Due to an increasing incidence of diabetes and its
complications, there is an urgent need to identify therapeutic
approaches for diabetes. Both type 1 and type 2 diabetes patients
eventually require lifelong insulin replacement therapy. However,
long-term exogenous insulin replacement therapy is not only
associated with numerous side effects, but can also not delay
and/or prevent the development of diabetes and the emergence of its
complications. In addition, islet transplantation has been studied
but progress in this field is slow due to a lack of islet donors,
organ transplant rejection and low survival rates following
transplantation in humans (1).
Therefore, the realization of endogenous islet cells regeneration
has become the focus of all researchers. At present, a fundamental
method of treating diabetes is regenerative therapy, which
generally involves the development of feasible therapeutic methods
that induce an expansion of adult human β-cells (2). In previous studies, a number of
mechanisms had been proposed to explain the proliferation of
β-cells, including the proliferation of pancreatic duct stem cells
(3), the replication of β-cell
progenitor cells (4) and the
endogenesis of residual β-cells (5).
Insight into the network of β-cell signaling pathways that promote
β-cell proliferation may help to overcome key difficulties in
diabetes therapy. As research methods for the evaluation of
cellular signaling pathways have improved, an increasing number of
pathways have been identified that are associated with β-cell
proliferation, including insulin, growth factors and hormones.
However, difficulties remain in various aspects of diabetes
research, including specific molecular mechanisms remaining unclear
and differences between animal and human trials. Despite these
limitations, more research into the related signal pathways may aid
to develop therapeutic strategies for the treatment of
diabetes.
Insulin signal transduction pathways within
pancreatic β-cells are a complex system, and are mainly composed of
insulin receptors (IR), including the insulin receptor (IR)-related
receptor and insulin-like growth factor 1 (IGF-1) receptor, and
substrates belonging to the insulin receptor substrates (IRS)
family, including IRS, IR tyrosine kinase, phosphoinositide
linositol-3-kinase (PI3K) and its downstream effectors such as
protein kinase B [PKB; also known as serine/threonine kinase
(AKT)], mitogen activated protein kinase (MAPK), MAPK kinase
(MAPKK/MEK), raf protein and rat sarcomas (Ras) proteins (small G
proteins) (6–8). Activated IR stimulates IRS, which is
present in two major forms in β-cells; IRS-1 and IRS-2 (9). IRS-1 is generally distributed in islet
β-cells, and closely regulates glucose-dependent insulin secretion
(9). By contrast, IRS-2 is mainly
located in the peripheral region of the pancreatic islets and at
low levels in β-cells, and positively moderates the compensation
mechanism of β-cells (9). In terms
of regulatory mechanisms, IRS-1 is activated through the activation
of PI3K and calcium channels, among others, while IRS-2 is
activated by PI3K, MAPK, Ras and other pathways, and they
ultimately modulate the growth, proliferation and mitosis of
β-cells (10,11).
The PI3K signaling pathway serves a dominant role in
the regulation of β-cell function (12), which is orchestrated by an intricate
network of mediators. AKT/PKB, acting as a downstream target of
PI3K, regulates the proliferation of β-cells through modulation of
its multiple downstream genes, which include Forkhead box protein
O1 (FOXO1) (13), glycogen synthase
kinase-3 (GSK3) (14) and mammalian
target of rapamycin (mTOR) (8),
among others.
FOXO1 is a member of the phosphorylase Fox gene
family and its transcription activity is modulated through
phosphorylation of PI3K/AKT signaling proteins (15). In β-cells, FOXO1 has been identified
as a weak inhibitor of pancreatic and duodenal homeobox 1 (Pdx1)
(15), which participated in the
maintenance of common features of β-cells, including the regulation
of target gene expression for genes such as glucose transporter 2
(GLUT2), insulin secretion and maintenance of appropriate cell
function (16). This indicates the
pivotal role of FOXO1 in the maintenance of β-cell function through
modulation of Pdx1.
GSK3 has been demonstrated to be a downstream target
of multiple signaling pathways and is associated with cell
proliferation and cell cycle regulation (17). It has been reported that stimulation
of GSK3 negatively regulates β-cell function by maintaining the
sub-cellular localization and stability of Pdx1 (18). Furthermore, upon activation of mTOR,
GSK3 is suppressed by FOXO1, which is activated through
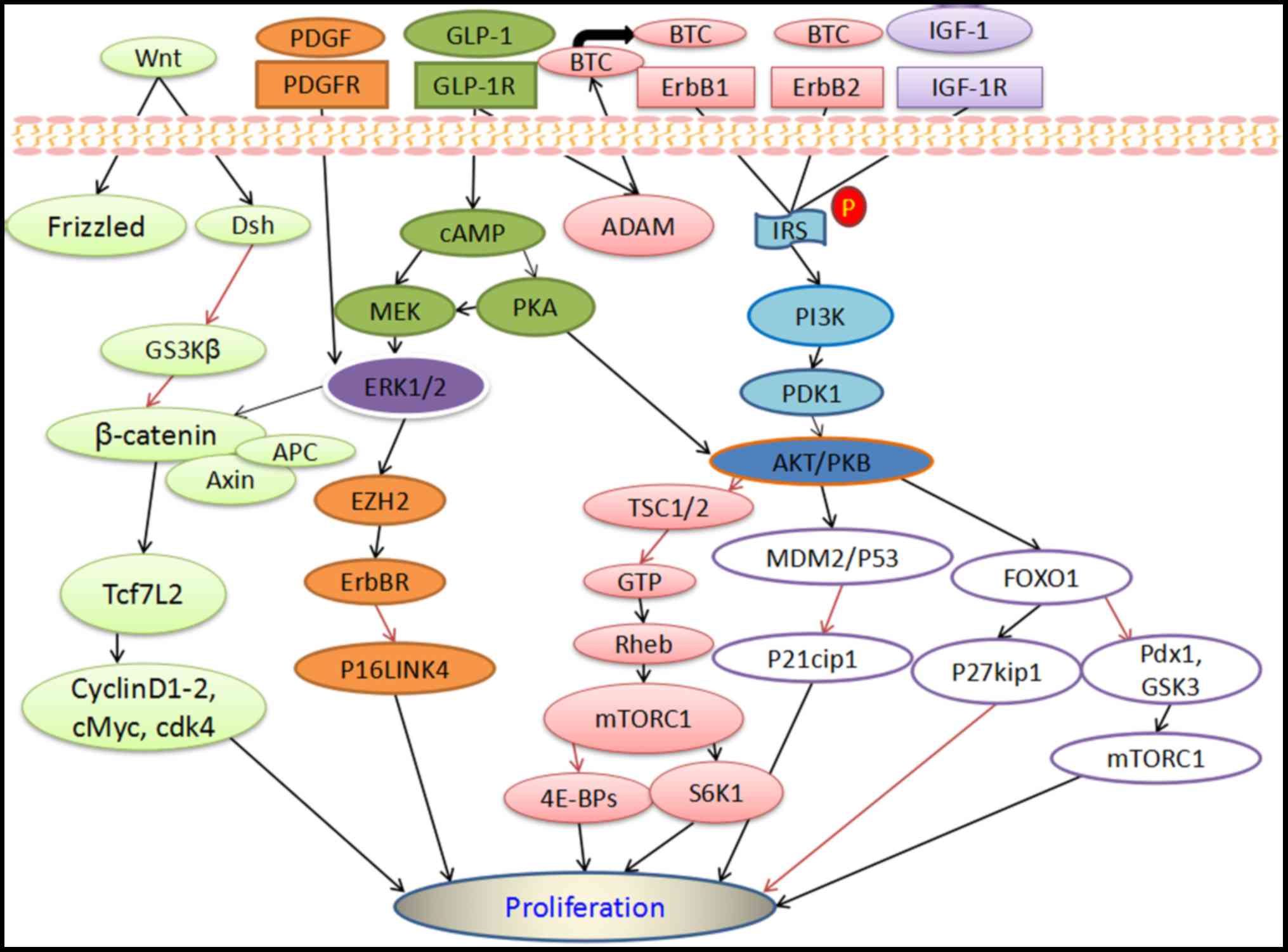
phosphorylated PI3K-AKT (Fig. 1)
(19). mTOR, as a member of the
serine/threonine protein kinase family, is a fundamental component
within two mTOR complexes, mTORC1 and mTORC2 (20–22). The
related signaling proteins of mTOR include PI3K-AKT, AMP activated
protein kinase and growth factors, which influence multiple
metabolic indices such as cellular metabolism, stem cell
differentiation and dedifferentiation, pancreatic β-cell function,
insulin secretion and resistance and cellular survival or death
(21–23). Maiese (24) reported that downregulation of mTOR
and mTORC1 activated PI3K-AKT through a feedback mechanism, which
blocked all anticipated clinical benefits. PI3K-AKT suppressed
activation of tuberous sclerosis complex 1/2 (TSC1/2), which was an
upstream negative regulator of mTORC1, and then S6K1 and 4E-BPs
promoted mRNA translation and synthesis of ribosomes, and
eventually increased β-cell proliferation (Fig. 1) (7,8).
