Introduction
Rheumatoid arthritis (RA) is a systemic, chronic,
inflammatory autoimmune disease that affects joints and other
tissues, including the skin and kidneys (1). RA affects a large number of individuals
worldwide, with nearly three times as many females as men being
diagnosed with RA (2). Studies have
suggested that the pathogenesis of RA is triggered by failure in
the maintenance of peripheral tolerance to self-antigens, which
leads to the activation and expansion of autoreactive T and B cells
(3,4). These pathogenic T cells and
autoantibodies in turn cause joint inflammation that eventually
leads to irreversible articular cartilage damage as well as the
breakdown of osseous tissue (3,4). Various
proinflammatory cytokines serve important roles in RA development
(4). Patients with RA exhibit
increased numbers of interleukin (IL)-17-producing T helper (h)17
cells in the peripheral blood in comparison with healthy subjects
(5,6). Additionally, IL-17 was elevated in the
synovial fluid of patients with RA and was involved in the
pathogenesis of RA (7,8). Regulatory T cells (Tregs) were revealed
to be crucial for peripheral self-tolerance, as they actively
suppress auto-reactive lymphocytes as well as other immune cells
and express suppressive cytokines, including IL-10 and transforming
growth factor (TGF)-β (9). It has
been reported that Tregs serve an important role in the
pathogenesis of RA by regulating autoimmunity and inflammation
(10,11). Inducible T-cell costimulator (ICOS)
is a co-receptor belonging to the cluster of differentiation (CD)28
immunoglobulin (Ig) super family that provide co-stimulatory
signals to T cells during antigen-mediated activation (12).
Reports on murine models for autoimmune diabetes and
allergic asthma have described the important roles of ICOS in the
generation, maintenance and suppressive functions of Tregs
(13,14). Notably, ICOS+ Tregs
exhibited a stronger suppressive activity compared with
ICOS− Tregs (15).
Furthermore, ICOS+ Tregs released IL-10 to suppress
dendritic cell function and TGF-β to suppress T cell function,
while ICOS− Tregs mainly secreted TGF-β (16). The blockade or absence of ICOS
inhibited the production of IL-10 and abrogated the inhibitory
function of Tregs (17,18). Although an increasing number of
studies have indicated that ICOS signaling is necessary for Treg
suppression in various diseases (13,14),
little is known about the roles of ICOS+ Tregs in RA.
Various studies have reported that a Th17/Treg imbalance is
associated with the development of RA (19–21).
However, the precise association between ICOS+ Tregs and
Th17 in patients with RA has yet to be determined. The current
study examined the ICOS+ Tregs in a cohort of Chinese
patients with RA, particularly in patients with inactive RA (iRA).
The proportions of ICOS+ Tregs in patients with RA were
assessed according to DAS28 scores. The current study also explored
the expression levels of ICOS+ Tregs from patients with
iRA and the levels of suppressive cytokines, including IL-10,
TGF-β, IL-35 and IL-17. The present study was performed to clarify
whether ICOS+ Tregs may perform inflammatory and
inhibitory functions, and if abnormal ICOS+ Tregs
numbers and functions may contribute to the pathogenesis of RA.
Patients and methods
Patients and healthy controls
The current study, which took place at the
Department of Rheumatology of Nanfang Hospital, Southern Medical
University (Guangzhou, China) from January 2015 to January 2016,
examined 33 Chinese patients diagnosed with RA according to the
American College of Rheumatology (ACR) criteria (22). These patients included 31 females and
2 males (age, 49.76±1.73 years). Information concerning the
patient's medical history, number of classification criteria
fulfilled and laboratory findings were obtained at the time of the
clinic visit. Whole blood samples were collected and disease
activity was measured using the 28-joint disease activity index
(DAS28) (23). Based on the DAS28,
10 patients possessed iRA (DAS28 score ≤3.3) and 23 possessed aRA
(DAS28 score >3.3) during the study. Additionally, 17 healthy
subjects were recruited to serve as the normal control (NC) group,
the details of which are presented in Table I. All participants provided written
consent during their enrollment. The Ethics Committee of Nanfang
Hospital approved the current study.
 | Table I.Demographic features of normal
controls and RA patients. |
Table I.
Demographic features of normal
controls and RA patients.
| Variable | RA | NC |
|---|
| Total number | 33 | 17 |
| Sex
(male/female) | 2/31 | 1/16 |
| Age (years) | 49.76±1.73 | 33.35±1.39 |
| DAS28>3.3 | 23 | N/A |
| DAS28≤3.3 | 10 | N/A |
Antibodies and reagents
Anti-CD4-fluoresceinisothiocyanate (FITC; cat. no.
11-0048-42), anti-ICOS-phycoerythrin (PE)-Cy7 (cat. no.
85-25-9948-42) and anti-TGF-β-PE (cat. no. 12-9829-42) antibodies
were purchased from (eBioscience; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA). Anti-CD25-PerCP-Cy5.5 (cat. no. 560503), Mouse
IgGl-PE-Cy7 (cat. no. 25-4714-41), Mouse IgGl-PE (cat. no.
12-4714-41), Mouse IgGl-PerCP-Cy5.5 (cat. no. 550795),
anti-Foxp3-PE-CF594 (cat. no. 562421), Mouse IgGl-PE-CF594 (cat.
no. 562292), anti-IL-10-APC (cat. no. 554707) and anti-IL-17-V450
(cat. no. 560610), Mouse IgGl-V450 (cat. no. 560373) antibodies
were purchased from BD Biosciences (Franklin Lakes, NJ, USA).
