Introduction
Naringin is a dihydroflavonoid compound with
numerous biological and pharmacological roles, including
anti-inflammatory, anticancer, antiulcer, analgesic and
anti-hypertensive activities (1–4).
Naringin exhibits estrogenic activity and can decrease blood
cholesterol, reduce thrombus formation, and improve local
microcirculation and nutritional supply (5–9).
Therefore, naringin is widely used in clinical practice to prevent
cardiovascular and cerebrovascular diseases and as an adjunctive
treatment for bone fracture healing (10–13). A
Previous study has indicated that naringin can promote bone marrow
stem cell differentiation into mature bone. However, it inhibits
differentiation of MSCs into adipocytes (14). Furthermore, naringin can inhibit bone
loss and increase bone density (15).
Bone MSCs are adult multipotent cells derived from
the fetal mesoderm that exhibit a high proliferation, self-renewal
and differentiation potential (16–18).
MSCs participate in bone fracture repair (19). Directional migration of MSCs and
differentiation into mature bone serve essential roles in the local
healing of bone fractures (20).
Additionally, MSCs exhibit autocrine and paracrine functions and
may secrete a number of cytokines, growth factors, chemokines and
other regulators that participate in cellular functions (21). Therefore, differential secretion of
various cytokines by MSCs under the effects of suitable medication
can alter the microenvironment of MSCs in bone fractures as well as
the traits and function of other associated cell types, thereby
affecting bone fracture healing.
The Ras signaling pathway is a common molecular
signal transduction pathway in cells. Binding of the receptor
protein tyrosine kinase to signaling molecules forms a dimer and
results in autophosphorylation and activation of the receptor
protein tyrosine kinase and Ras (22). Activated Ras induces a protein kinase
phosphorylation cascade (23,24).
Previous studies have shown that activation of the Ras signaling
pathway is associated with the proliferation and migration of
numerous cell types, including cancer cells (18,25,26). A
previous report by the authors of the present study indicated that
activation of Ras-associated protein-1 promotes directional
migration of MSCs (16). Therefore,
the authors of the present study hypothesized that naringin may
potentiate the migration of MSCs via the activation of Ras and this
hypothesis was supported by the following experiments.
A scratch assay was used in the present study to
preliminarily detect the effect of naringin on MSC migration. To
confirm the hypothesis of the present study and further investigate
the molecular mechanism of naringin on MSCs, Quantibody®
arrays (RayBiotech, Inc., Norcross, GA, USA) were used to detect
the cytokines secreted by MSCs. It was hypothesized that naringin
may affect MSC cytokine secretion and that chemokine secretion by
naringin-induced MSCs is the mechanism by which this drug promotes
MSC migration. By analyzing the results of the
Quantibody® array, differentially synthesized chemokines
secreted by naringin-induced MSCs were identified. The Ras
signaling pathway activity was selected as it associated with MSC
migration. The aim of the present study was to provide novel
candidate agents for clinical orthopedics and theoretical basis for
the future improvement of adjunctive medication for bone fracture
healing.
Materials and methods
Reagents
MSCs (cat. no. HUXMA-01001) and Human MSC basal
culture medium (cat. no. HUXMA-9001c) were purchased from Cyagen
Biosciences, Inc. (Santa Clara, CA, USA). Naringin (Fig. 1) at a purity >95% (Sigma-Aldrich;
Merck KGaA, Darmstadt, Germany) was dissolved in physiological
saline, according to the manufacturer's protocol. Ras inhibitor
(Selleck Chemicals, Houston, TX, USA), a transferase inhibitor of
GTPase HRas and GTPase KRas, was used at a concentration of 5
µM.
Isolation and culture expansion of
human bone marrow MSCs
Adherent MSCs were trypsinized and passaged when the
cell confluence reached ~80%. Cells at passages 3–5 were used in
the following experiments. Typically, these cells exhibited the
capacity of differentiation into osteoblasts, adipoblasts and
chondrocytes.
Transwell migration assay
Cell migration was carried out with Transwell
chambers (pore size, 8-µm diameter; Costar; Corning Incorporated,
Corning, NY, USA). Human MSC basal culture medium was added into
the wells of a 24-well plate and subsequently serum-starved MSCs
(1×105) suspended in 100 µl of complete medium
containing 0.1% FCS were added into the upper chamber. Prior to the
addition of naringin, the Transwell plate, with MSCs in the upper
chamber and medium containing 0.1% FCS only in the lower chamber,
was first incubated at 37°C for 1 h. Following the addition of
naringin (1.0 or 10 µM) and the inhibitor (10 µM naringin + 5 µM
inhibitor), the plate was incubated at 37°C for 3 h, followed by
membrane fixation with 4% paraformaldehyde and staining with 0.1%
crystal violet at 25°C for 20 min. The membrane was subsequently
washed and the cells on the underside of the membrane were observed
under a light microscope (magnification, ×400). Numbers of cells
were counted in five to ten random fields of view for each
membrane.
Cell scratch assay
Cell migration was determined using a scratch assay.
The cells were cultivated to 90% confluence on 12-well plates at
25°C and treated with naringin (0.1, 1.0 and 10 µM), or controls
(without naringin). In further experiments, MSCs were pretreated
with 10 µM naringin. Following 6 h, the supernatant of the MSCs was
extracted to culture a new batch of MSCs and a scratch assay was
performed using a 1 ml micropipettor to form the scratch. The MSCs
were recorded at 0 and 6 h. Subsequently, cell scrapers (Corning
Incorporated) were utilized to scratch the confluent cells. The
extent of cellular growth was observed at 0 and 6 h. All
experiments were repeated three times.
