Introduction
Angiogenesis is an important factor that influences
the development of atherosclerotic plaques and is closely
associated with local neovascularization and plaque stability
(1). It has been demonstrated that
excessive angiogenesis is associated with intraplaque hemorrhage,
suggesting that neovascularization may contribute to lesion
progression and rupture (2).
Rupture-prone vulnerable plaques exhibit high lipid infiltration,
increased numbers of inflammatory cells, a thin fibrous cap, and
extensive endometrial and adventitial neovascularization (3–5). Studies
have generally focused on the intimal accumulation of lipids and
inflammatory cells (6,7); however, the results of a recent study
suggest that endothelial cells also serve a critical role during
advanced plaque angiogenesis (8).
LncRNAs are transcripts that are >200 nucleotides
long, which regulate transcription via chromatin modulation,
post-transcriptional regulation, organization of protein complexes,
cell signaling and allosteric regulation (9,10). They
serve important roles in physiological processes, including the
differentiation, proliferation, apoptosis and invasion of induced
pluripotent stem cells (11,12). Studies have demonstrated that several
lncRNAs, including lncRNA-p21 (13),
metastasis associated lung adenocarcinoma transcript 1 (14) and lnc-Ang362 (15), are associated with the progression of
atherosclerosis and relevant cellular processes. Furthermore, it
has been confirmed that lncRNA is able to facilitate endothelial
angiogenic function (16).
MicroRNAs (miRNAs) are endogenous small
single-strand non-coding RNAs ~22 nucleotides long (17). Certain miRNAs are able to regulate
angiogenic processes and negatively modulate angiogenesis (18). For example, miRs-221 and −222 are
able to regulate the development and differentiation of endothelial
cells (ECs) but suppress pro-angiogenic activation and migration
(19). Furthermore, the EC-derived
miRNA 17–92 cluster negatively regulates arteriogenesis via
miRNA-19 repression of Wnt signaling (20). miR-21 is highly expressed in ECs and
studies have demonstrated that miR-21 expression suppresses the
expression of phosphatase and tensing homolog (PTEN) but
upregulates vascular endothelial growth factor (VEGF) expression
(21). Furthermore, miR-21 has been
identified as a modulator of migration and tubulogenesis in HUVECs
(22).
The present study measured the expression of lncRNAs
in tumor necrosis factor (TNF)-α-induced HUVECs and demonstrated
that TCONS_00024652 was overexpressed. Functional experiments
indicated that TCONS_00024652 silencing leads to the inhibition of
angiogenesis and the expression of downstream miR-21. It was
therefore suggested that TCONS_00024652 silencing induces the
subsequent upregulation of miR-21.
Materials and methods
Cell culture
Human umbilical vein endothelial cells (HUVECs) were
obtained from the American Type Culture Collection (cat. no.
CRL-1730, Manassas, VA, America) and cultured in Dulbecco's
modified Eagle's medium (DMEM; Gibco; Thermo Fisher Scientific,
Inc., Waltham, MA, USA) with 10% (v/v) fetal bovine serum (FBS;
Hyclone; GE Healthcare, Logan, UT, USA) in a humidified incubator
at 37°C with 5% CO2. HUVECs were seeded at
3×105 cells/well in 6-well plates and treated with
varying concentrations (0.5, 1 or 2 ng/ml for 24 h) or for varying
times (1 ng/ml for 6, 12 or 24 h) with recombinant human TNF-α
(Peprotech, Rock Hill, NJ, USA). Following treatment, cells were
lysed and RNA or protein was extracted for further experiments.
Cell transfection
All small interfering (si)RNAs used in the current
study were synthesized by Shanghai GenePharma Co., Ltd. (Shanghai,
China), including si-lncRNA, mimic siRNAs, inhibitor siRNAs and
scramble negative control (NC) siRNAs. Cells at 70% confluence were
transfected with 100 nM siRNAs using Lipofectamine 2000
(Invitrogen; Thermo Fisher Scientific, Inc.), following the
manufacturer's protocol. The si-TCONS_00024652-1 sequences were:
5′-GUGAAUUCAACAGACUGAAUU-3′ and the si-TCONS_00024652-2 sequence
was 5′-GUGAUUUACCAGUCAGUGAAU-3′. The sequences of miR-21 mimics,
inhibitor and negative control were as follow: miR-21 mimics
forward, 5′-UGGCAGUCUCUUAGAUGGUGG-3′ and reverse,
5′-GCCAUGCAAGACACUGCCAGG-3′; miR-21 inhibitor forward,
5′-UUCUAGCCUACACCGUCCCGAA-3′ and reverse,
5′-UGGUAGUCGACUCCGCACCTGA-3′; negative control forward:
5′-UUCUCGGAACGUGUCACGUTT-3′ and reverse:
5′-UUCAAGGCCCGUGUCAUGUGG-3′. Subsequent experiments were performed
48 h following transfection.
Cell proliferation assay
EC proliferation was measured using a Cell Counting
Kit-8 (CCK-8; Dojindo Molecular Technologies, Kuamoto, Japan).
Following transfection, cells were seeded in 96-well plates at
density of 5×103/well. CCK-8 solution was added and
cells were incubated at 37°C for 90 min. Subsequently, absorbance
was measured using an Infinite 200 Pro microplate reader (Tecan
Group Ltd., Mannedorf, Switzerland) at 450 nm.
