Introduction
Myocardial infarction is a leading cause of
mortality in humans worldwide, which results from myocardial
ischemia (1,2). Timely reperfusion is considered as an
effective therapy to limit the infarction size, although
reperfusion of the ischemic myocardium will increase the number of
patients with heart failure due to the cardiomyocyte death
(3). The mitochondria are involved
in the reperfusion injury, and the opening of mitochondrial
permeability transition pore (MPTP) contributes to the infarction
(4,5). In addition, intracellular
Ca2+ overload and oxidative stress in mitochondria are
also associated with myocardial infarction (6). In the development of effective
therapeutic approaches for myocardial infarction, natural compounds
serve an important role, including berberine (7), tanshinone IIA (8) and lycopene (9).
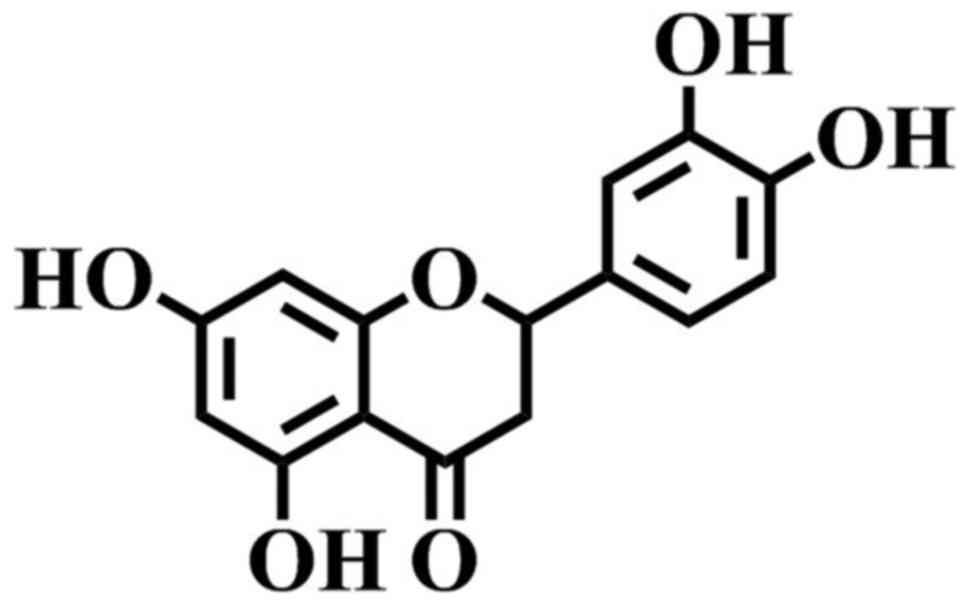
Eriodictyol (Fig. 1)
is a flavonoid identified in numerous medicinal plants, such as
Bauhinia ungulata (10),
Arcytophyllum thymifolium (11), Elsholtzia bodinieri (12) and Clinopodium chinense
(13). Pharmacological
investigations have revealed that eriodictyol possesses several
bioactivities, including neuroprotection (14–16),
renoprotection (17) and lung
protection (18), exerted via
anti-inflammation and anti-oxidation. In addition, its
anti-inflammatory and anti-oxidative capacities have drawn
attention to its therapeutic potential (19–23).
With the aim to investigate bioactive phytochemicals
for the treatment of myocardial infarction, the protective effects
of eriodictyol on H9c2 cardiomyocyte injury induced by
hypoxia/reoxygenation are investigated in the present study. The
protective effect of eriodictyol is reported in vitro.
Materials and methods
Chemicals and reagents
Eriodictyol was purchased from YuanYe Biotechnology
Co., Ltd. (Shanghai, China). Dulbecco's modified Eagle's medium
(DMEM), fetal bovine serum (FBS) and calcein acetoxymethyl
(calcein-AM) were obtained from Thermo Fisher Scientific, Inc.
(Waltham, MA, USA). Dimethyl sulfoxide (DMSO) and
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT)
and were purchased from Sigma-Aldrich (Merck AG, Darmstadt,
Germany). Reactive oxygen species (ROS), lactate dehydrogenase
(LDH) and bicinchoninic acid (BCA) assay kits were supplied by
Nanjing Jiancheng Bioengineering Institute (Nanjing, China). JC-1,
Fluo-3 AM, ATP detection kit and caspase-3 assay kit, as well as
cleaved caspase-3 (cat. no. AC033), B-cell lymphoma-2 (Bcl-2; cat.
no. AB112), Bcl-2-associated X protein (Bax; cat. no. AB026) and
actin (cat. no. AA128) antibodies (all 1:1,000) were purchased from
Beyotime Institute of Biotechnology (Nantong, China).
Cell culture and treatment
The H9c2 cardiomyocytes, a rat embryonic cardiac
myoblast line, were obtained from the Cell Bank of Chinese Academy
of Sciences (Shanghai, China) and cultured as previously described
(9). Briefly, H9c2 cells were
cultured in DMEM containing 10% FBS and 1% penicillin/streptomycin
under humid condition with 5% CO2 and 95% air at 37°C.
Next, cells at the logarithmic phase were incubated in 96-well
plates at a density of 1×105/ml. The cells were then
divided into the control group (CG), model group (MG) and three
eriodictyol groups, which were pretreated with 1, 10 and 50 µM
eriodictyol in DMSO for 4 h. In order to establish the
hypoxia/reoxygenation model, H9c2 cells in the MG and eriodictyol
groups were subjected to an atmosphere with 95% N2 and
5% O2 at 37°C for 4 h, and then cultured under a
condition of 95% air and 5% O2 at 37°C for a further 4
h. The CG cells were cultured under normoxic conditions.
Cell viability assay
To evaluate the protective effect of eriodictyol on
the H9c2 cardiomyocyte injury induced by hypoxia/reoxygenation, an
MTT assay was conducted. Following the aforementioned treatments,
the cells were incubated with 0.2 ml MTT for 4 h at 37°C.
Subsequently, 200 µl DMSO was added into each well to dissolve the
formazan crystals, and the absorbance was recorded on an iMark
microplate reader (Bio-Rad Laboratories, Inc., Hercules, CA, USA)
at 490 nm. The experiments were repeated three times.
Determination of LDH activity
The extracellular LDH activity was determined using
the LDH assay kit, according to the manufacturer's instructions.
Following treatment and incubation, the H9c2 cardiomyocyte culture
medium was centrifuged at 400 × g and room temperature for 5 min,
and then 20 µl supernatant was mixed with 20 µl
2,4-dinitrophenylhydrazine. The mixture was incubated at 37°C for
15 min. Next, 250 µl NaOH (0.4 M) was added into the reaction
system and incubated for a further 15 min at 37°C. Subsequent to
keeping at room temperature for 5 min, the absorbance was recorded
on a microplate reader at 450 nm. The activity of LDH was
calculated based on the absorbance as previously reported (24) and is expressed as U/l.
