Introduction
Aneurysmal subarachnoid hemorrhage (SAH) is a
neurological disease associated with high rates of mortality; only
~50% patients survive this clinical condition (1). A large proportion of the survivors may
also suffer from neurological disorders, including cognitive
impairment and dementia later in life (2). Extensive investigations have focused on
managing early post-morbidity and improving long-term care
following this type of cerebral stroke. For example, brain imaging
studies have suggested that SAH causes white matter damage, and
monitoring the extent of this lesion may be of prognostic value
(3,4). The factors and mechanism underlying
SAH-induced neurological deficits remain to be fully elucidated;
mechanical and ischemic stress, and neuroinflammation are expected
to occur during and following subarachnoid bleeding (5).
The transactive response DNA-binding protein of 43
(TDP-43) has an important physiological role in the nervous system.
TDP-43 regulates mRNA transport and stability, and is involved in
microRNA biosynthesis, apoptosis, cell division and the regulation
of neuronal plasticity. TDP-43 is mainly localized in the nucleus
under normal conditions. By interacting with RNAs, TDP-43 may
regulate the alternative splicing of genes associated with the
development of neurodegenerative diseases (6). Alterations in the expression of TDP-43
have been associated with Alzheimer's disease, Lewy body-related
diseases, Pick's disease and Huntington's disease (7,8). TDP-43
may exhibit neurotoxicity via phosphorylation, ubiquitination,
insolubility and the formation of ectopic cytoplasmic inclusions,
which has been defined as TDP-43 proteinopathy (6). Aggregated TDP-43 may also occur in
axons, causing neuronal pathology in amyotrophic lateral sclerosis
and other conditions (9–11). Similarly, the accumulation of TDP-43
in neurons may inhibit dendritic growth, and impair dendritic
complexity and neurocircuitry communication (12).
The present study aimed to investigate whether
alterations in the protein expression of TDP-43 occur in human
subjects suffering from SAH via western blot analysis of
cerebrospinal fluid (CSF). Marked elevations in the expression
levels of TDP-43 were observed within CSF samples from patients
with SAH relative to healthy controls in the present study. To
determine the potential cellular origin of the SAH-induced
upregulation of TDP-43, a rat model of SAH was established, which
was also used to investigate SAH-induced behavioral deficits
Additionally, alterations in the expression of TDP-43 within the
brains of experimental animals were examined relative to sham
controls.
Materials and methods
Ethics statement
The present study was performed in compliance with
Chinese legislations and guidelines of the National Institutes of
Health involving human subjects and for the use and care for
experimental animals. All animal experiments were approved by the
Ethics Committee of Central South University Xiangya School of
Medicine (Changsha, China). Clinical information and CSF samples
from patients were obtained with written informed consent prior to
enrollment in the study. The experiments involving human subjects
were approved by the Ethical Committee of The Third Xiangya
Hospital of Central South University (Changsha, China).
CSF samples from patients with SAH and
controls
All SAH subjects (4 males, each with aneurysmal SAH;
age range, 57–69 years) and controls (3 males, each with migraines;
age range, 30–43 years) were inpatients at the Department of
Neurology of The Third Xiangya Hospital of Central South
University. Patients with ruptured aneurysms did not have any
underlying disease prior to the incident and were treated with
interventional therapy. CSF was obtained from March 2017 to April
2017 by lumbar puncture at 48 h post-SAH; a 10-ml sample was
collected from each patient (n=4). Control CSF samples (n=3) were
obtained from patients clinically diagnosed with a migraine. CSF
samples were centrifuged at 3,200 × g for 20 min at 4°C. Following
this, 8 ml supernatant was placed in an ultrafiltration tube
(Amicon® Ultra-15 10K Centrifugal Filter Devices;
Amicon; EMD Millipore, Billerica, MA, USA) and centrifuged at a
speed of 4,000 × g for 20 min at 4°C, leading to a 4-fold
concentration increase.
Animals and grouping
A total of 72 Sprague-Dawley (SD) male rats (8 weeks
old) weighing 220–250 g were used in the present study. The animals
were obtained from and housed in the Animal Center of Central South
University, with free access to food and water during the entire
period of experiment. Rats were housed in a temperature (25°C) and
humidity (40–50%) controlled room with a 12 h light/dark cycle. The
rats were randomly divided into the sham (n=8) and SAH groups, and
were decollated 0.5, 3, 10, 24, 48 and 72 h (n=8/group),
respectively following surgery. A total of three rats surviving 48
h post-surgery were used for immunohistochemical analysis; 10 rats
succumbed to mortality following SAH and three rats were excluded
from analysis due to having an SAH grade ≤7.
Surgery and SAH grading
The endovascular perforation model of SAH was
established according to a previously described method (13). Briefly, under anesthesia via
inhalation of 3% isoflurane, a 4-0 monofilament nylon suture was
introduced from the stump of the external carotid artery to the
right internal carotid artery. By further extension, perforation
via the suture was performed at the bifurcation of the anterior
cerebral artery and middle cerebral artery. The sham group rats
underwent the same procedure but without arterial perforation.
Following recovery from anesthesia, the animals were housed for the
aforementioned durations; prior to sacrifice, the behavior of
animals was assessed by an experimenter in a blinded manner using
the SAH Grand scoring system (14).