Anti-human-EBI3-APC (cat. no. IC6456A), anti-human-p35-PE (cat. no.
IC2191P) antibodies were purchased from R&D Systems, Inc.
(Minneapolis, MN, USA). Additional reagents included Mouse IgGl-APC
(cat. no. 70-CMG105-10; MultiSciences, ZheJiang, China), OptiLyse C
lysing solution (Beckman Coulter, Inc., Brea, CA, USA), PBS, fetal
bovine serum (FBS; both Thermo Fisher Scientific, Inc.), Foxp3
Staining Buffer Set kit (BD Biosciences, NJ, USA) and Monensin
(eBioscience; Thermo Fisher Scientific, Inc.), phorbol myristate
acetate (PMA) (cat. no. P-8139; Sigma-Aldrich; Merck KGaA,
Darmstadt, Germany) ionomycin (cat. no. I-0634; Sigma-Aldrich;
Merck KGaA), brefeldin A (BFA; cat. no. B-7651; Sigma-Aldrich;
Merck KGaA) and Ficoll (Tianjin Haoyang 4E Products Technology Co.,
Ltd., Tianjin, China).
Flow cytometry
For fresh staining, 100 µl fresh heparinized
peripheral blood was initially stained with anti-CD4-FITC,
anti-CD25-PerCP-Cy5.5, anti-ICOS-PE-Cy7, and the aforementioned
isotypes (Mouse IgGl-PE-Cy7 and Mouse IgGl-PerCP-Cy5.5; all at a
dilution of 1:20) for 20 min at 4°C following the lysis of red
blood cells using OptiLyse C lysing solution. The cells were washed
twice with 1 ml washing buffer (PBS with 2% FBS) and the Foxp3
Staining Buffer Set kit was used to perform intracellular staining
of Foxp3 according to the manufacturer's protocol. Acquisitions
were performed on a BD LSRFortesa flow cytometer (BD Biosciences),
collecting ≥0.1 million events for each sample. The data was
analyzed with BD FACSDiva software v8.0.2. (BD Biosciences).
Intracellular cytokine detection
To detect intracellular cytokines, including IL-10,
IL-17, TGF-β and IL-35 fresh peripheral blood cells were stimulated
with PMA (40 ng/ml) and ionomycin (1 µg/ml) in the presence of BFA
(20 µg/ml) at 37°C for 5 h. Following stimulation, cells were
collected and stained for anti-CD4-FITC, anti-CD25-PerCP-Cy5.5,
anti-ICOS-PE-Cy7 and indicated intracellular cytokines such as
anti-Foxp3-PE-CF594, anti-TGF-β-PE, anti-IL-10-APC,
anti-IL-17-V450, anti-human-EBI3-APC and anti-human-p35-PE (all
1:20) respectively. Samples were collected using a BD LSRFortesa
flow cytometer and data was analyzed using BD FACSDiva software
v8.0.2.
Peripheral blood mononuclear cells
culture and cytokine analysis
Ficoll density gradient centrifugation (400 × g, 30
min, 21°C) was used to isolate fresh peripheral mononuclear blood
cells (PBMCs) from 5 ml heparin-treated venous blood samples. PBMCs
were cultured for 6 days in 12-well culture plates pre-coated with
1 µg/ml purified NA/LE Mouse Anti-Human CD3 (cat. no. 555336; BD
Pharmingen; BD Biosciences) with the addition of 300 U/ml
recombinant human IL-2 (cat. no. 200-02; Perprotech, Inc., London,
UK) for T-cell receptor (TCR) stimulation. Aliquots of PBMCs
(1×106) in 2 ml RPMI 1640 (cat. no. 72400047; Gbico;
Thermo Fisher Scientific, Inc.) were supplemented with 10%
heat-inactivated fetal bovine serum (Gbico; Thermo Fisher
Scientific, Inc.), glutamine (2 mM), penicillin (200 U/ml) and
streptomycin (100 µg/ml). Cells were either stained and analysis or
treated with PMA and ionomycin in the presence of BFA for 5 h prior
to harvesting, performing intracellular cytokine staining and
conducting fluorescence-activated cell sorting analysis.
Statistical analysis
All statistical analyses were performed using Prism
v5.0 GraphPad software (GraphPad Software, Inc., La Jolla, CA,
USA). Quantitative data were expressed as mean ± standard error of
the mean. One-Way analysis of variance followed by a Kruskal-Wallis
H test was performed to compare three groups. Two-group comparisons
were examined by a Mann-Whitney U test. Spearman's correlation was
used to assess correlations. P<0.05 indicated that the
difference between groups was statistically significant.