Detection of intracellular
Ca2+
In order to monitor the cytosolic Ca2+
content in H9c2 cardiomyocytes, the Fluo-3 AM molecular
fluorescence probe was employed. Following treatment as described
earlier, H9c2 cardiomyocytes were incubated with Fluo-3 AM at 37°C
for 30 min and washed twice with phosphate-buffered saline (PBS) to
remove any extracellular dye. Subsequent to incubation for a
further 30 min, the fluorescence intensity was measured on a
SpectraMax M5 microplate reader (Molecular Devices, LLC, Sunnyvale,
CA, USA) at an excitation wavelength of 488 nm and emission
wavelength of 525 nm.
Measurement of ROS generation
The production of intracellular ROS was detected by
a fluorescence method (25) with the
ROS assay kit, according to the manufacturer's protocol. Following
treatment, the medium was replaced and the cells were loaded with
10 µM 2,7-dichlorodihydrofluorescein diacetate (DCFH-DA). After
incubation at 37°C for 30 min, the cells were rinsed with PBS and
the fluorescence intensity was recorded on a fluorescence
microplate reader at an excitation wavelength of 480 nm and
emission wavelength of 525 nm.
Assessment of mitochondrial membrane
potential (MMP)
The fluorescent probe JC-1 was used to detect the
MMP in H9c2 cardiomyocytes. In normal mitochondria, the JC-1
monomer aggregates in the matrix, whereas JC-1 maintains its
monomeric form in mitochondria with reduced MMP. The fluorescence
of JC-1 aggregates is measured at an excitation wavelength of 530
nm and emission wavelength of 590 nm. In the current investigation,
cardiomyocytes were loaded with JC-1 (100 µM) at 37°C for 20 min
and then washed with PBS. The fluorescence intensity of the JC-1
aggregate was recorded on a fluorescence microplate reader, and the
MMP was determined as the ratio of the JC-1 fluorescence intensity
to that of the control group.
Opening of MPTP
The opening of MPTP was evaluated by determining the
release of mitochondrial calcein, as previously described (26). Briefly, cardiomyocytes were incubated
with 2 µM calcein-AM and 1 mM CoCl2 at room temperature
for 30 min. Next, free calcein-AM and CoCl2 were washed
away with Hank's balanced salt solution, and cells were incubated
with CoCl2 for a further 20 min at 37°C in order to
quench the fluorescence of free cytosolic calcein. The fluorescence
of mitochondrial calcein in the cardiomyocytes was recorded on a
fluorescence microplate reader at 490 nm for excitation and 515 nm
for emission. The quenching of fluorescence in H9c2 cardiomyocytes
indicated the opening of MPTP.
Level of intracellular ATP
The intercellular ATP in H9c2 cardiomyocytes was
determined by the firefly luciferase method (25) with the ATP detection kit according to
the manufacturer's protocol. Luciferin generates fluorescence under
the catalysis of firefly luciferase, and the process consumes ATP
quantitatively. The treated H9c2 cardiomyocytes were lysed on ice
with 200 µl lysis reagent from the assay kit. The lysed cells were
then centrifuged at 12,000 × g for 4 min at 4°C, and 100 µl
supernatant was mixed with 100 µl ATP monitoring reagent.
Subsequently, the luminescence was detected on a microplate reader,
and the level of intracellular ATP was derived from the standard
curve.
Caspase-3 activity
The activity of caspase-3 was quantified through a
colorimetric detection kit, following the manufacturer's
instructions. Briefly, subsequent to the aforementioned treatments,
the H9c2 cardiomyocytes were lysed and centrifuged at 16,000 × g
for 10 min at 4°C. The supernatant was then incubated with the
substrate Ac-DEVD-pNA at 37°C for 2 h, and the absorbance was
measured on a microplate reader at 405 nm. The relative caspase-3
activity was expressed as a percentage of the control group, as
previous described (27).
Western blot analysis
The protein expression levels of caspase-3, Bcl-2
and Bax in pretreated H9c2 cardiomyocytes were analyzed by western
blot analysis. In brief, the cells were lysed with lysis reagent
containing 20 mM Tris-HCl (pH 7.4), 150 mM NaCl, 1% Triton X-100
and 1 mM phenylmethylsulfonyl fluoride on ice for 30 min. Next, the
lysate was centrifuged at 12,000 × g for 15 min at 4°C, and the
supernatant was collected for the analysis of cleaved caspase-3,
Bcl-2 and Bax levels. The total protein concentration in the
samples was determined by BCA assay kit. Subsequently, the protein
was separated by electrophoresis on a 15% SDS-polyacrylamide gel
and transferred to polyvinylidene difluoride membranes. Following
blocking with 5% skimmed milk at room temperature for 1 h, the
membranes were incubated overnight at 4°C with primary antibodies
against cleaved caspase-3, Bcl-2, Bax and actin. The membranes were
was with TBST three times, treated with the respective secondary
antibodies conjugated to horseradish peroxidase (cat. no. LDAN0310;
1:1,000; Shanghai Lengton Bioscience Co., Ltd., Shanghai, China) at
room temperature for 1 h and detected by an Enhanced ECL
Chemiluminescent Substrate kit (cat. no. 36222ES60; Shanghai Yeasen
Biotechnology Co., Ltd., Shanghai, China). Actin was used as the
internal control.
Statistical analysis
All results are expressed as the means ± standard
deviation. GraphPad Prism (version 5.0; GraphPad Software, Inc., La
Jolla, CA, USA) was employed to analyze the results. Statistical
differences between different groups were compared by one-way
analysis of variance followed by Dunnett's test for multiple
comparisons and Student's t-test for single comparisons. P<0.05
was considered to indicate a statistically significant
difference.