RNA extraction and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
Total RNA was extracted from HUVECs using total RNA
extraction reagent (Shanghai Genepharma, Co. Ltd.) and cDNA was
synthesized from the extracted RNA using a FastQuant RT kit
[Tiangen Biotech (Beijing) Co., Ltd., Beijing, China]. qPCR was
then performed using SuperReal PreMix Plus [Tiangen Biotech
(Beijing), Co., Ltd., Beijing, China]. GAPDH was used as an
internal control. All primers were synthesized by Sangon Biotech
Co., Ltd. (Shanghai, China). The sequences of the primers used were
as follows: miR-21 stem-loop RT primer,
5′-CTCAACTGGTGTCGTGGAGTCGGCAATTCAGTTGAGTCAACATC-3′; miR-21 forward,
5′-ACACTCCAGCTGGCTAGCTTATCAGACTGATG-3′ and reverse,
5′-CTCAACTGGTGTCGTGGA-3′; GAPDH forward,
5′-GTCAACGGATTTGGTCTGTATT-3′ and 5′-AGTCTTCTGGGTGGCAGTGAT-3′.
Primer sequences for lncRNA are listed in Table I. Thermal cycling conditions were as
follows: 94°C for 15 min; followed by 40 cycles of 94°C for 15 sec,
60°C for 32 sec, 72°C for 60 sec. Relative expression levels of
gene were quantified based target gene/GAPDH and calculated using
the 2−ΔΔCq method (23).
 | Table I.lcnRNA primers used in reverse
transcription-quantitative polymerase chain reaction analysis. |
Table I.
lcnRNA primers used in reverse
transcription-quantitative polymerase chain reaction analysis.
| Name | Sequence
(5′-3′) |
|---|
| ATG9B forward |
GGTCTGCCATCTATTACTT |
| ATG9B reverse |
TCTACGCATTATAGTCACAA |
| LncRNA-p21
forward |
ATAGTAGTTGGAGACTTCA |
| LncRNA-p21
reverse |
ACTGTATATTCAATGTTGGC |
| TCONS_00004897
forward |
GGATGGCACTCCTAGTCTGC |
| TCONS_00004897
reverse |
GTGCAGGCCTGAGTCTTTA |
| TCONS_00024652
forward |
ACGCTAACTGGCACCTTGTT |
| TCONS_00024652
reverse |
TGGGGATTACTGGGGTAGAT |
| LNC00305
forward |
TAAAGGATGCGTAGGGATGG |
| LNC00305
reverse |
TTCATGATCACGCCCTCATA |
| SENCR forward |
TTTACTTTAACAGACCAGAA |
| SENCR reverse |
CTCCTTTGTTGAATCCAT |
| MALAT1 forward |
TTATCCTTGGAAGAGTATT |
| MALAT1 reverse |
TAAGAAGTCACATTATTGG |
| ANRIL forward |
TTGATGAGAAGAATAAGCC |
| ANRIL reverse |
CTCCTTTGATGTGTGTTT |
| Tie-1AS
forward |
CCGGGGGTATACTACGGTC |
| Tie-1AS
reverse |
CTCTAGAGGGGGTAGAGG |
| TCONS_00004013
forward |
GCGCTAACTGGCACCTTGAT |
| TCONS_00004013
reverse |
TTTGGATTACTGGGGTATTG |
| U6 forward |
GCTTCGGCAGCACATATACTAAAAT |
| U6 reverse |
CGCTTCACGAATTTGCGTGTCAT |
Bioinformatics analysis
RNA-RNA and protein-RNA interaction networks were
analyzed using StarBase v2.0 (http://starbase.sysu.edu.cn/). The database contains
unique features, including determination of ceRNA functional
networks based on miRNA-target interactions, provides comprehensive
miRNA-lncRNA interactions and drafts interaction maps between
miRNAs and circRNAs (24).
Luciferase assay
To examine interactions between TCONS_00024652 and
miR-21, wild-type pmirGLO-TCONS_00024652 and diverse mutant
pmirGLO-TCONS_00024652 with pmirGLO-promotor vector (GenePharma,
Shanghai, China) were constructed. miR-21 mimics, mimic NC, miR-21
inhibitor and inhibitor NC were purchased from GenePharma Co., Ltd.
(Shanghai, China), 293T cells were plated at a density of
8×103 cells/well in 96-well plates and cultured at 37°C
with 5% CO2 overnight. Cells were then co-transfected
with the wild-type or mutant luciferase reporter plasmids, and
miR-21 mimic or NC using Lipofectamine 2000. Following 48 h,
luciferase activity was measured using the Bright-Glo™
Luciferase assay system (Promega Corporation, Madison, WI, USA) and
normalized to that of Renilla luciferase.
Wound healing assay
HUVECs at 80% confluence were plated at a density of
3×105 cells/well in 6-well plates and cultivated in DMEM
without FBS at 37°C overnight. The monolayer of HUVECs was
scratched with a 1 ml of a 200 µl pipette tip to form wound gaps.
Cells were washed by PBS and incubated with 1 ng/ml TNF-α at 37°C
to induce cell migration for 24 h. Images were captured using light
microscope (magnification, ×200). Cells were photographed prior to
and 24 h following scratching and the migration distance were
calculated by Image J 1.48 software (National Institutes of Health,
Bethesda, MD, USA).
Tube formation assay
A tube formation assay was performed to detect the
angiogenesis ability of HUVECs. Briefly, cells were harvested
following treatment and seeded into a 96-well plate
(1×104 cells/well) coated with 10 mg/ml Matrigel (BD
Biosciences, Franklin Lakes, NJ, USA). Following 7 h incubation at
37°C, capillary tube structures were observed and representative
images were captured using a light microscope (magnification,
×400). The branching length was determined using Image-Pro plus 6.0
(Media Cybernetics, Rockville, MD, USA).