Results
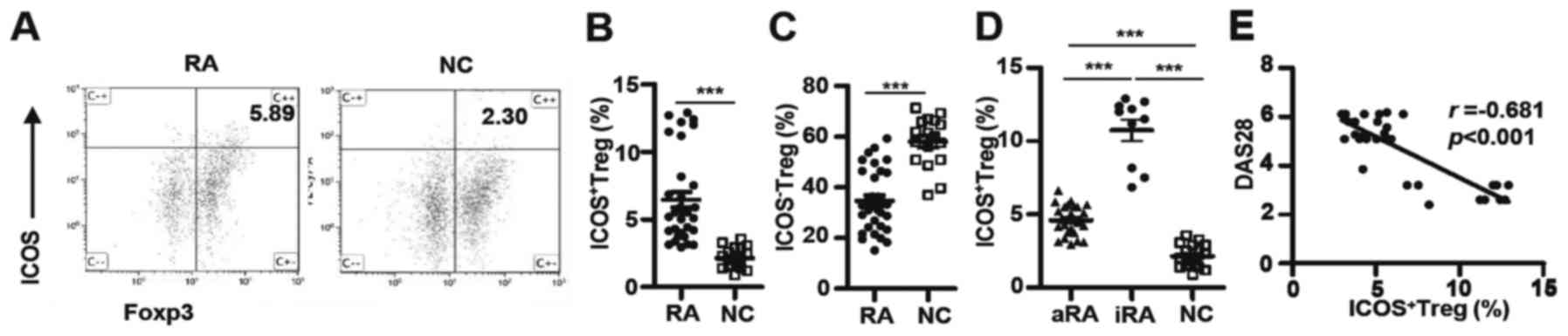
ICOS+ Tregs frequencies are
higher in patients with RA but negatively correlate with disease
activity
Tregs are essential for the maintenance of
peripheral tolerance, for preventing autoimmunity and for limiting
chronic inflammatory diseases (24).
Although it has been reported that the number of Tregs are
decreased in patients with RA (3)
and ICOS+ Tregs may be involved in the pathogenesis of
numerous tumors and autoimmune diseases (9), it is unknown whether or not
ICOS+ Tregs are associated with RA. To address this
issue, the percentages of ICOS+ Tregs identified in a
cohort of Chinese patients with RA and NCs were evaluated.
ICOS+ Tregs percentages were significantly increased in
patients with RA compared with the NCs (6.46±0.56% vs. 2.15±0.20%;
P<0.0001; Fig. 1A and B). By
contrast, the percentages of ICOS− Tregs in patients
with RA were considerably decreased (34.77±2.10% vs. 58.08±2.40%;
P<0.0001; Fig. 1C). Patients with
RA were further divided into aRA and iRA groups according to the
DAS28 scores. As indicated in Fig.
1D, the percentages of ICOS+ Tregs were
significantly increased in the aRA group (4.60±0.23%; P<0.001)
and the iRA group (10.74±0.73%; P<0.001) compared with the NC
group (2.15±0.20%). Additionally, the iRA group exhibited twice the
number of ICOS+ Tregs as the aRA group (P<0.0001).
Notably, DAS28 scores and ICOS+ Tregs percentages were
negatively correlated in patients with RA (r=−0.681; P<0.001;
Fig. 1E). Thus, ICOS+
Tregs were elevated in patients with RA; however, the magnitude of
increase was negatively correlated with disease activity in all
patients.
ICOS+ Tregs/Th17 ratios are
disturbed in patients with RA
Th17 cells serve important roles in autoimmune
inflammation (25). Consistent with
previous studies (5,6), it was revealed that Th17 cell
percentages in the patients with RA were significantly larger
compared with the NCs (6.04±0.84% vs. 0.43±0.05%; P<0.0001;
Fig. 2A). Additionally, Th17
percentages were ~6.5 times higher in patients with aRA
(8.09±0.91%) compared with that in patients with iRA (1.34±0.18%;
P<0.0001) and ~7.5 times higher than in the NCs (0.43±0.05%;
P<0.0001; Fig. 2B). Although the
number of Th17 cells was significantly lower in the iRA group
compared with that in the aRA group, Th17 percentages in the iRA
group were remained significantly high compared with the NCs
(P<0.0001).
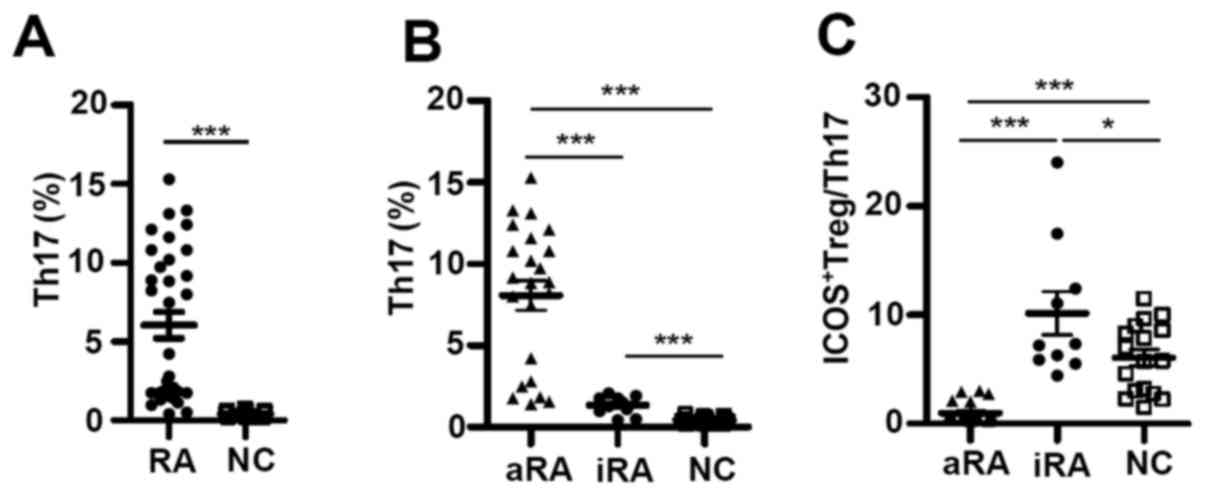 | Figure 2.ICOS+ Tregs/Th17 ratios
are disturbed in patients with RA. (A) The percentage of Th17 in
patients with RA (n=33) and in NCs (n=17; Mann-Whitney U test). (B)
The percentage of Th17 and (C) ratio of ICOS+ Tregs/Th17
in aRA patients (n=23), iRA patients (n=10), and NCs (n=17). Data
are presented as mean ± standard error of the mean. *P<0.05,
***P<0.0001. ICOS, inducible T-cell costimulator; RA, rheumatoid
arthritis; NCs, normal controls; aRA, active RA; iRA, inactive RA;
Tregs, regulatory T cells; Th17, T helper 17 cells. |
It has been reported that Tregs can inhibit Th17
activity, and a skewed Th17/Treg balance may be a characteristic
feature of chronic inflammatory diseases such as RA (21). However, the association between
ICOS+ Tregs and Th17 in RA remains controversial. Given
the increases of ICOS+ Tregs and Th17 cells in patients
with RA, their relative abundance in patients with RA and NCs were
analyzed. The ratios between ICOS+ Tregs and Th17 cells
were significantly lower in the aRA group (0.97±0.19) compared with
the NCs (6.07±0.77; P<0.001). However, the ratios were
significantly higher in the iRA group (10.17±1.99) in comparison
with either the aRA group (0.97±0.19; P<0.0001) or the NCs
(6.07±0.77; P<0.05; Fig. 2C).