Western blotting
Cells were harvested and lysed in
radioimmunoprecipitation assay buffer (Cyagen Biosciences, Inc.)
supplemented with protease inhibitors. Protein concentrations were
determined using a Novagen® Bicinchoninic Acid Protein
Assay kit (Merck KGaA). Following determination of the
concentration (5.534 µg/µl), protein samples (60 µg) were separated
by 10% SDS-PAGE and transferred to polyvinylidene difluoride
membranes. Membranes were blocked in 5% skim milk for 1 h at 25°C.
And incubated with antibodies against guanosine-5′-triphosphate
(GTP)-Ras (cat. no. 16117; 1:500; Active Ras Detection kit; Thermo
Fisher Scientific, Inc., Waltham, MA, USA), Ras (cat. no. ab52939;
1:5,000; Abcam, Cambridge, UK), and β-actin (cat. no. dc-130301;
1:2,000; Santa Cruz Biotechnology, Inc., Dallas, TX, USA),
overnight at 4°C. Following incubation with horseradish
peroxidase-conjugated goat anti-rabbit secondary antibodies (cat.
no. 31210; 1:5,000; Thermo Fisher Scientific, Inc.), immunoreactive
proteins were visualized using a Pierce™ Fast Western Blot kit, ECL
Substrate (cat. no. 35055; Thermo Fisher Scientific, Inc.).
Fluorescence intensity was measured using the ImageQuant™ LAS4000
system (GE Healthcare, Chicago, IL, USA; measurement parameter,
high resolution). Relative quantification of the bands was
performed using ImageJ software (version 5.0; National Institutes
of Health, Bethesda, MA, USA).
Antibody array
Soluble proteins in the medium of the MSCs were
measured using the Quantibody® Human Cytokine Array
G1000 (RayBiotech, Inc.; cat. no. AAH-CYT-G1000), according to the
manufacturer's protocol. Quantibody® arrays can detect
120 proteins. MSCs were plated 3 days prior to the experiment in
Human MSC basal culture medium and were 75–90% confluent. The
treatment group was stimulated with 10 µM naringin and compared
with the non-stimulated culture for 7 days. The cell lysis solution
was collected and filtered. Medium was also hybridized to the
arrays and subsequently used for normalization. A total of 10
technical and biological replicates were conducted and demonstrated
a very high positive correlation (Pearson correlation coefficient
>0.9; data not shown). Hybridization was conducted overnight at
4°C. All slides were scanned using the ImageQuant™ LAS4000 system
and analyzed using GenePix Pro 6.0 software (Molecular Devices,
LLC, Sunnyvale, CA, USA). The median score was used and averaged
across triplicates for each array. The results were subsequently
normalized using internal controls and the values for cytokines in
clear medium containing 10% FBS were subtracted.
Bioinformatics analysis