Neurobehavioral assessment
The behaviors of the animals were assessed at 10,
24, 48 and 72 h post-surgery, respectively, but not at the earlier
time points due to the effects of anesthesia (15). The modified Garcia score test
constitutes 18-point sensory assessments comprising spontaneous
activity, side impact, vibrational touch, limb symmetry, climbing
and forelimb walking. A beam balance test was performed to measure
the ability of an animal to walk on a narrow wooden beam (22.5 mm
diameter) within 60 sec.
Western blot analysis
Protein extraction and western blot analysis were
performed as previously described (16). Left hemispheres were collected and
extracted using radioimmunoprecipitation buffer (Beyotime Institute
of Biotchenology, Haimen, China) and protein concentrations were
measured using a BCA assay kit (Beyotime Institute of
Biotechnology). Lysates (10 µl) from the left cerebral hemisphere
(perforation side) were separated using 10% SDS-PAGE and
transferred onto a polyvinylidene difluoride membrane. Blocking of
nonspecific binding was performed using 5% skimmed milk for 2 h at
room temperature. Immunoblotting was performed overnight at 4°C by
incubation with the following primary antibodies: Rabbit polyclonal
antibodies against TDP-43 cat. no. BM5082; Wuhan Sanying
Biotechnology, Wuhan, China, 1:1,000) and mouse anti-GADPH (cat.
no. 60004-1-Ig; Wuhan Sanying Biotechnology; 1:5,000), followed by
incubation with fluorescent secondary antibodies (cat. no.
SA00001-9; Wuhan Sanying Biotechnology; 1:10,000) for 90 min at
room temperature. The immunoblotting signals were detected using
the Odyssey® CLx Infrared Imaging System (LI-COR
Biosciences, Lincoln, Nebraska USA). To assess the protein
expression in human CSF, 20 µl of concentrated CSF sample from
patients with SAH and controls were used according to the
aforementioned protocol.
Immunocytochemistry
Histology and immunohistochemistry were performed as
previously described (17). The
brain tissues of the SAH and sham groups were perfused with 4%
paraformaldehyde at 48 h post-surgery, post-fixed for 24 h in the
same fixative and cryoprotected via 15 and 30% sucrose prior to
sectioning in a cryostat at 35-µm thickness. The sections were
rinsed in 0.01 M phosphate-buffered saline (PBS; pH 7.4) three
times, and treated with 3% hydrogen peroxide for 30 min to inhibit
endogenous peroxidase activity and then 5% normal horse serum (cat.
no. S-2000; Vector Laboratories, Inc., Burlingame, CA, USA) for 2 h
at room temperature. The sections were then incubated with rabbit
polyclonal antibodies against TDP-43 (1:1,000) overnight at 4°C,
and biotinylated horse anti-rabbit IgG (cat. no. BA-1100; Vector
Laboratories, Inc.; 1:400) for 2 h at 37°C. The immunolabeling was
visualized with ABC reagents (Vector Laboratories, Inc., 1:400),
0.05% DAB and 0.003% H2O2. The immunolabeled
sections were mounted on micro-glass slides, dehydrated and
cover-slipped prior to light microscopic examination by ordinary
light microscope (Olympus BX51; Olympus Corporation, Tokyo,
Japan).
Double immunofluorescence
Double immunofluorescence labeling of rat brain
sections was performed as previously described (18). The sections were washed with PBS and
0.01% Triton X-100, and then blocked with 5% donkey serum (cat. no.
017-000-121; Jackson ImmunoResearch Laboratories, Inc., West Grove,
PA, USA) for 2 h at room temperature. Subsequently, the sections
were incubated overnight at 4°C with rabbit anti-TDP-43 (1:500) in
combination with mouse anti-neuron-specific nuclear antigen (NeuN;
cat. no. MAB377; EMD Millipore; 1:1,000), mouse anti-glial
fibrillary acidic protein (GFAP; cat. no. ab10062; Abcam,
Cambridge, UK; 1:1,000) or mouse anti-ionized calcium-binding
adaptor molecule 1 (Iba-1; cat. no. ab15690; Abcam; 1:500)
antibodies. The sections were then incubated with Alexia 488 and
594-conjugated donkey anti-rabbit and anti-mouse IgG antibodies
(cat. nos. 711-545-152 and 715-585-15, respectively; Jackson
ImmunoResearch Laboratories, Inc.; 1:400) for 2 h at room
temperature. As a negative control, the primary antibodies were
omitted and the same staining procedures were performed. All
sections were incubated with 4,6-diamidino-2-phenylindole for 10
min to visualize cell nuclei via fluorescence microscopy.
Data acquisition, quantification and
statistical analysis
Densitometric data of western blot analysis were
obtained using Odyssey® CLx Infrared Imaging System
(LI-COR Biosciences). The expression levels of TDP-43 were
normalized to the signal of the internal standard (GADPH). Data are
expressed as the mean ± standard deviation. Mean values were
statistically analyzed with GraphPad Prism software (Version 7.0;
GraphPad Software, Inc., La Jolla, CA, USA) using analysis of
variance followed by a Turkey's post hoc test. P<0.05 was
considered to indicate a statistically significant difference.