Results
Effect of eriodictyol on H9c2
cardiomyocyte viability
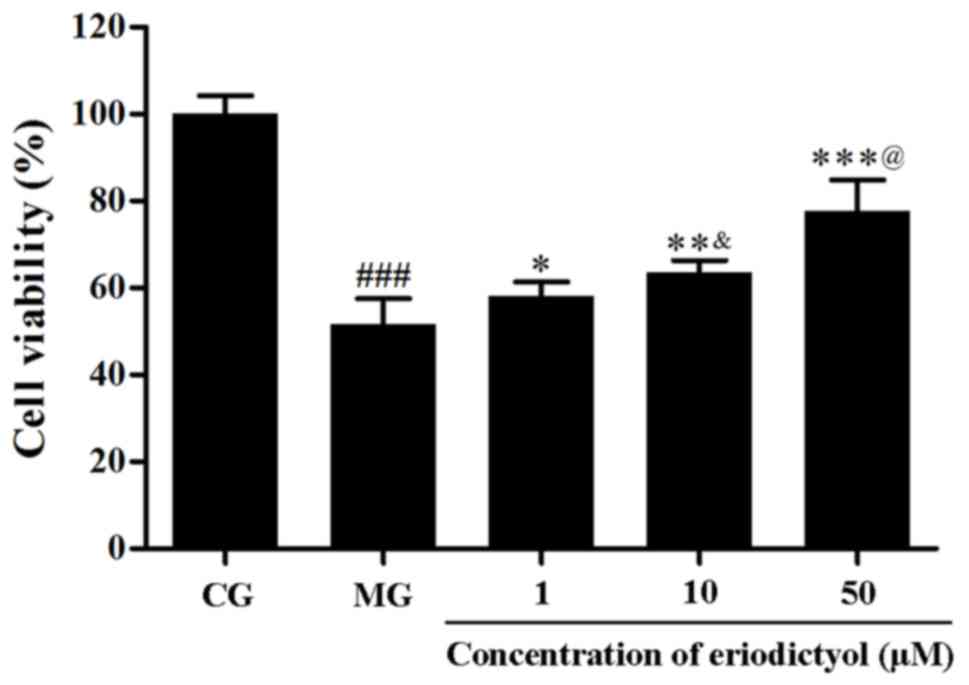
As shown in Fig. 2,
the MTT assay demonstrated that the viability of H9c2
cardiomyocytes decreased when subjected to the
hypoxia/reoxygenation (P<0.001). Upon treatment with different
dosages of eriodictyol, the survival of H9c2 cardiomyocytes was
significantly improved and the cells viability was 77.75±7.06% of
the cell viability of CG when cells were treated with 50 µM
eriodictyol (P<0.001). The viability in H9c2 cardiomyocytes
treated with 50 µM eriodictyol (77.75±7.06%) was significantly
higher than the cells treated with 10 µM eriodictyol (63.56±2.75%;
P<0.001) and the latter was significantly increased compared
with the group treated with 1 µM eriodictyol (58.16±3.17%;
P<0.05). These results indicate the potential cardioprotective
effect of eriodictyol on H9c2 cells in a dose-dependent manner at
the range of 1–50 µM eriodictyol.
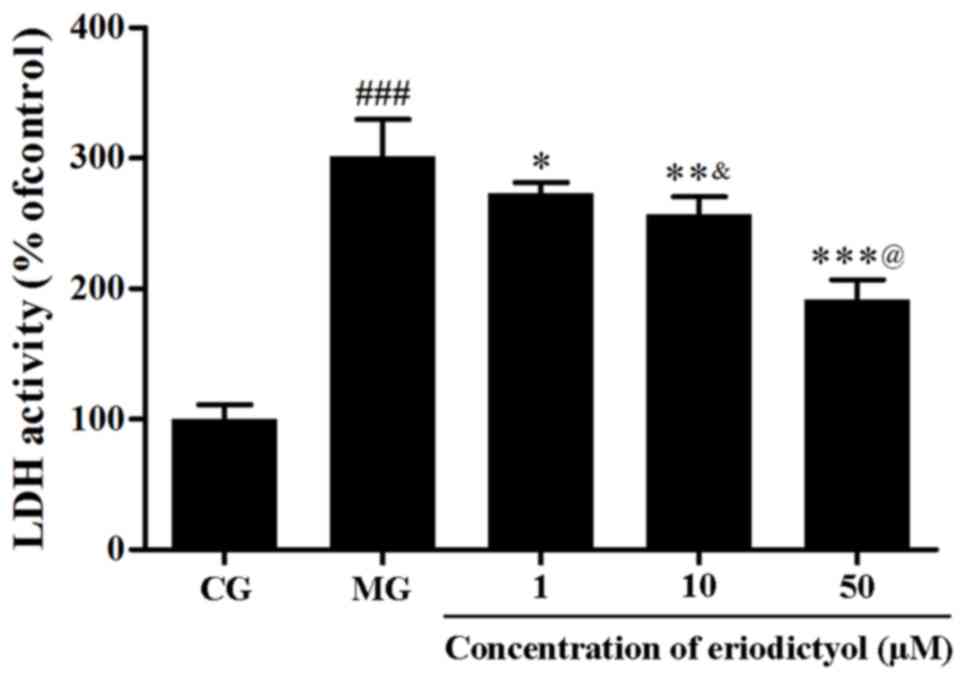
Effect of eriodictyol on LDH
activity
The leakage of LDH from the cytoplasm is associated
with cell death. In this investigation, the activity of LDH in the
culture medium of the MG (301.01±28.81%) was significantly higher
as compared with that in the CG (P<0.001), which further
demonstrated that the viability of H9c2 cells was affected by
hypoxia/reoxygenation. When pretreated with eriodictyol, the
activity of LDH in the 10 µM group with was reduced to
256.81±13.75%, which was significantly higher than the 50 µM group
(191.56±13.75%; P<0.001) and lower than the 1 µM group
(272.94±8.44%; P<0.05). The results indicated the release of LDH
from the cytosol of H9c2 cardiomyocytes was evidently decreased in
a dosage-dependent manner (Fig.
3).
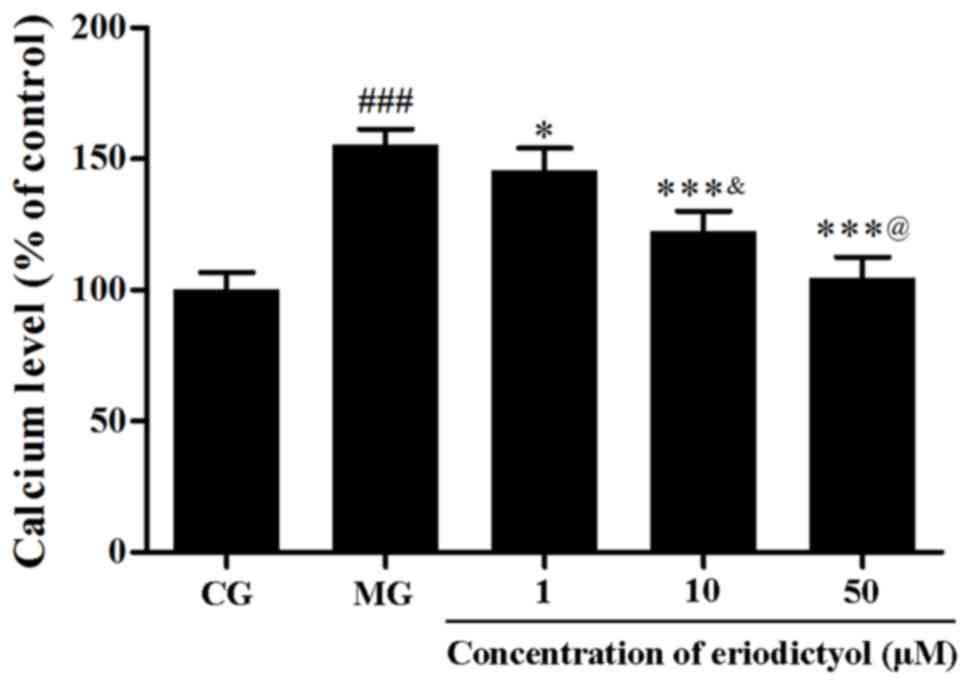
Effect of eriodictyol on intracellular
Ca2+
To assess the intracellular Ca2+ content,
the fluorescence probe Fluo-3 AM was used. Following the induction
of hypoxia/reoxygenation, the level of intracellular
Ca2+ was markedly elevated to 155.28±6.13% as compared
with the CG (P<0.001; Fig. 4).