Taken together, these data reveal that
ICOS+ Tregs and Th17 were increased in patients with RA;
however, their respective increases differed depending on the
status of the disease. This suggests that imbalances between
ICOS+ Tregs and Th17 may contribute to disease
progression and remission.
Anti-inflammatory cytokines increase
in RA, particularly iRA, whereas pro-inflammatory cytokine IL-17
increases in aRA
Cytokines produced by various synovial cell
populations serve significant roles in the pathogenesis of RA
(26). To further examine whether
cytokine profiles of ICOS+ Tregs were dysregulated in
patients RA, T cells were stimulated with PMA and ionomycin for 5 h
in the presence of BFA, followed by intracellular staining to
detect IL-10, TGF-β, IL-35 (anti-inflammatory cytokines) and IL-17
(a pro-inflammatory cytokine). As demonstrated in Fig. 3A and B, ICOS+ Tregs from
the iRA group contained twice the number of IL-10-producing cells
than those from the aRA group (9.75±1.32% vs. 4.09±0.32%;
P<0.005) and 5 times more IL-10-producing cells compared with
those from the NCs (1.90±0.59%; P<0.0005). The percentages of
IL-10-producing ICOS+ Tregs from patients with aRA were
higher compared with those in NCs (P<0.005).
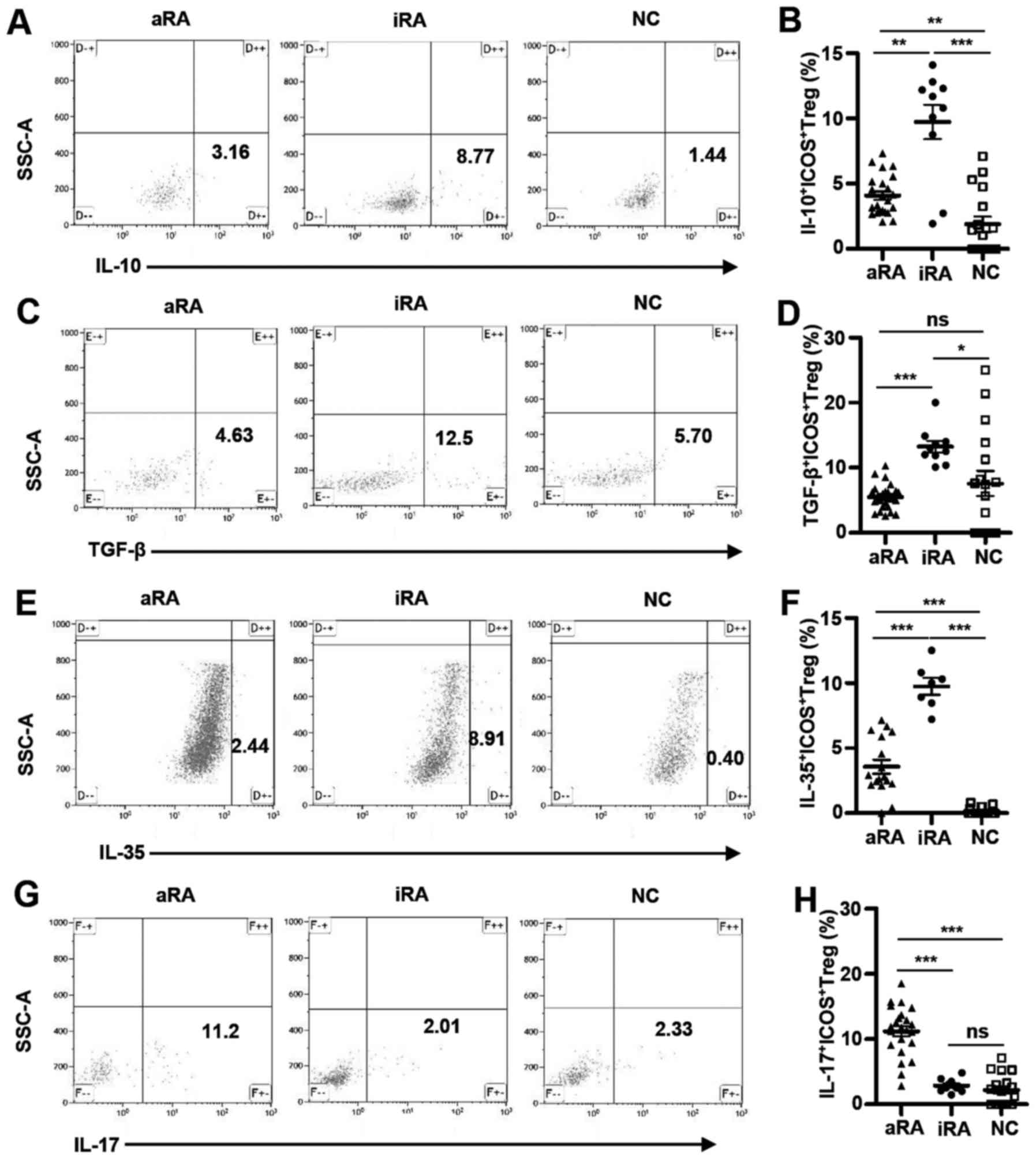 | Figure 3.Anti-inflammatory cytokines increase
in RA, particularly iRA, while a pro-inflammatory cytokine
increases in aRA. The numbers of cytokine-secreting ICOS+ Tregs
were measured in patients with aRA (n=23), patients with iRA (n=10)
and NCs (n=17). (A) Representative dot plots of IL-10 staining (B)
and the percentage of IL-10+ICOS+ Tregs. (C)
Representative dot plots of TGF-β staining (D) and the percentage
of TGF-β+ICOS+ Tregs. (E) Representative dot
plots of IL-35 staining and (F) the percentage of
IL-35+ICOS+ Tregs. (G) Representative dot
plots of IL-17 staining and (H) the percentage of
IL-17+ICOS+ Tregs. Data are presented as mean
± standard error of the mean. *P<0.05, **P<0.005,
***P<0.0005. ICOS, inducible T-cell costimulator; RA, rheumatoid
arthritis; NCs, normal controls; aRA, active RA; iRA, inactive RA;
Tregs, regulatory T cells; SSC-A, side scatter area; IL,
interleukin; TGF, transforming growth factor; ns, not
significant. |
Similarly, the percentages of TGF-β-producing
ICOS+ Tregs were increased in patients with iRA in
comparison with patients with aRA (13.25±0.89% vs. 5.52±0.43%;
P<0.0001) and the NCs (7.58±1.93%; P<0.05; Fig. 3C and D). However, no significant
differences in TGF-β-producing ICOS+ Tregs were
identified between the aRA group and the NCs.
IL-35, which serves a role in immune suppression of