Statistical analysis
All statistical analyses were performed using
GraphPad Prism 6.0 software (GraphPad, Software Inc., La Jolla, CA,
USA) and SPSS version 19.0 (IBM Corp., Armonk, NY, USA). Unpaired
Student's t test or one-way analysis of variance followed by least
significant difference tests were performed to determine
differences among groups. Heterogeneous data was analyzed using
one-way analysis of variance followed by Dunnett's tests. All data
are presented as the mean ± standard deviation and P<0.05 was
considered to indicate statistically significant difference.
Results
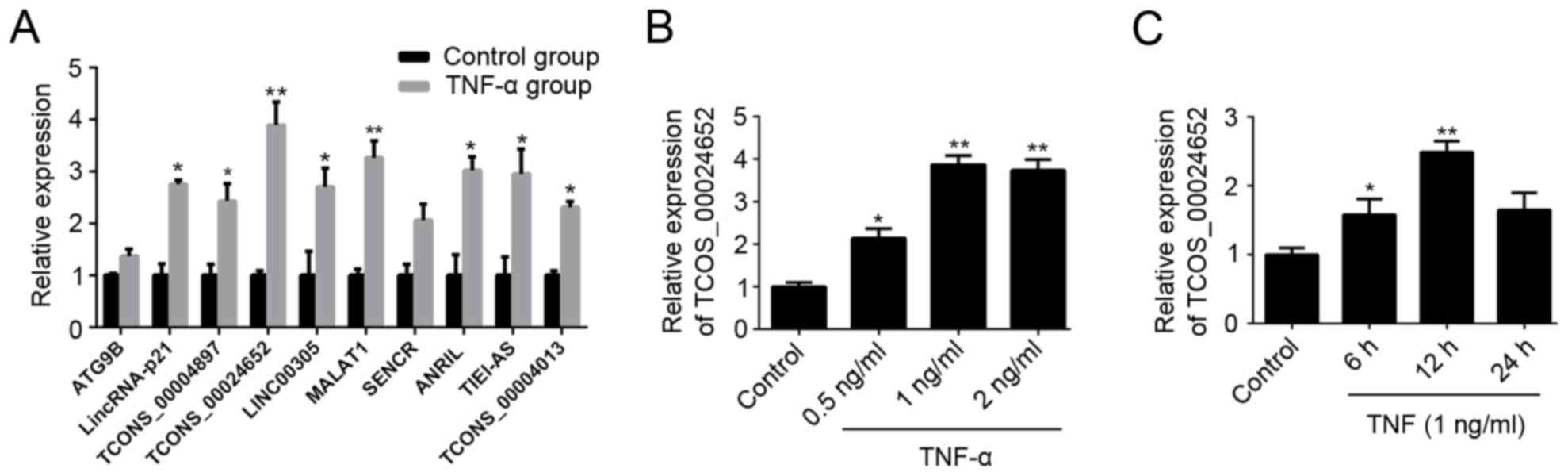
LncRNA expression profiles indicate
that TCONS_00024652 is upregulated following TNF-α treatment in
HUVECs
During the initial phase of the current study,
HUVECs were stimulated with TNF-α and levels of lncRNAs associated
with the regulation of vascular function (25,26) and
atherosclerosis (27) were assessed
using RT-qPCR. The results revealed that expression of a majority
of lncRNAs was significantly increased in these HUVECs following
TNF-α treatment; TCONS_00024652 exhibited the largest changes
(Fig. 1A). Therefore, it was
selected as the target lncRNA in subsequent experiments. RT-qPCR
was performed to further validate the up-regulation of
TCONS_00024652 following stimulation with TNF-α (Fig. 1B and C). These results indicate that
TNF-α treatment up-regulates the expression of TCONS_00024652 in a
dose-dependent manner.
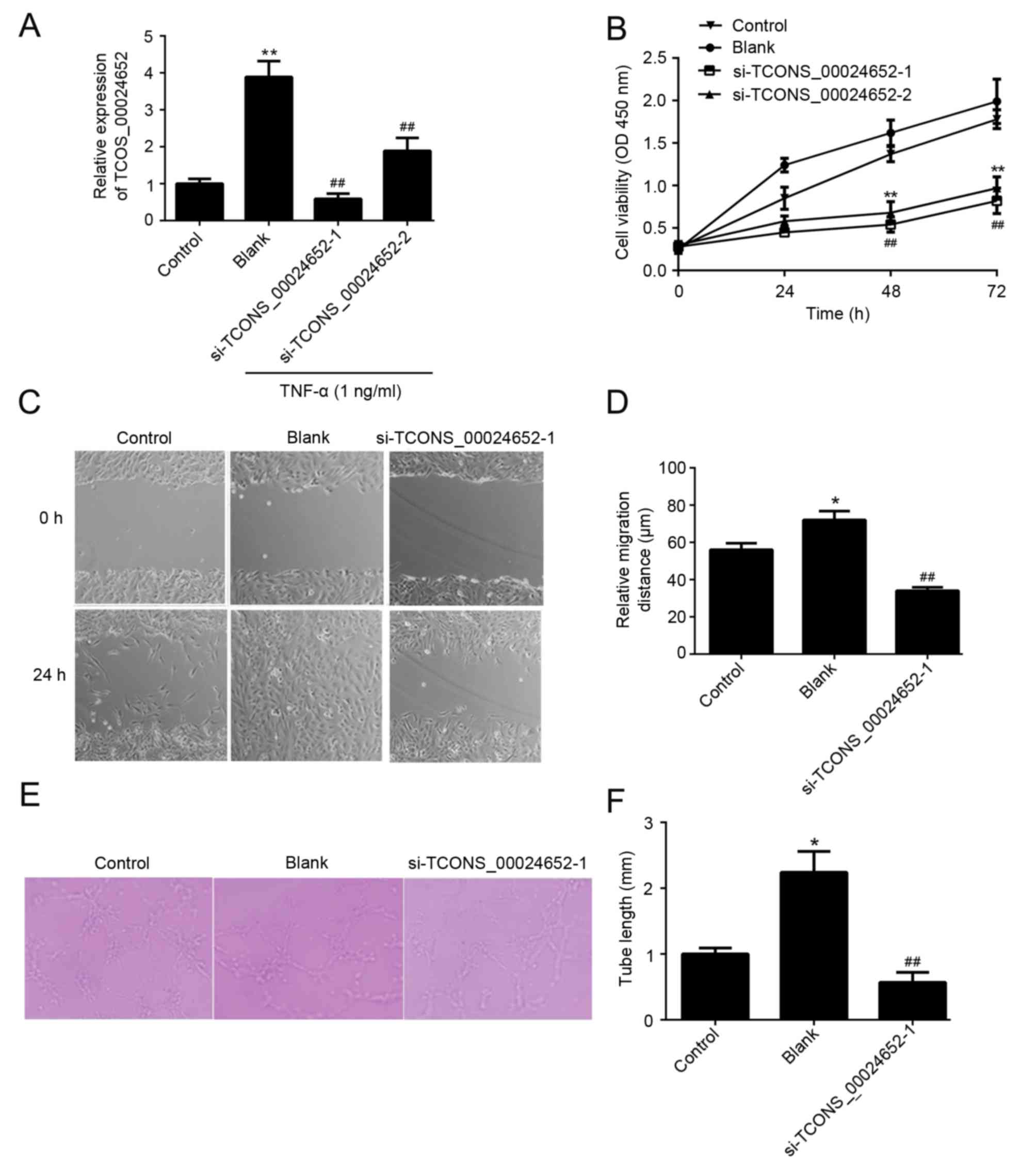