The analysis of cytokine-cytokine receptor
interaction pathways were performed using the Kyoto Encyclopedia of
Genes and Genomes database, (KEGG; http://www.kegg.jp/kegg/kegg2.html). The search
function of the website was utilized to search for the
cytokine-cytokine receptor interaction of CXCL5, CXCL6 and CCL20
for the species of Homo sapiens.
Statistical analysis
Analysis of variance and Student-Newman-Keuls test
(one-way analysis of variance) were used for data analysis. Data
were presented as the mean ± standard deviation (n=3 for each
experiment). P<0.05 was considered to indicate a statistically
significant difference. SPSS version 17.0 statistical software was
utilized to analyze data (SPSS, Inc., Chicago, IL, USA).
Results
Effects of naringin on the migration
of MSCs
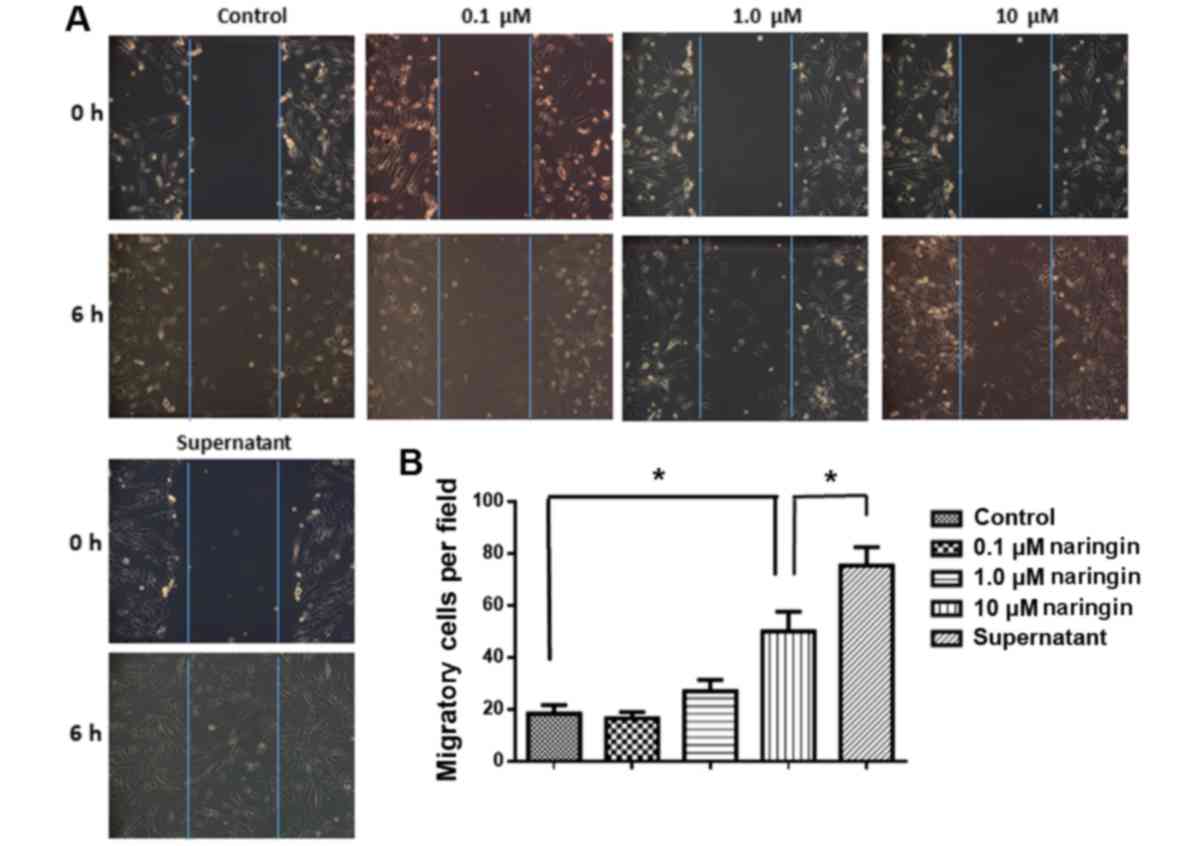
A scratch assay was performed to investigate the
effect of naringin on the migration of MSCs. The MSCs of all groups
moved towards the blank area to a certain extent after 6 h,
indicating a certain level of migration ability. Compared with the
control group, MSCs cultured with 10 µM naringin significantly
surpassed the baseline (P<0.05), indicating that the migration
ability of MSCs increased with naringin stimulation (Fig. 1).
MSCs exhibit strong autocrine and paracrine
functions (27) and, therefore, it
was hypothesized that MSCs could alter the composition of the
supernatant following treatment with naringin, making the
supernatant a possible chemoattractant. The supernatant may have
increased the migration of MSCs. Therefore, in another experiment,
MSCs were pretreated with 10 µM naringin. After 6 h, the
supernatant of the MSCs was extracted to culture a new batch of
MSCs and a scratch assay was performed. An increased number of MSCs
from the supernatant treatment crossed the baseline compared with
the group of MSCs directly cultured with naringin (P<0.05). The
above results indicated that treatment with the supernatant could
significantly increase the migration ability of MSCs (Fig. 1). The present study also aimed to
determine whether MSCs could synthesize factors that promote
cellular migration following treatment with naringin. An antibody
array experiment was subsequently conducted to identify these
factors.
Effects of naringin on cellular
chemokine synthesis by MSCs
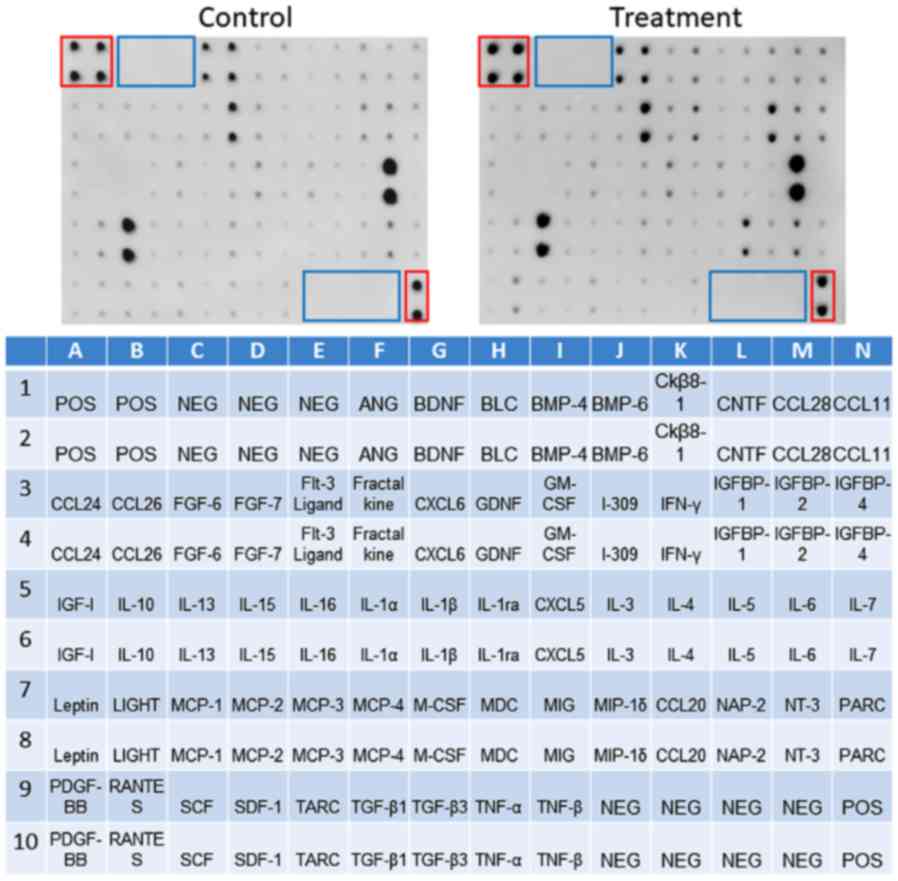
To determine the influence of stimulation with
naringin on human MSCs at the protein level, Quantibody®
arrays were performed. The treatment group was stimulated with 10
µM naringin and compared with the non-stimulated culture for 7 days
(Figs. 2 and 3).