Results
Protein expression levels of TDP-43
are elevated in the CSF of patients with SAH
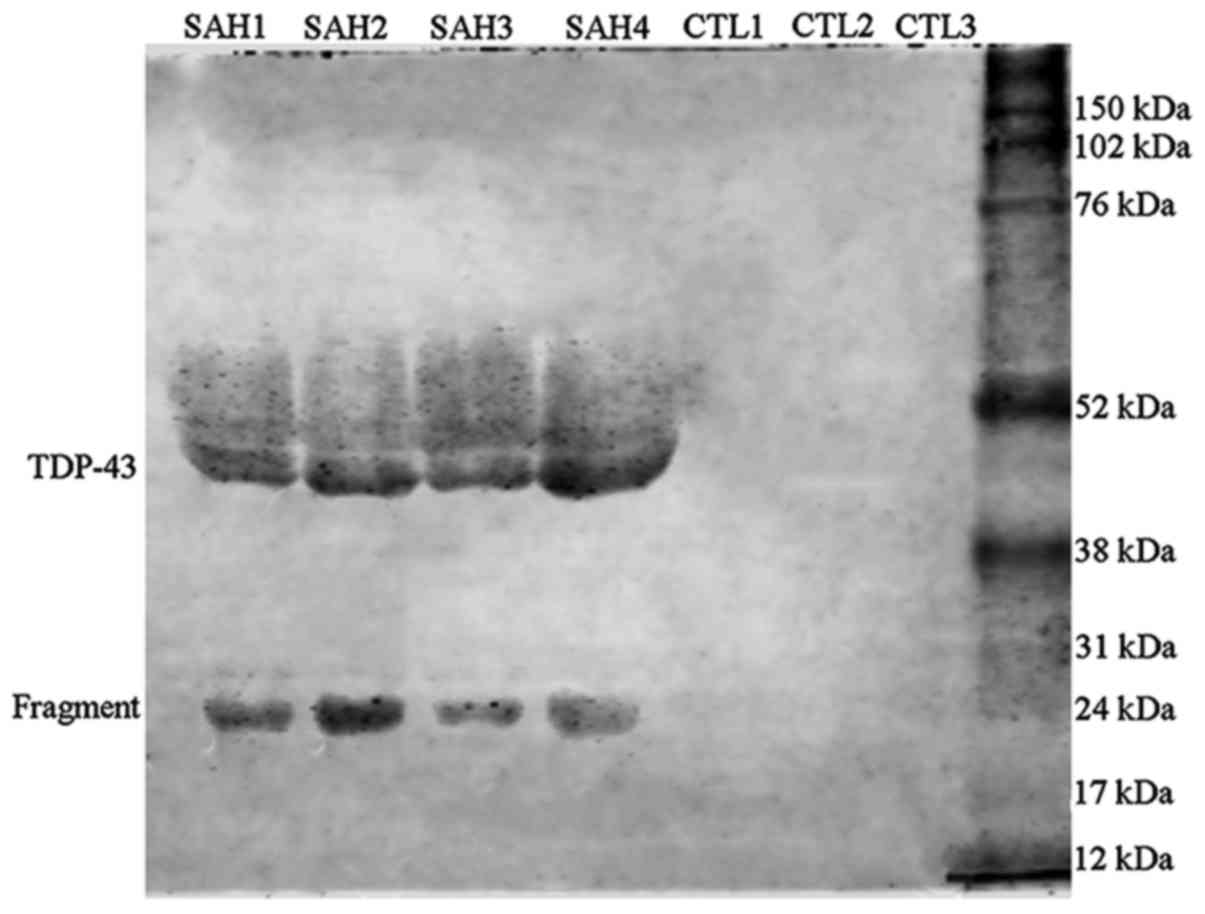
Initially, the present study analyzed the protein
expression of TDP-43 in the blank CSF of patients with SAH by
western blot analysis, which revealed an elevation of this protein,
however, the blotted bands were not readily detected. Therefore, a
freeze enrichment method was applied to concentrate the CSF samples
~4-fold. Subsequent immunoblotting using the concentrated samples
resulted in detectable bands of TDP-43 proteins in the CSF of
patients with SAH. By contrast, the protein expression of TDP-43 in
the concentrated CSF from all control human subjects appeared to be
low (Fig. 1). As the alterations in
the protein expression of TDP-43 of patients with SAH relative to
controls were qualitative, due to distinct bands of the SAH group
compared with in the control, quantification was not performed.