TCONS_00024652 knockdown inhibits the
proliferation, migration and angiogenesis of HUVECs
The function of TCONS_00024652 in cell proliferation
and angiogenesis was investigated. To ensure the effectiveness of
siRNAs in gene inhibition, two TCONS_00024652 siRNAs
(si-TCONS_00024652-1 and si-TCONS_00024652-2) were designed and
transfected into HUVECs to determine the effect of TCONS_00024652
on atherosclerosis progression. Transfection efficiency was
determined using RT-qPCR and the results indicated that the
transfection efficiency of si-TCONS_00024652-1 was higher than that
of si-TCONS_0024652-2 (Fig. 2A).
Furthermore, the results of the CCK-8 assay indicated that
si-TCONS_00024652-1 was more effective at reducing the
proliferation of HUVECs compared with si-TCONS_00024652-2 (Fig. 2B). Therefore, si-TCONS_00024652-1
alone was used in subsequent experiments. The results of the wound
healing assay indicated that TCONS_00024652 silencing significantly
decreased the migration of cells compared with the blank control
(Fig. 2C and D). Furthermore,
TCONS_00024652 knockdown significantly reduced the ability of
endothelial cells to form lumens (Fig.
2E and F). Taken together, these results indicate that
TCONS_00024652 promotes the proliferation and angiogenesis of
endothelial cells.
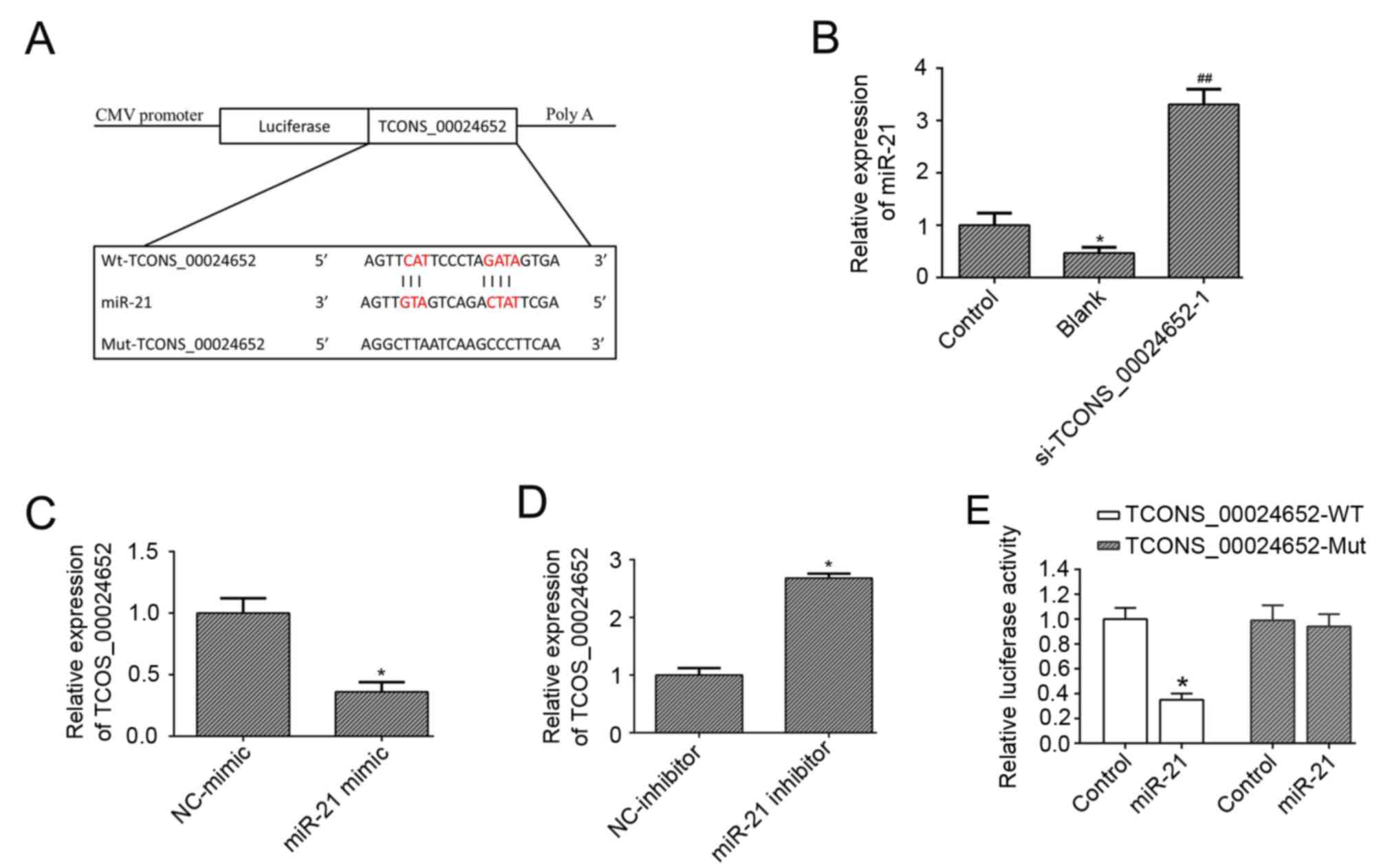
TCONS_00024652 directly targets
miR-21
Previous studies have demonstrated that miR-21
possesses anti-proliferative (28)
and anti-angiogenesis (24,29) properties; therefore it was
