Introduction
Nasopharyngeal carcinoma (NPC) is a malignant tumor
affecting the head and neck and has a particularly high incidence
in Southern China (1,2). The onset and development of NPC is
closely associated with genetic factors, Epstein-Barr viral
infection, air pollution and excessive contact with trace elements,
including nickel (3). Poorly
differentiated squamous cell carcinoma is the most common type of
NPC, whereas the incidence of other types of NPC, including highly
differentiated squamous cell carcinoma, adenocarcinoma and
vesicular nucleus cell carcinoma, is much lower (4). Most patients with NPC are sensitive to
radiotherapy, which is the preferred modality of treatment
(5).
The overall survival rate of patients with NPC has
been increasing; however the prognosis of patients with advanced
NPC remains poor and the 5-year survival rate of such patients is
only 30–40% (6,7). Tumor recurrence and distant metastasis
are the primary factors affecting the prognosis of patients with
NPC (8). It has been demonstrated
that 70–80% of patients with NPC already have lymph node metastasis
upon diagnosis and 60% of patients with NPC experience neck lymph
node enlargement as their initial symptom (9). Therefore, it is important to determine
the molecular mechanism of invasion and metastasis of NPC, as this
may improve the early diagnosis and treatment of patients with
NPC.
Tight junctions are important structures that
maintain cell connections and polarity and are composed of a
variety of proteins (10). The
disordered expression of tight junctions is closely associated with
the proliferation, invasion, metastasis and apoptosis of tumors
(11,12). Claudin (CLDN) 3 and CLDN4 are
important proteins in the intestinal epithelial cell barrier and
their disordered expression induces tumor metastasis (13,14).
CLDN3 expression is downregulated in early gastric cancer, whereas
CLDN4 overexpression is closely associated with the development of
gastric cancer (15,16). Furthermore, CLDN6, CLDN7 and CLDN9
promote the proliferation, invasion and metastasis of gastric
cancer cells (17,18).
CLDN1 is one of the key structural proteins in the
tight junction protein family and is distributed across the surface
of the cell membrane in the form of a transmembrane structure that
crosses the membrane four times. CLDN1 interacts with the
isoenzymes of creatine kinase, tight junction proteins ZO1, ZO2,
ZO3 and proteins containing the PDZ domain, to pass signals inside
and outside cells and maintain the physical barrier function of
tight junctions (19,20). The expression of CLDN1 is upregulated
in NPC and is associated with lymph node metastasis and clinical
staging; however, the function of CLDN1 in NPC remains unclear
(21).
The epithelial-mesenchymal transition (EMT) is a
process during which epithelial cells lose their polarity and gain
interstitial cell properties, including increased invasiveness and
metastasis, and decreased rates of apoptosis. The EMT is a key step
in tumor metastasis and inhibiting the EMT thus inhibits tumor
invasion and metastasis (22).
Previous studies have demonstrated that numerous genes and
signaling pathways are involved in the EMT in epithelial tumors.
Inhibiting enhancer of zeste homolog 2 expression and enhancing
forkhead box protein C2 expression promotes the EMT in ovarian
cancer cells (23). In addition,
microRNA-630 regulates the invasion and metastasis of esophageal
squamous carcinoma cells by inhibiting the EMT (24). Tight junction proteins are associated
with the EMT in tumors (25);
however, to the best of our knowledge, the effect of CLDN1 on the
EMT in NPC cells has not yet been reported. It has also been
demonstrated that the Wnt/β-catenin signaling pathway serves an
important role in the EMT and β-catenin aggregation in cytoplasm
stimulates the entry of β-catenin into the nucleus. This triggers
the transcription of EMT-related genes and the onset of the EMT
(26). The present study assessed
the biological effects and molecular mechanisms of CLDN1 in NPC,
with the aim of determining the effect of tight junction proteins
in NPC.
Materials and methods
Cells
The human nasopharyngeal epithelial cell line NP69
was purchased from the Cell Bank of Chinese Academy of Sciences
(Shanghai, China). The human nasopharyngeal carcinoma cell line
NPC-TW01 was purchased from the American Type Culture Collection
(Manassas, VA, USA). All cells were cultured in RPMI-1640 medium
containing 10% fetal bovine serum (both obtained from BD
Biosciences, Franklin Lakes, NJ, USA) at 37°C and 5%
CO2. Cells were passaged once they reached 80–90%
confluence. The medium was replenished every 2 days.
CLDN1 silencing
To determine the biological functions of CLDN1 in
NPC, NPC-TW01 cells exhibiting silenced CLDN1 expression were
constructed. NPC-TW01 cells were seeded into 24-well plates at a
density of 1×105 cells/well. NPC-TW01 cells were divided
into a small interfering RNA (siR)-negative control (NC) group and
an siR-CLDN1 group (25 pmol/µl; Hanbio Biotechnology Co., Ltd.,
Shanghai, China). Cells in the siR-CLDN1 group were transfected
with LV-GFP-PURO-siR-CLDN1 lentivirus (MOI=20; Hanbio Biotechnology
Co., Ltd.) using Lipofectamine 3000 (Thermo Fisher Scientific,
Inc., Waltham, MA, USA). Following 12 h cultivation, RPMI-1640
medium was replenished. Subsequently, cells in siR-NC and siR-CLDN1
groups were transfected with siR-NC and siR-β-catenin (rescue
group; 25 pmol/µl; Hanbio Biotechnology Co., Ltd.), respectively.
On the next day, RPMI-1640 medium was replenished. Following 48 h
of transfection, the cells were used for subsequent
experiments.
CDN1 overexpression
NPC-TW01 cells were divided into an NC group and a
CLDN1 group and transfected with a LV-GFP-PURO-CLDN1 lentivirus.
Following 12 h cultivation, RPMI-1640 medium was replenished.
Subsequently, cells in the NC and CLDN1 groups were transfected
with pcDNA3.1-NC and pcDNA3.1-β-catenin (rescue group; 0.5 µg DNA;
Hanbio Biotechnology Co., Ltd.), respectively. On the next day,
RPMI-1640 medium was replenished. Following 48 h of transfection,
the cells were used for subsequent experiments.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Cells were trypsinized and lysed with 1 ml TRIzol
(Thermo Fisher Scientific Inc., Waltham, MA, USA). Following lysis,
total RNA was extracted using the phenol chloroform method
(27). RNA purity was determined at
A260/A280 using an ultraviolet spectrophotometer (Nanodrop ND1000;
Thermo Scientific, Inc.). cDNA was obtained from 1 µg RNA following
reverse transcription with a PrimeScript RT Reagent kit (Takara
Biotechnology, Co., Ltd., Dalian, China) and stored at −20°C. qPCR
was performed using a SYBR Green RT-qPCR kit (Takara Biotechnology
Co., Ltd.) and GAPDH was used as an internal reference. The qPCR
system (20 µl) consisted of 10 µl SYBR EX Taq-Mix, 0.5 µl forward
primer (CLDN1, 5′-CTGGGAGGTGCCCTACTTTG-3′; GAPDH,
5′-AGAAGGCTGGGGCTCATTTG-3′), 0.5 µl reverse primer (CLDN1,
5′-ACACGTAGTCTTTCCCGCTG-3′; GAPDH, 5′-AGGGGCCATCCACAGTCTTC-3′), 1
µl cDNA and 8 µl ddH2O. qPCR was performed in triplicate
for each sample. The thermocycling conditions for qPCR were:
Initial denaturation at 95°C for 5 min; 40 cycles of denaturation
at 95°C for 1 min, annealing at 60°C for 30 sec, elongation at 72°C
for 20 sec and final extension at 72°C for 1 min. The
2−ΔΔCq method was used to determine the relative
expression of mRNA (28).
