Introduction
Coronary heart disease (CHD) is the leading cause of
mortality in African-American men and women (1). Genetic factors have been demonstrated
to serve a significant role in the development of CHD (2). In addition, the involvement of
epigenetic modifications has been suggested in the development and
the progression of CHD (3–6). DNA methylation is a stable epigenetic
modification that results in the addition of a methyl group to the
5′ carbon of cytosine and primarily occurs at CpG dinucleotide
sequences in the mammalian genome (7). Notably, gene promoter hypermethylation
typically silences gene transcription, and aberrant gene
methylation has been indicated to be involved in the pathogenesis
of various diseases, including CHD (4,8,9) and type 2 diabetes (10,11).
Cyclin dependent kinase inhibitor 2B (CDKN2B)
is located on chromosome 9p21, which has been associated with CHD
in a number of genome-wide association studies (GWASs) (12,13).
CDKN2B encodes a cyclin-dependent kinase inhibitor that
regulates cell cycle G1 progression (14,15).
Cancer cells with hypermethylated CDKN2B are typically
associated with aberrantly accelerated proliferation (16). Atherosclerotic plaques are the major
contributing factor in CHD pathogenesis and are caused by
overproliferation of vascular smooth muscle cells and macrophages
(4,17). Notably, CDKN2B loss in mice
promoted atherosclerosis by increasing the size and complexity of
the lipid-laden necrotic core through impaired efferocytosis
(17). Furthermore, CDKN2B
has been suggested as a candidate gene of CHD (18,19).
CDKN2B hypermethylation has been indicated to be
significantly associated with the elevated expression of its
antisense noncoding RNA, antisense noncoding RNA in the INK4 locus
(ANRIL), and an increased risk of CHD (4). Several studies have revealed the
potential roles of CDKN2B in CHD (14,17,20).
Sex-specific associations have been indicated in
various aspects of CHD. For example, women have been demonstrated
to have a proportionally lower prevalence of disease and tend to
develop it later in life compared with men, and the difference of
incidence, development and surgical treatment of CHD between males
and females was indicated in previous studies (21–23).
Prior to menopause, women have relatively more protection against
CHD compared with men of the same age range (24–26). A
previous GWAS indicated sex differences in DNA methylation on 470
autosomal sites, including sites in CDKN2B (27). These epigenetic differences are
associated with differential mRNA and microRNA expression levels
and organ functions (27). In
addition, conventional cardiovascular pharmacological agents have
been indicated to induce their therapeutic effects on CHD through
various mechanisms, for example, by affecting serum levels of
vascular calcification inhibitors, which reduce cardiac workload
and increase coronary blood flow (28,29).
In the present study, a case-control study was
performed to investigate whether CDKN2B promoter methylation
contributes to the risk of CHD in a sex-dependent pattern, and
whether estrogen and conventional cardiovascular pharmacological
agents are able to recover CDKN2B expression by reversing
CDKN2B promoter methylation.
Materials and methods
Samples
CHD and non-CHD control samples were obtained from
patients at Ningbo First Hospital (Ningbo, China) between May 2008
and April 2010. A total of 36 CHD cases (18 males and 18 females,
mean age, 62.5±5.5) and 36 age- and sex-matched controls were
included in the present study. All individuals were Han Chinese
from Ningbo city in Eastern China diagnosed according to the World
Health Organization criteria (30).
The inclusion criteria utilized were as follows: Angiographic
evidence of >50% stenosis in one or more major coronary
arteries. Patients were excluded from the current study if they had
congenital heart disease, autoimmune disease, cardiomyopathy or
severe liver or kidney disease. All peripheral blood samples from
patients (5 ml) were collected in 3.2% citrate sodium-treated tubes
and stored at −80°C. The study protocol was approved by the Ethics
Committee in Ningbo First Hospital and all methods were performed
in accordance with the relevant guidelines and regulations.
Written, informed consent forms were obtained from all
subjects.
Bisulfite pyrosequencing
Human blood genomic DNA was extracted and quantified
as described previously (31). The
DNA methylation assay comprised of sodium bisulfite DNA conversion
(EpiTech Bisulfite kits; Qiagen AB, Sollentuna, Sweden), polymerase
chain reaction (PCR) amplification (Pyromark PCR kit; Qiagen) and
pyrosequencing (Pyromark Gold Q24 Reagents; Qiagen AB), which were
performed in accordance with the manufacturer's protocol. The
Pyromark PCR Master Mix was used in PCR amplification. PCR primers
were designed using PyroMark Assay Design software v2.0.1.15 (both
Qiagen). The sequences of the primers utilized were as follows:
CDKN2B forward, 5′-TAGGGGGAGGAGTTTAAGGGG-3′ and reverse,
5′-biotin-ACACTCTTCCCTTCTTTCC-3′; CDKN2B sequencing primer;
5′-GGGGTAGTGAGGATT-3′. The thermocycling conditions were as
follows: 1 cycle at 94°C for 15 sex, 45 cycles at 94°C for 20 sec,
58°C for 30 sec, 72°C for 60 sec and an extension stage at 72°C for
3 min.