hypothesized that miR-21 may be a downstream target in
TNF-α-induced HUVEC activation. Bioinformatics analysis indicated
that miR-21 was a potential target of TCONS_00024652 (Fig. 3A). To explore the effect of
TCONS_00024652 on miR-21, its expression was measured in HUVECs
following TCONS_00024652 knockdown. The results demonstrated that
miR-21 expression was significantly increased compared with the
blank control (Fig. 3B).
Furthermore, the expression of TCONS_00024652 was significantly
decreased in HUVECs transfected with the miR-21 mimic (Fig. 3C) and increased in cells transfected
with the miR-21 inhibitor (Fig. 3D).
The results of the luciferase reporter assay confirmed that miR-21
interacts with TCONS_00024652, indicating that luciferase activity
was significantly decreased following transfection of miR-21 into
TCONS_0024652-WT (Fig. 3E). Overall,
these data indicate that TCONS_00024652 downregulates miR-21
expression.
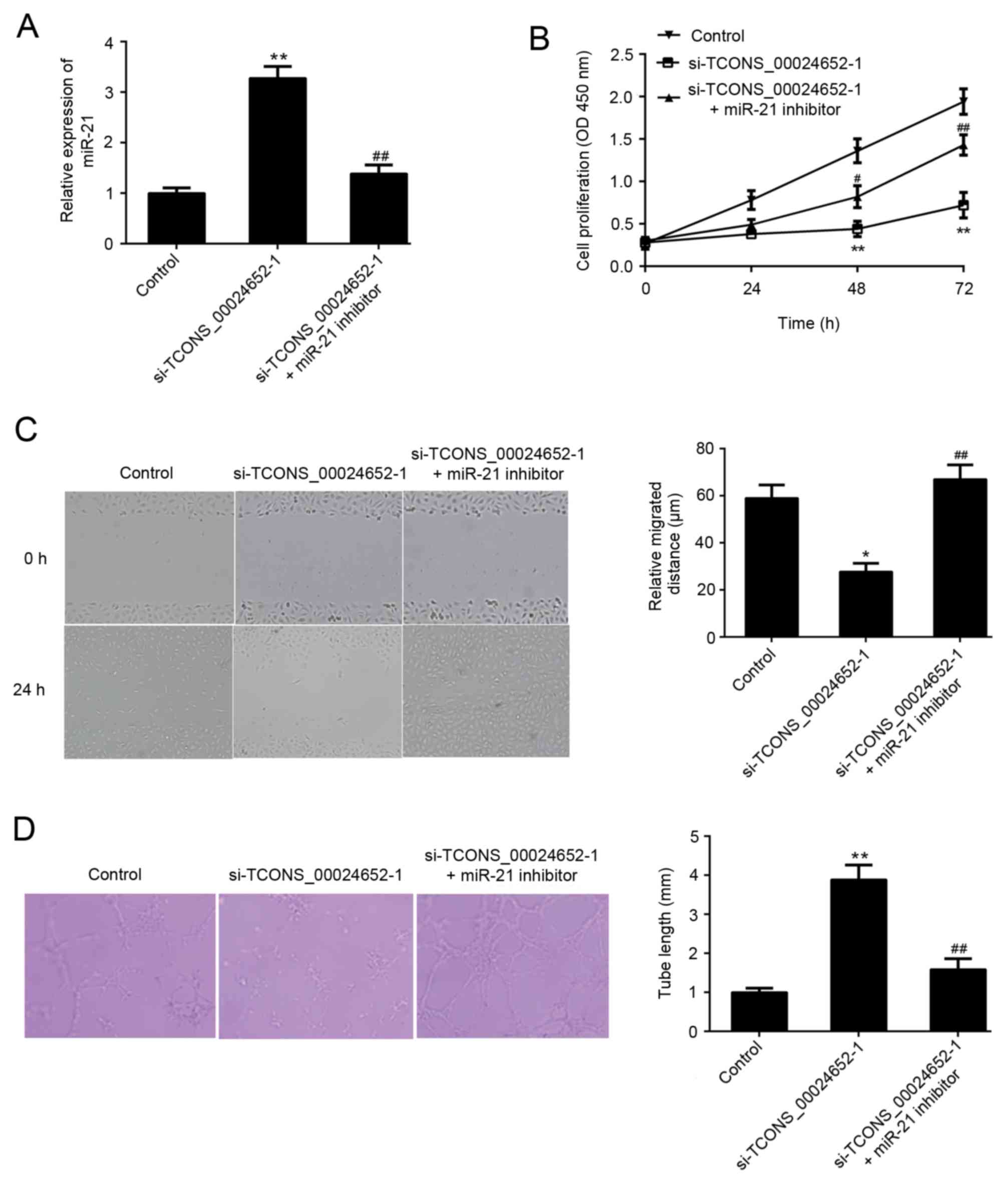
miR-21 inhibition reverses the
inhibitory effect of TCONS_00024652 knockdown on proliferation,
migration and angiogenesis in HUVECs
The aforementioned results demonstrated that there
is an association between TCONS_00024652 and miR-21. Thus, it was
hypothesized that miR-21 may neutralize the effects of
TCONS_00024652 on HUVECs. A series of experiments were performed to
confirm that TCONS_00024652 affects cell proliferation and
angiogenesis by regulating miR-21 expression in endothelial cells.