Furthermore, P21cip1 and p27kip1 were inhibitors of the cell cycle
(Fig. 1) (7). PI3K-AKT suppressed P21cip1 by
activating MDM2/P53 gene, and increased β-cell proliferation, while
it enhanced P27kip1 by activating FOXO1 gene, and hindered β-cell
proliferation (Fig. 1) (7). A recent study demonstrated that the
immortalized mouse β-cell line (exclusively expressing IR A
isoform) was sensitive to the mitogenic response induced by IGF-1,
which further activated β-cell proliferation, primarily through
mTORC1 (Fig. 1) (25). Furthermore, in partial pancreatectomy
(60%), mTORC1 exerted a compensatory effect on β-cell mass mainly
through the cyclin D2 pathway in early stages (26).
Recently, studies have focused on the mechanism
behind inulin-mediated fat and glucose metabolism, though few have
investigated protein metabolism (13). For instance, it has been reported
that hyperglycemia promoted compensatory β-cell proliferation to
meet the insulin demand induced by hyperglycemia (27). Lipid metabolism has previously been
investigated in pregnant female mice (10), whereby mice were randomly assigned to
receive either a high-fat diet (MO-HF) or standard chow (MO-SC)
without advance preparation. It was observed that MO-HF induced a
failure in β-cell proliferation and survival by downregulating
IRS1, PI3K and GLUT2 protein, while upregulating insulin protein
and FOXO1 when compared with MO-SC mice (10,27).
Furthermore, a high-fat diet has been demonstrated to negatively
impact on β-cell proliferation, leading to the development of
insulin resistance and D2M characteristics (28,29).
Rhee et al (30) also
suggested that preadipocyte factor 1 (Pref-1) may independently
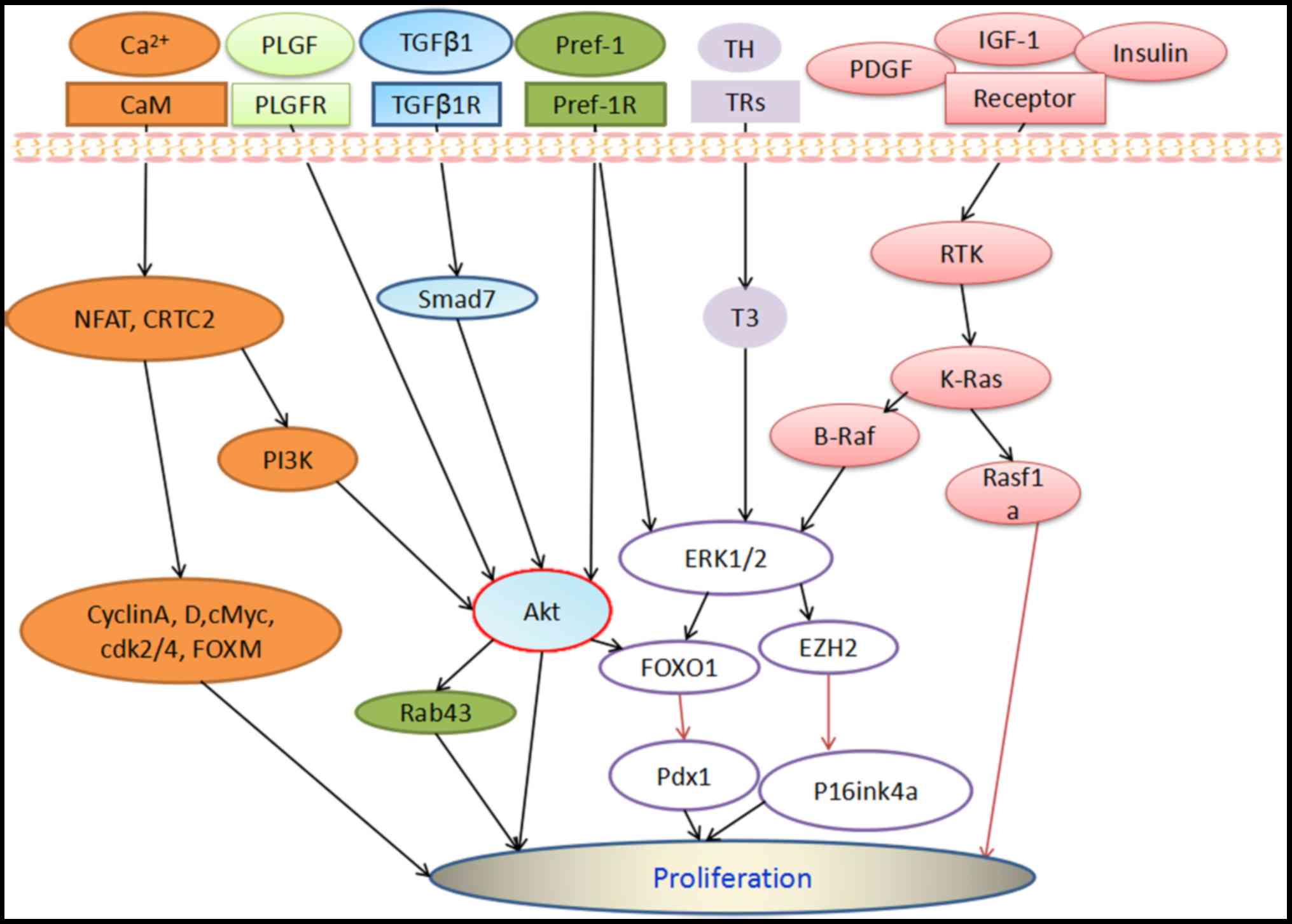
stimulate insulin secretion via AKT-Rab43 (Fig. 2). Finally, there has been dispute
regarding the interplay between protein metabolism and the related
signaling pathways of β-cell proliferation. Chen et al
(13) argued that FOXO1 may inhibit
the expression of albumin, while other proteins such as phycocyanin
have been demonstrated to promote β-cell proliferation by
regulating PI3K-AKT signaling and downstream FOXO1, and ameliorate
diabetes mice by stimulating glucokinase expression and insulin
signaling in the pancreas (11,31). It
has also been reported that Exendin-4 may enhance β-cell
proliferation in some instances by stimulating PI3K-AKT signaling,
potentially via an intermediate ligand-binding activation step
involving corresponding receptors (14).
As an important second messenger, calcium
participates in apoptosis and regulates the synthesis of enzymes
and hormones, including that of insulin. The maintenance of normal
Ca2+ concentration in the body is termed calcium
homeostasis, and is critical to cell survival (32). Previous results have indicated that
disruption of calcium homeostasis may accelerate β-cell death and
lead to type 1 diabetes mellitus (D1M) (32). Furthermore, PI3K signaling stimulated
the autocrine function of pancreatic β-cells to moderate β-cell
proliferation by inhibiting endoplasmic reticulum Ca2+
ATPase, which increased the intracellular Ca2+
concentration and subsequently activated calcium-mediated signaling
pathways (33,34). In calcium-mediated signaling,
increases in intracellular calcium activate calmodulin (CaM), which
has two substrates, namely carbohydrate response element-binding
(CREB)-regulated transcription coactivator-2 (CRTC2) and
transcription factors of the nuclear factor of activated T cells
(NFAT) family (35–38). Notably, dephosphorylated NFAT
translocated to the nucleus and combined with the promoters of
cell-cycle activating agents, including cyclin A and D, cMyc,
cyclin-dependent kinase (cdk) 2 and 4, FOX protein M1 (FOXM1;
Fig. 2) and the promoter of IRS-2,
which participated in PI3K-AKT and Ras-Raf-MAPK signaling to
activate β-cell proliferation (35–37). By
contrast, NFAT function may be improved by interaction with
transcription factors, including CREB, cAMP-response element
modulator and activator protein 1 (38). Changes in cytosolic Ca2+
concentration have been documented to exert a significant effect on
insulin secretion by modulating gene expression from electrically
excitable pancreatic β-cells (39).
In pancreatic β-cells, intracellular high calcium concentration
triggers exocytosis and leads to insulin secretion (40). In addition, previous data suggest
that Ca2+ may be drawn into pancreatic islets either by
the opening L-type voltage-dependent Ca2+ channels or
crosstalk between intracellular Ca2+ concentration and
protein kinase (PK) A/L/C to stimulate insulin secretion (40,41).
Indeed, Ca2+/CaM-dependent protein kinase 2 (CaMKK2) is
considered to be a molecular regulator of insulin function, and
Ca2+/CaM/CaMKK2 has been documented to serve a crucial
role in the modulation of holistic body metabolism (42,43).