Cell lines
Recent studies have suggested that CDKN2B is
associated with the occurrence and development of several types of
cancer (32,33). Notably, 293 cells are typically
applied to study the transforming and oncogenic properties of
cancer-associated genes as a model (34). Thus, 293 cell lines were selected for
the present study. Three cell lines, including 293 cells
(https://www.atcc.org/Products/All/CRL-11268.aspx),
human aortic endothelial cells (HAEC; http://www.atcc.org/Products/All/PCS-100-011.aspx) and
human primary coronary artery smooth muscle cells (HPCASMC;
http://www.atcc.org/Products/All/PCS-100-021.aspx)
were used in the present study. Cell lines were purchased from
American Type Culture Collection (Manassas, VA, USA) and were
cultured using dulbecco's modified eagle's medium (DMEM) with 10%
fetal bovine serum (FBS) and penicillin/streptomycin (Invitrogen;
Thermo Fisher Scientific, Inc., Waltham, MA, USA) in an incubator
at 37°C with 5% CO2 for 24 h.
Treatment with 5-aza-2′-deoxycytidine
(DAC), estrogen and cardiovascular pharmacological agents
Cells were cultured at a density of 1×106
cells/well in 6-well plates at 37°C for 24 h and the media (DMEM
with FBS and penicillin/streptomycin) was replaced following 4–8 h.
To determine the potential regulatory roles of DNA methylation in
CDKN2B gene transcription, HAEC, HPCASMC and 293 cells were
treated with DAC (Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) at
0.5, 1.0 and 2.0 µM at 37°C for 3 days. Following this incubation
step, total RNA was isolated from the cells and subjected to
reverse transcription. To examine the effects of estrogen
treatment, the cells were treated with estrogen (17-β-estradiol;
Sigma-Aldrich; Merck KGaA) at different concentrations (10, 100 and
1,000 nM), and the total RNA and genomic DNA were isolated. In
aformentioned comparisons, these three cell lines with the EtOH
treatment were considered as the control. In addition, 10 µM
simvastatin, 10 µM trimetazidine dihydrochloride and 50 µM
γ-carboxy-L-glutamic acid (all Sigma-Aldrich; Merck KGaA) were used
to treat HAEC and HPCASMC for 1, 6, 12 and 24 h. Notably,
γ-carboxy-L-glutamic acid is an effective ingredient of isosorbide
mononitrate, which is a common cardiovascular drug (35,36).
Following treatments, total RNA and genomic DNA were extracted from
cells. 293 cells were not subjected to treatment as HAEC and
HPCASMC were more specific to cardiovascular disease and thus would
produce more meaningful results.
Total RNA extraction and reverse
transcription-quantitative PCR (RT-qPCR)
TRIzol reagent (Invitrogen; Thermo Fisher
Scientific, Inc.) was used for total RNA isolation. Subsequently, 1
µg RNA was treated with a High Capacity cDNA Reverse Transcription
kit (Applied Biosystems, Thermo Fisher Scientific, Inc.) for 2 h at
37°C for cDNA synthesis according to the manufacturer's protocol.
qPCR was performed using SYBR green master PCR mix (Applied
Biosystems, Thermo Fisher Scientific, Inc.). CDKN2B
transcription was normalized to GAPDH transcription levels
and the PCR products were quantified using the 2−ΔΔCq
method (37). The thermocycling
conditions were as follows: denaturation at 94°C for 5 min,
followed by 40 cycles of amplification (30 sec of denaturation at
94°C, 30 sec of annealing at 58°C and 30 sec of extension at 72°C).
The following primers were used: CDKN2B forward,
5′-GTTAAGTTTACGGCCACCGG-3′ and reverse, 5′-ACCTTCTCCACTAGTCCC-3′;
and GAPDH forward, 5′-TGGTATGGAAGGACTCA-3′ and reverse,
5′-CCAGTAGAGGCAGGGATGAT-3′.
Bisulfite sequencing
Genomic DNA was isolated from cells with or without
estrogen (10, 100 and 1,000 nM) and cardiovascular drug treatments
(10 µM simvastatin, 10 µM trimetazidine dihydrochloride and 50 µM
γ-carboxy-L-glutamic acid) using a genomic DNA isolation kit
(Qiagen, Inc., Valencia, CA, USA). Bisulfite conversion reagents
(EpiTect Bisulfite kit; Qiagen, Inc.) were used to convert genomic
DNA. Products were amplified with the following primers for
CDKN2B promoter: CDKN2B_F,
5′-TTGGTTTAGTTGAAAAYGGAATT-3′; and CDKN2B_R,
5′-AACRCCTAACRCRAACRCAACC-3′. PCR was performed as follows: 95°C
for 5 min; followed by 35 cycles of 95°C for 30 sec, 52°C for 30
sec, 72°C for 30 sec, with a final extension of 2 min at 72°C. PCR
products were cloned using a commercial CloneJET PCR Cloning kit
(Thermo Fisher Scientific, Inc.) according to manufacturer's
instructions. A total of 15–20 independent bacterial clones were
isolated and sequenced for each PCR fragment to calculate the
cytosine methylation levels.