Western blotting
At 72 h after transfection, cells (1×106)
were mixed with precooled radio immunoprecipitation assay lysis
buffer (1 ml; 50 mM Tris-base, 1 mM EDTA, 150 mM NaCl, 0.1% sodium
dodecyl sulfate, 1% Triton X-100 and 1% sodium deoxycholate;
Beyotime Institute of Biotechnology, Shanghai, China) and lysed for
30 min on ice. Following sonication in an ice bath performed three
times each at 5 sec (with a 20 sec interval), the mixture was kept
on ice for 1 min and centrifuged at a speed of 12,000 × g at 4°C
for 15 min. The supernatant was used to determine protein
concentration using a bicinchoninic acid protein concentration
determination kit [RTP7102; Real-Times (Beijing) Biotechnology Co.,
Ltd., Beijing, China]. Protein samples (20 µg/lane) were then mixed
with 5X SDS loading buffer prior to denaturation in a boiling water
bath for 10 min. Samples were then subjected to 10% SDS-PAGE at 100
V. Subsequently, resolved proteins were transferred to
polyvinylidene difluoride membranes on ice (300 mA, 2 h). Following
blocking with 50 g/l skim milk at room temperature for 1 h,
membranes were incubated with polyclonal rabbit anti-human
E-cadherin 1 (1:1,000; cat. no. ab15148; Abcam, Cambridge, UK),
polyclonal rabbit anti-human vimentin (1:2,000; cat. no. ab45939;
Abcam), polyclonal rabbit anti-human CLDN1 (1:1,000; cat. no.
ab15098; Abcam), polyclonal rabbit anti-human β-catenin (1:1,000;
cat. no. AF0066; Beyotime Institute of Biotechnology), monoclonal
mouse anti-human H3 (1:1,000; cat. no. AF0009; Beyotime Institute
of Biotechnology), and monoclonal mouse anti-human GAPDH (1:4,000;
cat. no. AF0006; Beyotime Institute of Biotechnology) primary
antibodies at 4°C overnight. Following 5 washes with
phosphate-buffered saline with Tween 20 (5 min/wash), membranes
were incubated with goat anti-rabbit (cat. no. A0208; for target
proteins) or goat anti-mouse (cat. no. A0216; for GAPDH)
horseradish peroxidase-conjugated secondary antibodies (1:3,000;
Beyotime Institute of Biotechnology) for 1 h at room temperature.
Membranes were then washed 5 times with phosphate-buffered saline
with Tween-20 (5 min/wash). Membranes were developed using an
enhanced chemiluminescence detection kit (Sigma-Aldrich; Merck
KGaA, Darmstadt, Germany). Image lab software v3.0 (Bio-Rad
Laboratories, Inc., Hercules, CA, USA) was used to acquire and
analyze imaging signals. The expression of E-cadherin 1, vimentin
CLDN1 and β-catenin were expressed relative to that of GAPDH, while
nucleo-β-catenin was expressed relative to that of H3.
Cell-counting kit 8 (CCK-8) assay
All cells were seeded in 96-well plates at a density
of 3,000/well. At 24, 48 and 72 h, the medium was discarded and
cells were washed twice with phosphate-buffered saline, followed by
the addition of RPMI-1640 medium (BD Biosciences) and 10% CCK-8
reaction reagent (Beyotime Institute of Biotechnology). Following
incubation at 37°C for 1 h, the absorbance of each well was
measured at 450 nm for plotting cell proliferation curves. Each
sample was tested in triplicate.
Cell migration and invasion
assays
Transwell chambers (8 µm diameter and 24 wells;
Corning Inc., Corning, NY, USA) were used to evaluate the migration
ability of cells. Cell suspension (200 µl; 2×105 cells)
in serum-free RPMI-1640 medium (BD Biosciences) was added to the
upper chamber. In the lower chamber, 500 µl RPMI-1640 medium
supplemented with 10% fetal bovine serum was added. Following 24 h
incubation, cells in the upper chamber were wiped using a cotton
swab. Then, the chamber was fixed using 4% formaldehyde for 10 min
at room temperature and then stained using Giemsa stain at room
temperature for 1 min. Following 3 washes, cells that migrated to
the other side of the chamber were counted in five fields using a
light microscope (magnification, ×100) to evaluate migration
ability.
The same number of cells as the migration assay were
seeded in the upper chamber for the invasion assay. Matrigel (BD
Biosciences) was used to determine the invasive ability of cells.
Matrigel was diluted with serum-free RPMI-1640 medium at a ratio of
1:1. In the upper chamber, 100 µl diluted Matrigel was added and
kept at 37°C for 1 h. In the lower chamber, 500 µl RPMI-1640 medium
supplemented with 10% fetal bovine serum was added. Following 72 h
incubation, cells in the upper chamber were wiped using cotton
swab. The chamber was fixed using 4% formaldehyde for 10 min at
room temperature and then stained using Giemsa stain at room
temperature for 1 min. Following 3 washes, cells that moved to the
other side of the chamber were counted using a microscope to
evaluate invasive ability.
Statistical analysis
Results were analyzed using SPSS 17.0 statistical
software (IBM Corp., Armonk, NY, USA). The data were expressed as
the mean ± standard deviation. Differences between two groups were
compared using Student's t-test and those among multiple groups
were compared using one-way analysis of variance. Subsequently, a
post hoc multiple comparison test (Tukey's honest significance
test) was performed. P<0.05 was determined to indicate a
statistically significant difference.
Results
CLDN1 expression is elevated in NPC
cell lines
To determine the expression of CLDN1 in NPC cell
lines, RT-qPCR and western blotting were performed. The results
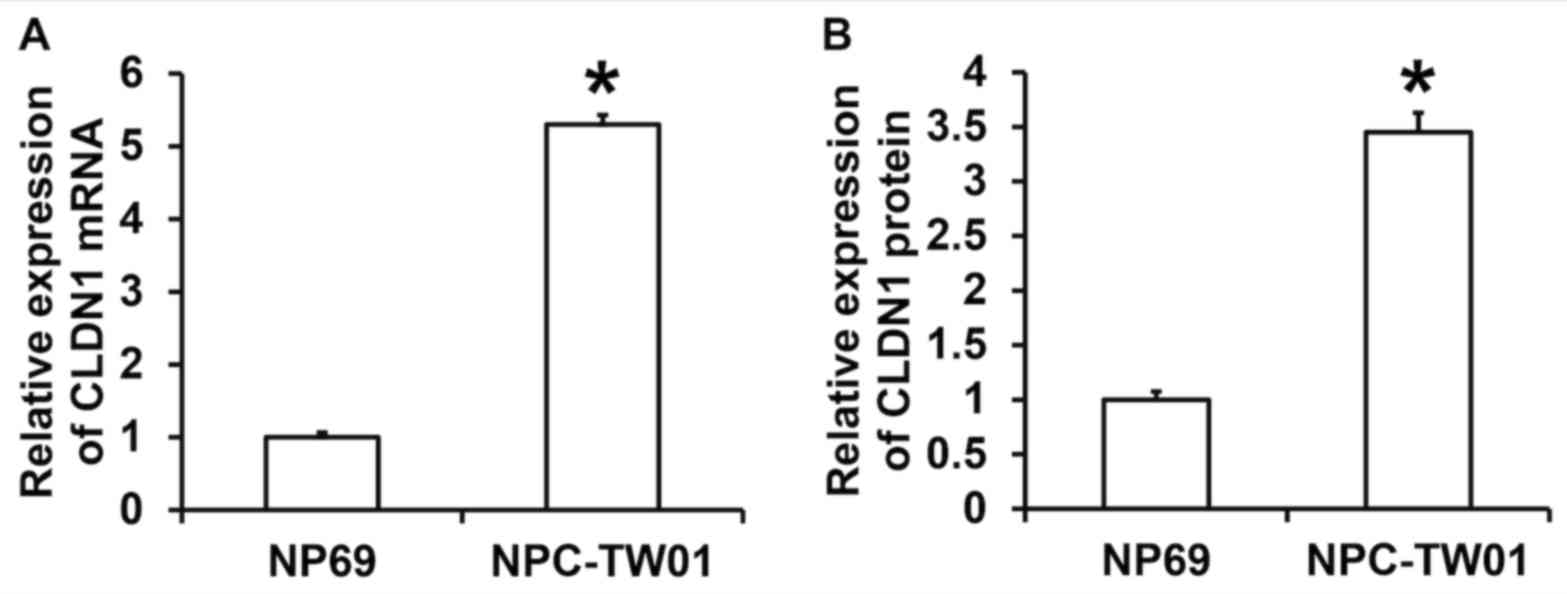
demonstrated that the expression of CLDN1 mRNA in NPC-TW01 cells
was significantly higher than in NP69 cells (P<0.05; Fig. 1A). The results of western blotting
demonstrated that the expression of CLDN1 protein in NPC-TW01 cells
was significantly higher than in NP69 cells (P<0.05; Fig. 1B). These results demonstrate that
CLDN1 expression is significantly elevated in NPC cells.