In cells stably transfected with si-TCONS_00024652, the expression
of miR-21 was significantly higher than in the control group; this
increase was reversed following transfection with miR-21 inhibitor
(Fig. 4A). The co-transfection of
the miR-21 inhibitor significantly reversed the inhibitory effects
of si-TCONS_00024652 on HUVEC proliferation, migration and invasion
(Fig. 4B-D). These results
demonstrate that TCONS_00024652 affects endothelial cell
proliferation and angiogenesis via the regulation of miR-21
expression, suggesting that TCONS_00024652 acts as a competing
endogenous RNA (ceRNA) for miR-21.
Discussion
Studies investigating the effects of inflammation on
atherosclerotic plaques have tended to focus on their innate immune
and adaptive immune responses. However, it has been demonstrated
that inflammation is closely associated with neovascularization in
plaques and that this may be another mechanism which increases the
vulnerability of plaques (30).
Chemokine and inflammatory factors secreted by plaque inflammatory
cells are able to directly or indirectly promote angiogenesis; this
process is referred to as ‘inflammatory angiogenesis’ (31). The biological effect of TNF-α depends
on its concentration and the type of cell it is acting on. High
concentrations of TNF-α activate tumor necrosis factor receptor
(TNFR) 1 to induce inflammation and stimulate the apoptosis of
endothelial cells; however low concentrations of TNF-α promote
endothelial cell proliferation, migration and microvascular
production via TNFR2 (32). The
present study successfully used low concentrations of TNF-α to
induce endothelial angiogenesis and also confirmed the important
role that inflammation serves in angiogenesis.
Previous studies have demonstrated the critical role
that lncRNAs serve during the progression of various diseases
(33,34), including cardiovascular disease
(35). It has been demonstrated that
smooth muscle and endothelial cell enriched
migration/differentiation-associated lncRNA regulates endothelial
cell differentiation and the angiogenic capacity of HUVEC (36). Furthermore, a conserved natural
noncoding antisense RNA in the tie-1 locus lncRNA-myocardial
infarction associated transcript serve important roles in the
pathological mechanism of atherosclerosis and may function as
molecular sponges in VECs (37,38). In
the present study, the gene expression profile of the lncRNA
TCONS_00024652 was screened and its functional importance in
TNF-α-induced HUVECs was determined. TCONS_00024652 silencing
reduced the proliferation and angiogenesis of TNF-α-induced HUVECs,
suggesting that TCONS_00024652 serves an angiogenesis-promoting
role in TNF-α-stimulated HUVECs.
In addition to regulating transcription, it has been
demonstrated that many lncRNAs function as endogenous decoys for
miRNAs. For example, LOC100129973, an important regulator of
endothelial cell apoptosis, contains sites that bind to miR-4707-5p
and miR-4767 (39). XLOC_008466
functions as an oncogene in non-small cell lung cancer by
regulating the miR-874-matrix metalloproteinase/X-linked inhibitor
of apoptosis protein axis (40). In
the present study, bioinformatics was used to search for target
miRNAs to reveal the downstream regulatory pathway by which
TCONS_00024652 acts on HUVECs. The results indicated that miR-21
directly targets TCONS_00024652. The rescue experiments and
luciferase reporter assay confirmed that TCONS_00024652 and miR-21
had a pairing region. Furthermore, it was demonstrated that
TCONS_00024652 targets miR-21, decreasing its expression, thus
acting as a ceRNA or molecular sponge. This indicates that
TCONS_00024652 induces the proliferation and angiogenesis of VECs
and stimulates the formation of atherosclerotic plaques. Although
the results of the present study clearly indicate that there is an
interaction between miR-21 and TCONS_00024652 that results in the
regulation of HUVEC proliferation and angiogenesis, their effects
on downstream target proteins remains uncertain. The instability of
atherosclerotic plaques is associated with immature angiogenesis
and the regulation of the interaction between VEGF and other
factors may result in the recruitment of pericytes and promotion of
neovascularization by the basement membrane (41). Future studies are required to
elucidate the mechanisms of action of TCONS_00024652 and miR-21 on
atherosclerotic plaques.