murine Treg, is a dimeric protein composed of IL-12α and IL-27β
chains that is encoded by IL12A and EBI3 genes, respectively
(27). IL-35 producing
ICOS+ Tregs were detected from fresh PBMCs after 5 h of
PMA and ionomycin stimulation. However, as Fig. 3E and F demonstrated, after
stimulation of PBMCs with anti-CD3 and IL-2 in vitro for 6
days, IL-35-producing cells could be detected in ICOS+
Tregs and were significantly increased in the iRA group in
comparison with the aRA group (9.74±0.65% vs. 3.57±0.53%;
P<0.0001). The NCs exhibited a small number of IL-35-producing
ICOS+ Tregs in comparison with the aRA group
(P<0.0001) ICOS+ Tregs from the NCs (0.17±0.07%)
possessed significantly fewer IL-35-producing cells compared with
those from the iRA group (P<0.0005).
In contrast to these suppressive cytokines, the aRA
group contained higher percentages of IL-17-producing cells within
ICOS+ Tregs compared with the iRA group (11.23±0.79% vs.
2.89±0.31%, respectively) or the NCs (2.22±0.58%; both P<0.0001;
Fig. 3G and H). No significant
differences in the percentage of IL-17-producing ICOS+
Tregs were identified between the iRA and NC groups.
Together, these data reveal that the aRA group had
increased IL-17-expressing ICOS+ Tregs but decreased
IL-10, TGF-β and IL-35-expressing ICOS+ Tregs in
comparison with the iRA group. Additionally, in comparison with the
NCs, the aRA group had increased IL-17 and slightly increased IL-10
and IL-35, but decreased TGF-β-expressing ICOS+ Tregs.
Among these three groups, the iRA group had the highest IL-10,
TGF-β and IL-35-expressing ICOS+ Tregs. ICOS+
Tregs may perform inflammatory and inhibitory functions by
expressing IL-17, IL-10, TGF-β and IL-35.
Cytokine expression profile prior to
and following TCR stimulation are similar
TCR signaling serves several important roles in
Tregs function. Given the number of abnormal ICOS+ Tregs
and cytokine expression in patients with RA, PBMCs were stimulated
from patients with aRA and iRA, and NCs with anti-CD3 and IL-2
in vitro for 6 days. The number of ICOS+ Tregs
and cytokine expression were subsequently analyzed. As indicated in
Fig. 4A, the number of
ICOS+ Tregs was highest in the iRA group (16.24±1.67%)
and lowest in the NC group (3.40±0.42%; P<0.0001). In addition,
IL-10-producing cells in ICOS+ Tregs were higher in the
iRA group (16.93±1.69) compared with that in the aRA (3.72±0.41;
P<0.0001) and NC groups (0.80±0.33; P<0.0001; Fig. 4B). TGF-β-producing cells in
ICOS+ Tregs were increased in the iRA group (23.96±1.08)
compared with that in the aRA (7.54±0.80; P<0.0001) and NC
groups (9.21±1.10; P<0.0001; Fig.