CLDN1 silencing inhibits the
proliferation of NPC cells whereas CLDN1 overexpression promotes
the proliferation of NPC cells
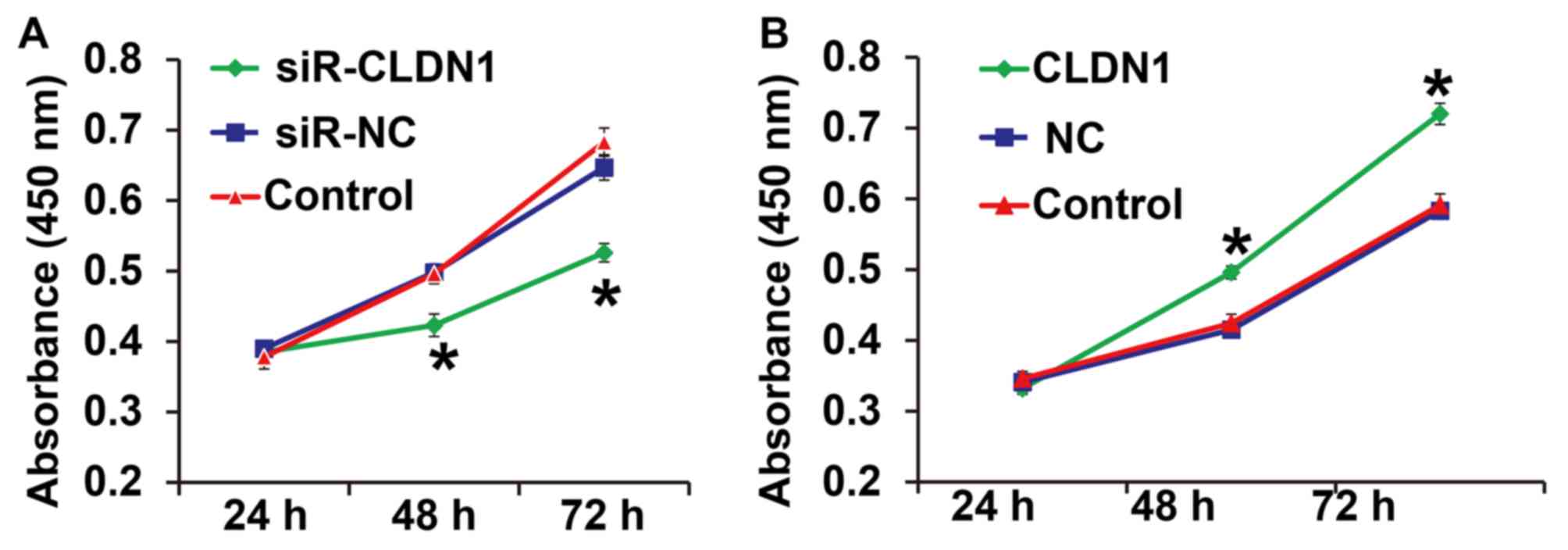
To measure cell proliferation, a CCK-8 assay was
performed. The results demonstrated that the proliferation of
NPC-TW01 cells transfected with siR-CLDN1 was significantly lower
compared with cells in the siR-NC group at 48 and 72 h (P<0.05;
Fig. 2A). By contrast, the
proliferation of NPC-TW01 cells transfected with CLDN1 was
significantly higher than that of cells in the NC group at 48 and
72 h (P<0.05; Fig. 2B). These
results demonstrate that CLDN1 silencing inhibits the proliferation
of NPC cells, whereas CLDN1 overexpression promotes the
proliferation of NPC cells.
CLDN1 promotes the migration and
invasion of NPC cells
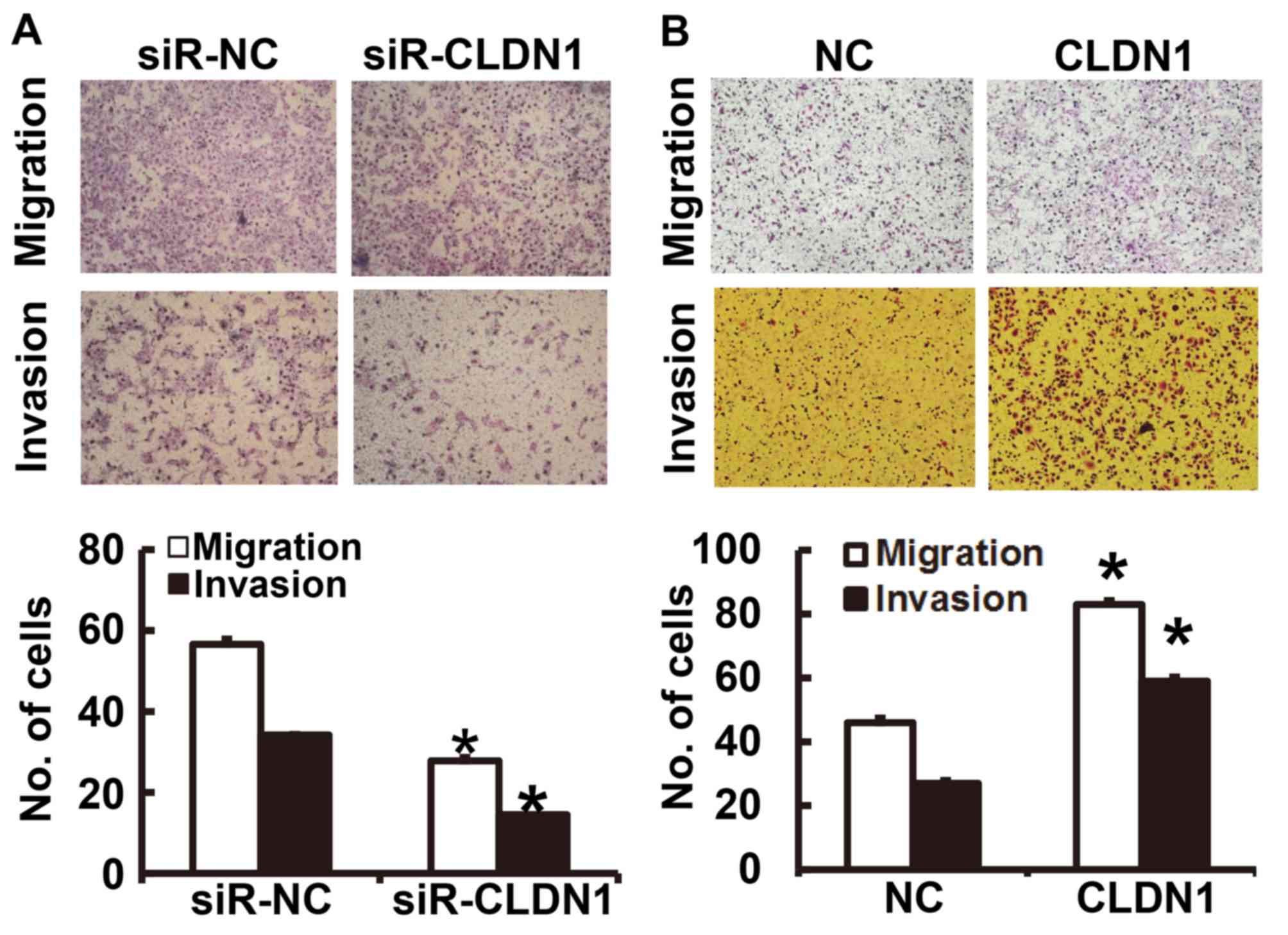
To determine the migration and invasion of NPC
cells, a Transwell assay was employed. The results of the migration
and invasion assays demonstrated that the number of NPC-TW01 cells
transfected with siR-CLDN1 that passed through the Transwell
chamber membrane was significantly lower than that of the siR-NC
group (P<0.05; Fig. 3A). By
contrast, the number of NPC-TW01 cells overexpressing CLDN1 that
passed through the Transwell chamber membrane was significantly
higher than those in the NC group (P<0.05; Fig. 3B). These results suggest that CLDN1
promotes the migration and invasion of NPC cells.