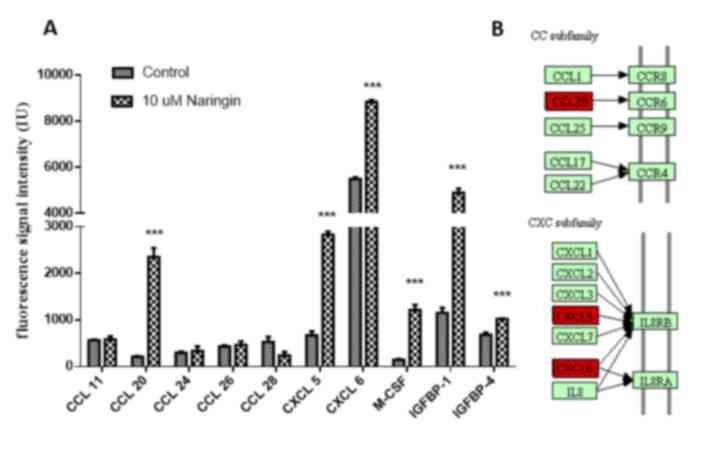 | Figure 3.Cytokine-cytokine receptor interaction
of cytokines synthesized by MSCs. (A) Following treatment with
naringin, an increased cytokine synthesis of CXCL-6, CCL-20, M-CSF,
IGFBP-1, IGFBP-4 and CXCL5 was observed. (B) Cytokine-cytokine
receptor interaction between the synthesized cytokines. The
differentially secreted cytokines are presented in their
interaction pathway including IL-8RB, IL-8RA and CCR6 according to
the cytokine-cytokine receptor interaction pathway (04060 hsa) in
the Kyoto Encyclopedia of Genes and Genomes database (red:
significantly upregulated). ***P<0.001. CXCL, C-X-C motif
chemokine; CCL, C-C motif chemokine; IL-8RB, C-X-C chemokine
receptor type 2; IL-8RA, C-X-C chemokine receptor type 1; CCR, C-C
chemokine receptor; IL8, interleukin-8; MSC, mesenchymal stromal
cells. |
Statistical analysis demonstrated identification of
seven cytokines that were differentially synthesized between the
two groups. Following treatment with naringin, an increased
cytokine synthesis (relative level, >1.5-fold change) of C-X-C
motif chemokine (CXCL)6, C-C motif chemokine (CCL)20, macrophage
colony-stimulating factor (M-CSF), insulin-like growth
factor-binding protein (IGFBP)-1, IGFBP-4 and CXCL5 was observed
(relative level, >1.5 fold change). Decreased cytokine synthesis
(relative level, <0.5-fold change) was observed for CCL28
(Fig. 3A).
Based on the search criteria for differential
secretion used in the present study, Statistical analysis resulted
in identification of three cytokines (relative level, >1.5-fold
control). Increased secretion of cytokines CXCL5, CXCL6 and CCL20
followed induction with naringin. The differentially secreted
cytokines are presented in their interaction pathways, including
IL-8RB, IL-8RA and CCR6 according to the cytokine-cytokine receptor
interaction pathway (hsa04060; http://www.kegg.jp/kegg-bin/show_pathway?hsa04060+6364)
in the Kyoto Encyclopedia of Genes and Genomes database (Fig. 3B).
Naringin induces MSCs to activate the
Ras signaling pathway and enhance MSC migration
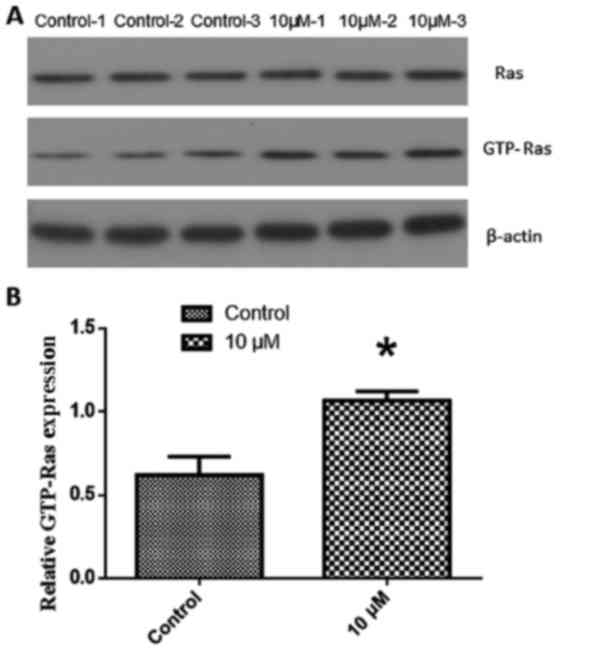
To verify the hypothesis that the Ras signaling
pathway of MSCs could be activated by treatment with naringin,
western blot analysis was performed. Compared with the control
group, the expression of GTP-Ras in MSCs increased significantly
following treatment with 10 µM of naringin (P<0.05). These
results indicated that naringin could activate the Ras signaling
pathway in MSCs (Fig. 4).