4C). Although the number of IL-10-producing cells in
ICOS+ Tregs from the iRA group was higher compared with
that of the NCs, the two groups had similar numbers of
TGF-β-producing cells. In comparison with suppressive cytokine
expression, the number of IL-17-producing cells in ICOS+
Tregs was highest in the aRA group, and no significant difference
was identified between the iRA and NC groups (Fig. 4C). Thus, the differences in
ICOS+ Tregs observed in freshly isolated PBMCs from the
three groups remained virtually unchanged following in vitro
TCR and IL-2 stimulation.
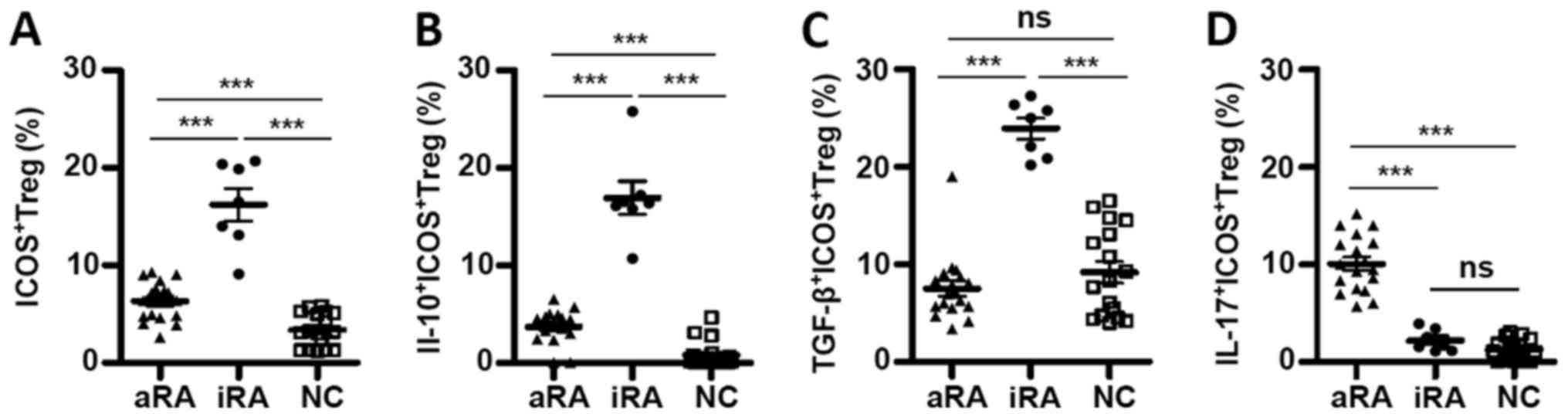 | Figure 4.Cytokine expression profile prior to
and following TCR stimulation are similar. PBMCs were stimulated
with anti-cluster of differentiation 3 and IL-2 in vitro for
6 days. The percentage of (A) ICOS+(B)
IL-10+ICOS+ (C)
TGF-β+ICOS+ and (D)
IL-17+ICOS+ in Tregs in patients with aRA
(n=18), patients with iRA (n=7) and NCs (n=17). Data are presented
as mean ± standard error of the mean. ***P<0.0001. ICOS,
inducible T-cell costimulator; RA, rheumatoid arthritis; NCs,
normal controls; aRA, active RA; iRA, inactive RA; Tregs,
regulatory T cells; IL, interleukin; TGF, transforming growth
factor; ns, not significant. |
Discussion
It has been reported that Th17 cells promote
autoimmunity and inflammation, and diseases associated with these
conditions such as RA (8), whereas
Tregs are involved in maintaining immune responses and preventing
autoimmunity (11,28). The balance between activated
responder T cells and Tregs may influence the extent of
immunoregulation in RA (29).
Additionally, humans and mice contain ICOS+ and
ICOS− Tregs, which exhibit different properties. The
current study demonstrated the amount of ICOS+ Tregs
increased in a cohort of Chinese patients with RA. The increase in
ICOS+ Tregs was larger in patients with iRA compared
with those in patients with aRA, and the frequencies of
ICOS+ Tregs in patients with RA were negatively
correlated with DAS28 scores. It was reported that IL-17, TGF-β and
IL-6 levels detected by ELISA were increased in peripheral blood
serum of patients with RA (30).
Although the serum or plasma cytokines may reflect the status of
disease in a patient, since several types of cells can secrete the
total peripheral suppressive cytokines, the current study focused
only on the cytokines produced by Tregs. The data demonstrated that
the ICOS+ Tregs from patients with iRA expressed
multiple suppressive cytokines, including IL-10, TGF-β and IL-35,
but they expressed low levels of IL-17. The patients with aRA
exhibited ICOS+ Tregs that expressed decreased levels of
suppressive cytokines, but also had noticeably increased IL-17
levels. Data from the current study suggests that an abnormal
number of ICOS+ Tregs and abnormal ICOS+ Treg
functions may contribute to the pathogenesis of RA.