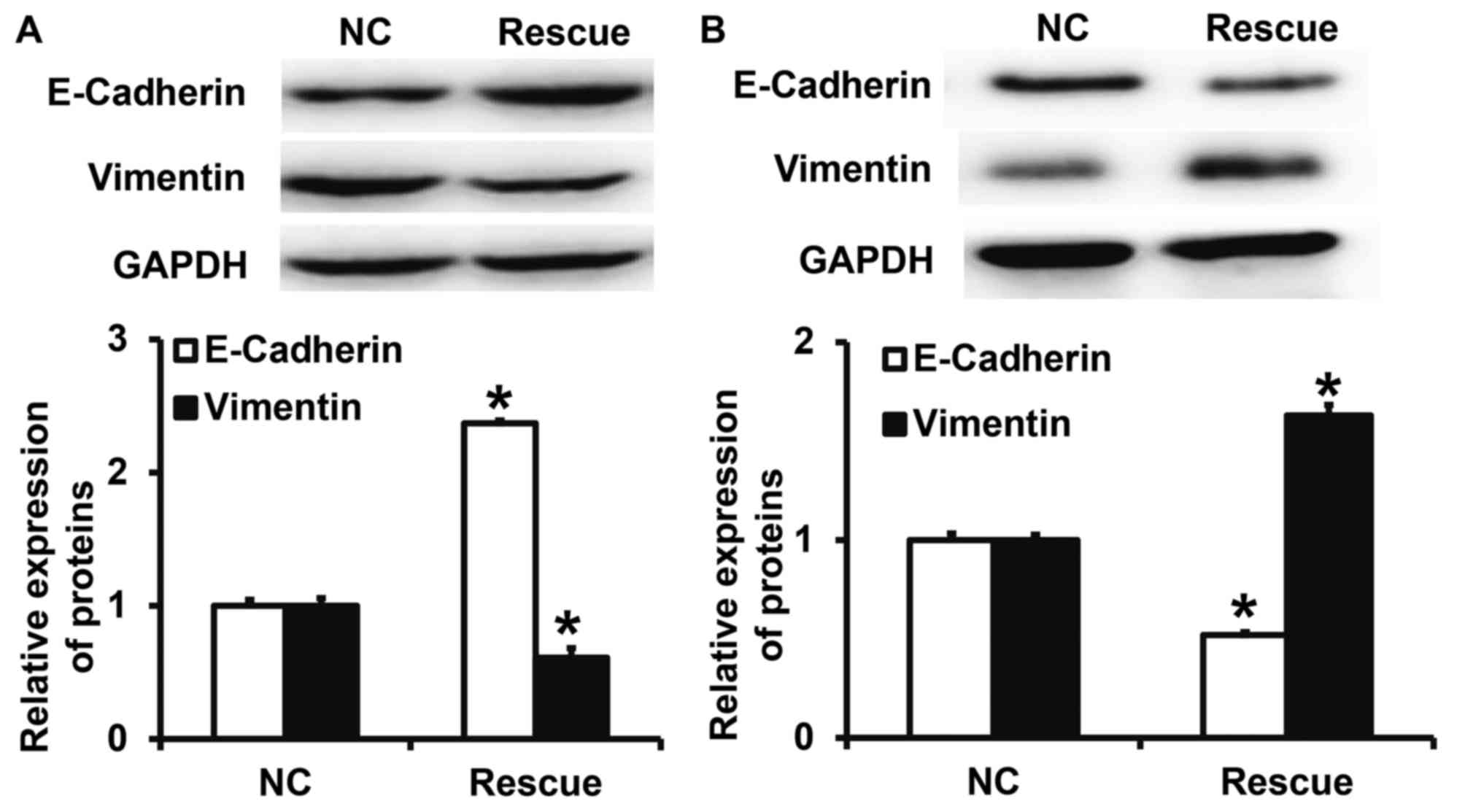
CLDN1 overexpression promotes the EMT
in NPC cells
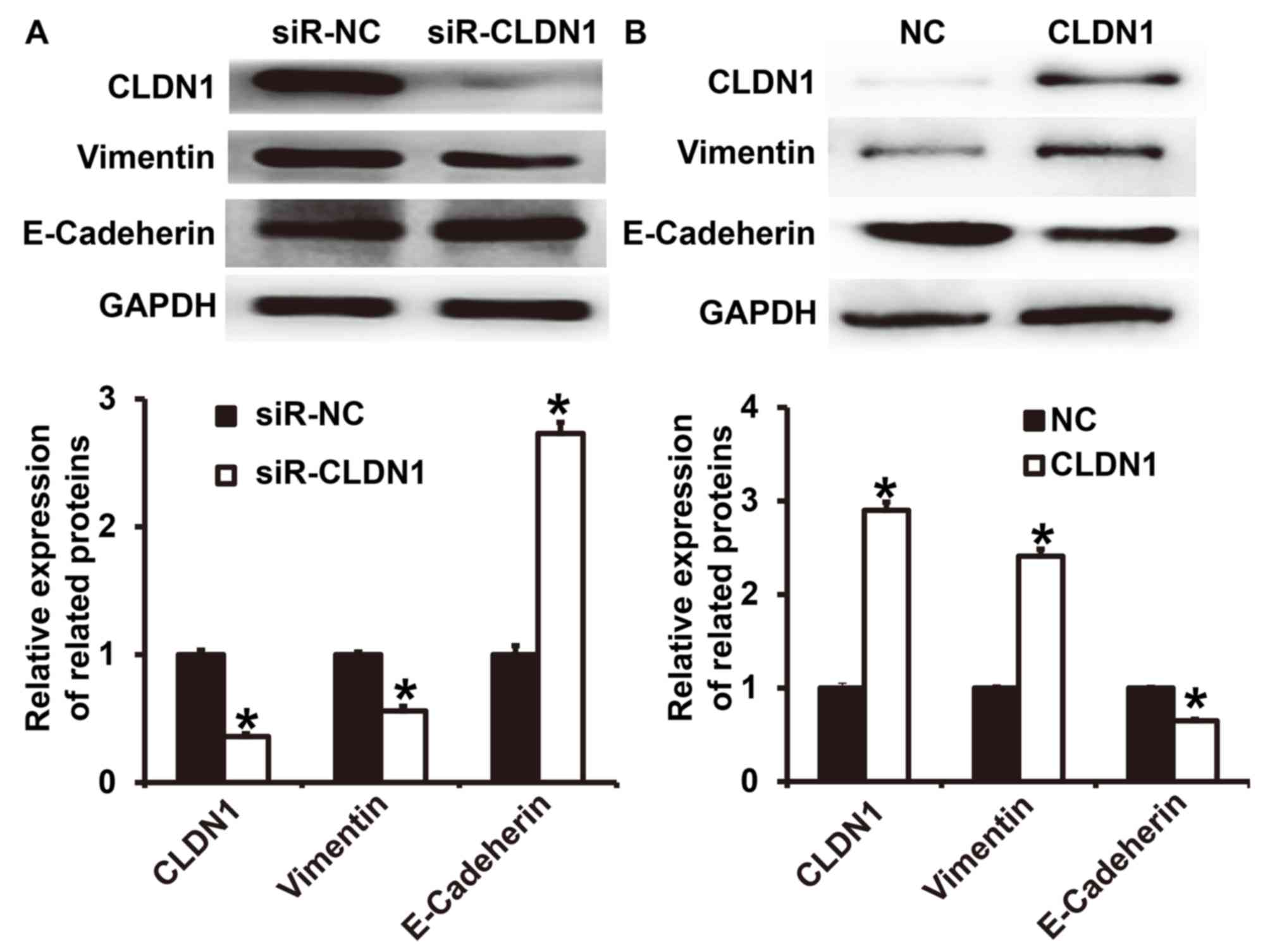
To determine the expression of the two EMT markers
E-cadherin and vimentin following transfection of NPC-TW01 cells
with either siR-CLDN1 or CLDN1, western blotting was performed. The
results demonstrated that the expression of vimentin following
transfection with siR-CLDN1 was significantly lower than that of
the siR-NC group (P<0.05; Fig.