Induction of SAH induces behavioral
changes
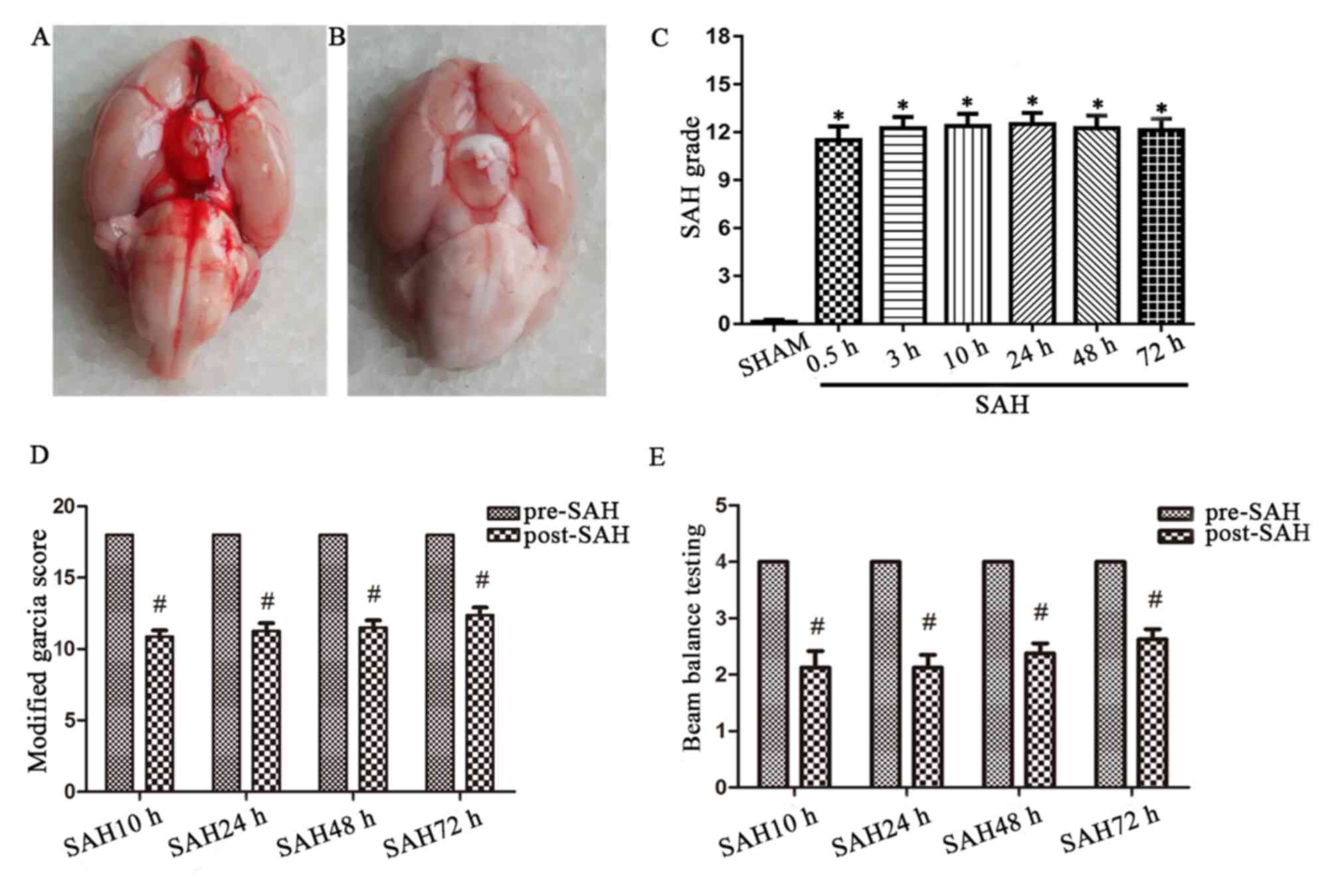
In the present study, a total of 61 male rats were
subjected to SAH surgery, among them, 10 animals succumbed to
mortality, accounting for a mortality rate of 16.3%. No rats
succumbed to mortality in the sham group. Postmortem examination
indicated subarachnoid blood clots around the Circle of Willis and
ventral surface of the brainstem within the brains from the SAH
group (Fig. 2A); blood infiltration
was not observed in the sham-operated group (Fig. 2B). There were significant differences
of the SAH scores between the sham group and each SAH group
(Fig. 2C). The modified Garcia score
and the balance beam score were decreased 10 h following the
induction of SAH, but subsequently improved (Fig. 2D and E).
Protein levels of TDP-43 in rat brain
following the induction of SAH
The present study comparatively assayed the protein
expression levels of TDP-43 in the lysates of the ipsilateral
cortex to vascular perforation in the SAH and sham groups. Within
the rats of the SAH group surviving 0.5–75 h post-surgery, the
expression levels of TDP-43 appeared to increase with longer
durations of survival, relative to the sham control (P<0.05;
Fig. 3A). Increases in the
expression levels of TDP-43 peaked at 48 h and remained elevated at
72 h post-surgery (Fig. 3A and B).
No significant differences were observed in the expression levels
of TDP-43 between the ipsilateral and contralateral cortices of the
SAH group surviving 48 h following surgery (Fig. 3C and D).
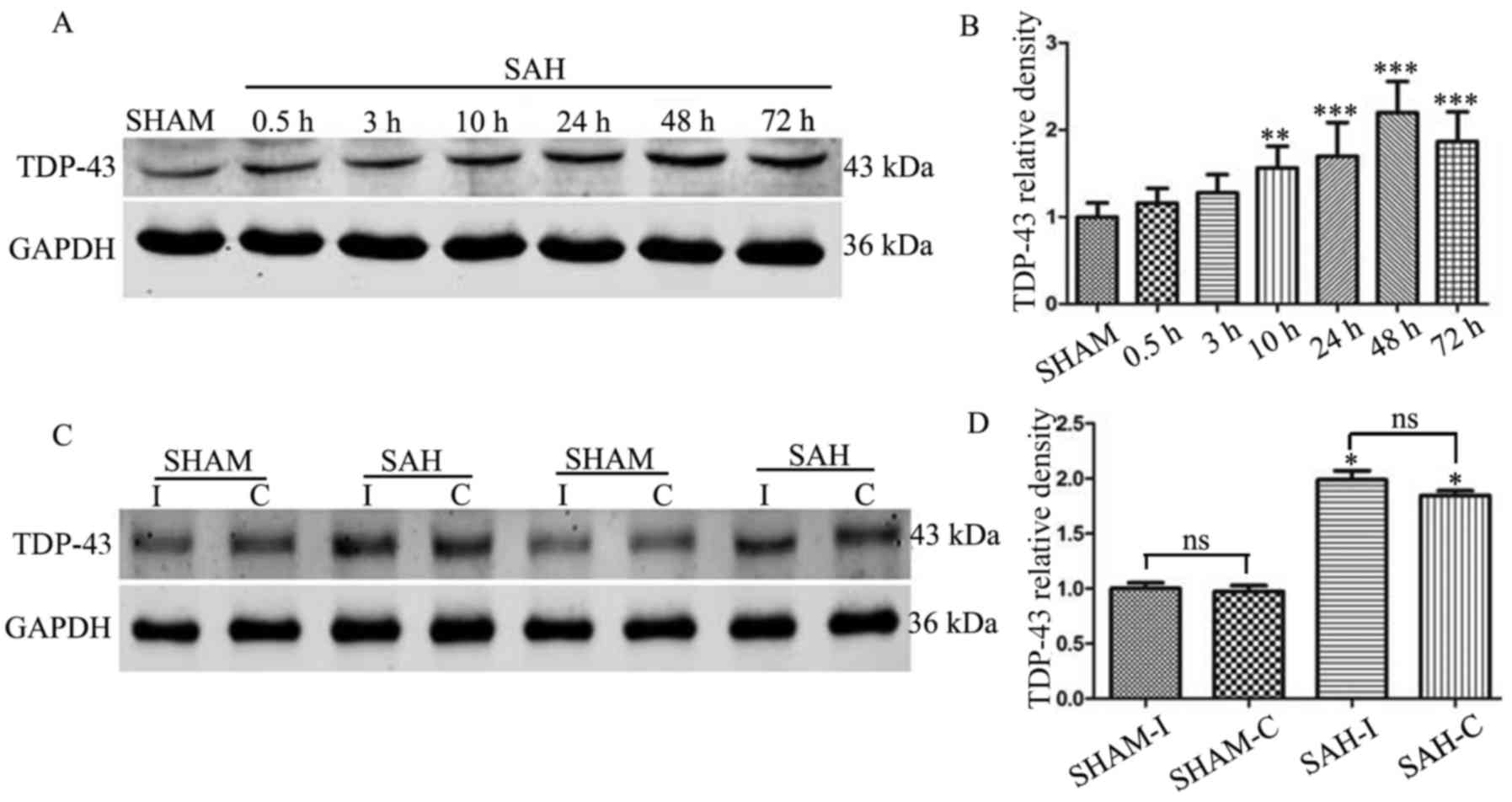 | Figure 3.Western blot analysis of the
expression of TDP-43 in rat brain tissue. (A) Blotted TDP-43 bands
of the sham and SAH rat groups surviving for different durations
(0.5, 3, 10, 24, 48 and 72 h). (B) Relative to the control,
expression levels of TDP-43 gradually increased, peaked at 48 h,
and remained elevated in the experimental groups (**P<0.01 and
***P<0.001 vs. the sham group). (C) Expression levels of TDP-43
of the I and C hemispheric cortex at 48 h following SAH. (D) No
significant difference were observed between the two sides (n=8).
ns, not significant. *P<0.05 vs. the sham group. C,
contralateral; I, ipsilateral; SAH, subarachnoid hemorrhage;