Collectively these observations indicate that calcium-mediated
signaling pathways may be therapeutic targets for the modulation of
β-cell proliferation in diabetes.
Three types of Ras protein are expressed in humans,
namely H-Ras, K-Ras, and N-Ras. K-Ras is considered to be the most
important form, acting as a downstream target of multiple cellular
signaling pathways that regulate cell growth, proliferation and
differentiation (44). Chamberlain
et al (45) demonstrated that
K-Ras stimulated the anti-proliferative ras association domain
family 1A (Rasf1a) pathway, which diminished cell proliferation
(Fig. 2). Therefore, K-Ras may serve
a negative regulatory role in the proliferation of β-cells, and
downregulation of the K-Ras gene may enhance proliferation by
reducing Rassf1a activity.
ERKs (also known as classical MAPKs), are diffusely
expressed cellular signaling molecules that participate in the
modulation of cell proliferation (46). In pancreatic β-cells, ERK1 (MAPK3)
and ERK2 (MAPK1) have been documented to be the major expressed
forms of ERK (5). In general, the
binding of a ligand to its cognate tyrosine kinase or G protein
coupled receptor stimulates the small GTPase Ras, resulting in the
formation of activated Ras-GTP, which phosphorylates c-Raf and
subsequently activates MEK, and ultimately activates ERK1/2
(5). Furthermore, a number of growth
factors and hormones have been reported to induce β-cell
proliferation via the activation of ERK1/2. For instance, it was
observed that stimulation of platelet derived growth factor
receptor required activated ERK1/2, leading to upregulation of the
polycomb group protein EZH2 (enhancer of zeste homolog 2, EZH2),
inhibition of P16LINK4 tumor suppressor protein and ultimately
adult β-cell expansion (Fig. 1)
(47). ERK signaling was also
demonstrated to serve a pivotal role in 3,5,3′-Triidothyronine
(T3)-induced β-cell proliferation (Fig. 2) (46). In addition, reactive oxygen
species-facilitated β-cell differentiation by regulating the ERK1/2
pathway (48), and the
anti-apoptotic function of visfatin was mediated by stimulation of
the ERK1/2 signaling pathway (49).
In a study by Ozaki et al (50), a mouse model of diabetes was
established in high-fat diet KKAy mice and leptin
receptor-deficient (db/db) mice. In these mice it was observed that
obesity-associated insulin resistance could be treated with an
inhibitor of MEK (50), indicating
that regulation of ERK signaling may be an effective treatment for
type 2 diabetes (D2M).
Several growth factors have been implicated in the
modulation of Β-cell proliferation, including IGF-1, members of the
epidermal growth factor (EGF) family, nerve growth factor (NGF),
members of the transforming growth factor (TGF)-β superfamily,
placental growth factor (PLGF) and their corresponding receptors
(IGF-1R, EGFR, NGFR, TGFβR and PLGFR, respectively) (51). These mitogen signals have been
associated with multiple downstream pathways, including
PI3K-AKT-mTOR, ERK (Figs. 1 and
2) and janus kinase-signal
transducers and activators of transcription (JAK-STAT), in the
modulation of β-cell proliferation and survival (51).
Multiple downstream proteins of the EGF family,
including EGF, β-cellulin (BTC), TGF-α, heparin-binding (HB)-EGF
and epiregnin, have been implicated in β-cell proliferation
(52). For instance, BTC-dependent
mitogenic signals were involved in the upregulation of IRS and
activation of both EGF/ErbB1 and ErbB2 receptors in β-cells
(Fig. 1) (53). BTC has also been linked to the
IRS-PI3K signaling pathway, which signals through
3′-phosphoinositide-dependent protein kinase 1 (PDK1) to regulate
AKT and FOXO1 (Fig. 1) (53). As members of the EGF superfamily, EGF
and TGF-α have been reported to transiently activate ERK1/2, though
the interaction was not strong enough to induce β-cell
proliferation (53). Furthermore,
constitutively active EGFR protected β-cells against damage and
even death (54). It has also been
reported that excessive nutrients may promote β-cell expansion via
crosstalk between EGFR signaling, FOXM1-mediated cell proliferation
and mTOR activation (55). Further
study into the mechanism of β-cell survival has demonstrated that
β-cell proliferation may be regulated via EGF and Raf-1/MEK/ERK
signaling, which primarily increased the half-life of β-cells
(51).
NGF regulates the development of neuronal cells and
may control the function of non-neuronal cells (56,57). In
islet β-cells, NGF is among the peptides stored by β-cells within
secretary vesicles, and it may interact with β-cell receptors to
trigger autocrine and paracrine effects in response to appropriate
stimulation (57). NGF primarily
acts via two membrane receptors, p75 neurotrophin receptor (P75
NTR) and tropomyosin receptor kinase A (trkA). trkA functions upon
its phosphorylation, and mainly modulates the activation of signals
involved in cell survival, differentiation and proliferation
(58), while P75 NTR recruits
specific intracellular effectors depending on its NGF-bound and
unbound forms (59). A recent study
suggested that islet NGF served a pivotal role in fine-tuning
insulin secretion, to maintain both basal insulin secretion and
biphasic secretion of glucose (60).
Various members of the TGFβ superfamily, including
TGFβ, inhibins, activins, follistains, mullerian inhibitor
substance and bone morphogenetic proteins, have been demonstrated
to function in β-cells (5), though
the exact underlying mechanisms remain unclear. The TGFβ
superfamily, which has been implicated in numerous developmental
processes, including proliferation, differentiation and growth
arrest, served a crucial role in modulating pancreatic endocrine
development and maturation (61–64). In
particular, interplay between the inhibitor TGFβ signaling Smad7
and intracellular mediators of TGFβ signaling were involved in
coordinating this process (61). A
recent investigation into TGFβ signaling demonstrated that
macrophages released TGFβ1 to trigger Smad7 expression, which then
facilitated β-cell proliferation (65). Smad7 was also increased in β-cells
after partial pancreatic duct ligation (PDL) (65). In general, the functions of TGFβ
superfamily members affects downstream Smad proteins in β-cells,
which typically suppresses proliferation (64). Therefore, suppression of TGFβ
signaling may promote β-cell proliferation (65–70). In
cultured human pancreatic duct cells, TGFβ1 induced the
epithelial-mesenchymal transition and inhibited differentiation
into β-cells, and bound with the downstream targets ERK1/2, AKT and
Ras (71). The underlying mechanisms
of pancreatic islet cell expansion may involve an upregulation in
TGFβ signaling (68). Blockade of
TGFβ signaling with short hairpin RNA (shRNA) against TGFβ1R, such
as Alk5 shRNA, suppressed the differentiation and proliferation of
β-cell-derived (BCD) cells, which may be associated with the
AKT-FOXO1 pathway (Fig. 2) (72).
Finally, gestational PLGF may stimulate endothelial
cells within the pancreatic islets to release growth factors that
activate the PI3K-AKT pathway (Fig.
2), thus enhancing β-cell proliferation (73). A previous study demonstrated that
Pref-1 independently facilitated the phosphorylation of AKT and
ERK1/2, and triggered alterations in the expression of FOXO1 and
Pdx1 (Fig. 2) (29).
Hormones including thyroid hormone (TH), growth
hormone (GH), glucagon like peptide 1 (GLP-1), IGF-1, and prolactin
(PRL) and their cognate receptors (TR, GHR, GLP-1R, IGF-1R and
PRLR, respectively) have been critically associated with β-cell
survival, growth, proliferation, differentiation and insulin
secretion, and may be involved in PI3K-AKT, JAK-STAT and ERK
crosstalk (74). A stimulatory
effect of T3 on cellular growth was observed on TRs in several cell
lines. TR expression in the pancreas suggested that pancreatic
β-cell proliferation might be induced by T3 (46). The study demonstrated that T3 induced
pancreatic β-cell proliferation via the ERK signaling pathway
(46). The current review has mainly
focused on the incretin hormone GLP-1, which is primarily released
from intestinal L-cells in response to hormones, nutrients and
neurons, and may serve a significant role in β-cell proliferation
and insulin secretion (75,76). In general, GLP-1 combined with its
receptor (GLP-1R) may enhance insulin secretion. However, it has
been reported that activation of GLP-1R requires a host of complex
proteins interactions (77). Recent
studies have identified multiple novel GLP-1R interactors,
including the Ras superfamily proteins Rab5b and Rab5c, ATPase
H+-transporting lysosomal accessory protein 2 (ATO6Ap2)
and progesterone receptor membrane component 1, which served dual
roles by regulating both GLP-1R signaling and insulin secretion
through EGF receptor and cAMP signaling (77,78). In
β-cells, GLP-1 might activate various signaling pathways, including
cAMP-PKA, PI3K-AKT, ERK1/2 and Wnt (Fig.
