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Greisen SR, Rasmussen TK,
Stengaard-Pedersen K, Hetland ML, Hørslev-Petersen K, Hvid M and
Deleuran B: Increased soluble programmed death-1 (sPD-1) is
associated with disease activity and radiographic progression in
early rheumatoid arthritis. Scand J Rheumatol. 43:101–108. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Zhang Z, Zhao S, Zhang L, Crew R, Zhang N,
Sun X and Jiang Y: A higher frequency of CD4(+)CXCR5(+) T
follicular helper cells in patients with newly diagnosed
Henoch-Schönlein purpura nephritis. Int Immunopharmacol. 32:8–15.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Zhang L, Wang Y, Shi X, Zou H and Jiang Y:
A higher frequency of CD4+CXCR5+ T follicular helper cells in
patients with newly diagnosed IgA nephropathy. Immunol Lett.
158:101–108. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Zhang N, Tai J, Qu Z, Zhang Z, Zhao S, He
J, Zhang S and Jiang Y: Increased CD4+CXCR5+ T follicular helper
cells in diabetic nephropathy. Autoimmunity. 49:405–413. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Krishnamoorthy V, Kannanganat S,
Maienschein-Cline M, Cook SL, Chen J, Bahroos N, Sievert E, Corse
E, Chong A and Sciammas R: The IRF4 gene regulatory module
functions as a read-write integrator to dynamically coordinate T
helper cell fate. Immunity. 47(481–497): e72017.
|
|
43
|
Fu SH, Yeh LT, Chu CC, Yen BL and Sytwu
HK: New insights into Blimp-1 in T lymphocytes: A divergent
regulator of cell destiny and effector function. J Biomed Sci.
24:492017. View Article : Google Scholar : PubMed/NCBI
|