Luciferase reporter gene assays
The CDKN2B promoter fragment containing seven
CpG sites was generated using PCR as aforementioned, and the primer
sequences of CDKN2B were as follows: Forward,
5′-GGGGCAGTGAGGACT-3′ and reverse, 5′-GCCTGGATTGCTTCT-3′. The
subsequent PCR product was cloned into pCR2.1 (included in the kit)
using a T-A Cloning Kit (Invitrogen; Thermo Fisher Scientific,
Inc.) and sequenced. Plasmids containing CDKN2B promoter
region were amplified and digested with XhoI and KpnI
(New England Biolabs, Ipswich, MA, USA). The target DNA fragment
containing the reporter gene was cloned into pGL3-Luciferase
reporter plasmid (Promega Corporation, Madison, WI, USA).
Subsequently, constructed pGL3-CDKN2B-Luciferase plasmid was
transfected into 293 cells using Lipofectamine 2000 Reagent
(Invitrogen; Thermo Fisher Scientific, Inc.) according to
manufacturer's protocol. A total of 24 h following transfection,
cells were lysed with Tropix lysis buffer (Applied Biosystems;
Thermo Fisher Scientific, Inc.). Luciferase and β-galactosidase
activities were measured using a Luciferase Assay System (Applied
Biosystems; Thermo Fisher Scientific, Inc.) according to
manufacturer's protocol. β-Galactosidase activity was used to
normalize transfection efficiency.
Statistical analysis
SPSS package software (version 16.0; SPSS, Inc.,
Chicago, IL, USA) was used to determine the association between
CDKN2B promoter methylation with CHD and various biochemical
factors. Comparisons of CDKN2B methylation were performed
using GraphPad Prism 5 Software (GraphPad Software, Inc., La Jolla,
CA, USA). Correlations of CDKN2B methylation with age and
biochemical indicators were performed using R statistical software
(R V.3.3.2; http://www.r-project.org/). All P-values were adjusted
for age, smoking, diabetes and hypertension. Relative expression
data were presented as the mean ± standard deviation. Statistical
analysis for RT-qPCR data was performed using one-way analysis of