1) (79,80). In a study of the hyperglycemic
response, GLP-1 reinforced glucose-stimulated insulin secretion
principally through activation of cAMP-PKA signaling in the β-cell
line INS-1E (81). GLP-1 binding to
GLP-1R has been reported to activate cAMP-PKA, and via ADAM
proteins GLP-1 triggered the secretion of BTC, which acted upon
ErbB1/2 (Fig. 1) (53). Furthermore, a recent report suggested
that geniposide may enhance insulin secretion by activating GLP-1R
in tandem with adenylyl cyclase (AC)/cAMP signaling (82). Yang et al (83) demonstrated that the peroxisome
proliferator-activated receptor β/δ agonist, GW501516 (GW), in
tandem with its transcriptional modulation of GLP-1R, may protect
against lipotoxic apopotosis in β-cells. In addition, conditional
chicken ovalbumin upstream promoter transcription factor II, as a
novel mediator of GLP-1 signaling required for GLP-1 activation,
depended on β-catenin signaling, and its expression was regulated
by TCF7L2, which increased β-cell mass during the neonatal stage
(84).
The present review has described a number of key
hormone pathways implicated in β-cell proliferation, though further
research is required to elucidate the full involvement of hormones
in β-cell proliferation.
The wnt signaling pathway is generally composed of
extracellular wnt ligands, disheveled protein, Frizzled receptor,
β-catenin, axin, GSK3β and adenomatous polyposis colis protein
(Fig. 1) (85,86). The
wnt pathway may be divided into three types: A canonical
wnt/β-catenin pathway, a planar cell polarity (PCP) pathway and a
non-canonical pathway, which includes wnt/Ca2+,
activated phospholipase C, PKC and small GTPase (87,88).
In terms of function, wnt signaling molecules have
been demonstrated to influence pancreatic endocrine development and
regulate the postnatal β-cell functions of insulin secretion,
proliferation, survival and differentiation (89). Bader et al (90) concluded that wnt/PCP genes were
responsible for functional β-cell heterogeneity and induced β-cell
maturation. Fltp, as a wnt/PCP effector and firefly luciferase, is
considered to be a marker gene that may aid to elucidate the
molecular mechanisms of islet cell plasticity and heterogeneity,
and may be a target in endocrine cells for functional β-cell
regeneration in diabetes (91).
Wnt4, a regulator of β-cell proliferation, has been documented to
primarily regulate canonical wnt signaling (85,86).
Krutzfeldt et al (91)
demonstrated that the expression and upregulation of wnt4 was
common in the β-cells of obese mice. Furthermore, stimulation with
exendin-4 enhanced wnt4 expression in β-cells (87). It has also been reported that wnt3a
modulated the proliferation of β-cells and enhanced β-cell function
by activating the wnt/β-catenin pathway, indicating crosstalk with
PI3K signaling (92). Rulifson et
al (93) observed that wnt3a
induced the expression of Cdk4 and cyclin D-1 and −2, and
stimulated enhanced β-cell proliferation in vitro. Regarding
stem cells, canonical wnt signaling facilitated stem cell
proliferation and survival via the expression of β-catenin-related
downstream targets, such as cMyc, which was conducive to the
transplantation of pluripotent stem cells in D1M (94–96).
Afelik et al (97) also
documented that wnt7b was a modulator of pancreatic progenitor cell
development. T cell factor 7-like 2 (TCF7L2) has been identified as
a significant component of wnt/β-catenin signaling (Fig. 1), and a potent factor in the
pathogenesis and progression of D2M (98). Notably, the dominant-negative form or
‘at-risk’ alleles of TCF7L2 served a deleterious role in β-cell
proliferation and insulin secretion (99). Furthermore, the functional
consequences of reduced high-mobility group protein B1 in BCD
(INS1) and human colon cancer (HCT116) cells were reductions in
TCF7L2 mRNA expression, TCF7L2 transcriptional regulatory activity
and glucose triggered insulin secretion (100). A recent study also documented that
geniposide promoted β-cell proliferation and survival by modulating
the β-catenin/TCF7L2 signaling pathway (101).
JAK-STAT is a relatively novel signaling pathway,
which has been associated with multiple cytokines, such as
interleukin (IL)-1, −2 and −6, and hormones such as leptin, GH,
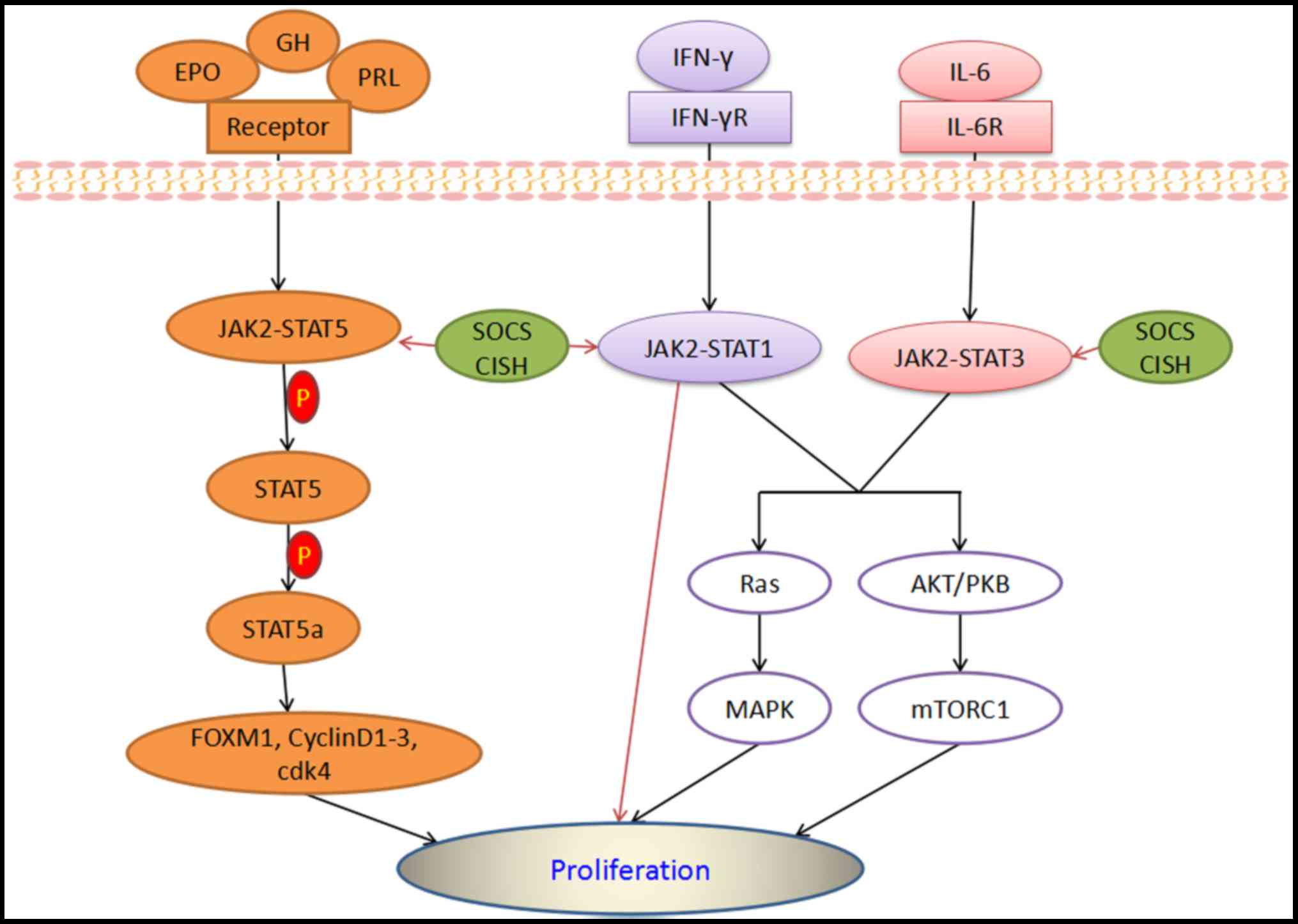
placental lactogens, erythropoietin (EPO) and PRL (Fig. 3) (102–105).
JAK-STAT participates in the regulation of key biological
processes, including cell proliferation, differentiation and
apoptosis (102). Furthermore, JAKs
may serve as a fundamental switch that must be activated through a
cytokine to trigger STAT proteins, which enables downstream target
activation and subsequent regulation of target gene transcription
(106). It has been reported that
activated STAT proteins may also trigger suppressors of cytokine
signaling (SOCS), protein repressors of activated STAT and
transcription of cytokine-inducible SH2 domain-containing proteins
(CISH; Fig. 3) (107,108).
SOCS proteins are considered to be pivotal negative modulators of
JAK-STAT signaling by terminating its upstream signals, of which
four-SOC1, SOC2, SOC3 and CISH have been reported to serve as
primary factors in the JAK-STAT pathway (106,109).