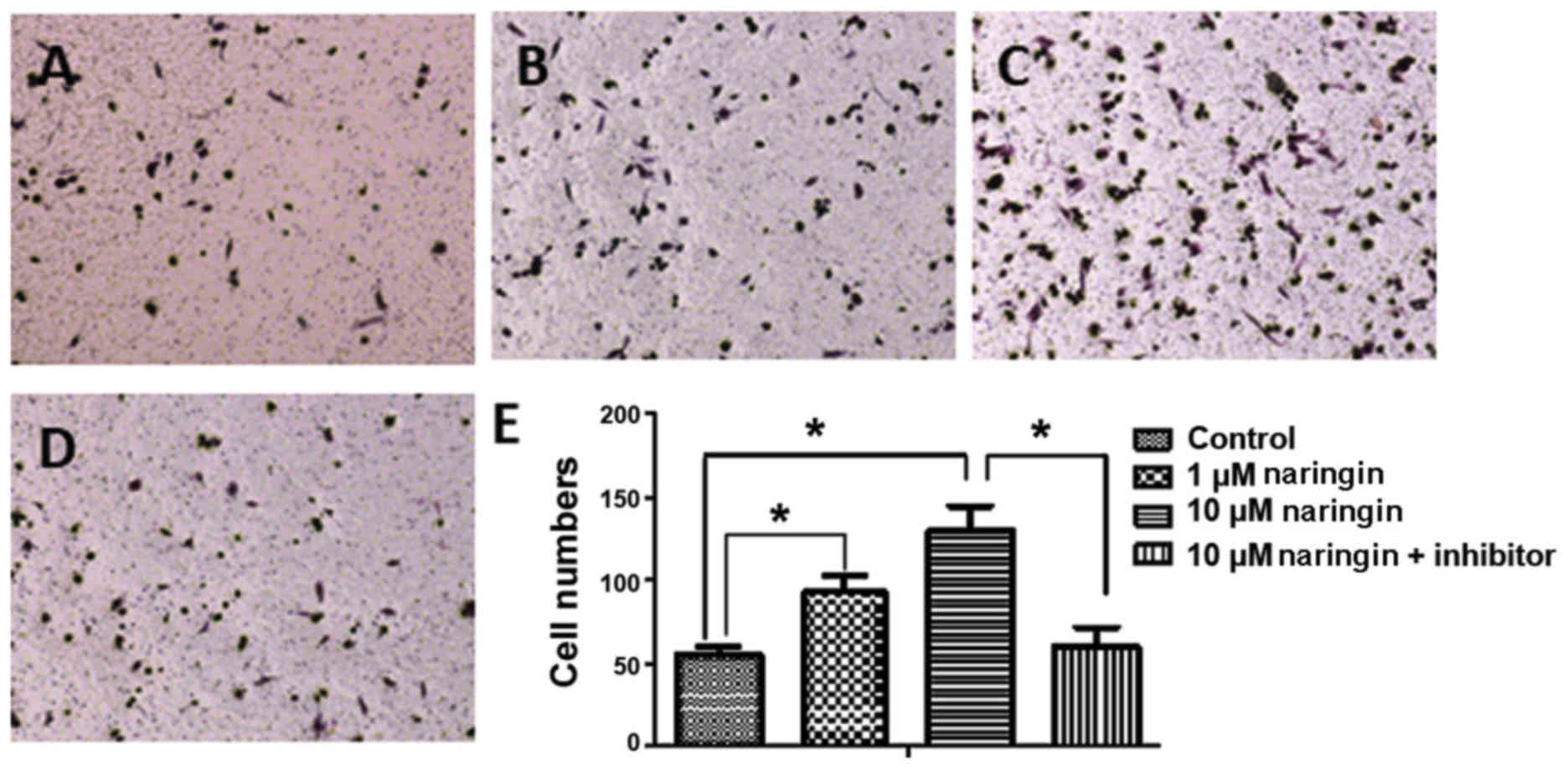
To investigate whether activation of the Ras
signaling pathway could increase the migration of MSCs, a Transwell
migration assay was performed. Compared with the control group, the
number of MSCs migrating towards the other side of the membrane
increased significantly following treatment with 1 or 10 µM
naringin. However, following treatment with the Ras inhibitor, cell
migration was significantly reduced compared with the group treated
with 10 µM naringin. This result suggested that activation and
inhibition of the Ras signaling pathway significantly enhanced and
suppressed the migration of MSCs, respectively (Fig. 5).
Discussion
In the first scratch assay, it was observed that the
migration of MSCs treated with an appropriate concentration of
naringin was enhanced compared with the control group, suggesting
that naringin promoted the migration of MSCs. In another
experiment, the supernatant of MSCs induced with naringin was used
to culture a separate batch of MSCs. The migration of MSCs treated
with the supernatant was enhanced compared with that of the MSCs of
the control group and the group directly treated with an identical
concentration of naringin. The above results suggested that the
supernatant may contain certain cytokines that promote MSC
migration. Therefore, it was hypothesized that, following treatment
with naringin, MSCs may synthesize certain cytokines to further
enhance their mobility. However, this hypothesis requires further
verification, since the present experiment did not control for all
the potentially confounding variables.