variance followed by a Dunnett's post hoc test. P<0.05 was
considered to indicate a statistically significant difference.
Results
Significant association of CDKN2B
promoter methylation with CHD in women
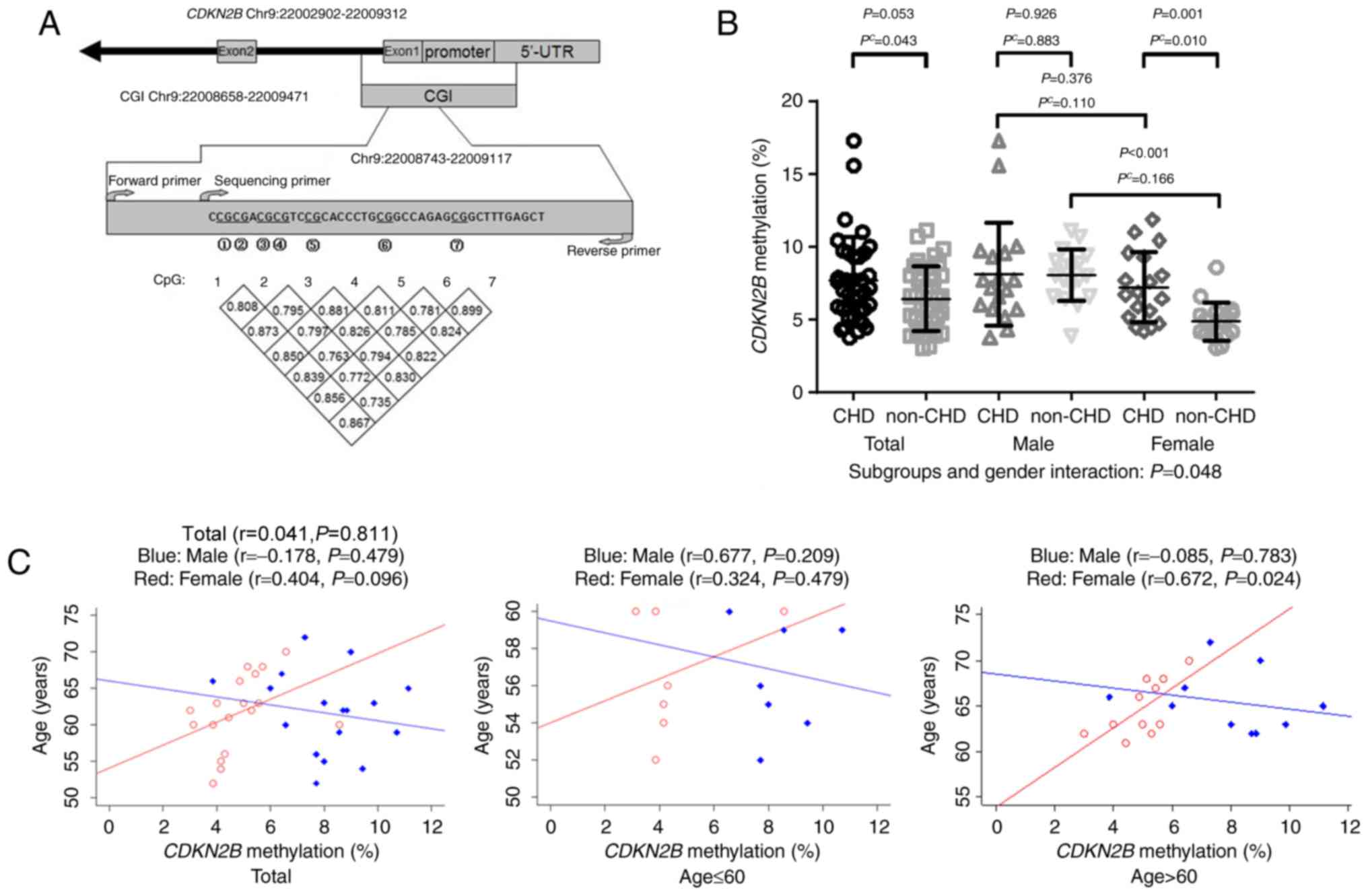
The bisulfite pyrosequencing assay was used to
measure a total of seven CpGs on the CDKN2B promoter among
36 cases and 36 matched controls. Significant correlations were
observed among DNA methylation levels of seven CpGs (r>0.7,
Fig. 1A). Subsequently CDKN2B
methylation was represented as the mean methylation level of the
seven CpGs. As indicated in Table I
and Fig. 1B, significantly increased
CDKN2B methylation levels (7.66±3.02%) were observed in
patients with CHD compared with non-CHD subjects (6.43±2.23%,
adjusted P=0.043). In addition, a significant association of CHD
with sex was indicated regarding CDKN2B methylation
(adjusted P=0.048; Fig. 1B). Further
analysis demonstrated a significant female-specific association of
CDKN2B methylation with CHD [women with CHD (7.21±2.40%)
compared with women without CHD (4.83±1.31%), adjusted P=0.010;
Fig. 1B]. However, there was no
significant correlation between CDKN2B methylation and age
in the whole cohort (total, r=0.041, adjusted P=0.811; Fig. 1C), further analysis regarding age
indicated a significant association with CHD in women >60 years
old (women, r=0.672, adjusted P=0.024; Fig. 1C).
 | Table I.Comparison of cyclin dependent kinase
inhibitor 2B methylation levels within subgroups. |
Table I.
Comparison of cyclin dependent kinase
inhibitor 2B methylation levels within subgroups.
|
| DNA methylation
(%) |
|
|
|---|
|
|
|
|
|
|---|
| Variable | CHD (n=36) mean ±
SD | Non-CHD (n=36) mean
± SD | P-value | Sex subgroup
interaction P-value |
|---|
| DNA methylation
site |
| CpG
1 | 8.89±3.97 | 7.44±2.99 | 0.066 | 0.058 |
| CpG
2 | 5.94±2.03 | 5.14±1.78 | 0.042 | 0.239 |
| CpG
3 | 6.11±2.81 | 4.61±1.68 | 0.012 | 0.088 |
| CpG