Although ICOS+ cells account for a minor
population within Tregs, emerging evidence has indicated that
ICOS+ Tregs exhibit a strong suppressive function and
that a decline in these Tregs may contribute to autoimmune disease
in humans (13–16). Higher levels of ICOS expression in
Tregs have been identified to correlate with improved control of
autoimmune disease, diabetes and atherosclerosis (14,18). The
current study revealed that ICOS+ Tregs were increased
in patients with RA in comparison with NCs, suggesting that the
ICOS+ Tregs count itself may not be the sole indicator
for disease occurrence. However, patients with RA exhibit an
increased number of Th17 cells, which cause decreased
ICOS+ Tregs/Th17 ratios. The data from the current study
suggest that imbalances between ICOS+ Tregs and Th17 may
contribute to RA pathogenesis and that the ICOS+
Tregs/Th17 ratio in patients with RA may be more indicative of
disease progression than the ICOS+ Tregs percentage
itself.
Vocanson et al (31) reported that
CD4+CD25+FoxP3+ T cells
upregulated ICOS expression during in vivo sensitization,
and specifically suppress hapten-reactive CD8+ T cells
in vivo and in vitro. The aforementioned study
further demonstrated that ICOS+ Tregs distinguish
themselves from ICOS− FoxP3+ Tregs through
their expression of IL-10, IL-17 and interferon-γ. In the current
study, it was demonstrated that ICOS+ Tregs exhibit a
special cytokine profile characterized by the expression of
inflammatory cytokine IL-17 and anti-inflammatory cytokines IL-10,
TGF-β and IL-35. Notably, ICOS+ Tregs from patients with
iRA exhibited more anti-inflammatory cytokines compared with
patients with aRA. In contrast, ICOS+ Tregs from
patients with aRA expressed more IL-17 compared with those from the
iRA group. This data suggests that ICOS+ Tregs
contribute to RA pathogenesis via at least two mechanisms. On one
hand, ICOS+ Tregs produce inhibitory cytokines to
suppress inflammation. On the other hand, ICOS+ Tregs
may express proinflammatory cytokines that exacerbate RA. The
balance between these two functions may dictate disease
progression. Elevated pathogenic IL-17 expression and decreased
suppressive cytokine-producing ICOS+ Tregs may promote
disease development in patients with aRA. In contrast, predominance
of suppressive cytokine-producing ICOS+ Tregs over
IL-17-producing ICOS+ Tregs may reduce inflammation and
help ensure an inactive state in patients with RA.
Acknowledgements
The authors would like to thank Dr Xiaoping Zhong
(Division of Allergy and Immunology, Department of Pediatrics, Duke
University Medical Center, Durham, NC27710, USA) for their helpful
advice and assistance.
Funding
The present study was supported by the Nanfang
Hospital Foundation of SMU (grant no. 201362) and the Guangdong
Natural Science Foundation (grant no. 2017A030313508).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
HXW, XK, WL and Y-RQ conceived and designed the
experiments. XK, SC, HL, XL, XY performed the experiments. HXW, SC,
XK and WL analyzed the data. HXW, XK and SC wrote the manuscript.
Y-RQ had full access to all of the data in the study and takes
responsibility for the integrity of the data and the accuracy of
the data analysis.
Ethics approval and consent to
participate
Ethics approval was received from Nanfang Hospital,
Southern Medical University. All participants gave written consent
during their enrollment.
Patient consent for publication
All patients gave written consent during their
enrollment for publication.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Firestein GS: Evolving concepts of
rheumatoid arthritis. Nature. 423:356–361. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Del Junco DJ, Annegers JF, Coulam CB and
Luthra HS: The relationship between rheumatoid arthritis and
reproductive function. Br J Rheumatol. 1 Soppl 28:(33): 42–45.
1989.
|
|
3
|
Picerno V, Ferro F, Adinolfi A, Valentini
E, Tani C and Alunno A: One year in review: The pathogenesis of
rheumatoid arthritis. Clin Exp Rheumatol. 33:551–558.
2015.PubMed/NCBI
|
|
4
|
McInnes IB and Schett G: The pathogenesis
of rheumatoid arthritis. N Engl J Med. 365:2205–2219. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Yue C, You X, Zhao L, Wang H, Tang F,
Zhang F, Zhang X and He W: The effects of adalimumab and
methotrexate treatment on peripheral Th17 cells and IL-17/IL-6
secretion in rheumatoid arthritis patients. Rheumatol Int.
30:1553–1557. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Lu TT, Zhu P, Li XY and Fan CM: Functional
status of T helper cells in rheumatoid arthritis and effect of
etanercept. Xi Bao Yu Fen Zi Mian Yi Xue Za Zhi. 24:495–497.
2008.(In Chinese). PubMed/NCBI
|
|
7
|
Miossec P: Interleukin-17 in fashion, at
last: Ten years after its description, its cellular source has been
identified. Arthritis Rheum. 56:2111–2125. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Shahrara S, Huang Q, Mandelin AN II and
Pope RM: TH-17 cells in rheumatoid arthritis. Arthritis Res Ther.
10:R932008. View
Article : Google Scholar : PubMed/NCBI
|
|
9
|
Campbell DJ: Control of regulatory T cell
migration, function, and homeostasis. J Immunol. 195:2507–2513.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Byng-Maddick R and Ehrenstein MR: The
impact of biological therapy on regulatory T cells in rheumatoid
arthritis. Rheumatology (Oxford). 54:768–775. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Cooles FA, Isaacs JD and Anderson AE: Treg
cells in rheumatoid arthritis: An update. Curr Rheumatol Rep.