4A), however, the expression of E-cadherin following
transfection with siR-CLDN1 was significantly higher than that of
the siR-NC group (P<0.05). By contrast, the expression of
vimentin following transfection with CLDN1 was significantly higher
than that of the NC group (P<0.05; Fig. 4B), however, the expression of
E-cadherin following transfection with CLDN1 was significantly
lower than that of the NC group (P<0.05). These results indicate
that the overexpression of CLDN1 promotes the EMT in NPC cells.
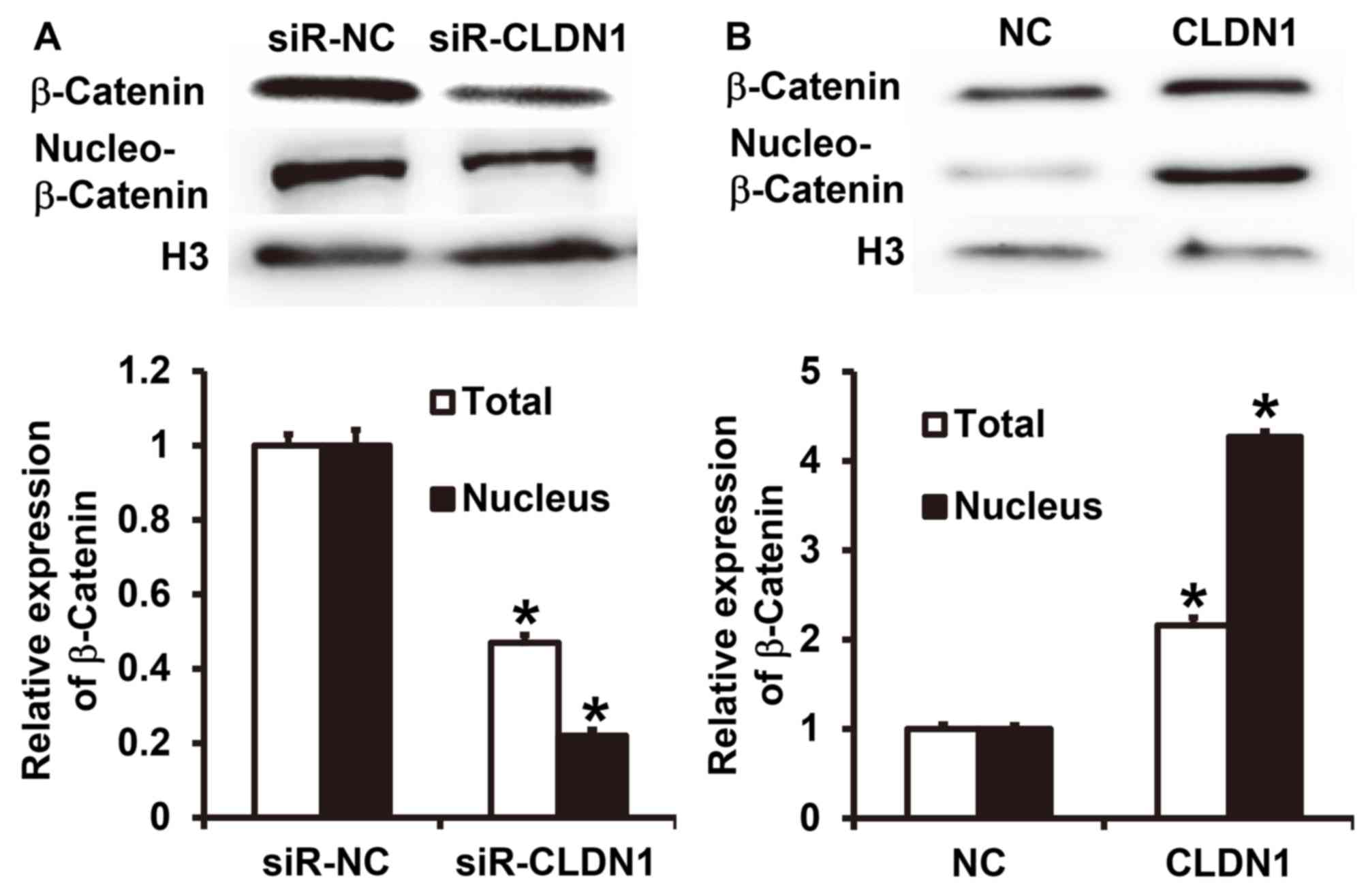
CLDN1 promotes β-catenin expression
and entry into the nucleus and induces the EMT via the β-catenin
signaling pathway
To measure the expression of β-catenin, western
blotting was performed. The results demonstrated that CLDN1
silencing significantly inhibited the total expression of β-catenin
and its nuclear translocation in NPC-TW01 cells (P<0.05;
Fig. 5A). By contrast,
overexpression of CLDN1 significantly enhanced the expression of
β-catenin and its nuclear translocation in NPC-TW01 cells