To confirm whether the synthesis of specific
cytokines by MSCs was enhanced following treatment with naringin,
and to identify the cytokines responsible for enhancing the
mobility of MSCs, a Quantibody® array was performed. The
results demonstrated that the synthesis of CXCL5, CXCL6 and CCL20
increased significantly. The CCL subfamily contains >20 members
with two neighboring cysteine residues in the N-terminal domain;
these predominantly stimulate mononuclear cells and certain
subpopulations of T cells (28).
CCL20, also known as macrophage inflammatory protein-3, exerts its
function by binding to its receptor, C-C chemokine receptor type 6
(29). CCL20 primarily stimulates
dendritic cells and numerous types of immune cells, and may also
regulate bone mass accrual (30).
The characteristic molecular structure of the C-X-C motif subfamily
consists of an amino acid between two cysteine residues at the
N-terminus (31). The C-X-C motif
subfamily proteins are produced by activated mononuclear cells,
endothelial cells, fibroblasts and megakaryocytes, and the majority
of members can stimulate neutrophils, T cells and B cells (32–34).
CXCL5, also known as epithelial-derived neutrophil-activating
peptide 78, is primarily produced by cells upon stimulation by
interleukin-1 or tumor necrosis factor-α (35). Studies have shown that CXCL5
secretion affects differentiation and migration of MSCs (36). CXCL6, also known as granulocyte
chemotactic protein 2, exerts its stimulatory effect on white blood
cells and promotes angiogenesis and antibacterial activity by
interacting with chemokine receptors C-X-C chemokine receptor type
(CXCR)1 and CXCR2 (37). CXCL6 also
induces angiogenesis and promotes tumor growth (38). The above studies suggested that
differentially secreted chemokines in naringin-induced MSCs may
affect MSCs and other associated cell types, including immune
cells, in the bone fracture microenvironment. Naringin may promote
MSC migration by stimulating chemokine synthesis and secretion.
Increased concentrations of chemokines in the bone fracture
microenvironment may in turn promote the directional migration of
MSCs and associated cells. Further experiments are required to
elucidate the specific roles of individual chemokines during MSC
migration. Subsequent studies should investigate the role of
naringin in the promotion of MSC osteogenic differentiation.
The present study also aimed to identify the
signaling pathways that regulate MSC migration using the
Quantibody® assays. The differentially secreted
cytokines (insulin-like growth factor-binding protein and M-CSF)
were involved in the transmission of the Ras signaling pathway
(39,40). This indicated that the Ras signaling
pathway may regulate stem cell migration. Therefore, the authors of
the present study hypothesized that naringin may facilitate MSC
migration by activating the Ras signaling pathway. To test this
hypothesis, western blotting experiment were conducted to detect
expression of relevant proteins in naringin-induced MSCs. The
results indicated that following treatment with naringin GTP-Ras
content in MSCs significantly increased. The expression level of
GTP-Ras represents the activation of Ras (24) and, therefore, the results of the
present study indicated that naringin can be considered a
stimulator of the Ras signaling pathway in MSCs. In subsequent
experiments, MSCs were treated with a Ras inhibitor and the
Transwell assay revealed that MSC migration was significantly
decreased compared with the group treated with 10 µM naringin. The
above results indicated that naringin activated the Ras signaling
pathway, thereby promoting MSC migration.
In the present study, a preliminary analysis of the
Quantibody® array results was performed, and these
results require further verification. Several of the results merit
further study. For example, CCL-5 also serves a role in the nuclear
factor-κB signaling pathway (41).
It can be hypothesized that naringin may affect this signaling
pathway. However, whether this signaling pathway is activated or
inhibited requires experimental confirmation. The nuclear factor-κB
signaling pathway serves an important role in MSC differentiation
into mature bone (42) and,
therefore, its activity under the effects of naringin should be
further investigated to advance the understanding of the molecular
mechanism by which naringin affects MSCs.
Acknowledgements
Not applicable.
Funding
The present study was supported by a grant from the
Hangzhou Medical and Health Science and Technology Plan Project
(grant no. 2018B012) and Zhejiang Chinese Medical University
Scientific Research Fund Project (grant no. 2018ZY26).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
All authors have contributed significantly and are
in agreement with the content of the manuscript. FL designed the
experiments and wrote the manuscript. GH designed the experiments
was a major contributor in writing the manuscript. YZ performed
cell experiments and performed data analysis. All authors read and
approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Chtourou Y, Aouey B, Aroui S, Kebieche M
and Fetoui H: Anti-apoptotic and anti-inflammatory effects of
naringin on cisplatin-induced renal injury in the rat. Chem Biol
Interact. 243:1–9. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Yoshinaga A, Kajiya N, Oishi K, Kamada Y,
Ikeda A, Chigwechokha PK, Kibe T, Kishida M, Kishida S, Komatsu M
and Shiozaki K: NEU3 inhibitory effect of naringin suppresses
cancer cell growth by attenuation of EGFR signaling through GM3
ganglioside accumulation. Eur J Pharmacol. 782:21–29. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kandhare AD, Ghosh P and Bodhankar SL:
Naringin, a flavanone glycoside, promotes angiogenesis and inhibits
endothelial apoptosis through modulation of inflammatory and growth
factor expression in diabetic foot ulcer in rats. Chem Biol
Interact. 219:101–112. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ikemura M, Sasaki Y, Giddings JC and
Yamamoto J: Preventive effects of hesperidin, glucosyl hesperidin
and naringin on hypertension and cerebral thrombosis in
stroke-prone spontaneously hypertensive rats. Phytother Res.