4 | 4.83±2.25 | 4.00±1.64 | 0.057 | 0.168 |
| CpG
5 | 9.19±4.03 | 7.97±3.07 | 0.149 | 0.278 |
| CpG
6 | 9.03±3.19 | 8.11±2.97 | 0.142 | 0.005 |
| CpG
7 | 9.61±4.09 | 7.72±3.53 | 0.031 | 0.023 |
| Mean ± SD DNA
methylation (%) | 7.66±3.02 | 6.43±2.23 | 0.043 | 0.048 |
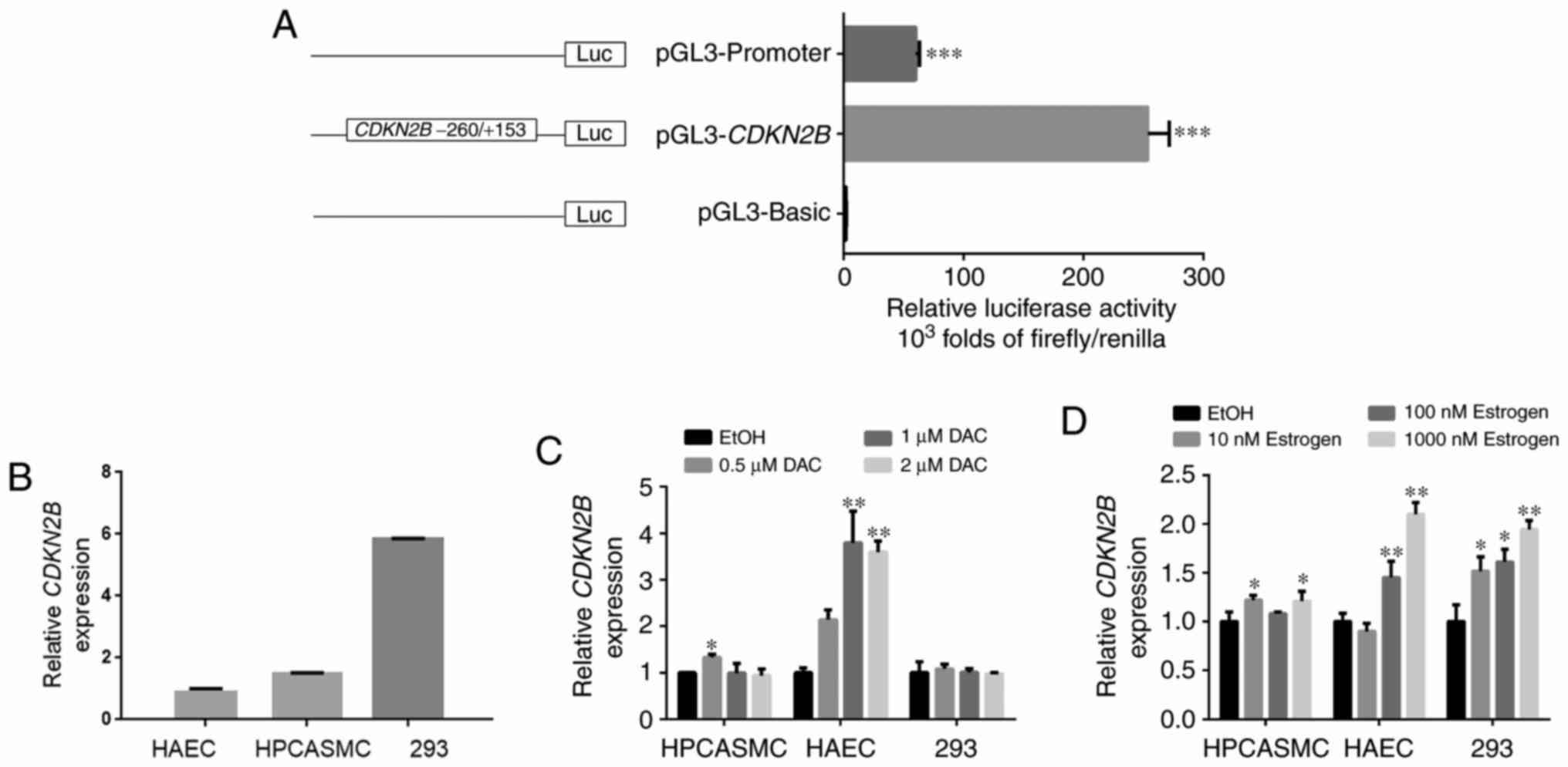
CDKN2B promoter fragment enhances
luciferase gene activity
The dual-luciferase reporter assay is a standard
method that utilizes Firefly and Renilla luciferase to explore the
promoter activity of target fragment (38). The CDKN2B promoter fragment
containing seven CpG sites was amplified and cloned into pGL3
luciferase plasmid in the present study. The construct was
transfected into 293 cells. Notably, HPCASMC and HAEC were
difficult transfect and were therefore excluded from this
experiment. Results suggested that the CDKN2B promoter
fragment significantly enhanced the luciferase activity by >200
fold compared with pGL3-Basic in 293 cells (P<0.001; Fig. 2A). The present results indicated that
CDKN2B promoter enhanced the luciferase activity in 293
cells.
Methylation inhibitor DAC enhances
CDKN2B gene transcription
Endogenous CDKN2B transcription levels were
determined in HPCASMC, HAEC and 293 cells (Fig. 2B). Furthermore, cell lines were
incubated with different concentrations of DAC, a DNA
methyltransferase inhibitor. Results revealed that DAC (0.5 µM/l)
significantly upregulated CDKN2B transcription levels compared with
EtOH treatment in HPCASMC (P<0.05; Fig. 2C). In addition, DAC (1 µM/l and 2
µM/l) significantly upregulated CDKN2B transcription levels
compared with EtOH in HAEC (1 µM/l, P<0.01 and 2 µM/l,
P<0.01; Fig. 2C). However, no
significant differences in CDKN2B transcription levels were
detected in 293 cells, which may have been due to the high
endogenous CDKN2B expression. In light of these findings, it
was speculated that DNA methylation may serve an important role in
the regulation of CDKN2B transcription in HAEC and
HPCASMC.