TDP-43, transactive response DNA-binding protein of 43; GAPDH,
glyceraldehyde-3-phosphate dehydrogenase. |
TDP-43 is expressed in neurons and
glial cells within SAH rat brains
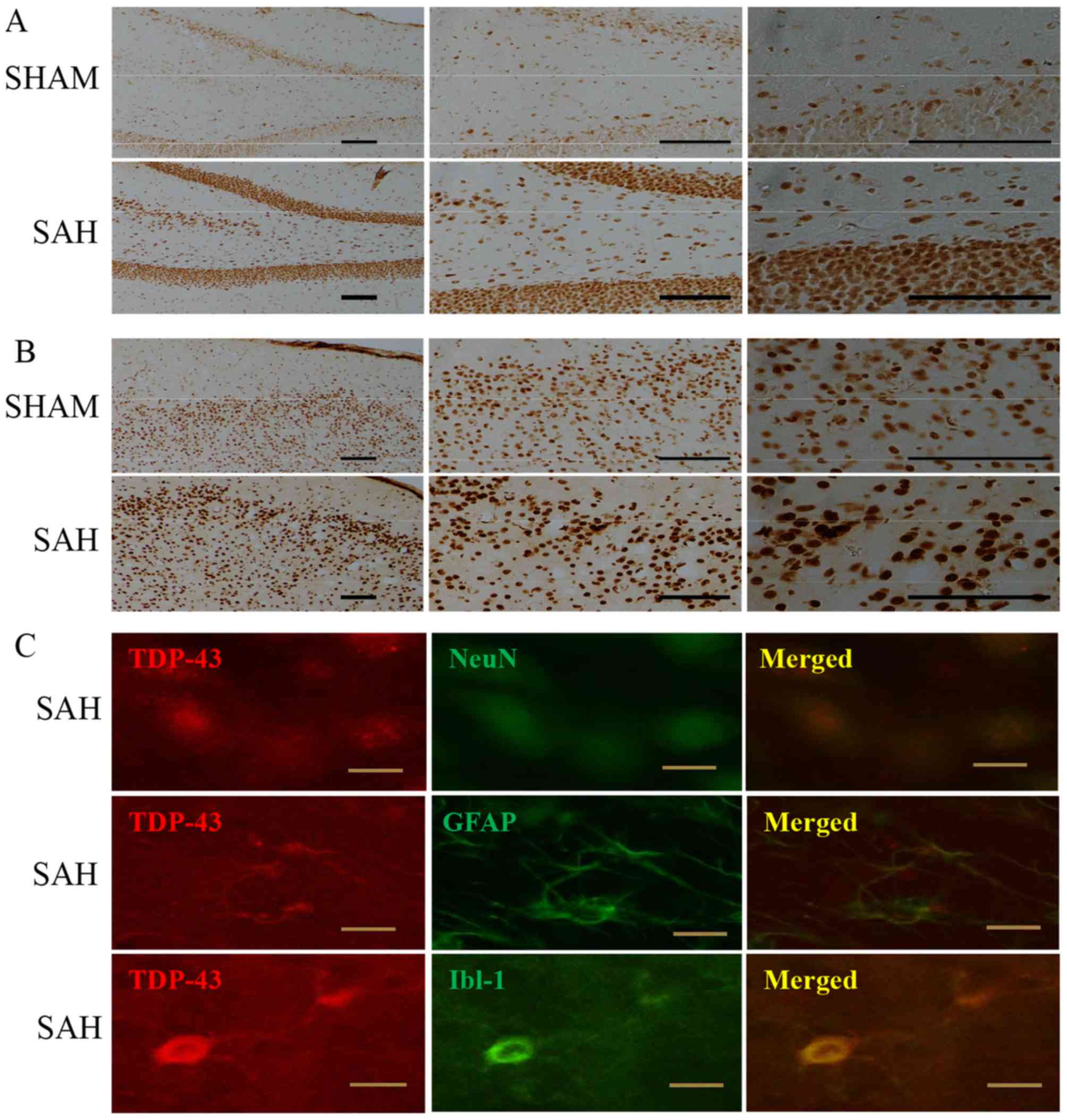
The immunoreactivity of TDP-43 in coronal sections
across the entire rostrocaudal extension of the brain was assessed
using the ABC-DAB method in the present study. Sections from the
sham-operated and SAH animals were processed collectively under
identical conditions. The immunolabeling intensity of the cortical
and subcortical areas, predominantly nucleus-like, appeared to be
more marked in the SAH group compared with that in the sham group
of animals. The difference in labeling intensities between the two
groups were markedly apparent in the hippocampal formation
(Fig. 4A) and the ventral cortical
regions around the sites of accumulated blood clotting (Fig. 4B).
To characterize the types of cells that exhibited
TDP-43 immunoreactivity, sections from rats surviving 48 h
following the induction of SAH were used for confocal double
immunofluorescence analysis. The immunofluorescence suggested that
TDP-43 was co-localized with NeuN, a marker of mature neurons,
GFAP, a marker of astrocytes and Iba-1, a marker of microglia, in
the cerebral cortex, hippocampal formation and subcortical areas
(Fig. 4C).
Discussion
As one type of acute cerebral stroke, SAH has a high
early mortality rate and can cause devastating long-term
neurological outcomes. Therefore, it is important to identify
potential prognostic biomarkers, and understand the neurobiological
and pathological mechanism induced by subarachnoid bleeding. TDP-43
has critical biological roles in the central nervous system (CNS),
and its dysregulation has been associated with numerous chronic
neurodegenerative diseases (6–12). The
present study aimed to investigate whether TDP-43 may be used as
clinical biomarker in patients with SAH, and whether it may be
associated with neurological and cognitive deficits among survivors
of SAH.
As a pilot study, performed assessing TDP-43 in the
CSF for predictive prognosis in patients suffering acute SAH
attack, the present study reported an elevation of this protein in