26:1272–1277. 2012. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
Xiao Y, Li LL, Wang YY, Guo JJ, Xu WP,
Wang YY and Wang Y: Naringin administration inhibits platelet
aggregation and release by reducing blood cholesterol levels and
the cytosolic free calcium concentration in hyperlipidemic rabbits.
Exp Ther Med. 8:968–972. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Mao Z, Gan C, Zhu J, Ma N, Wu L, Wang L
and Wang X: Anti-atherosclerotic activities of flavonoids from the
flowers of Helichrysum arenarium L. MOENCH through the pathway of
anti-inflammation. Bioorg Med Chem Lett. 27:2812–2817. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zamanian M, Hajizadeh M, Shamsizadeh A,
Moemenzadeh M, Amirteimouri M, Elshiekh M and Allahtavakoli M:
Effects of naringin on physical fatigue and serum MMP-9
concentration in female rats. Pharm Biol. 55:423–427. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wang S, Bao YR, Li TJ, Yu T, Chang X, Yang
GL and Meng XS: Mechanism of fructus aurantii flavonoids promoting
gastrointestinal motility: From organic and inorganic endogenous
substances combination point of view. Pharmacogn Mag. 13:372–377.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Guo D, Wang J, Wang X, Luo H, Zhang H, Cao
D and Chen L: Double directional adjusting estrogenic effect of
naringin from Rhizoma drynariae (Gusuibu). J Ethnopharmacol.
138:451–457. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Toth PP, Patti AM, Nikolic D, Giglio RV,
Castellino G, Biancucci T, Geraci F, David S, Montalto G, Rizvi A
and Rizzo M: Bergamot reduces plasma lipids, atherogenic small
dense LDL and subclinical atherosclerosis in subjects with moderate
hypercholesterolemia: A 6 months prospective study. Front
Pharmacol. 6:2992016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Baskaran G, Salvamani S, Ahmad SA,
Shaharuddin NA, Pattiram PD and Shukor MY: HMG-CoA reductase
inhibitory activity and phytocomponent investigation of Basella
alba leaf extract as a treatment for hypercholesterolemia. Drug Des
Devel Ther. 9:509–517. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Nyane NA, Tlaila TB, Malefane TG, Ndwandwe
DE and Owira PMO: Metformin-like antidiabetic, cardio-protective
and non-glycemic effects of naringenin: Molecular and
pharmacological insights. Eur J Pharmacol. 803:103–111. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Song N, Zhao Z, Ma X, Sun X, Ma J, Li F,
Sun L and Lv J: Naringin promotes fracture healing through
stimulation of angiogenesis by regulating the VEGF/VEGFR-2
signaling pathway in osteoporotic rats. Chem Biol Interact.
261:11–17. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Fan J, Li J and Fan Q: Naringin promotes
differentiation of bone marrow stem cells into osteoblasts by
upregulating the expression levels of microRNA-20a and
downregulating the expression levels of PPARγ. Mol Med Rep.
12:4759–4765. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Li N, Jiang Y, Wooley PH, Xu Z and Yang
SY: Naringin promotes osteoblast differentiation and effectively
reverses ovariectomy-associated osteoporosis. J Orthop Sci.
18:478–485. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Lin F, Xue D, Xie T and Pan Z: HMGB1
promotes cellular chemokine synthesis and potentiates mesenchymal
stromal cell migration via Rap1 activation. Mol Med Rep.
14:1283–1289. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lin F, Zhang W, Xue D, Zhu T, Li J, Chen
E, Yao X and Pan Z: Signaling pathways involved in the effects of
HMGB1 on mesenchymal stem cell migration and osteoblastic
differentiation. Int J Mol Med. 37:789–797. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Feng L, Xue D, Chen E, Zhang W, Gao X, Yu
J, Feng Y and Pan Z: HMGB1 promotes the secretion of multiple
cytokines and potentiates the osteogenic differentiation of
mesenchymal stem cells through the Ras/MAPK signaling pathway. Exp
Ther Med. 12:3941–3947. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhang H, Kot A, Lay YE, Fierro FA, Chen H,
Lane NE and Yao W: Acceleration of fracture healing by
overexpression of basic fibroblast growth factor in the mesenchymal
stromal cells. Stem Cells Transl Med. 6:1880–1893. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Li X, He L, Yue Q, Lu J, Kang N, Xu X,
Wang H and Zhang H: MiR-9-5p promotes MSC migration by activating
β-catenin signaling pathway. Am J Physiol Cell Physiol.
313:C80–C93. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Luan XY, Zhang GB, Hu YM, Yu GH, Wang MY,
Duan QY, Duan X and Zhang XG: Effects of MSC on cell cycle,
phenotype and cytokine secretion of T cells activated by PHA. Xi
Bao Yu Fen Zi Mian Yi Xue Za Zhi. 23:402–405. 2007.(In Chinese).