The progressive loss of pancreatic β-cells leads to
D1M, which is closely associated with autoimmune assault (110). During the inflammatory progressive
stage, overexpression of JAK-STAT molecules in pancreatic islets
has been reported to contribute to β-cell dysfunction (111). A recent study documented that
BRD0476, a novel inhibitor of β-cell apoptosis, impeded
inteferon-γ-induced JAK2 and STAT1 signaling to facilitate β-cell
survival (112). Unlike general
JAK-STAT pathway suppressors, BRD0476 blocked JAK-STAT signaling
pathways via ubiquitin-specific peptidase 9X (USP9X) without
inhibiting the kinase function of any JAK (112). With regard to PRL, it has been
observed that human β-cells failed to expand in response to PRL,
potentially due to a deficiency of PRL receptors, among other
explanations (5). Notably, a failure
of β-cell expansion could be overcome by overexpression of murine
STAT5a, due to the resulting upregulation and nuclear translocation
of cdk4 and cyclins D1-3 (Fig. 3).
This STAT5 signaling has been linked with β-cell cycle activity
(113). Furthermore, EPO, acting
via the EPO receptor, has been reported to stimulate β-cell
proliferation through JAK2-STAT5 (Fig.
3) (114). De Groef et
al (115) demonstrated that PDL
stimulated the expression of multiple cytokines that served as
positive ligands of STAT3 signaling in β-cells. In addition, β-cell
cycling was enhanced by activating STAT3 with IL-6 (Fig. 3). STAT3 played an important role in
maintaining the stability of β-cells by regulating the cell cycle
and protecting against DNA damage (115).
Toll like receptor 4 (TLR4) is principally expressed
in the innate immune system, and is considered to be involved in DM
(116). In D2M, the expression of
TLR4 may increase due to activation of the nuclear factor-κB
(NF-κB) signaling pathway. While NF-κB has not been associated with
the inflammatory reaction in diabetes, it may have an important
influence on glucose metabolism and insulin reaction. NF-kB binds
to the promoters of innate immune genes, particularly those of
pro-inflammatory cytokines, including tumor necrosis factor-α
(TNF-α) and IL-1β, which is considered to exert positive feedback
on TLR4 expression (117–119). Sepehri et al (119) reported that TLR4 was upregulated in
the peripheral blood mononuclear cells of patients with D2M
patients, which was independent of sex, age, body mass index and
blood sugar content. Furthermore, TLR4 is considered to be a
significant candidate in the related complications of D2M,
including nephropathy (120) and
atherosclerosis (121).
Cellular signaling pathways are substantial targets,
and are capable of stimulating β-cell proliferation, as
demonstrated in various islets and β-cell systems. As such, they
may be useful in future treatment strategies for diabetes. Cellular
signaling pathways involving interactions with ligands, receptors
and the proliferative machinery of β-cells are complex systems. The
present review evaluated the potential underlying mechanisms of
insulin, growth factor, Wnt, JAK-STAT and TLR4 signaling. While
various cellular signaling mechanisms were discussed, not all
signaling pathways implicated in β-cell proliferation were
included, such as those involving islet cell autoantigen-512,
inhibin, activin, muscarinic and octeocalicn.
Cellular signaling molecules and their downstream
targets form an intricate network, which may directly or indirectly
regulate β-cell proliferation and survival. While an increased
number of studies have investigated cellular signaling,
difficulties remain in their applications. For instance, the
expression of cellular signals in β-cells varies among different
species, ages and tissues. Furthermore, various cellular signals
have not been investigated, and their specific functions and
mechanisms within β-cells remain unclear and disputed by
researchers. Though notably, excessive activation of signaling
pathways may have negative impacts on β-cells.
In conclusion, multiple other therapeutic strategies
may be used for the treatment of diabetes, including stem cell
differentiation induction and islet transplantation. Cellular
signaling pathways may have untapped potential in the induction of
β-cell proliferation. Identification of appropriate molecular
targets within these pathways may aid to develop novel strategies
in the treatment of diabetes and improve the outcome for
patients.
Not applicable.
The present work was supported by the Yunnan
Province Project Education Fund (grant no. 2015J117).
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
WJJ wrote the manuscript, collected images and
analyzed the data; YCP processed the images; KMY analyzed the data,
processed the images and revised the manuscript. All authors have
read and approved the manuscript.
Not applicable.
Not applicable.
The authors declare that they have no competing
interests.
|
1
|
Farney AC, Sutherland DE and Opara EC:
Evolution of islet transplantation for the last 30 years. Pancreas.
1:8–20. 2016. View Article : Google Scholar
|
|
2
|
Tiwari S, Roel C, Wills R, Casinelli G,
Tanwir M, Takane KK and Fiaschi-Taesch NM: Early and late G1/S
cyclins and Cdks act complementarily to enhance authentic human
β-cell proliferation and expansion. Diabetes. 10:3485–3498. 2015.
View Article : Google Scholar
|
|
3
|
Wang GS, Rosenberg L and Scott FW: Tubular
complexes as a source for islet neogenesis in the pancreas of
diabetes-prone BB rats. Lab Invest. 5:675–688. 2005. View Article : Google Scholar
|
|
4
|
Dor Y, Brown J, Martinez OI and Melton DA:
Adult pancreatic beta-cells are formed by self-duplication rather
than stem-cell differentiation. Nautre. 429:41–46. 2004. View Article : Google Scholar
|
|
5
|
Stewart AF, Hussain MA, Garcia-Ocana A,
Vasavada RC, Bhunshan A, Bernal-Mizrachi E and Kulkarni RN: Human
β-cell proliferation and intracellular signaling: Part 3. Diabetes.
4:1872–1885. 2015. View Article : Google Scholar
|
|
6
|
Saltiel AR and Kahn CR: Insulin signaling
and the regulation of glucose and lipid metabolism. Nature.
414:799–806. 2001. View
Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kulkarni RN, Mizrachi EB, Ocana AG and
Stewart AF: Human β-cell proliferation and intracellular signaling:
Driving in the dark without a road map. Diabetes. 9:2205–2213.
2012. View Article : Google Scholar
|
|
8
|
Pende M, Kozma SC, Jaquet M, Oorschot V,
Burcelin R, Le Marchand-Brustel Y, Kluperman J, Thorens B and
Thomas G: Hypoinsulinaemia, glucose intolerance and diminished
beta-cell size in S6K1-deficient mice. Nature. 408:994–997. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Arajuo EP, Amaral ME, Souza CT, Bordin S,
Ferreira F, Saad MJ, Boschero AC, Maqalhaes EC and Velloso LA:
Blockade of IRS1 in isolated rat pancreatic islets improves
glucose-induced insulin secretion. FEBS Lett. 531:437–442. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Bringhenti I, Ornellas F,
Mandarim-de-Lacerda CA and Aguila MB: The insulin-signaling pathway
of the pancreatic islet is impaired in adult mice offspring of
mothers fed a high-fat diet. Nutrition. 32:1138–1143. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ou Y, Ren Z, Wang J and Yang X:
Phycocyanin ameliorates alloxan-induced diabetes mellitus in mice:
Involved in insulin signaling pathway and GK expression. Chem Biol
Interact. 247:49–54. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kaneko K, Ueki K, Takahashi N, Hashimoto
S, Okamoto M, Awazawa M, Okazaki Y, Ohsugi M, Inabe K, Umehara T,
et al: Class IA phosphatidylinositol 3-kinase in pancreatic β cells
controls insulin secretion by multiple mechanisms. Cell Metab.
12:619–632. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Chen Q, Lu M, Monks BR and Birnabaum MJ:
Insulin is required to maintain albumin expression by inhibiting
forkhead box o1 protein. J Biol Chem. 291:2371–2378. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wang C, Chen X, Ding X, He Y, Gu C and
Zhou L: Exendin-4 promotes beta cell proliferation via PI3K/Akt
signaling pathway. Cell Physiol Biochem. 35:2223–2232. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Katoh M and Katoh H: Human FOX gene family
(Review). Int J Oncol. 25:1495–1500. 2004.PubMed/NCBI
|
|
16
|
Seyer P, Vallois D, Poitry-Yamate C,
Schutz F, Metref S, Tarussio D, Maechler P, Staels B, Lanz B,
Grueter R, et al: Hepatic glucose sensing is required to preserve β
cell glucose competence. J Clin Invest. 123:1662–1676. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Huang L, Jiang X, Gong L and Xing D:
Photoactivation of Akt1/GSK3β Isoform-Specific signaling axis
promotes pancreatic β-cell regeneration. J Cell Biochem.
116:1741–1754. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Boucher MJ, Selander L, Carlsson L and
Edlund H: Phosphorylation marks IPF1/PDX1 protein for degradation
by glycogen synthase kinase 3-dependent mechanisms. J Biol Chem.