concentrated CSF samples from patients 72 h following disease
onset. Initially, a classical method of centrifugation of CSF was
used prior to ELISA and western blot analyses, which failed to
reliably detect the protein. It has been reported that TDP-43
protein and a derivative may be detected by immunoblotting in the
CSF from patients with SAH only, but not in that from control
subjects, according to a previously described protocol (19). It appears that this approach may
allow the investigation and follow-up of a large sample size to
evaluate the prognostic value of CSF-associated TDP-43 regarding
mortality risk and long-term neurological outcomes following SAH.
Using the same method, a previous study reported that, within 24 h
of disease onset, TDP-43 may be detected within the CSF of patients
following traumatic brain injury (19). In addition, increased expression
levels of TDP-43 in biological fluids, including CSF and plasma,
have been detected in patients with amyotrophic lateral sclerosis
(ALS) and fronto-temporal lobar degeneration (20). Extracellular TDP-43 may be the result
of cell death induced by TDP-43 inclusion bodies in the cytoplasm
(21). In addition, TDP-43 can be
released from cells by secretory vesicles, known as exosomes
(22,23).
Using a rat experimental model, the present study
demonstrated that, following the induction of SAH, the expression
levels of TDP-43 in brain tissues were rapidly increased and this
elevation persisted for up to 72 h post-injury.
Immunohistochemistry also indicated increased TDP-43 immunolabeling
in the nuclei and cytoplasm of the cortical, hippocampal and
subcortical neurons. Double immunofluorescence suggested that the
expression of TDP-43 was notable in neurons, and may co-localize
with additional proteins in astrocytes and microglia. Of note, the
present study used phosphorylated-TDP-43 antibodies for the
immunoblotting and immunohistochemical analysis of human CSF
samples and rat brain tissues, which failed to obtain a clear
signal. The results of the present study suggested that the acute
upregulation of TDP-43 may largely involve the regular form of
TDP-43 protein. Future investigations aim to investigate the mRNA
expression of TDP-43 in the SAH model.