15:3522013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Dong C, Juedes AE, Temann UA, Shresta S,
Allison JP, Ruddle NH and Flavell RA: ICOS co-stimulatory receptor
is essential for T-cell activation and function. Nature.
409:97–101. 2001. View
Article : Google Scholar : PubMed/NCBI
|
|
13
|
Busse M, Krech M, Meyer-Bahlburg A, Hennig
C and Hansen G: ICOS mediates the generation and function of
CD4+CD25+Foxp3+ regulatory T cells conveying respiratory tolerance.
J Immunol. 189:1975–1982. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Herman AE, Freeman GJ, Mathis D and
Benoist C: CD4+CD25+ T regulatory cells dependent on ICOS promote
regulation of effector cells in the prediabetic lesion. J Exp Med.
199:1479–1489. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Chen Y, Shen S, Gorentla BK, Gao J and
Zhong XP: Murine regulatory T cells contain hyperproliferative and
death-prone subsets with differential ICOS expression. J Immunol.
188:1698–1707. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ito T, Hanabuchi S, Wang YH, Park WR,
Arima K, Bover L, Qin FX, Gilliet M and Liu YJ: Two functional
subsets of FOXP3+ regulatory T cells in human thymus and periphery.
Immunity. 28:870–880. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kohyama M, Sugahara D, Sugiyama S, Yagita
H, Okumura K and Hozumi N: Inducible costimulator-dependent IL-10
production by regulatory T cells specific for self-antigen. Proc
Natl Acad Sci USA. 101:pp. 4192–4197. 2004; View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Gotsman I, Grabie N, Gupta R, Dacosta R,
MacConmara M, Lederer J, Sukhova G, Witztum JL, Sharp AH and
Lichtman AH: Impaired regulatory T-cell response and enhanced
atherosclerosis in the absence of inducible costimulatory molecule.
Circulation. 114:2047–2055. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Nistala K and Wedderburn LR: Th17 and
regulatory T cells: Rebalancing pro-and anti-inflammatory forces in
autoimmune arthritis. Rheumatology (Oxford). 48:602–606. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Niu Q, Cai B, Huang ZC, Shi YY and Wang
LL: Disturbed Th17/Treg balance in patients with rheumatoid
arthritis. Rheumatol Int. 32:2731–2736. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wang W, Shao S, Jiao Z, Guo M, Xu H and
Wang S: The Th17/Treg imbalance and cytokine environment in
peripheral blood of patients with rheumatoid arthritis. Rheumatol
Int. 32:887–893. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Aletaha D, Neogi T, Silman AJ, Funovits J,
Felson DT, Bingham CO III, Birnbaum NS, Burmester GR, Bykerk VP,
Cohen MD, et al: 2010 rheumatoid arthritis classification criteria:
An american college of rheumatology/european league against
rheumatism collaborative initiative. Ann Rheum Dis. 69:1580–1588.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Prevoo ML, van't Hof MA, Kuper HH, van
Leeuwen MA, van de Putte LB and van Riel PL: Modified disease
activity scores that include twenty-eight-joint counts. Development
and validation in a prospective longitudinal study of patients with
rheumatoid arthritis. Arthritis Rheum. 38:44–48. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Sakaguchi S, Yamaguchi T, Nomura T and Ono
M: Regulatory T cells and immune tolerance. Cell. 133:775–787.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Bedoya SK, Lam B, Lau K and Larkin J III:
Th17 cells in immunity and autoimmunity. Clin Dev Immunol.
2013:9867892013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kirkham BW, Lassere MN, Edmonds JP, Juhasz
KM, Bird PA, Lee CS, Shnier R and Portek IJ: Synovial membrane
cytokine expression is predictive of joint damage progression in
rheumatoid arthritis: A two-year prospective study (the DAMAGE
study cohort). Arthritis Rheum. 54:1122–1131. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Collison LW, Workman CJ, Kuo TT, Boyd K,
Wang Y, Vignali KM, Cross R, Sehy D, Blumberg RS and Vignali DA:
The inhibitory cytokine IL-35 contributes to regulatory T-cell
function. Nature. 450:566–569. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wing K and Sakaguchi S: Regulatory T cells
exert checks and balances on self tolerance and autoimmunity. Nat
Immunol. 11:7–13. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
van Amelsfort JM, Jacobs KM, Bijlsma JW,
Lafeber FP and Taams LS: CD4(+)CD25(+) regulatory T cells in
rheumatoid arthritis: Differences in the presence, phenotype, and
function between peripheral blood and synovial fluid. Arthritis
Rheum. 50:2775–2785. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Pan F, Xiang H, Yan J, Hong L, Zhang L,
Liu Y, Feng X and Cai C: Dendritic cells from rheumatoid arthritis
patient peripheral blood induce Th17 cell differentiation via
miR-363/Integrin αv/TGF-β axis. Scand J Immunol. 85:441–449. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Vocanson M, Rozieres A, Hennino A, Poyet
G, Gaillard V, Renaudineau S, Achachi A, Benetiere J, Kaiserlian D,
Dubois B and Nicolas JF: Inducible costimulator (ICOS) is a marker
for highly suppressive antigen-specific T cells sharing features of
TH17/TH1 and regulatory T cells. J Allergy Clin Immunol.
126:280–289. 2010. View Article : Google Scholar : PubMed/NCBI
|