Estrogen increases CDKN2B
transcription and alters its promoter methylation
Cells were incubated with different concentrations
of estrogen. Results suggested that estrogen significantly
upregulated CDKN2B transcription levels in HAEC and 293
cells compared with EtOH treatment (HAEC, 100 nM/l:
P<0.01 and 1,000 nM/l: P<0.01; 293, 10 nM/l:
P<0.05, 100 nM/l: P<0.05 and 1,000 nM/l: P<0.01; Fig. 2D). Specific concentrations of
estrogen also significantly increased CDKN2B transcription
levels in HPCASMC (HPCASMC, 10 nM/l: P=0.015 and 1,000 nM/l:
P=0.030; Fig. 2D).
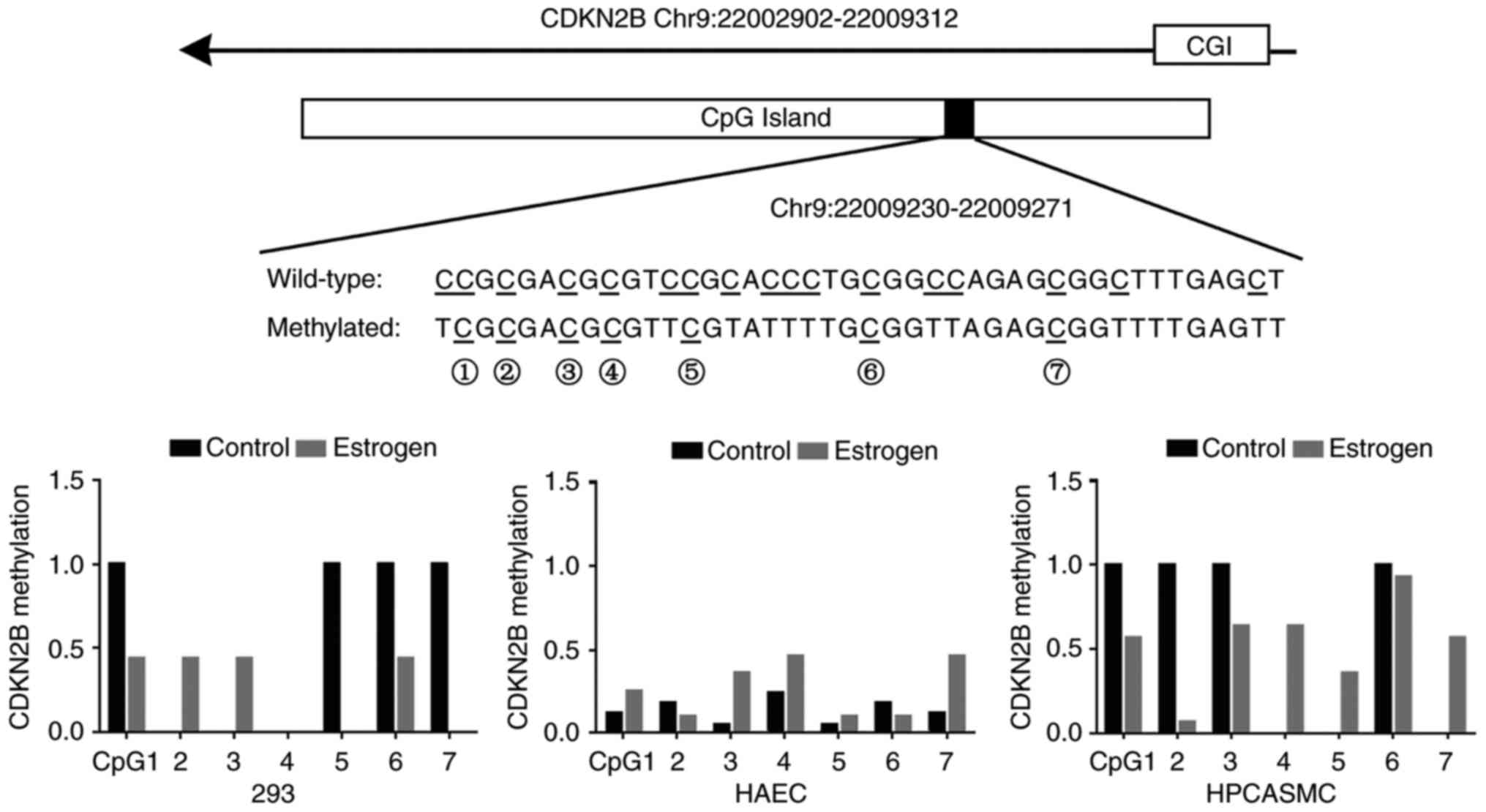
Methylation levels of seven CDKN2B CpGs were
varied in HAEC, HPCASMC and 293 cells. There were four fully
methylated CpGs (CpG-1, 5, 6 and 7) and three unmethylated CpGs
(CpG-2, 3 and 4) in 293 cells, and four fully methylated CpGs
(CpG-1, 2, 3 and 6) and three unmethylated CpGs (CpG-4, 5 and 7) in
HPCASMC. The methylation levels of the seven CpGs were <0.3 in
HAEC (Fig. 3). In some cases,
estrogen treatment reduced the methylation levels of previously
hypermethylated CpGs and increased the methylation levels of
previously hypomethylated CpGs in HPCASMC and 293 cells (Fig. 3). Notably, estrogen treatment was
able to increase the methylation levels of the majority of CpG
sites in HAEC.