(P<0.05; Fig. 5B). These results
suggest that CLDN1 promotes β-catenin expression and entry into the
nucleus, and facilitates the EMT via the β-catenin signaling
pathway.
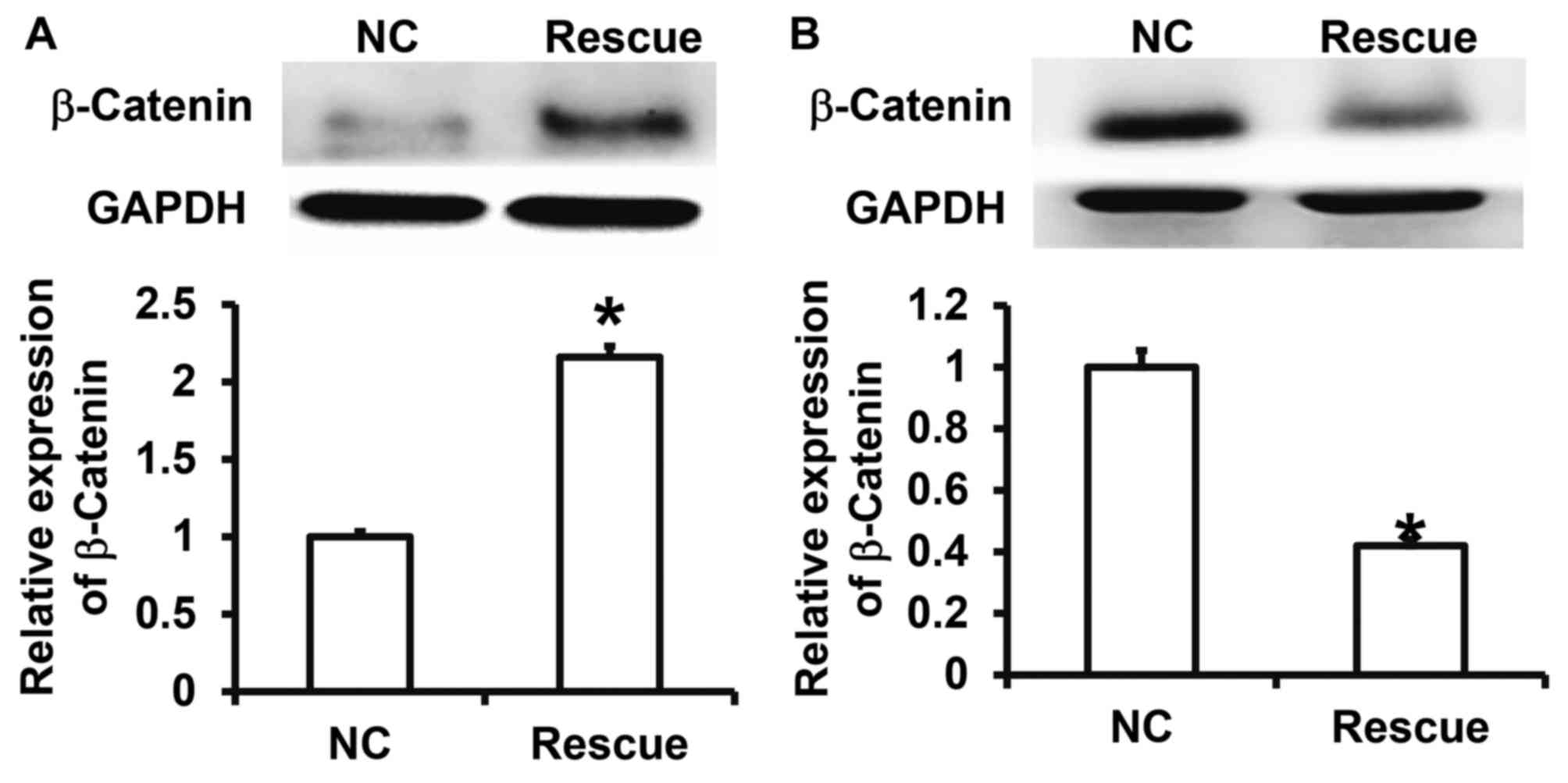
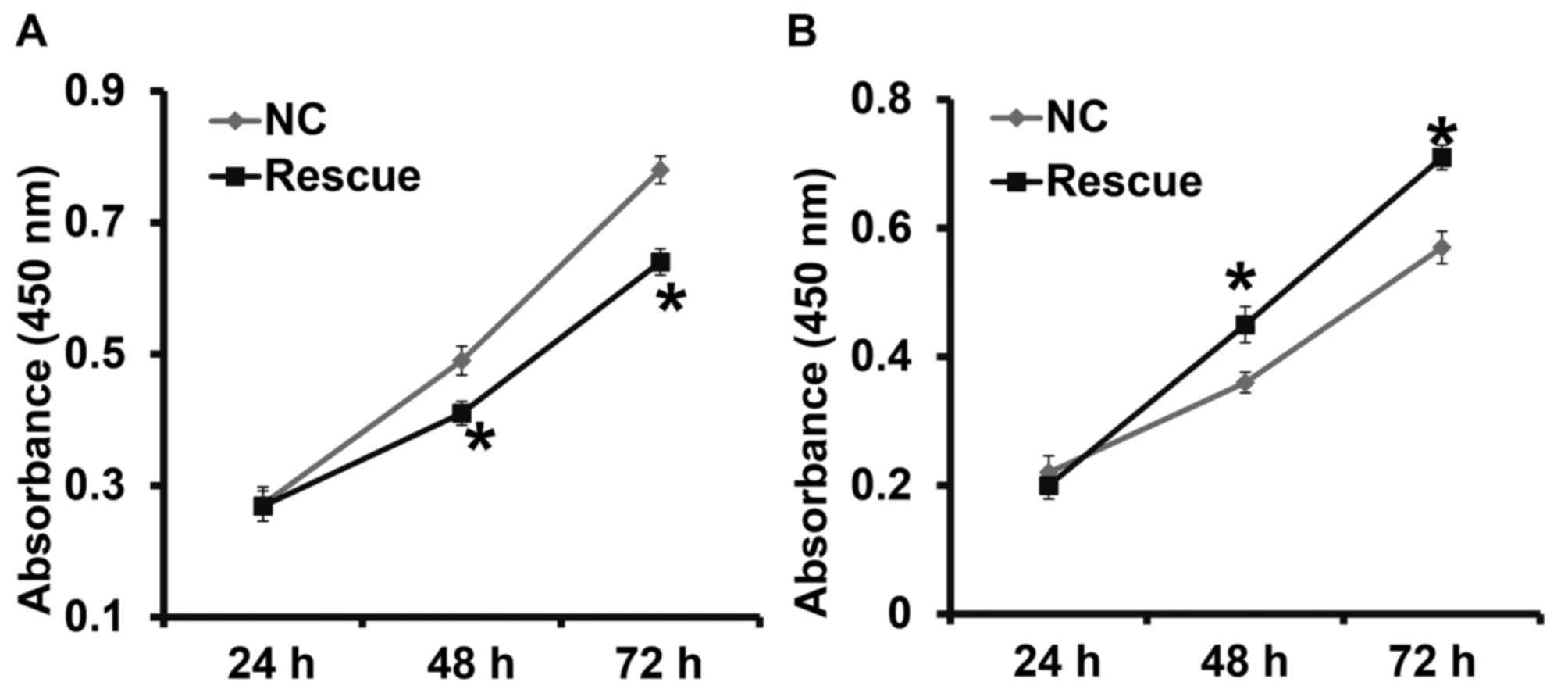
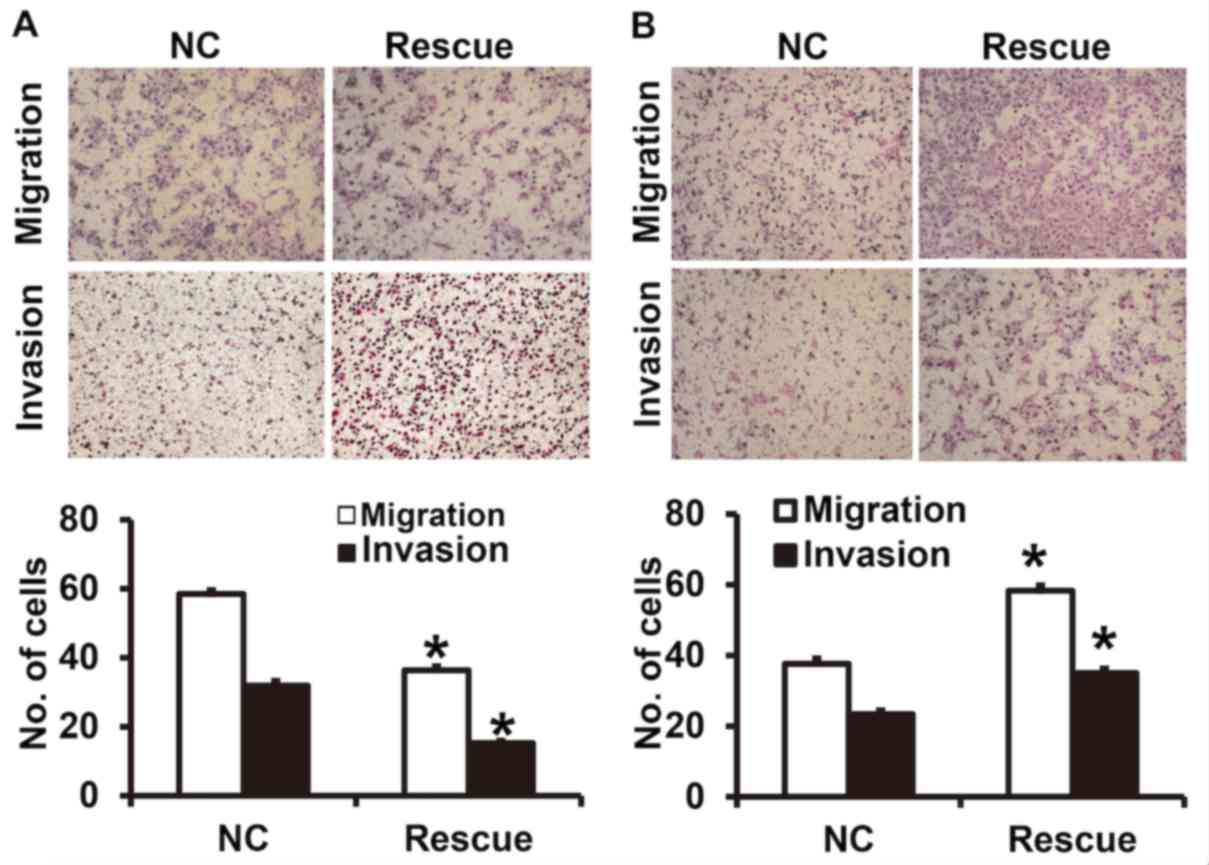
CLDN1 exerts its oncogene function via
the β-catenin signaling pathway
To confirm that CLDN1 induces its effects via the
β-catenin signaling pathway, NPC-TW01 cells exhibiting CLDN1
silencing and overexpression were transfected with β-catenin siRNA
and overexpression plasmids, respectively, to increase and decrease
β-catenin expression (Fig. 6). The
results demonstrated that the upregulation of β-catenin expression
significantly promoted the proliferation (Fig. 7), invasion and migration (Fig. 8) and the EMT (Fig. 9) of NPC-TW01 cells with silenced
CLDN1 expression. By contrast, downregulation of β-catenin
expression inhibited the proliferation, invasion and migration, and
EMT of NPC-TW01 cells exhibiting CLDN1 overexpression (Figs. 6–9).