PubMed/NCBI
|
|
22
|
Rocks O, Peyker A and Bastiaens PI:
Spatio-temporal segregation of Ras signals: One ship, three
anchors, many harbors. Curr Opin Cell Biol. 18:351–357. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Thatcher JD: The Ras-MAPK signal
transduction pathway. Sci Signal. 3:tr1. 2010. View Article : Google Scholar
|
|
24
|
Khosravi-Far R and Der CJ: The Ras signal
transduction pathway. Cancer Metastasis Rev. 13:67–89. 1994.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhou J, Zhang S, Chen X, Zheng X, Yao Y,
Lu G and Zhou J: Palbociclib, a selective CDK4/6 inhibitor,
enhances the effect of selumetinib in RAS-driven non-small cell
lung cancer. Cancer Lett. 408:130–137. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Li X, Li D, Wikstrom JD, Pivarcsi A,
Sonkoly E, Ståhle M and Landén NX: MicroRNA-132 promotes fibroblast
migration via regulating RAS p21 protein activator 1 in skin wound
healing. Sci Rep. 7:77972017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Gao D, Xie J, Zhang J, Feng C, Yao B, Ma
K, Li J, Wu X, Huang S and Fu X: MSC attenuate diabetes-induced
functional impairment in adipocytes via secretion of insulin-like
growth factor-1. Biochem Biophys Res Commun. 452:99–105. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Van Coillie E, VanDamme J and Opdenakker
G: The MCP/eotaxin subfamily of CC chemokines. Cytokine Growth
Factor Rev. 10:61–86. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Baba M, Imai T, Nishimura M, Kakizaki M,
Takagi S, Hieshima K, Nomiyama H and Yoshie O: Identification of
CCR6, the specific receptor for a novel lymphocyte-directed CC
chemokine LARC. J Biol Chem. 272:14893–14898. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Doucet M, Jayaraman S, Swenson E, Tusing
B, Weber KL and Kominsky SL: CCL20/CCR6 signaling regulates bone
mass accrual in mice. J Bone Miner Res. 31:1381–1390. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Hieshima K, Imai T, Opdenakker G, Van
Damme J, Kusuda J, Tei H, Sakaki Y, Takatsuki K, Miura R, Yoshie O
and Nomiyama H: Molecular cloning of a novel human CC chemokine
liver and activation-regulated chemokine (LARC) expressed in liver.
Chemotactic activity for lymphocytes and gene localization on
chromosome 2. J Biol Chem. 272:5846–5853. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Kim GW, Han MS, Park HR, Lee EJ, Jung YK,
Usmani SE, Ulici V, Han SW and Beier F: CXC chemokine ligand 12a
enhances chondrocyte proliferation and maturation during
endochondral bone formation. Osteoarthritis Cartilage. 23:966–974.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Le Y, Zhou Y, Iribarren P and Wang J:
Chemokines and chemokine receptors: Their manifold roles in
homeostasis and disease. Cell Mol Immunol. 1:95–104.
2004.PubMed/NCBI
|
|
34
|
Graham GJ and Locati M: Regulation of the
immune and inflammatory responses by the ‘atypical’ chemokine
receptor D6. J Pathol. 229:168–175. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Chang MS, McNinch J, Basu R and Simonet S:
Cloning and characterization of the human neutrophil-activating
peptide (ENA-78) gene. J Biol Chem. 269:25277–25282.
1994.PubMed/NCBI
|
|
36
|
Nedeau AE, Bauer RJ, Gallagher K, Chen H,
Liu ZJ and Velazquez OC: A CXCL5- and bFGF-dependent effect of
PDGF-B-activated fibroblasts in promoting trafficking and
differentiation of bone marrow-derived mesenchymal stem cells. Exp
Cell Res. 314:2176–2186. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Catusse J, Struyf S, Wuyts A, Weyler M,
Loos T, Gijsbers K, Gouwy M, Proost P and Van Damme J: Rabbit
neutrophil chemotactic protein (NCP) activates both CXCR1 and CXCR2
and is the functional homologue for human CXCL6. Biochem Pharmacol.
68:1947–1955. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Verbeke H, Struyf S, Berghmans N, Van
Coillie E, Opdenakker G, Uyttenhove C, Van Snick J and Van Damme J:
Isotypic neutralizing antibodies against mouse GCP-2/CXCL6 inhibit
melanoma growth and metastasis. Cancer Lett. 302:54–62. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Kuemmerle JF and Zhou H: Insulin-like
growth factor-binding protein-5 (IGFBP-5) stimulates growth and
IGF-I secretion in human intestinal smooth muscle by Ras-dependent
activation of p38 MAP kinase and Erk1/2 pathways. J Biol Chem.
277:20563–20571. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Bradley EW, Ruan MM, Vrable A and Oursler
MJ: Pathway crosstalk between Ras/Raf and PI3K in promotion of
M-CSF-induced MEK/ERK-mediated osteoclast survival. J Cell Biochem.
104:1439–1451. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Pham TH, Kim MS, Le MQ, Song YS, Bak Y,
Ryu HW, Oh SR and Yoon DY: Fargesin exerts anti-inflammatory
effects in THP-1 monocytes by suppressing PKC-dependent AP-1 and
NF-kB signaling. Phytomedicine. 24:96–103. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Wang YJ, Zhang HQ, Han HL, Zou YY, Gao QL
and Yang GT: Taxifolin enhances osteogenic differentiation of human
bone marrow mesenchymal stem cells partially via NF-κB pathway.
Biochem Biophys Res Commun. 490:36–43. 2017. View Article : Google Scholar : PubMed/NCBI
|