However, in comparison with the model group, eriodictyol reduced
the overload of intracellular Ca2+ to 145.68±8.45,
122.41±7.64 and 102.39±8.17% upon pretreatment with 1, 10 and 50
µM, respectively (Fig. 4). These
results provided evidence that eriodictyol was able to reduce the
overload of intracellular Ca2+ in the
hypoxia/reoxygenation cell model.
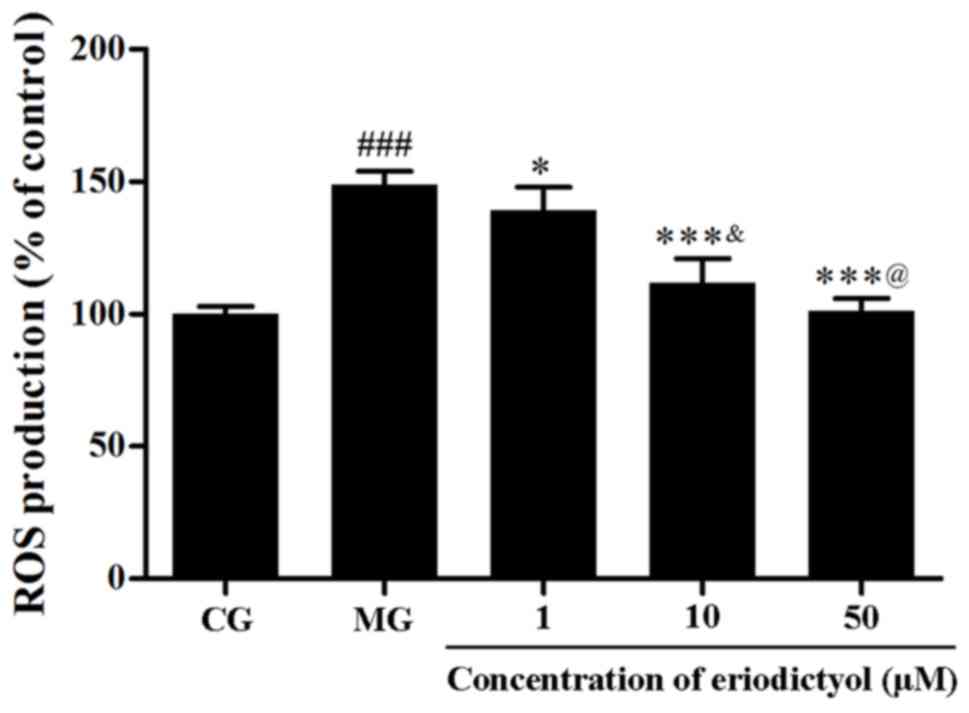
Effect of eriodictyol on ROS
generation
The intracellular ROS generation was determined
through the fluorescence intensity of DCFH-DA. The results
indicated that the relative fluorescence intensity in the MG
(148.84±5.16%) was significantly higher compared with that in the
CG (P<0.001). By contrast, when cells were treated with
eriodictyol, the fluorescence intensity was significantly decreased
to 139.25±8.68% (P<0.05), 111.89±9.10 and 101.33±4.57% (both
P<0.001) compared with the MG group (Fig. 5). The fluorescence intensity of 10 µM
(111.89±9.10%) was lower compared with 1 µM (139.25±8.68%;
P<0.05) and higher compared with 50 µM (101.33±4.57%;
P<0.05). This indicated that eriodictyol was able to
downregulate the generation of intracellular ROS.
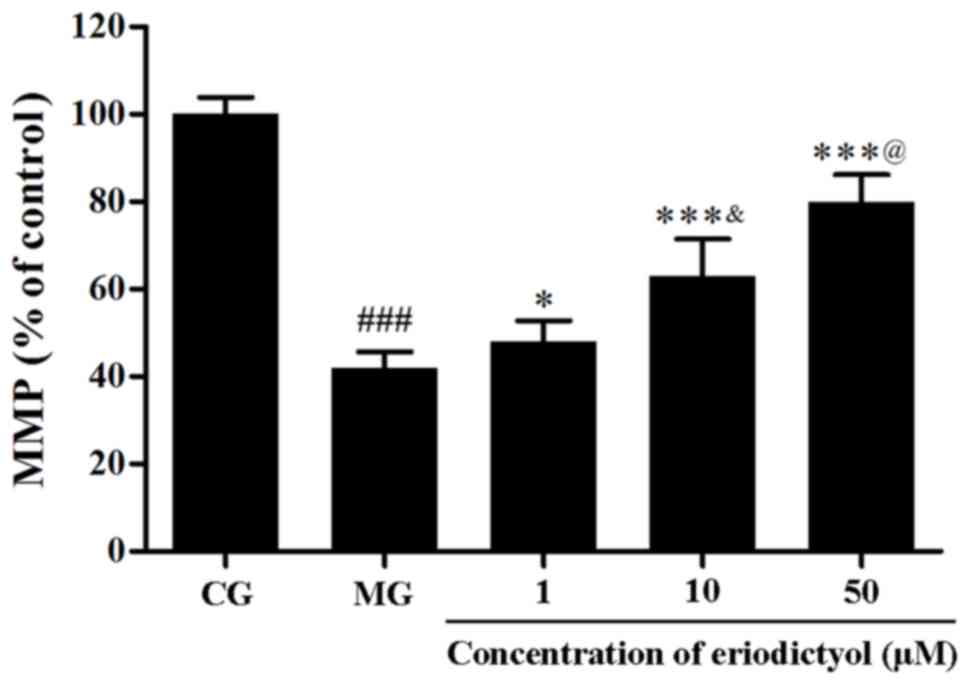
Effect of eriodictyol on MMP
In order to determine the MMP in H9c2
cardiomyocytes, the fluorescent probe JC-1 was used. Compared with
the CG, the fluorescence intensity in the MG (41.89±3.75%) was
significantly decreased (P<0.001), which indicated the collapse
of MMP in H9c2 cardiomyocytes following hypoxia/reoxygenation.