These results indicate that CLDN1 exerts its oncogene function via
the β-catenin signaling pathway.
Discussion
CLDN1 is a tumor marker that exhibits abnormal
expression in various different tumors and participates in the
regulation of biological processes, including cell proliferation,
apoptosis, invasion and migration (29). Nakagawa et al (30) indicated that CLDN1 overexpression
promotes the invasion and migration of colon cancer cells and is
negatively correlated with patient prognosis. Fortier et al
(31) demonstrated that deletion of
the keratin 8 and 18 genes upregulates the expression of CLDN1,
thus stimulating the proliferation, invasion and migration of HepG2
cells. Jian et al (32)
indicated that the function of CLDN1 in promoting the invasion and
migration of osteosarcoma cells is closely associated with its
detachment from the cell membrane and entry into the nucleus,
suggesting that the intracellular location of CLDN1 is associated
with tumor migration and invasion. It has also been reported that
the expression of CLDN1 is elevated in gastric cancer tissues and
that it inhibits the anoikis of gastric cancer cells via the
β-catenin signaling pathway (33).
These studies suggest that CLDN1 is closely associated with tumor
invasion and metastasis and that the EMT is a key process in the
migration of epithelial tumor cells. Certain studies have
demonstrated that CLDN1 is closely associated with the EMT. For
example, CLDN1 promotes the EMT in hepatocytes via the c- Abelson
murine leukemia viral oncogene homolog
1-extracellular-signal-regulated kinase signaling pathway (34). In addition, the downregulation of
CLDN1 facilitates the EMT of rat hepatocytes induced by
transforming growth factor β (35).
The function of CLDN1 in the EMT may differ among different cells.
The results of the present study demonstrate that CLDN1 expression
is upregulated in NPC cell lines and promotes the proliferation,
the EMT, invasion and migration of NPC cells, which is consistent
with its effects in other tumors.
As a type of multifunctional protein, β-catenin is
widely distributed in different types of cells, including
epithelial cells, fibroblasts and osteoblasts, and promotes the
proliferation, differentiation and apoptosis of these cells
(35). It has been demonstrated that
the expression of β-catenin is upregulated in different types of
tumor and promotes the EMT in these tumor cells, indicating that it
is a key molecular target for inhibiting tumor metastasis. Oridonin
inhibits the EMT in pancreatic cancer cells by downregulating the
activity of the Wnt/β-catenin signaling pathway (36). Furthermore, the long non-coding RNA
UCA1 promotes the EMT in breast cancer cells by activating the
Wnt/β-catenin pathway (26) and Yi
et al (37) determined that
Wnt/β-catenin promotes the EMT and induces chemotherapy resistance
in glioma. Wnt/β-catenin is not only a key signaling pathway that
promotes the EMT, but also regulates tumor cell proliferation.
Santos et al (38) reported
that Sox9 enhances the proliferation of gastric cancer cells by
activating the Wnt/β-catenin pathway. Furthermore, Lu et al
(39) indicated that karyopherin β1
promotes the proliferation of glioma cells by activating the
Wnt/β-catenin pathway. The results of these studies suggest that
the Wnt/β-catenin signaling pathway induces important regulatory
effects on the EMT and tumor proliferation. The results of the
present study demonstrated that downregulating and overexpressing
CLDN1 in NPC cells upregulates and downregulates the expression and
nuclear entry of β-catenin, respectively. The downregulation of
β-catenin inhibits the cancer-promoting function of CLDN1,
suggesting that CLDN1 promotes the proliferation, EMT, invasion and
migration of NPC cells by activating the Wnt/β-catenin signaling
pathway.
In conclusion, the results of the present study
demonstrate that CLDN1 promotes the proliferation, EMT, invasion
and metastasis of NPC cells by activating the Wnt/β-catenin
signaling pathway. Therefore, CLDN1 is an oncogene that may be a
potential molecular therapeutic target for treating NPC.
Acknowledgements
The present study was supported by West China
Hospital, Sichuan University, China. The authors wish to thank
Professor Ping Li for his help.
Funding
No funding was received.
Availability of data and materials
The analyzed data sets generated during the study
are available from the corresponding author on reasonable
request.
Authors' contributions
The final version of the manuscript has been read
and approved by all authors, and each author believes that the
manuscript represents honest work. XW, JX, CZ and YJ collaborated
to design the study. XW, JX, CZ, CJZ, ZH, FW, YY and FF were
responsible for the experiments. XW, JX, CZ and YJ analyzed the
data. All authors collaborated to interpret results and develop the
manuscript.
Ethics approval and consent to
participate
All procedures performed in the current study were
approved by the Ethics Committee of Sichuan University. Written
informed consent was obtained from all patients or their
families.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Chen F, Chen C, Qu Y, Xiang H, Ai Q, Yang
F, Tan X, Zhou Y, Jiang G and Zhang Z: Selenium-binding protein 1
in head and neck cancer is low-expression and associates with the
prognosis of nasopharyngeal carcinoma. Medicine (Baltimore).
95:e45922016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Wang HZ, Cao CN, Luo JW, Yi JL, Huang XD,
Zhang SP, Wang K, Qu Y, Xiao JP, Li SY, et al: High-risk factors of
parotid lymph node metastasis in nasopharyngeal carcinoma: A
case-control study. Radiat Oncol. 11:1132016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Zhang M, Wei S, Su L, Lv W and Hong J:
Prognostic significance of pretreated serum lactate dehydrogenase
level in nasopharyngeal carcinoma among Chinese population: A
meta-analysis. Medicine (Baltimore). 95:e44942016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Sung WW, Chu YC, Chen PR, Liao MH and Lee
JW: Positive regulation of HIF-1A expression by EBV oncoprotein
LMP1 in nasopharyngeal carcinoma cells. Cancer Lett. 382:21–31.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Lin PJ, Twu CW, Liu YC, Lin TY, Wang WY
and Lin JC: Comparison the clinical outcomes with altered versus
conventional fractionated radiotherapy plus concurrent chemotherapy
for advanced nasopharyngeal carcinoma. Head Neck. Feb 1–2018.(Epub
ahead of print). View Article : Google Scholar
|
|
6
|
Zhang L, Huang Y, Hong S, Yang Y, Yu G,
Jia J, Peng P, Wu X, Lin Q, Xi X, et al: Gemcitabine plus cisplatin
versus fluorouracil plus cisplatin in recurrent or metastatic
nasopharyngeal carcinoma: A multicentre, randomised, open-label,
phase 3 trial. Lancet. 388:1883–1892. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Shi P, Zhong J, Hong J, Huang R, Wang K
and Chen Y: Automated Ki-67 quantification of immunohistochemical
staining image of human nasopharyngeal carcinoma xenografts. Sci
Rep. 6:321272016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wu VW, Leung WS, Wong KL, Chan YK, Law WL,
Leung WK and Yu YL: The impact of positron emission tomography on
primary tumour delineation and dosimetric outcome in intensity
modulated radiotherapy of early T-stage nasopharyngeal carcinoma.
Radiat Oncol. 11:1092016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zou ZW, Ma C, Medoro L, Chen L, Wang B,
Gupta R, Liu T, Yang XZ, Chen TT, Wang RZ, et al: LncRNA ANRIL is
up-regulated in nasopharyngeal carcinoma and promotes the cancer
progression via increasing proliferation, reprograming cell glucose
metabolism and inducing side-population stem-like cancer cells.