In conclusion, the results of the current study
indicate that the increased expression of the
endothelium-associated lncRNA TCONS_00024652 promotes plaque
angiogenesis and the progression of atherosclerosis via the
TCONS_00024652/miR-21 signaling pathway, suggesting that
TCONS_00024652 may be an important molecular marker that stabilizes
atherosclerotic plaques and slows down the progression of
atherosclerosis.
Acknowledgements
The authors wish to thank Dr. Liu from the Basic
Medical Research Center of Xinjiang Medical University for his
technical support throughout the study.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
MH performed the experiments and prepared the
manuscript. AA designed and organized the project and helped with
the preparation of the manuscript. BD analyzed the data and
performed statistics. JN collected and interpreted the data. All
authors read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
lncRNA
|
long noncoding RNA
|
|
VECs
|
vascular endothelial cells
|
|
TNF-α
|
tumor necrosis factor α
|
|
HUVECs
|
human umbilical vein endothelial
cells
|
|
PTEN
|
phosphatase and tensin homolog
|
|
TNFR1
|
tumor necrosis factor receptor 1
|
References
|
1
|
Camaré C, Pucelle M, Nègre-Salvayre A and
Salvayre R: Angiogenesis in the atherosclerotic plaque. Redox Biol.
12:18–34. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Weng J: Activation of CD137 signaling
promotes angiogenesis in atherosclerosis via modulating endothelial
Smad1/5-NFATc1 pathway. J Am Heart Assoc. 6:234–247. 2017.
View Article : Google Scholar
|
|
3
|
Sun Z: Atherosclerosis and Atheroma Plaque
Rupture: Imaging Modalities in the Visualization of Vasa Vasorum
and Atherosclerotic Plaques. Sci World J. 23:1–12. 2014. View Article : Google Scholar
|
|
4
|
Mulligan-Kehoe MJ: The vasa vasorum in
diseased and nondiseased arteries. Am J Physiol Heart Circ Physiol.
298:H295–H305. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Moguillansky D, Leng X, Carson A, Lavery
L, Schwartz A, Chen X and Villanueva FS: Quantification of plaque
neovascularization using contrast ultrasound: A histologic
validation. Eur Heart J. 32:646–653. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wu MY and Li CJ: New insights into the
role of inflammation in the pathogenesis of atherosclerosis. Int J
Mol Sci. 18:E20342017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Pirillo A, Bonacina F, Norata GD and
Catapano AL: The interplay of lipids, lipoproteins and immunity in
atherosclerosis. Curr Atheroscler Rep. 20:122018. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Sedding DG, Boyle EC, Demandt JAF, Sluimer
JC, Dutzmann J, Haverich A and Bauersachs J: Vasa vasorum
angiogenesis: Key player in the initiation and progression of
atherosclerosis and potential target for the treatment of
cardiovascular disease. Front Immunol. 9:7062018. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Tsai MC, Manor O, Wan Y, Mosammaparast N,
Wang JK, Lan F, Shi Y, Segal E and Chang HY: Long Noncoding RNA as
modular scaffold of histone modification complexes. Science.
329:689–693. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Schmitz SU, Grote P and Herrmann BG:
Mechanisms of long noncoding RNA function in development and
disease. Cell Mol Life Sci. 73:2491–2509. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Taylor DH, Chu ETJ, Spektor R and Soloway
PD: Long non-coding RNA regulation of reproduction and development.
Mol Reprod Dev. 82:932–956. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Hecht PM, Ballesteros-Yanez I, Grepo N,
Knowles JA and Campbell DB: Noncoding RNA in the transcriptional
landscape of human neural progenitor cell differentiation. Front
Neurosci. 9:3922015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wu G, Cai J, Han Y, Chen J, Huang ZP, Chen
C, Cai Y, Huang H, Yang Y, Liu Y, et al: LincRNA-p21 regulates
neointima formation, vascular smooth muscle cell proliferation,
apoptosis and atherosclerosis by enhancing p53 activity.
Circulation. 130:1452–1465. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Puthanveetil P, Chen S, Feng B, Gautam A
and Chakrabarti S: Long non-coding RNA MALAT1 regulates
hyperglycaemia induced inflammatory process in the endothelial
cells. J Cell Mol Med. 19:1418–1425. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Leung A, Trac C, Jin W, Lanting L, Akbany
A, Sætrom P, Schones DE and Natarajan R: Novel long non-coding RNAs
are regulated by angiotensin II in vascular smooth muscle cells.
Circ Res. 113:266–278. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Leisegang MS, Fork C, Josipovic I, Richter
FM, Preussner J, Hu J, Miller MJ, Epah J, Hofmann P, Günther S, et
al: Long noncoding RNA MANTIS facilitates endothelial angiogenic
function. Circulation. 136:65–79. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Bhaskaran M and Mohan M: MicroRNAs:
History, biogenesis and their evolving role in animal development
and disease. Vet Pathol. 51:759–774. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Chen LJ, Lim SH, Yeh YT, Lien SC and Chiu
JJ: Roles of microRNAs in atherosclerosis and restenosis. J Biomed
Sci. 19:79–91. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Chistiakov DA, Sobenin IA, Orekhov AN and
Bobryshev YV: Human miR-221/222 in physiological and
atherosclerotic vascular remodeling. Biomed Res Int. 15:1216–1229.