However, when pretreated with 1, 10 and 50 µM eriodictyol, the
fluorescence intensity was significantly elevated to 48.00±4.74%
(P<0.05), 62.92±8.56 and 79.89±6.24% (both P<0.001),
respectively, compared with the MG group (Fig. 6). Meanwhile, among these groups
treated with eriodictyol, the 10 µM group was significantly higher
than the 1 µM group lower than the 50 µM (both P<0.05). These
findings revealed that the collapse of MMP in
hypoxia/reoxygenation-treated H9c2 cardiomyocytes was attenuated by
eriodictyol.
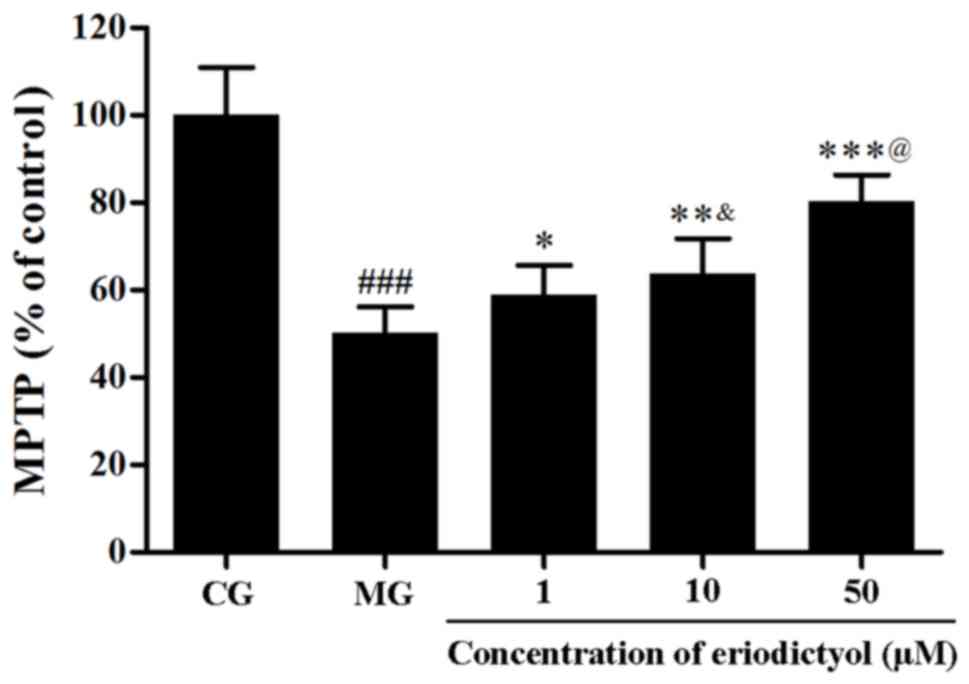
Effect of eriodictyol on the opening
of MPTP
The MPTP opening was evaluated through the
fluorescence intensity of free calcein in mitochondria. As shown in
Fig. 7, under hypoxia/reoxygenation,
the fluorescence intensity of mitochondrial calcein was
approximately half that of the CG (50.11±6.00%; P<0.001), which
demonstrated that the MPTP opened. Upon treatment with 1, 10 and 50
µM eriodictyol, the fluorescence intensity increased significantly
to 58.83±6.84% (P<0.05), 63.75±8.00% (P<0.01) and 80.40±5.92%
(P<0.001), respectively, compared to the MG group (Fig. 7). Compared with the CG group
(100±11.83%), the ATP level in the MG group (52.62±5.87%) was
significantly decreased (P<0.001). Eriodictyol (1, 10 and 50 µM)
pre-treatement increased ATP levels to 59.69±4.95% (P<0.05),
69.23±3.27% (P<0.01) and 80.52±9.52% (P<0.001), respectively,
compared with the MG group. These results implied that eriodictyol
inhibited the MPTP opening.
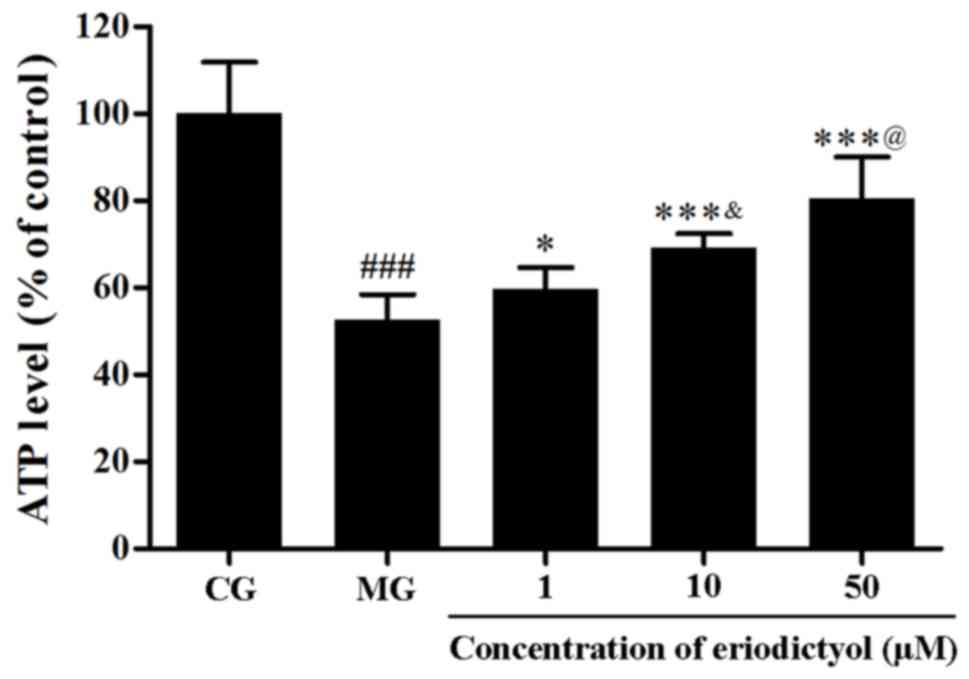
Effect of eriodictyol on ATP
depletion
The level of intracellular ATP represents the
function of mitochondria. To detect the depletion of intracellular
ATP, the firefly luciferase method was used. Hypoxia/reoxygenation
in H9c2 cardiomyocytes resulted in the decline of the intracellular
ATP level, whereas treatment with eriodictyol enhanced the ATP
level. The ATP level in the 10 µM eriodictyol group was
significantly higher than that in the 1 µM group as well as lower
than 50 µM group (both P<0.05; Fig.