Oncotarget. 7:61741–61754. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Pearce SC, Al-Jawadi A, Kishida K, Yu S,
Hu M, Fritzky LF, Edelblum KL, Gao N and Ferraris RP: Marked
differences in tight junction composition and macromolecular
permeability among different intestinal cell types. BMC Biol.
16:192018. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Gonzalez-Mariscal L, Posadas-Torrentera Y,
Miranda J, Uc P, Ortega-Olvera JM and Hernandez S: Strategies that
target tight junctions for enhanced drug delivery. Curr Pharm Des.
22:1305–1311. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Salvador E, Burek M and Forster CY: Tight
junctions and the tumor microenvironment. Curr Pathobiol Rep.
4:135–145. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Chaouche-Mazouni S, Scherpereel A, Zaamoum
R, Mihalache A, Amir ZC, Lebaïli N, Delaire B and Gosset P: Claudin
3, 4 and 15 expression in solid tumors of lung adenocarcinoma
versus malignant pleural mesothelioma. Ann Diagn Pathol.
19:193–197. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kudinov AE, Deneka A, Nikonova AS, Beck
TN, Ahn YH, Liu X, Martinez CF, Schultz FA, Reynolds S, Yang DH, et
al: Musashi-2 (MSI2) supports TGF-β signaling and inhibits claudins
to promote non-small cell lung cancer (NSCLC) metastasis. Proc Natl
Acad Sci USA. 113:pp. 6955–6960. 2016; View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kwon MJ, Kim SH, Jeong HM, Jung HS, Kim
SS, Lee JE, Gye MC, Erkin OC, Koh SS, Choi YL, et al: Claudin-4
overexpression is associated with epigenetic derepression in
gastric carcinoma. Lab Invest. 91:1652–1667. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Okugawa T, Oshima T, Chen X, Hori K,
Tomita T, Fukui H, Watari J, Matsumoto T and Miwa H:
Down-regulation of claudin-3 is associated with proliferative
potential in early gastric cancers. Dig Dis Sci. 57:1562–1567.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Troy TC, Arabzadeh A, Yerlikaya S and
Turksen K: Claudin immunolocalization in neonatal mouse epithelial
tissues. Cell Tissue Res. 330:381–388. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Katoh M and Katoh M: CLDN23 gene,
frequently down-regulated in intestinal-type gastric cancer, is a
novel member of CLAUDIN gene family. Int J Mol Med. 11:683–689.
2003.PubMed/NCBI
|
|
19
|
Chen JJ, Zhong M, Dou TH, Wu ZY and Tang
WJ: rs17501976 polymorphism of CLDN1 gene is associated with
decreased risk of colorectal cancer in a Chinese population. Int J
Clin Exp Med. 8:1247–1252. 2015.PubMed/NCBI
|
|
20
|
Benczik M, Galamb A, Koiss R, Kovács A,
Járay B, Székely T, Szekerczés T, Schaff Z, Sobel G and Jeney C:
Claudin-1 as a biomarker of cervical cytology and histology. Pathol
Oncol Res. 22:179–188. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Hsueh C, Chang YS, Tseng NM, Liao CT,
Hsueh S, Chang JH, Wu IC and Chang KP: Expression pattern and
prognostic significance of claudins 1, 4 and 7 in nasopharyngeal
carcinoma. Hum Pathol. 41:944–950. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Cantelli G, Crosas-Molist E, Georgouli M
and Sanz-Moreno V: TGFB-induced transcription in cancer. Semin
Cancer Biol. 42:42–62. 2017. View Article : Google Scholar
|
|
23
|
Cardenas H, Zhao J, Vieth E, Nephew KP and
Matei D: EZH2 inhibition promotes epithelial-to-mesenchymal
transition in ovarian cancer cells. Oncotarget. 7:84453–84467.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Jin L, Yi J, Gao Y, Han S, He Z, Chen L
and Song H: MiR-630 inhibits invasion and metastasis in esophageal
squamous cell carcinoma. Acta Biochim Biophys Sin (Shanghai).
48:810–819. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Chung YH, Li SC, Kao YH, Luo HL, Cheng YT,
Lin PR, Tai MH and Chiang PH: MiR-30a-5p inhibits
epithelial-to-mesenchymal transition and upregulates expression of
tight junction protein claudin-5 in human upper tract urothelial
carcinoma cells. Int J Mol Sci. 18:E18262017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Xiao C, Wu CH and Hu HZ: LncRNA UCA1
promotes epithelial-mesenchymal transition (EMT) of breast cancer
cells via enhancing Wnt/beta-catenin signaling pathway. Eur Rev Med
Pharmacol Sci. 20:2819–2824. 2016.PubMed/NCBI
|
|
27
|
Cepollaro S, Della Bella E, de Biase D,
Visani M and Fini M: Evaluation of RNA from human trabecular bone
and identification of stable reference genes. J Cell Physiol.
233:4401–4407. 2017. View Article : Google Scholar
|
|
28
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
De Vicente JC, Fernandez-Valle A,
Vivanco-Allende B, Santamarta TR, Lequerica-Fernández P,
Hernández-Vallejo G and Allonca-Campa E: The prognostic role of
claudins −1 and −4 in oral squamous cell carcinoma. Anticancer Res.
35:2949–2959. 2015.PubMed/NCBI
|
|
30
|
Nakagawa S, Miyoshi N, Ishii H, Mimori K,
Tanaka F, Sekimoto M, Doki Y and Mori M: Expression of CLDN1 in
colorectal cancer: A novel marker for prognosis. Int J Oncol.
39:791–796. 2011.PubMed/NCBI
|
|
31
|
Fortier AM, Asselin E and Cadrin M:
Keratin 8 and 18 loss in epithelial cancer cells increases
collective cell migration and cisplatin sensitivity through
claudin1 up-regulation. J Biol Chem. 288:11555–11571. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Jian Y, Chen C, Li B and Tian X:
Delocalized Claudin-1 promotes metastasis of human osteosarcoma
cells. Biochem Biophys Res Commun. 466:356–361. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Huang J, Zhang L, He C, Qu Y, Li J, Zhang
J, Du T, Chen X, Yu Y, Liu B and Zhu Z: Claudin-1 enhances tumor
proliferation and metastasis by regulating cell anoikis in gastric
cancer. Oncotarget. 6:1652–1665. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Suh Y, Yoon CH, Kim RK, Lim EJ, Oh YS,
Hwang SG, An S, Yoon G, Gye MC, Yi JM, et al: Claudin-1 induces
epithelial-mesenchymal transition through activation of the
c-Abl-ERK signaling pathway in human liver cells. Oncogene.
32:4873–4882. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Kojima T, Takano K, Yamamoto T, Murata M,
Son S, Imamura M, Yamaguchi H, Osanai M, Chiba H, Himi T and Sawada
N: Transforming growth factor-beta induces epithelial to
mesenchymal transition by down-regulation of claudin-1 expression
and the fence function in adult rat hepatocytes. Liver Int.
28:534–545. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Liu QQ, Chen K, Ye Q, Jiang XH and Sun YW:
Oridonin inhibits pancreatic cancer cell migration and
epithelial-mesenchymal transition by suppressing Wnt/β-catenin
signaling pathway. Cancer Cell Int. 16:572016. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Yi GZ, Liu YW, Xiang W, Wang H, Chen ZY,
Xie SD and Qi ST: Akt and β-catenin contribute to TMZ resistance
and EMT of MGMT negative malignant glioma cell line. J Neurol Sci.
367:101–106. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Santos JC, Carrasco-Garcia E, Garcia-Puga
M, Aldaz P, Montes M, Fernandez-Reyes M, de Oliveira CC, Lawrie CH,
Araúzo-Bravo MJ, Ribeiro ML and Matheu A: SOX9 elevation acts with
canonical WNT signaling to drive gastric cancer progression. Cancer
Res. 76:6735–6746. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Lu T, Bao Z, Wang Y, Yang L, Lu B, Yan K,
Wang S, Wei H, Zhang Z and Cui G: Karyopherinβ1 regulates
proliferation of human glioma cells via Wnt/β-catenin pathway.
Biochem Biophys Res Commun. 478:1189–1197. 2016. View Article : Google Scholar : PubMed/NCBI
|