Although estrogen treatment did not result in a
similar methylation pattern among the three cell lines, the results
suggested that estrogen was able to increase CDKN2B gene
transcription among the three cell lines. The findings indicated
that estrogen may increase CDKN2B transcription by altering
CDKN2B methylation.
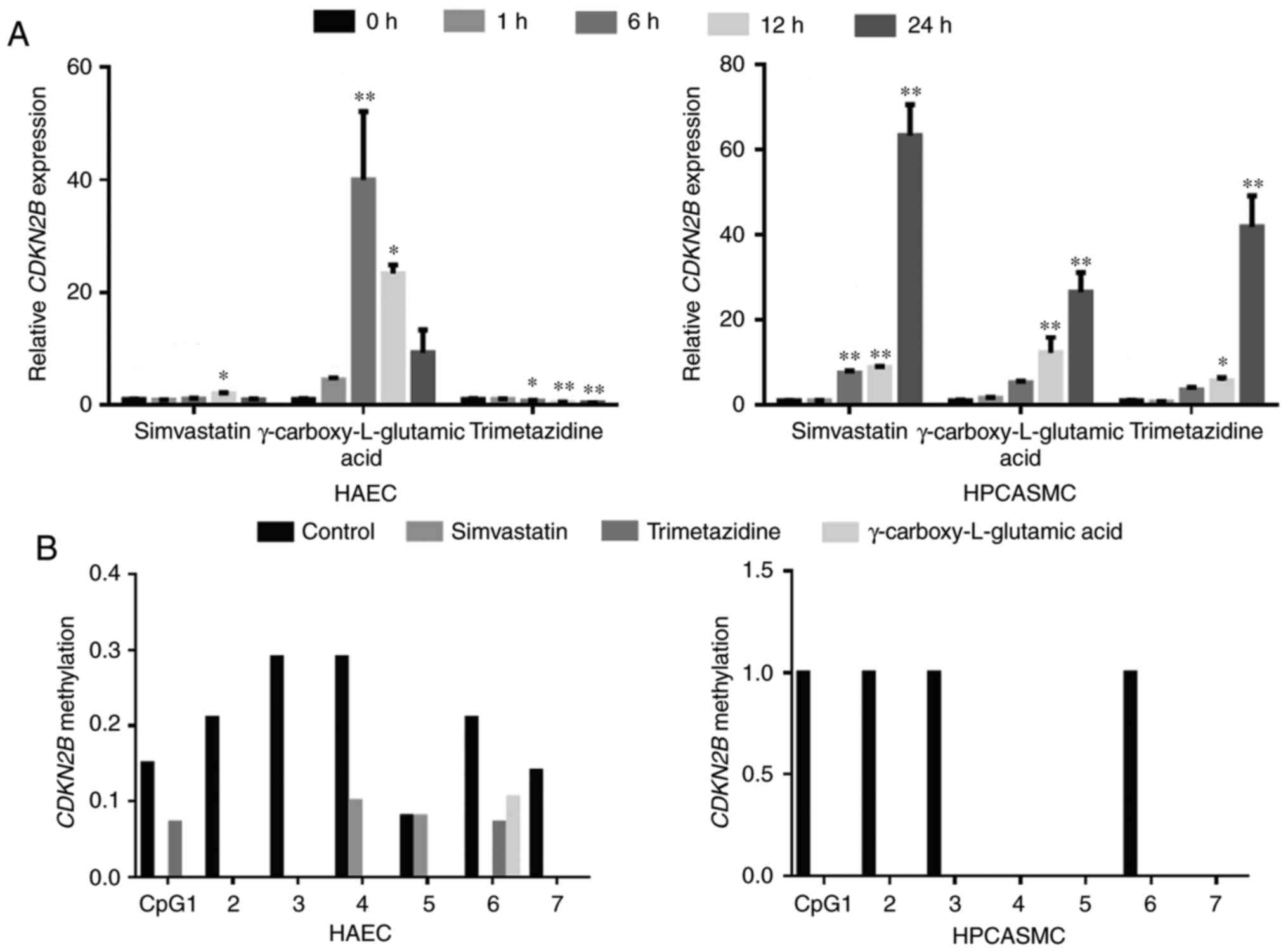
Cardiovascular pharmacological agents
increase CDKN2B transcription and reduce CDKN2B promoter
methylation concomitantly
Three cardiovascular pharmacological agents,
including simvastatin, γ-carboxy-L-glutamic acid and trimetazidine
dihydrochloride, were selected to explore their regulatory effects
on CDKN2B. Results revealed that 24-h treatment with these
agents significantly altered CDKN2B transcription levels in
HPCASMC and HAEC, with the exception of HAEC treated with
simvastatin and γ-carboxy-L-glutamic acid (P<0.05 and P<0.01;
Fig. 4A). Notably, CDKN2B
transcription levels in HPCASMC increased 63.3-fold following 24 h
treatment with simvastatin, 26.5-fold following 24 h treatment with
γ-carboxy-L-glutamic acid and 41.9-fold following 24 h treatment
with trimetazidine dihydrochloride, respectively (Fig. 4A). CDKN2B transcription levels
in HAEC increased 2.0-fold following 12 h treatment with
simvastatin and 40.0-fold following 6 h treatment with
γ-carboxy-L-glutamic acid treatment, respectively (Fig. 4A). Conversely, CDKN2B
transcription levels significantly decreased 2.2-fold following 24
h trimetazidine dihydrochloride treatment in HAEC (P<0.01;
Fig. 4A). Methylation assays
demonstrated that the cardiovascular pharmacological agents reduced
CDKN2B methylation levels in HAEC and HPCASMC (Fig. 4B). Treatment with simvastatin,
γ-carboxy-L-glutamic acid and trimetazidine dihydrochloride reduced
CDKN2B promoter methylation and increased CDKN2B
transcription concomitantly in HPCASMC. Additionally, treatment
with simvastatin and γ-carboxy-L-glutamic acid reduced
CDKN2B promoter methylation and increased CDKN2B
transcription concomitantly in HAEC.