Inflammation is considered to be an important factor
that mediates the upregulation and translocation of TDP-43 into the
cytoplasm under pathological conditions. Inflammatory factors,
including tumor necrosis factor-α and lipopolysaccharide, may
promote the cytoplasmic accumulation of TDP-43 in neurons and glial
cells, as suggested by the proteinopathy of TDP-43 in
CNS-associated inflammatory diseases, including ALS (24,25). It
has been reported that extracellular TDP-43 activates glial cells
and induces the microglial caspase-3 and interleukin-18 signaling
pathway, resulting in the formation of TDP-43 cytoplasmic inclusion
bodies via reverse translocation of endogenous TDP-43 from the
nucleus to the cytoplasm (21).
Therefore, inflammation induced by SAH may account for the observed
elevation in expression levels of TDP-43 within the CSF and brains
of patients with SAH and experimental rats, respectively.
Future investigations are required to determine
whether, and if so, how the early increases in expression levels of
TDP-43 may impact recovery or may be associated with certain
long-term neuronal dysfunction in patients and experimental animals
with SAH (26,27). TDP-43 has important physiological
effects in neuronal production and axon maintenance, however, it
can also cause abnormal axonal truncation and axonal transport
defects (10,11). Persistent upregulation induced by
injury and the accumulation of TDP-43 may be potentially harmful by
inhibiting dendritic growth and the recovery of neurocircuitry
(12).
Taken together, the clinical analyses of the present
study revealed that CSF-associated levels of TDP-43 were markedly
elevated in patients with SAH soon following the onset of disease.
The animal experiments performed in the present study revealed an
early upregulation of TDP-43 in the rat brain following the
surgical induction of SAH.
The present study analyzed alterations in the
expression of TDP-43 within the CSF and brain tissue following SAH,
which is considered to be a pathological marker protein of
neurodegenerative diseases. Of note, the aggregation and
phosphorylation of TDP-43 may lead to cytotoxicity and long-term
cognitive dysfunction (22).
Therefore, future investigations are required to examine the
long-term effects of the aggregated upregulation of TDP-43 relative
to neuronal damage and recovery in an experimental rat model of
SAH, and then to reduce aggregated TDP-43 via activation of the
heat shock factor 1/heat shock protein pathways (28). Reductions in aggregated TDP-43 in the
early stage following SAH may improve or reverse long-term
behavioral changes. Future investigations also aim to evaluate the
potential of using the CSF-associated expression of TDP-43 as a
prognostic biomarker for SAH clinically.
Acknowledgements
The authors wish to thank their department and
research team for their help and dedication.
Funding
The present study was supported by the Natural
Science Foundation of China (grant nos. ٨١٥٧١١٥٠ and 91632116) and
the New Xiangya Talent Project of the Third Xiangya Hospital of
Central South University (China; grant no. 20160302).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
FL and TBH designed the present study, analyzed the
data and wrote the manuscript; TBH and YCZ established the podocyte
injury model; JL and HXZ collected the CSF samples. KA and XXY
assisted with technical performance and contributed to writing the
manuscript. The final version of the manuscript has been read and
approved by all authors and each author considers the manuscript to
represents honest work.
Ethics approval and consent to
participate
The present study was performed in compliance with
Chinese legislations and guidelines of the National Institutes of
Health involving human subjects and for the use and care for
experimental animals. Written informed consent was obtained from
all patients. Experiments involving animals and humans were
approved by the Ethics Committees of Central South University
Xiangya School of Medicine and The Third Xiangya Hospital of
Central South University, respectively.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Al-Khindi T, Macdonald RL and Schweizer
TA: Cognitive and functional outcome after aneurysmal subarachnoid
hemorrhage. Stroke. 41:e519–e536. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Takata K, Sheng H, Borel CO, Laskowitz DT,
Warner DS and Lombard FW: Long-term cognitive dysfunction following
experimental subarachnoid hemorrhage: New perspectives. Exp Neurol.
213:336–344. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Egashira Y, Hua Y, Keep RF and Xi G: Acute
white matter injury after experimental subarachnoid hemorrhage:
Potential role of lipocalin 2. Stroke. 45:2141–2143. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Kummer TT, Magnoni S, Mac Donald CL,
Dikranian K, Milner E, Sorrell J, Conte V, Benetatos JJ, Zipfel GJ
and Brody DL: Experimental subarachnoid haemorrhage results in
multifocal axonal injury. Brain. 138:2608–2618. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Fern RF, Matute C and Stys PK: White
matter injury: Ischemic and nonischemic. Glia. 62:1780–1789. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Lee EB, Lee VM and Trojanowski JQ: Gains
or losses: Molecular mechanisms of TDP43-mediated
neurodegeneration. Nat Rev Neurosci. 13:38–50. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Schwab C, Arai T, Hasegawa M, Yu S and
McGeer PL: Colocalization of transactivation-responsive DNA-binding
protein 43 and huntingtin in inclusions of Huntington disease. J
Neuropathol Exp Neurol. 67:1159–1165. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Yokota O, Tsuchiya K, Arai T, Yagishita S,
Matsubara O, Mochizuki A, Tamaoka A, Kawamura M, Yoshida H, Terada
S, et al: Clinicopathological characterization of Pick's disease
versus frontotemporal lobar degeneration with
ubiquitin/TDP-43-positive inclusions. Acta Neuropathol.