8). Compared with the CG group (100±11.83%), the ATP level in
the MG group (52.62±5.87%) was significantly decreased
(P<0.001). Pretreated with eriodictyol (1, 10 and 50 µM),
compared with the MG group, the ATP levels were increased to
59.69±4.95% (P<0.05), 69.23±3.27% (P<0.001) and 80.52±9.52%
(P<0.001), respectively. These results indicated that
eriodictyol treatment was able to improve the depletion of
intracellular ATP in the cardiomyocytes.
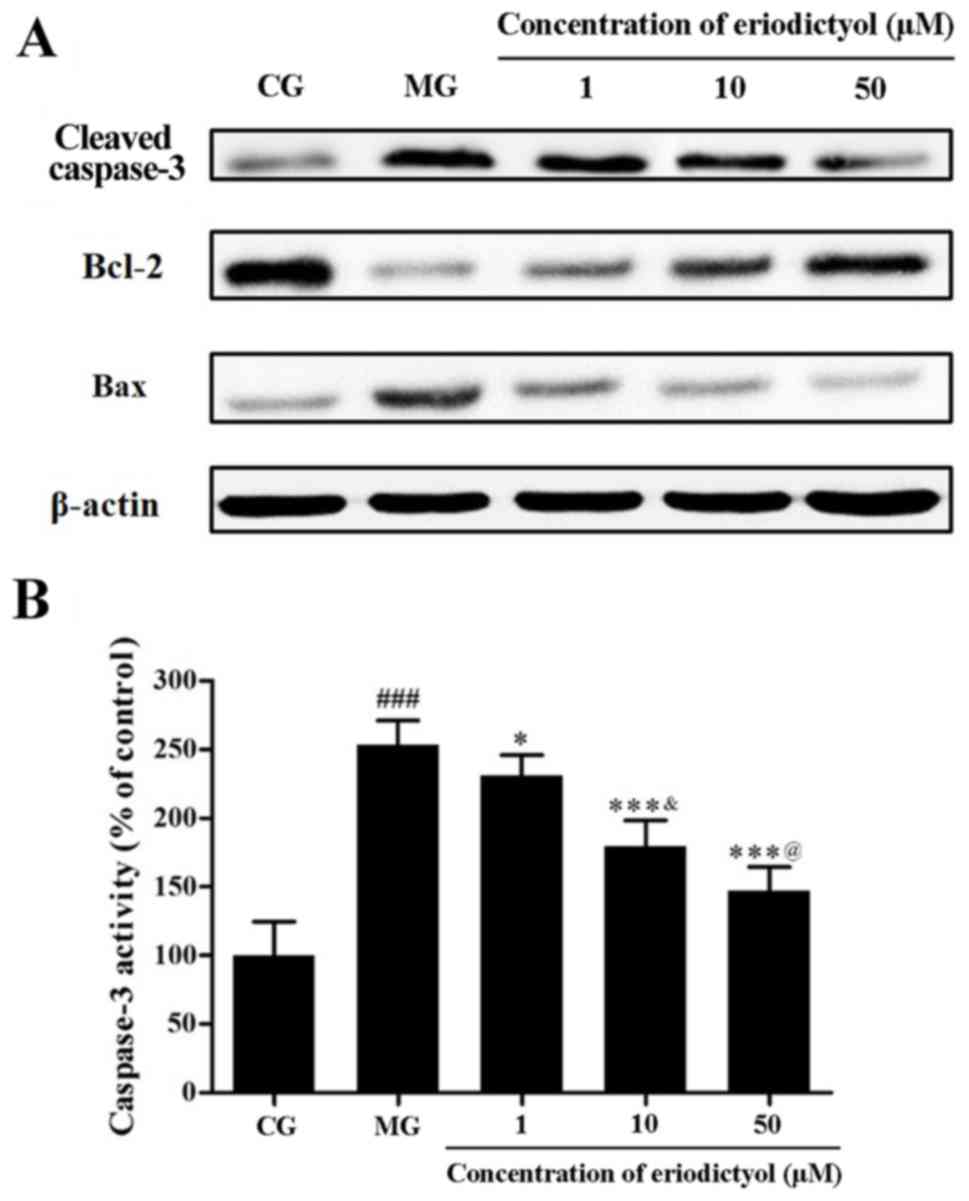
Effect of eriodictyol on caspase-3
activity and expression levels of caspase-3, Bcl-2 and Bax
As a member of the cysteinyl aspartate specific
protease family, caspase-3 serves a pivotal role in apoptosis
through hydrolyzed cleavage (28).
Western blot analysis revealed that hypoxia/reoxygenation promoted
the expression of caspase-3 in H9c2 cardiomyocytes, while
eriodictyol reduced this expression by different extents (Fig. 9A). In addition, colorimetric
detection further confirmed the activity of caspase-3
quantitatively. Compared with the control group, the caspase-3
activity was markedly elevated in H9c2 cardiomyocytes treated by
hypoxia/reoxygenation. However, in the presence of eriodictyol, the
increased activity of caspase-3 was inhibited accordingly.
Similarly, the activity of caspase-3 in the 10 µM eriodictyol group
was significantly lower than that in 1 µM group as well as higher
than 50 µM group (both P<0.05; Fig.
9B). The activity of caspase-3 in the MG group (253.38±17.80%)
was significantly decreased compared with the CG group
(100.00±24.74%; P<0.001). In contrast to the MG group, caspase-3
activity in the 1, 10 and 50 µM eriodictyol-treated groups was
significantly decreased to 230.80±15.03% (P<0.05), 179.40±18.85%
(P<0.001) and 147.09±17.41% (P<0.001), respectively.
Bcl-2 and Bax are the major members of the Bcl-2
protein family, which are involved in mitochondrion-mediated
apoptosis. The former demonstrates an anti-apoptotic effect,
whereas the latter exhibits a pro-apoptotic effect (29). In the present study, the expression
of Bcl-2 was downregulated by hypoxia/reoxygenation in contrast to
that in the control group. However, treatment with eriodictyol was
observed to upregulate Bcl-2 expression. Accordingly,
hypoxia/reoxygenation upregulated the expression of Bax, while
eriodictyol treatment suppressed this increased expression
(Fig. 9B).
Discussion
Myocardial ischemia and subsequent reperfusion is
the major cause of myocardial infarction. The oxygen deprivation
will lead to the breakdown of redox homeostasis and ROS
overproduction. As the major site of ROS production, mitochondria
serve an important role in the injury of myocardial ischemia and
reperfusion (30). In addition to
the overproduction of ROS, overload of intracellular
Ca2+ also affects the function of the mitochondria
(31). As the key determinant of
mitochondrial dysfunction, the MPTP will open (32). The electrochemical gradient across
the inner mitochondrial membrane (MMP) is necessary for
mitochondrial function (33).