281:6395–6403. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Morral N: Novel targets and therapeutic
strategies for type 2 diabetes. Trends Endocrinol Metab.
14:169–175. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Foster KG and Fingar DC: Mammalian target
of rapamycin (mTOR): Conducting the cellular signaling symphony. J
Biol Chem. 285:14071–14077. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Duzgn Z, Eroglu Z and Biray-Avci C: Role
of mTOR in glioblastoma. Gene. 575:187–190. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Maiese K: Novel nervous and multi-system
regenerative therapeutic strategies for diabetes mellitus with
mTOR. Neural Regen Res. 11:372–385. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wang J, Yang X and Zhang J: Bridges
between mitochondrial oxidative stress, ER stress and mTOR
signaling in pancreatic β cells. Cell Signal. 28:1099–1104. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Maiese K: Targeting molecules to medicine
with mTOR, autophagy and neurodegenerative disorders. Br J Clin
Pharmacol. 82:1245–1266. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Escribano O, Gomez-Hernandez A,
Diaz-Castroverde S, Nevado C, Garcia G, Otoro YF, Perdomo L, Beneit
N and Benito M: Insulin receptor isoform A confers a higher
proliferative capability to pancreatic β cells enabling glucose
availability and IGF-1 signaling. Mol Cell Endocrinol. 409:82–91.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Li W, Zhang H, Nie A, Ni Q, Li F, Ning G,
Li X, Gu Y and Wang Q: mTORC1 pathway mediates beta cell
compensatory proliferation in 60% partial-pancreatectomy mice.
Endocrine. 53:117–128. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
McCurdy CE and Klemm DJ: Adipose tissue
insulin sensitivity and macrophage recruitment: Does PI3K pick the
pathway. Adipocyte. 2:135–142. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Bringhenti I, Moraes-Teixeira JA, Cunha
MR, Ornellas F, Mandarim-de-lacerda CA and Aguila MB: Maternal
obesity during the preconception and early life periods alters
pancreatic development in early and adult life in male mouse
offspring. PLoS One. 8:e557112013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Cerf ME, Williams K, Chapman CS and Louw
J: Compromised beta-cell development and beta-cell dysfunction in
weanling offspring from dams maintained on a high-fat diet during
gestation. Pancreas. 34:347–353. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Rhee M, Lee H, Kim JW, Ham DS, Park HS,
Yang HK, Shin JY, Cho JH, Kim YB, Youn BS, et al: Preadipocyte
factor 1 induces pancreatic ductal cell differentiation into
insulin-producing cells. Sci Rep. 6:239602016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Gao Y, Liao G, Xiang C, Yang X, Cheng X
and Ou Y: Effects of phycocyanin on INS-1 pancreatic β-cell
mediated by PI3K/Akt/FoxO1 signaling pathway. Int J Biol Macromol.
83:185–194. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Ahn C, An BS and Jeung EB: Streptozotocin
induces endoplasmic reticulum stress and apoptosis via disruption
of calcium homeostasis in mouse pancreas. Mol Cell Endocrinol.
412:302–308. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Khan S, Yan-Do R, Duong E, Wu X, Bautista
A, Cheley S, MacDonald PE and Braun M: Autocrine activation of P2Y1
receptors couples Ca (2+) release in human pancreatic beta cells.
Diabetologia. 57:2235–2245. 2014. View Article : Google Scholar
|
|
34
|
Roper MG, Qian WJ, Zhang BB, Kulkarni RN,
Kahn CR and Kennedy RT: Effect of the insulin mimetic L-783,281 on
intracellular Ca2 and insulin secretion from pancreatic beta-cells.
Diabetes. 51 Suppl:S43–S49. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Demozay D, Tsunekawa S, Briaud I, Shah R
and Rhodes CJ: Specific glucose-induced control of insulin receptor
substrate-2 expression is mediated via Ca2+-dependent
calcineurin/NFAT signaling in primary pancreatic islet β-cells.
Diabetes. 60:2892–2902. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Goodyer WR, Gu X, Liu Y, Bottino R,
Crabtree GR and Kim SK: Neonatal β cell development in mice and
humans is regulated by calcineurin/NFAT. Dev Cell. 23:21–34. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Nguidjoe E, Sokolow S, Bigabwa S, Pachera
N, D'Amico E, Allagnat F, Vanderwinden JM, Sener A, Manto M,
Depreter M, et al: Heterozygous inactivation of the Na/Ca exchanger
increases glucose-induced insulin release, β-cell proliferation,
and mass. Diabetes. 60:2076–2085. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Lawrence MC, Naziruddin B, Levy MF,
Jackson A and McGlynn K: Calcineurin/nuclear factor of activated T
cells and MAPK signaling induce TNF-{alpha} gene expression in
pancreatic islet endocrine cells. J Biol Chem. 286:1025–1036. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Gilon P, Chae HY, Rutter GA and Ravier MA:
Calcium signaling in pancreatic β-cells in health and in type 2
diabetes. Cell Calcium. 5:340–361. 2014. View Article : Google Scholar
|
|
40
|
Kappel VD, Frederico MJ, Postal BG, Mendes
CP, Cazarolli LH and Silva FR: The role of calcium in intracellular
pathways of rutin in rat pancreatic islets: Potential insulin
secretagogue effect. Eur J Pharmacol. 702:264–268. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Castro AJ, Cazarolli LH, de-Carvalho FK,
da Luz G, Altenhofen D, dos Santos AR, Pizzolatti MG and Silva FR:
Acute effect of 3β-hidroxihop-22(29)ene on insulin secretion is
mediated by GLP-1, potassium and calcium channels for the glucose
homeostasis. J Sterois Biochem Mol Biol. 150:112–122. 2015.
View Article : Google Scholar
|
|
42
|
Marcelo KL, Ribar T, Means CR, Tsimelzon
A, Stevens RD, Llkayeva O, Bain JR, Hilsenbeck SG, Newgard CB,
Means AR and York B: Research resource: Roles for
calcium/calmodulin-dependent protein kinase kinase 2 (CaMKK2) in
systems metabolism. Mol Endocrinol. 30:557–572. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Markwardt ML, Seckinger KM and Rizzo MA:
Regulation of glucokinase by intracellular calcium levels in
pancreatic β cells. J Biol Chem. 291:3000–3009. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Nussinov R, Tsai CJ, Muratcioglu S, Jang
H, Gursoy A and Keskin O: Principles of K-Ras effector organization
and the role of oncogenic K-Ras in cancer initiation through G1
cell cycle. Expert Rev Proteomics. 12:669–682. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Chamberlain CE, Scheel DW, Mcglynn K, Kim
H, Miyatsuka T, Wang J, Nguyen V, Zhao S, Mavropoulos A, Abraham
AG, et al: Menin determines K-RAS proliferative outputs in
endocrine cells. J Clin Invest. 124:4093–4101. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Kim TK, Lee JS, Jung HS, Ha TK, Kim SM,
Han N, Lee EJ, Kim TN, Kwon MJ, Lee SH, et al: Triiodothyronine
induces proliferation of pancreatic β-cells through the MAPK/ERK
pathway. Exp Clin Endocrinol Diabetes. 122:240–245. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Chen H, Gu X, Liu Y, Wang J, Wirt SE,
Bottino R, Schorle H, Sage J and Kim SK: PDGF signaling controls
age-dependent proliferation in pancreatic β-cells. Nature.
478:349–355. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Hoarau E, Chandra V, Rustin P, Scharfmann
R and Duvillie B: Pro-oxidant/antioxidant balance controls
pancreatic β-cell differentiation through the ERK1/2 pathway. Cell
Death Dis. 5:e14872014. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Xiang RL, Mei M, Su YC, Li L, Wang JY and
Wu LL: Visfatin protects rat pancreatic β-cells against
IFN-γ-induced apoptosis through AMPK and ERK1/2 signaling pathways.
Biomed Environ Sci. 28:169–177. 2015.PubMed/NCBI
|
|
50
|
Ozaki KI, Awazu M, Tamiya M, Lwasaki Y,
Harada A, Kugisaki S, Tanimura S and Kohno M: Targeting the ERK
signaling pathway as a potential treatment for insulin resistance
and type 2 diabetes. Am J Physiol Endocrino Metab. 310:E643–E651.
2016. View Article : Google Scholar
|
|
51
|
Wang H, Gambosova K, Cooper ZA, Holloway
MP, Kassai A, Lzquierdo D, Cleveland K, Boney CM and Altura RA: EGF
regulates surviving stability through the Raf-1/ERK pathway in
insulin-secreting pancreatic β-cells. BMC Mol Biol. 11:662010.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Ernesto BM, Kulkarni RN, Scott DK,
Mauvais-Jarvis F, Stewart AF and Garcia-Ocana A: Human β-cell
proliferation and intracellular signaling part 2: Still driving in
the dark without a road map. Diabetes. 63:819–831. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Oh YS, Shin S, Lee YJ, Kim EH and Jun HS:
Betacellulin-induced beta cell proliferation and regeneration is
mediated by activation of ErbB-1 and ErbB-2 receptors. PLoS One.