117:429–444. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Onozato T, Nakahara A, Suzuki-Kouyama E,
Hineno A, Yasude T, Nakamura T, Yahikozawa H, Watanabe M, Kayanuma
K, Makishita H, et al: Axonal TDP-43 aggregates in sporadic
amyotrophic lateral sclerosis. Neuropathol Appl Neurobiol.
42:561–572. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ishiguro A, Kimura N, Watanabe Y, Watanabe
S and Ishihama A: TDP-43 binds and transports
G-quadruplex-containing mRNAs into neurites for local translation.
Genes Cells. 21:466–481. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Tripathi VB, Baskaran P, Shaw CE and
Guthrie S: Tar DNA-binding protein-43 (TDP-43) regulates axon
growth in vitro and in vivo. Neurobiol Dis. 65:25–34. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Herzog JJ, Deshpande M, Shapiro L, Rodal
AA and Paradis S: TDP-43 misexpression causes defects in dendritic
growth. Sci Rep. 7:156562017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Liu F, Hu Q, Li B, Manaenko A, Chen Y,
Tang J, Guo Z, Tang J and Zhang JH: Recombinant milk fat
globule-EGF factor-8 reduces oxidative stress via integrin
β٣/nuclear factor erythroid 2-related factor 2/heme oxygenase
pathway in subarachnoid hemorrhage rats. Stroke. 45:3691–3697.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Sugawara T, Ayer R, Jadhav V and Zhang JH:
A new grading system evaluating bleeding scale in filament
perforation subarachnoid hemorrhage rat model. J Neurosci Methods.
167:327–334. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Liu F, Chen Y, Hu Q, Li B, Tang J, He Y,
Guo Z, Feng H, Tang J and Zhang JH: MFGE8/Integrin β٣ pathway
alleviates apoptosis and inflammation in early brain injury after
subarachnoid hemorrhage in rats. Exp Neurol. 272:120–127. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Hu Q, Ma Q, Zhan Y, He Z, Tang J, Zhou C
and Zhang J: Isoflurane enhanced hemorrhagic transformation by
impairing antioxidant enzymes in hyperglycemic rats with middle
cerebral artery occlusion. Stroke. 42:1750–1756. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ren M, Li K, Wang D, Guo J, Li J, Yang G,
Long X, Shen W, Hu R, Wang X and Zeng K: Neurofibromin regulates
seizure attacks in the rat pilocarpine-induced model of epilepsy.
Mol Neurobiol. 53:6069–6077. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yan J, Manaenko A, Chen S, Klebe D, Ma Q,
Caner B, Fujii M, Zhou C and Zhang JH: Role of SCH79797 in
maintaining vascular integrity in rat model of subarachnoid
hemorrhage. Stroke. 44:1410–1417. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yang Z, Lin F, Robertson CS and Wang KK:
Dual vulnerability of TDP-43 to calpain and caspase-3 proteolysis
after neurotoxic conditions and traumatic brain injury. J Cereb
Blood Flow Metab. 34:1444–1452. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Noto Y, Shibuya K, Sato Y, Kanai K, Misawa
S, Sawai S, Mori M, Uchiyama T, Isose S, Nasu S, et al: Elevated
CSF TDP-43 levels in amyotrophic lateral sclerosis: specificity,
sensitivity, and a possible prognostic value. Amyotroph Lateral
Scler. 12:140–143. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Leal-Lasarte MM, Franco JM,
Labrador-Garrido A, Pozo D and Roodveldt C: Extracellular TDP-43
aggregates target MAPK/MAK/MRK overlapping kinase (MOK) and trigger
caspase-3/IL-18 signaling in microglia. FASEB J. 31:2797–2816.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Iguchi Y, Eid L, Parent M, Soucy G, Bareil
C, Riku Y, Kawai K, Takagi S, Yoshida M and Katsuno M: Exosome
secretion is a key pathway for clearance of pathological TDP-43.
Brain. 139:3187–3201. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zondler L, Feiler MS, Freischmidt A, Ruf
WP, Ludolph AC, Danzer KM and Weishaupt JH: Impaired activation of
ALS monocytes by exosomes. Immunol Cell Biol. 95:207–214. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Neumann M, Sampathu DM, Kwong LK, Truax
AC, Micsenyi MC, Chou TT, Bruce J, Schuck T, Grossman M, Clark CM,
et al: Ubiquitinated TDP-43 in frontotemporal lobar degeneration
and amyotrophic lateral sclerosis. Science. 314:130–133. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Correia AS, Patel P, Dutta K and Julien
JP: Inflammation Induces TDP-43 mislocalization and aggregation.
PLoS One. 10:E01402482015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Lucke-Wold BP, Turner RC, Logsdon AF,
Bailes JE, Huber JD and Rosen CL: Linking traumatic brain injury to
chronic traumatic encephalopathy: Identification of potential
mechanisms leading to neurofibrillary tangle development. J
Neurotrauma. 31:1129–1138. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Dewey CM, Cenik B, Sephton CF, Johnson BA,
Herz J and Yu G: TDP-43 aggregation in neurodegeneration: Are
stress granules the key? Brain Res. 1462:16–25. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Chen HJ, Mitchell JC, Novoselov S, Miller
J, Nishimura AL, Scotter EL, Vance CA, Cheetham ME and Shaw CE: The
heat shock response plays an important role in TDP-43 clearance:
Evidence for dysfunction in amyotrophic lateral sclerosis. Brain.
139:1417–1432. 2016. View Article : Google Scholar : PubMed/NCBI
|