Following the opening of MPTP, free solutes and proteins can be
distributed across the inner mitochondrial membrane and result in
the collapse of the MMP (34). The
dysfunction of mitochondria also leads to the depletion of ATP due
to MPTP opening (35), and finally
results in the cardiomyocyte apoptosis (36).
Caspases are cysteinyl aspartate specific proteases
with a central role in apoptosis, and their activation occurs
through cleavage at specific sites (37). As an effector enzyme, caspase-3 is
the key mediator responsible for promoting cell apoptosis (38). In addition, Bcl-2 and Bax are members
of the Bcl-2 protein family that participate in
mitochondrion-mediated apoptosis. Bcl-2 prevents apoptosis and
blocks the activation of caspase-3, while Bax promotes cell
apoptosis (39).
In the present study, mitochondrial dysfunction and
cell injury induced by hypoxia/reoxygenation were observed.
Pretreatment with eriodictyol increased the cell survival and
blocked the leakage of LDH from the cytosol, which indicates the
potential cardioprotective effect of eriodictyol. Further
experiments revealed that eriodictyol improved the dysfunction of
mitochondria through suppressing the overload of intracellular
Ca2+, preventing the overproduction of ROS, blocking the
opening of MPTP, increasing the MMP level and decreasing ATP
depletion. As important intracellular signaling molecules, there is
interplay between Ca2+ and ROS production.
Ca2+ may increase ROS production by enhancing metabolism
and ROS regulates Ca2+ homeostasis through reciprocal
redox (40). Meanwhile, the
interplay between ROS and Ca2+ triggers the opening of
MPTP opening, which leads to the collapse of MMP (41). In addition, as the main source of
ATP, mitochondria may fail to synthesize enough ATP to maintain
cellular function due to the dysfunction of MMP (42).
Furthermore, eriodictyol inhibited the apoptosis of
H9c2 cardiomyocytes through upregulating the expression of Bcl-2
and downregulating the expression levels of Bax and caspase-3, as
well as reducing the activity of caspase-3. As a bioactive
flavonoid, eriodictyol showed many protective effects through the
inhibition of oxidative stress (14,15,17,18). At
the same time, eriodictyol and its glucoside can protect against
cerebral ischemia injury in vitro and in vivo
(16,43). To the best of our knowledge, the
present study is the first to demonstrate the protective effects of
eriodictyol on cardiomyocytes injured by hypoxia/reoxygenation,
which indicates its potential application in the prevention of
myocardial ischemia and reperfusion injury.
In conclusion, the results of the present study
demonstrated the myocardial protective effects of eriodictyol and
relevant mechanisms in vitro. Eriodictyol can enhance the
survival of H9c2 cardiomyocytes injured by hypoxia/reoxygenation.
The mechanisms involve the improvement of mitochondrial dysfunction
and inhibition of apoptosis via the mitochondria-mediated signaling
pathway, including the upregulation of Bcl-2, downregulation of Bax
and inhibition of caspase-3. These results provided evidence for
further evaluations in vivo for the development of novel
therapeutic approaches for myocardial infarction.
References
|
1
|
Yellon DM and Hausenloy DJ: Realizing the
clinical potential of ischemic preconditioning and
postconditioning. Nat Clin Pract Cardiovasc Med. 2:568–575. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Yellon DM and Hausenloy DJ: Myocardial
reperfusion injury. New Eng J Med. 357:1121–1135. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Hausenloy DJ and Yellon DM: Targeting
myocardial reperfusion injury-the search continues. New Eng J Med.
373:1073–1075. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ong SB, Samangouei P, Kalkhoran SB and
Hausenloy DJ: The mitochondrial permeability transition pore and
its role in myocardial ischemia reperfusion injury. J Mol Cell
Cardiol. 78:23–34. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Hausenloya DJ, Duchenb MR and Yellon DM:
Inhibiting mitochondrial permeability transition pore opening at
reperfusion protects against ischaemia-reperfusion injury.
Cardiovasc Res. 60:617–625. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Pagliaro P, Moro F, Tullio F, Perrelli MG
and Penna C: Cardioprotective pathways during reperfusion: Focus on
redox signaling and other modalities of cell signaling. Antioxid
Redox Signal. 14:833–850. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zhao GL, Yu LM, Gao WL, Duan WX, Jiang B,
Liu XD, Zhang B, Liu ZH, Zhai ME, Jin ZX, et al: Berberine protects
rat heart from ischemia/reperfusion injury via activating
JAK2/STAT3 signaling and attenuating endoplasmic reticulum stress.
Acta Pharmacol Sin. 37:354–367. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Li Q, Shen L, Wang Z, Jiang HP and Liu LX:
Tanshinone IIA protects against myocardial ischemia reperfusion
injury by activating the PI3K/Akt/mTOR signaling pathway. Biomed
Pharmacother. 84:106–114. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Gao Y, Jia P, Shu W and Jia D: The
protective effect of lycopene on hypoxia/reoxygenation-induced
endoplasmic reticulum stress in H9C2 cardiomyocytes. Eur J
Pharmacol. 774:71–79. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
de Sousa LM, de Carvalho JL, da Silva HC,
Lemos TL, Arriaga AM, Braz-Filho R, Militão GC, Silva TD, Ribeiro
PR and Santiago GM: New cytotoxic bibenzyl and other constituents
from Bauhinia ungulata L. (Fabaceae). Chem Biodivers.
13:1630–1635. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Milella L, Milazzo S, De Leo M, Vera
Saltos MB, Faraone I, Tuccinardi T, Lapillo M, De Tommasi N and
Braca A: α-Glucosidase and α-amylase inhibitors from
arcytophyllum thymifolium. J Nat Prod. 79:2104–2112. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhong JD, Feng Y, Li HM, Xia XS and Li RT:
A new flavonoid glycoside from Elsholtzia bodinieri. Nat
Prod Res. 30:2278–2284. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zeng B, Chen K, Du P, Wang SS, Ren B, Ren
YL, Yan HS, Liang Y and Wu FH: Phenolic compounds from
Clinopodium chinense (Benth.) O. Kuntze and their inhibitory
effects on α-Glucosidase and vascular endothelial cells injury.