6:e238942011. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Hakonen E, Ustinov J, Eizirik DL, Sariola
H, Miettinen PJ and Otonkoski T: In vivo activation of the PI3K-Akt
pathway in mouse beta cells by the EGFR mutation L858R protects
against diabetes. Diabetologia. 57:970–979. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Zarrouki B, Benterki I, Fontes G, Peyot
ML, Seda O, Prentki M and Poitout V: Epidermal growth factor
receptor signaling promotes pancreatic β-cell proliferation in
response to nutrient excess in rats through mTOR and FOXM1.
Diabetes. 63:982–993. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Zhang CC, Yin X, Cao CY, Wei J, Zhang Q
and Gao JM: Chemical constituents from hericium erinaceus and their
ability to stimulate NGF-mediated neurite outgrowth on PC12 cells.
Bioorg Med Chem Lett. 25:5078–5082. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Rosenbaum T, Vidaltamayo R, Sanchez-Soto
MC, Zentella A and Hiriart M: Pancreatic beta cells synthesize and
secrete nerve growth factor. Proc Natl Acad Sci USA. 95:7784–7788.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Freund V and Frossard N: Expression of
nerve growth factor in the airways and its possible role in asthma.
Prog Brain Res. 146:335–346. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Roux PP and Barker PA: Neurotrophin
signaling through the P75 neurotrophin receptor. Prog Neurobiol.
67:203–233. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Pingitore A, Caroleo MC, Cione E,
Castanera-Gonzalez R, Huang GC and Perasud SJ: Fine tuning of
insulin secretion by release of nerve growth factor from mouse and
human islet β-cells. Mol Cell Endocrinol. 436:23–32. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
61
|
EI-Gohary Y, Tulachan S, Guo P, Welsh C,
Wiersch J, Prasadan K, Paredes J, Shiota C, Xiao X, Wada Y, et al:
Smad signaling pathways regulate pancreatic endocrine development.
Dev Biol. 378:83–93. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Feng Z, Zi Z and Liu X: Measuring TGF-β
ligand dynamics in culture medium. Methods Mol Biol. 1344:379–389.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Richardson CC, To K, Foot VL, Hauge-Evans
AC, Carmignac D and Christie MR: Increased perinated remodeling of
the pancreas in somatostatin-deficient mice: Potential role of
transforming growth factor-beta signaling in regulating beta cell
growth in early life. Horm Metab Res. 47:56–63. 2015.PubMed/NCBI
|
|
64
|
Xiao X, Wiersch J, EI-Gohary Y, Guo P,
Prasadan K, Paredes J, Welsh C, Shiota C and Gittes GK: TGFβ
receptor signaling is essential for inflammation-induced but not
β-cell workload-induced β-cell proliferation. Diabetes.
62:1217–1226. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Blum B, Roose AN, Barrrandon O, Maehr R,
Arvanites AC, Davidow LS, Davis JC, Peterson QP, Rubin LL and
Melton DA: Reversal of β cell de-differentiation by a small
molecule inhibitor of the TGFβ pathway. Elife. 3:e028092014.
View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Bruun C, Christensen GL, Jacobsen ML,
Kanstrup MB, Jensen PR, Fjordvang H, Mandrup-Poulsen T and
Billestrup N: Inhibition of beta cell growth and function by bone
morphogenetic proteins. Diabetologia. 57:2546–2554. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
67
|
EI-Gohary Y, Tulachan S, Wiersch J, Guo P,
Welsh C, Prasadan K, Paredes J, Shiota C, Xiao X, Wada Y, et al: A
smad signaling network regulates islet cell proliferation.
Diabetes. 63:224–236. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Lei C, Zhou X, Pang Y, Mao Y, Lu X, Li M
and Zhang J: TGF-β signaling prevents pancreatic beta cell death
after proliferation. Cell Prolif. 48:356–362. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Wu H, Mezghenna K, Marmol P, Guo T,
Moliner A, Yang SN, Berggren PO and Lbanez CF: Differential
regulation of mouse pancreatic islet insulin secretion and Smad
proteins by activin ligands. Diabetologia. 57:148–156. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Xiao X, Gaffar I, Guo P, Wiersch J,
Fischbach S, Peirish L, Song Z, El-Gohary Y, Prasadan K, Shiota G
and Gittes GK: M2 macrophages promote beta-cell proliferation by
up-regulation of SMAD7. Prov Natl Acad Sci USA. 111:E1211–E1220.
2014. View Article : Google Scholar
|
|
71
|
Shin JA, Hong OK, Lee HJ, Jeon SY, Kim JW,
Lee SH, Cho JH, Lee JM, Choi YH, Chang SA, et al: Transforming
growth factor-β induces epithelial to mesenchymal transition and
suppresses the proliferation and transdifferentiation of cultured
human pancreatic duct cells. J Cell Biochem. 112:179–188. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Toren-Haritan G and Efrat S: TGFβ pathway
inhibition redifferentiates human pancreatic islet β cells expanded
in vitro. PLoS One. 9:e01391682015. View Article : Google Scholar
|
|
73
|
Li J, Ying H, Cai G, Guo Q and Chen L:
Pre-Eclampsia-associated reduction in placental growth factor
impaired beta cell proliferation through PI3K signaling. Cell
Physiol Biochem. 36:34–43. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Huang Y and Chang Y: Regulation of
pancreatic islet beta-cell mass by growth factor and hormone
signaling. Prog Mol Biol Transl Sci. 121:321–349. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Lee YS and Jun HS: Anti-diabetic actions
of glucagon-like peptide-1 on pancreatic beta-cells. Metabolism.
63:9–19. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Tian L and Jin T: The incretin hormone
GLP-1 and mechanisms underlying its secretion. J Diabetes.
8:753–765. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Dai FF, Bhattacharjee A, Liu Y, Batchuluun
B, Zhang M, Wang XS, Huang X, Luu L, Zhu D, Gaisano H and Wheeler
MB: A novel GLP1 receptor interacting protein ATP6ap2 regulates
insulin secretion in pancreatic beta cells. J Biol Chem.
290:25045–25061. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Zhang M, Robitaille M, Showalter AD, Huang
X, Liu Y, Bhattacharjee A, Willard FS, Han J, Froese S, Wei L, et
al: Progesterone receptor membrane component 1 is a functional part
of the glucagon-like peptide-1 (GLP-1) receptor complex in
pancreatic β cells. Mol Cell Proteomics. 13:3049–3062. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Campbell JE and Drucker DJ: Pharmacology,
physiology, and mechanisms of incretin hormone action. Cell Metab.
17:819–837. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Lavine JA and Attie AD: Gastrointerstinal
hormones and the regulation of β-cell mass. Ann NY Acad Sci.
1212:41–58. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Kong X, Yan D, Wu X, Guan Y and Ma X:
Glucotoxicity inhibits cAMP-protein kinase A-potentiated
glucose-stimulated insulin secretion in pancreatic β-cells. J
Diabetes. 7:378–385. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Zhang Y, Ding Y, Zhong X, Guo Q, Wang H,
Gao J, Bai T, Ren L, Guo Y, Jiao X and Liu Y: Geniposide acutely
stimulates insulin secretion in pancreation β-cells by regulating
GLP-1 receptor/cAMP signaling and ion channels. Mol Cell
Endocrinol. 430:89–96. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Yang Y, Tong Y, Gong M, Lu Y, Wang C, Zhou
M, Yang Q, Mao T and Tong N: Activation of PPARβ/δ protects
pancreatic β cells from palmitate-induced apoptosis by upregulating
the expression of GLP-1 receptor. Cell Signal. 26:268–278. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Boutant M, Ramos OH, Tourrel-Cuzin C,
Movassat J, Llias A, Vallois D, Planchais J, Pegorier JP, Schuit F,
Petit PX, et al: COUP-TFII controls mouse pancreatic β-cell mass
through GLP-1-β-catenin signaling pathways. PLoS One. 7:e308472012.
View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Nelson WJ and Nusse R: Convergence of Wnt,
beta-catenin, and cadherin pathways. Science. 303:1483–1487. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Nusse R: Wnt signaling in disease and in
development. Cell Res. 15:28–32. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Heller C, Kuhn MC, Mulders-Opgenoorth B,
Schott M, Willenberg HS, Scherbaum WA and Schinner S: Exendin-4
upregulates the expression of wnt-4, a novel regulator of
pancreatic β-cell proliferation. Am J Physiol Endocrinol Metab.