2015.
|
|
20
|
Landskroner-Eiger S, Qiu C, Perrotta P,
Siragusa M, Lee MY, Ulrich V, Luciano AK, Zhuang ZW, Corti F,
Simons M, et al: Endothelial miR-17~92 cluster negatively regulates
arteriogenesis via miRNA-19 repression of WNT signaling. Proc Natl
Acad Sci USA. 112:12812–12817. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Haque R, Iuvone PM, He L, Choi KSC, Ngo A,
Gokhale S, Aseem M and Park D: The MicroRNA-21 signaling pathway is
involved in prorenin receptor (PRR)-induced VEGF expression in
ARPE-19 cells under a hyperglycemic condition. Mol Vision.
23:251–262. 2017.
|
|
22
|
Luo M, Tan X, Mu L, Luo Y, Li R, Deng X,
Chen N, Ren M, Li Y, Wang L, et al: MiRNA-21 mediates the
antiangiogenic activity of metformin through targeting PTEN and
SMAD7 expression and PI3K/AKT pathway. Sci Rep. 7:4521–4534.
2017.PubMed/NCBI
|
|
23
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Li JH, Liu S, Zhou H, Qu LH and Yang JH:
starBase v2.0: Decoding miRNA-ceRNA, miRNA-ncRNA and protein-RNA
interaction networks from large-scale CLIP-Seq data. Nucleic Acids
Res. 42:D92–D97. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Uchida S and Dimmeler S: Long noncoding
RNAs in cardiovascular diseases. Circ Res. 116:737–750. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhang BY, Jin Z and Zhao Z: Long
intergenic noncoding RNA 00305 sponges miR-136 to regulate the
hypoxia induced apoptosis of vascular endothelial cells. Biomed
Pharmacother. 94:238–243. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Li H, Zhu H and Ge J: Long noncoding RNA:
Recent updates in atherosclerosis. Int J Biol Sci. 12:898–910.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Xu L, Wu Z, Chen Y, Zhu Q, Hamidi S and
Navab R: MicroRNA-21 (miR-21) regulates cellular proliferation,
invasion, migration and apoptosis by targeting PTEN, RECK and Bcl-2
in lung squamous carcinoma, Gejiu City, China. PLoS One.
9:5324–5337. 2014.
|
|
29
|
Sabatel C, Malvaux L, Bovy N, Deroanne C,
Lambert V, Gonzalez ML, Colige A, Rakic JM, Noël A, Martial JA and
Struman I: MicroRNA-21 exhibits antiangiogenic function by
targeting rhob expression in endothelial cells. PLoS One.
6:6548–6560. 2011. View Article : Google Scholar
|
|
30
|
Osborn EA and Jaffer FA: Imaging
inflammation and neovascularization in atherosclerosis: Clinical
and translational molecular and structural imaging targets. Curr
Opin Cardiol. 30:671–680. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
De León H, Boué S, Schlage WK, Boukharov
N, Westra JW, Gebel S, Van Hooser A, Talikka M, Fields RB,
Veljkovic E, et al: A vascular biology network model focused on
inflammatory processes to investigate atherogenesis and plaque
instability. J Transl Med. 12:1852014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Luo D, Luo Y, He Y, Zhang H, Zhang R, Li
X, Dobrucki WL, Sinusas AJ, Sessa WC and Min W: Differential
functions of tumor necrosis factor receptor 1 and 2 signaling in
ischemia-mediated arteriogenesis and angiogenesis. Am J Pathol.
169:1886–1898. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Ulitsky I and Bartel DP: lincRNAs:
Genomics, evolution and mechanisms. Cell. 154:26–46. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Di Gesualdo F, Capaccioli S and Lulli M: A
pathophysiological view of the long non-coding RNA world.
Oncotarget. 5:10976–10996. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Gomes CPC, Spencer H, Ford KL, Michel LYM,
Baker AH, Emanueli C, Balligand JL and Devaux Y: Cardiolinc
network: The function and therapeutic potential of long Non-coding
RNAs in cardiovascular development and disease. Mol Ther Nucleic
Acids. 8:494–507. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Boulberdaa M, Scott E, Ballantyne M,
Garcia R, Descamps B, Angelini GD, Brittan M, Hunter A, McBride M,
McClure J, et al: A role for the Long Noncoding RNA SENCR in
commitment and function of endothelial cells. Mol Ther. 24:978–990.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Yan B, Yao J, Liu JY, Li XM, Wang XQ, Li
YJ, Tao ZF, Song YC, Chen Q and Jiang Q: lncRNA-MIAT regulates
microvascular dysfunction by functioning as a competing endogenous
RNA. Circ Res. 116:1143–1156. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Li K, Blum Y, Verma A, Liu Z, Pramanik K,
Leigh NR, Chun CZ, Samant GV, Zhao B, Garnaas MK, et al: A
noncoding antisense RNA in tie-1 locus regulates tie-1 function in
vivo. Blood. 115:133–139. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Lu W, Huang SY, Su L, Zhao BX and Miao JY:
Long Noncoding RNA LOC100129973 suppresses apoptosis by targeting
miR-4707-5p and miR-4767 in vascular endothelial cells. Sci Rep.
6:9533–9547. 2016.
|
|
40
|
Yang R, Li P, Zhang G, Lu C, Wang H and
Zhao G: Long non-coding RNA XLOC_008466 functions as an oncogene in
human non-small cell lung cancer by targeting miR-874. Cell Physiol
Biochem. 42:126–136. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Cerezo AB, Hornedo-Ortega R,
Alvarez-Fernandez MA, Troncoso AM and Garcia-Parrilla MC:
Inhibition of VEGF-induced VEGFR-2 activation and HUVEC migration
by melatonin and other bioactive indolic compounds. Nutrients.
9:E2492017. View Article : Google Scholar : PubMed/NCBI
|