Chem Biodivers. 13:596–601. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lou H, Jing X, Ren D, Wei X and Zhang X:
Eriodictyol protects against H2O2-induced
neuron-like PC12 cell death through activation of Nrf2/ARE
signaling pathway. Neurochem Int. 61:251–257. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Jing X, Shi H, Zhu X, Wei X, Ren M, Han M,
Ren D and Lou H: Eriodictyol attenuates β-amyloid 25-35
peptide-induced oxidative cell death in primary cultured neurons by
activation of Nrf2. Neurochem Res. 40:1463–1471. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ferreira Ede O, Fernandes MY, Lima NM,
Neves KR, Carmo MR, Lima FA, Fonteles AA, Menezes AP and Andrade
GM: Neuroinflammatory response to experimental stroke is inhibited
by eriodictyol. Behav Brain Res. 312:321–332. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Li CZ, Jin HH, Sun HX, Zhang ZZ, Zheng JX,
Li SH and Han SH: Eriodictyol attenuates cisplatin-induced kidney
injury by inhibiting oxidative stress and inflammation. Eur J
Pharmacol. 772:124–130. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhu GF, Guo HJ, Huang Y, Wu CT and Zhang
XF: Eriodictyol, a plant flavonoid, attenuates LPS-induced acute
lung injury through its antioxidative and anti-inflammatory
activity. Exp Ther Med. 10:2259–2266. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Lee JK: Anti-inflammatory effects of
eriodictyol in lipopolysaccharide-stimulated Raw 264.7 murine
macrophages. Arch Pharm Res. 34:671–679. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Rossato MF, Trevisan G, Walker CI, Klafke
JZ, de Oliveira AP, Villarinho JG, Zanon RB, Royes LF, Athayde ML,
Gomez MV and Ferreira J: Eriodictyol: A flavonoid antagonist of the
TRPV1 receptor with antioxidant activity. Biochem Pharmacol.
81:544–551. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Habtemariam S and Dagne E: Comparative
antioxidant, prooxidant and cytotoxic activity of sigmoidin A and
eriodictyol. Planta Med. 76:589–594. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Walker J, Reichelt KV, Obst K, Widder S,
Hans J, Krammer GE, Ley JP and Somoza V: Identification of an
anti-inflammatory potential of Eriodictyon angustifolium compounds
in human gingival fibroblasts. Food Funct. 7:3046–3055. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ferreira PS, Spolidorio LC, Manthey JA and
Cesar TB: Citrus flavanones prevent systemic inflammation and
ameliorate oxidative stress in C57BL/6J mice fed high-fat diet.
Food Funct. 7:2675–2681. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Chen C, He H, Luo Y, Zhou M, Yin D and He
M: Involvement of Bcl-2 signal pathway in the protective effects of
apigenin on anoxia/reoxygenation-induced myocardium injury. J
Cardiovasc Pharmacol. 67:152–163. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ji HJ, Wang DM, Hu JF, Sun MN, Li G, Li
ZP, Wu DH, Liu G and Chen NH: IMM-H004, a novel courmarin
derivative, protects against oxygen-and
glucose-deprivation/restoration-induced apoptosis in PC12 cells.
Eur J Pharmacol. 723:259–266. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wang M, Sun GB, Zhang JY, Luo Y, Yu YL, Xu
XD, Meng XB, Zhang MD, Lin WB and Sun XB: Elatoside C protects the
heart from ischaemia/reperfusion injury through the modulation of
oxidative stress and intracellular Ca2+ homeostasis. Int
J Cardiol. 185:167–176. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Li JZ, Yu SY, Wu JH, Shao QR and Dong XM:
Paeoniflorin protects myocardial cell from doxorubicin-induced
apoptosis through inhibition of NADPH oxidase. Can J Physiol
Pharmacol. 90:1569–1575. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Shalini S, Dorstyn L, Dawar S and Kumar S:
Old, new and emerging functions of caspases. Cell Death Differ.
22:526–539. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Ding H, Han R, Chen X, Fang W, Liu M, Wang
X, Wei Q, Kodithuwakku ND and Li Y: Clematichinenoside (AR)
attenuates hypoxia/reoxygenation-induced H9c2 cardiomyocyte
apoptosis via a mitochondria-mediated signaling pathway. Molecules.
21(pii): E6832016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Madungwe NB, Zilberstein NF, Feng Y and
Bopassa JC: Critical role of mitochondrial ROS is dependent on
their site of production on the electron transport chain in
ischemic heart. Am J Cardiovasc Dis. 6:93–108. 2016.PubMed/NCBI
|
|
31
|
Hurst S, Hoek J and Sheu SS: Mitochondrial
Ca2+ and regulation of the permeability transition pore.
J Bioenerg Biomembr. 49:27–47. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Weiss JN, Korge P, Honda HM and Ping P:
Role of the mitochondrial permeability transition in myocardial
disease. Circ Res. 93:292–301. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Marchetti P, Castedo M, Susin SA, Zamzami
N, Hirsch T, Macho A, Haeffner A, Hirsch F, Geuskens M and Kroemer
G: Mitochondrial permeability transition is a central coordinating
event of apoptosis. J Exp Med. 184:1155–1160. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Kroemer G: Mitochondrial control of
apoptosis: An overview. Biochem Soc Symp. 66:1–15. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Li YY, Xiao L, Qiu LY, Yan YF, Wang H,
Duan GL, Liao ZP and Chen HP: Sasanquasaponin-induced
cardioprotection involves inhibition of mPTP opening via
attenuating intracellular chloride accumulation. Fitoterapia.
116:1–9. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Whelan RS, Kaplinskiy V and Kitsis RN:
Cell death in the pathogenesis of heart disease: Mechanisms and
significance. Annu Rev Physiol. 72:19–44. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Budihardjo I, Oliver H, Lutter M, Luo X
and Wang X: Biochemical pathways of caspase activation during
apoptosis. Annu Rev Cell Dev Biol. 15:269–290. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Uchiyama T, Otani H, Okada T, Ninomiya H,
Kido M, Imamura H, Nogi S and Kobayashi Y: Nitric oxide induces
caspase-dependent apoptosis and necrosis in neonatal rat
cardiomyocytes. J Mol Cell Cardiol. 34:1049–1061. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Youle RJ and Strasser A: The Bcl-2 protein
family: Opposing activities that mediate cell death. Nat Rev Mol
Cell Biol. 9:47–59. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Yan Y, Wei CL, Zhang WR, Cheng HP and Liu
J: Cross-talk between calcium and reactive oxygen species
signaling. Acta Pharmacol Sin. 27:821–826. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Görlach A, Bertram K, Hudecova S and
Krizanova O: Calcium and ROS: A mutual interplay. Redox Biol.
6:260–271. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Jašová M, Kancirová I, Waczulíková I and
Ferko M: Mitochondria as a target of cardioprotection in models of
preconditioning. J Bioenerg Biomembr. 49:357–368. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Jing X, Ren D, Wei X, Shi H, Zhang X,
Perez RG and Lou H and Lou H: Toxicol Appl Pharmacol. 273:672–679.
2013. View Article : Google Scholar : PubMed/NCBI
|