301:E864–E872. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Xue G, Romano E, Massi D and MandaIa M:
Wnt/β-catenin signaling in melanoma: Preclinical rationale and
novel therapeutic insights. Cancer Treat Rev. 49:1–12. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Schinner S, Ulgen F, Papewalis C, Schott
M, Woelk A, Vidal-Puig A and Scherbaum WA: Regulation of insulin
secretion, glucokinase gene transcription and beta cell
proliferation by adipicyte-derived wnt signaling molecules.
Diabetologia. 51:147–154. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Bader E, Migliorini A, Gegg M, Moruzzi N,
Gerdes J, Roscioni SS, Bakhti M, Brandl E, Irmler M, Beckers J, et
al: Indentification of proliferative and mature β-cells in the
islets of Langerhans. Nature. 535:430–434. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Krutzfeldt J and Stoffel M: Regulation of
wingless-type MMTV integration site family (WNT) signaling in
pancreatic islets from wild-type and obese mice. Diabetologia.
53:123–127. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Gui S, Yuan G, Wang L, Zhou L, Xue Y, Yu
Y, Zhang J, Zhang M, Yang Y and Wang DW: Wnt3a regulates
proliferation, apoptosis and function of pancreatic NIT-1 β cells
via activation of IRS2/PI3K signaling. J Cell Biochem.
114:1488–1497. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Rulifson IC, Karnik SK, Heiser PW, Ten
Berge D, Chen H, Gu X, Taketo MM, Nusse R, Hebro M and Kim SK: Wnt
signaling regulates pancreatic beta cell proliferation. Proc Nati
Acad Sci USA. 104:6247–6252. 2007. View Article : Google Scholar
|
|
94
|
He X, Han W, Hu SX, Zhang MZ, Hua JL and
Peng S: Canonical wnt signaling pathway contributes to the
proliferation and survival in porcine pancreatic stem cells (PSCs).
Cell Tissue Res. 362:379–388. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Sarkar S and Mandal C, Sangwan R and
Mandal C: Coupling G2/M arrest to the wnt/β-catenin pathway
restrains pancreatic adcnocarcinoma. Endocr Relat Cancer.
21:113–125. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Zhang YQ, Morris JP, Yan W, Schofield HK,
Gurney A, Simeone DM, Millar SE, Hoey T, Hebrok M and Pasca di
Magliano M: Canonical wnt signaling is required for pancreatic
carcinogenesis. Cancer Res. 73:4909–4922. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Afelik S, Pool B, Schmerr M, Penton C and
Jensen J: Wnt7b is required for epithelial progenitor growth and
operates during epithelial-to-mesenchymal signaling in pancreatic
development. Dev Biol. 399:204–217. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Lyssenko V: The transcription factor
7-like 2 gene and increased risk of type 2 diabetes: An update.
Curr Opin Clin Nutr Metab Care. 11:385–392. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Takamoto I, Kubota N, Nakaya K, Kumagai K,
Hashimoto S, Kubota T, Inoue M, Kajiwara E, Katsuyama H, Obata A,
et al: TCF7L2 in mouse pancreatic beta cells plays a crucial role
in glucose homeostasis by regulating beta cell mass. Diabetologia.
57:542–553. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Zhou Y, Oskolkov N, Shcherbina L, Ratti J,
Kock KH, Su J, Martin B, Oskolkova MZ, Goransson O, Bacon J, et al:
HMGB1 binds to the rs7903146 locus in TCF7L2 in human pancreatic
islets. Mol Cell Endocrinol. 430:138–145. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Yao DD, Yang L, Wang Y, Liu C, Wei YJ, Jia
XB, Yin W and Shu L: Geniposide promotes beta-cell regeneration and
survival through regulating β-catenin/TCF7L2 pathway. Cell Death
Dis. 6:e17462015. View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Kiu H and Nicholson SE: Biology and
significance of the JAK/STAT signaling pathways. Growth Factors.
30:88–106. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
O'Shea JJ and Plenge R: JAK and STAT
signaling molecules in immunoregulation and immune-mediated
disease. Immunity. 36:542–550. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Zhuang S: Regulation of STAT signaling by
acetylation. Cell Signal. 25:1942–1931. 2013. View Article : Google Scholar
|
|
105
|
Brooks AJ, Dai W, O'Mara ML, Abankwa D,
Chhabra Y, Pelekanos RA, Gardon O, Tunny KA, Blucher KM, Morton CJ,
et al: Mechanism of activation of protein kinase JAK2 by the growth
hormone recetor. Science. 344:12497832014. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Liongue C, Taznin T and Ward AC: Signaling
via the CytoR/JAK/STAT/SOCS pathway: Emergence during evolution.
Mol Immunol. 71:166–175. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Linossi EM, Babon JJ, Hilton DJ and
Nicholson SE: Suppression of cytokine signaling: The SOCS
perspective. Cytokine Growth Factor Rev. 24:241–248. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Shuai K and Liu B: Regulation of
gene-activation pathways by PIAS protein in the immune system. Nat
Rev Immunol. 5:593–605. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Trengove MC and Ward AC: SOCS proteins in
development and disease. Am J Exp Clin Immunol. 2:1–29. 2013.
|
|
110
|
Fleyel T, Brorsson C, Nielsen LB, Miani M,
Bang-Berthelsen CH, Friedrichsen M, Overgaard AJ, Berchtold LA,
Wiberg A, Poulsen P, et al: CTSH regulates β-cell function and
disease progression in newly diagnosed type 1 diabetes patients.
Proc Natl Acad Sci USA. 111:10305–10310. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Stanley WJ, Litwak SA, Quah HS, Tan SM,
Kay TW, Tiganis T, de Haan JB, Thomas HE and Gurzov EN:
Inactivation of protein tyrosine phosphatases enhances interferon
signaling in pancreatic islets. Diabetes. 64:2489–2496. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Chou DH, Vetere A, Choudhary A, Scully SS,
Schenone M, Tang A, Gomez R, Burns SM, Lundh M, Vital T, et al:
Kinase-independent small-molecule inhibition of JAK-STAT signaling.
J Am Chem Soc. 137:7929–7934. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
113
|
Chen H, Kleinberger JW, Takane KK, Salim
F, Fiaschi-Taesch N, Pappas K, Parsons R, Jiang J, Zhang Y, Liu H,
et al: Augmented stat5 signaling bypasses multiple impediments to
lactogen-mediated proliferation in human β-cells. Diabetes.
64:3784–3797. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
114
|
Choi D, Schroer SA, Lu SY, Wang L, Wu X,
Liu Y, Zhang Y, Gaisano HY, Wagner KY, Wu H, et al: Erythropoietin
protects against diabetes through direct effects on pancreatic beta
cells. J Exp Med. 207:2831–2842. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
115
|
De-Groef S, Renmans D, Cai Y, Leuckx G,
Roels S, Staels W, GradwohI G, Baeyens L, Heremans Y, Martens GA,
et al: STAT3 modulates β-cell cycling in injured mouse pancreas and
protects against DNA damage. Cell Death Dis. 7:e22722016.
View Article : Google Scholar : PubMed/NCBI
|
|
116
|
Shao L, Zhang P, Zhang Y and Ma A:
Inflammatory unbalance of TLR3 and TLR4 in PCI patients with or
without type 2 diabetes mellitus. Immunol Lett. 161:81–88. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
117
|
Kruger B, Yin N, Zhang N, Yadav A, Coward
W, Lai G, Zang W, S Heeger P, Bromberg JS, Murphy B, et al:
Islet-expressed TLR2 and TLR4 sense injury and mediate early graft
failure after transplantation. Eur J Immunol. 40:2914–2924. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
118
|
Yang X, Haghiac M, Glazebrook P, Minium J,
Catalano PM and Hanguel-de Mouzon S: Saturated fatty acids enhance
TLR4 immune pathways in human trophoblasts. Hum Reprod.
30:2152–2159. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
119
|
Sepehri Z, Kiani Z, Nasiri AA, Mashhadi
MA, Javadian F, Haghighi A, Kohan F, Bahari A and Sargazi A: Human
Toll like receptor 4 gene expression of PBMCs in diabetes mellitus
type 2 patients. Cell Mol Biol. 61:92–95. 2015.PubMed/NCBI
|
|
120
|
Verzila D, Cappuccino L, D'Amato E,
Villaggio B, Gianiorio F, Mij M, Simonato A, Viazzi F, Salvidia G
and Garibotto G: Enhanced glomerular Toll-like receptor 4
expression and signaling in patients with type 2 diabetic
nephropathy and microalbuminuria. Kidney Int. 86:1229–1243. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
121
|
Baldan A, Ferronato S, Olivato S, Malerba
G, Scuro A, Veraldi GF, Gelati M, Ferrari S, Mariotto S, Pignati
PF, et al: Cyclooxygenase 2, toll-like receptor 4 and interleukin
1β mRNA expression in atherosclerotic plaques of type 2 diabetic
patients. Inflamm Res. 63:851–858. 2014. View Article : Google Scholar : PubMed/NCBI
|