Discussion
In the present study, it was identified that there
was a female-specific association of elevated CDKN2B
promoter methylation with CHD. Subsequent functional experiments
indicated that CDKN2B promoter methylation is important for
gene expression, which was further demonstrated to be susceptible
to estrogen and conventional cardiovascular pharmacological
agents.
CDKN2B has been suggested to regulate
efferocytosis and atherosclerosis (39). The deletion of CDKN2B promotes
the advanced development of atherosclerotic plaques (40). Furthermore, CDKN2B expression
is reduced in atherosclerotic plaques, indicating that
CDKN2B serves an essential role in the formation of
atherosclerotic plaques (39). In
the present study, the CDKN2B promoter fragment was able to
significantly enhance luciferase reporter gene activity.
Furthermore, methylation inhibitor DAC increased CDKN2B
transcription levels. These results suggest that CDKN2B
promoter methylation may be important in the regulation of
CDKN2B gene function.
Sex dimorphism of CHD has been observed in the
prevalence and the onset age of CHD (21,41).
Notably, the onset age of CHD in women is typically 10 years later
than that in men (21). Furthermore,
non-obstructive CHD and angina are more frequently identified in
women than in men (42,43). Previous results suggested an
interaction between sex and age impacted DNA methylation (44,45).
Furthermore, age has been indicated to increase the risk of CHD in
women compared with men (46). CHD
risk in men plateaus at the age of 45–50, whereas in women, CHD
risk continues to increase sharply until the age of 60–65 (47). In the present study, a
female-specific association of elevated CDKN2B promoter
methylation was indicated with the risk of CHD. Further analysis by
age suggested that the women >60 years of age had significantly
higher CDKN2B promoter methylation levels.
Studies have indicated that estrogen may inhibit
atherosclerotic plaque progression and vasodilation through its
anti-oxidative and anti-inflammatory properties (21,43,48).
Furthermore, it has been identified that estrogen may influence
neoplastic diseases via its effects on the levels of gene
expression and DNA methylation (49,50). In
the present study, CDKN2B transcription levels were
upregulated following estrogen treatment in HPCASMC, HAEC and 293
cells, in addition with promoter cytosine modifications in the
promoter region of CDKN2B. The present findings suggest that
estrogen may exert its regulatory role through promoter methylation
modification.
Cardiovascular pharmacological agents include
angiotensin-converting enzyme inhibitors, nitrates, statins and
β-adrenergic blockers (51–54). Abnormal gene methylation may
influence the curative effect of various kinds of drugs, including
anti-tumor drugs and chemotherapeutic drugs (55–57). In
the present study, treatment with three types of cardiovascular
pharmacological agents increased CDKN2B transcription levels
and reduced CDKN2B methylation levels concomitantly in
HPCASMC. Similar results were identified in HAEC, except
trimetazidine was demonstrated to decrease CDKN2B
transcription in HAEC and further study should be performed to
verify this result. This suggests that these agents may deliver
their effects through CDKN2B gene silencing.
In conclusion, the present study demonstrated the
role of DNA methylation in the regulation of CDKN2B
transcription and that CDKN2B transcription may be affected
by estrogen and cardiovascular pharmacological agents. Furthermore,
the present results provided an improved understanding of the
mechanisms by which CDKN2B may contribute to the risk of CHD
in women. Due to the moderate sample size, future studies with
extended samples are required to assess the significant association
of CDKN2B promoter methylation with CHD in females.
Acknowledgements
Not applicable.
Funding
The present study was supported by grants from the
National Natural Science Foundation of China (grant no. 81371469),
the Natural Science Foundation of Zhejiang Province (grant no.
LR13H020003) and from K. C. Wong Magna Fund in Ningbo University
(Ningbo, Zhejiang, China).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
XC and SD conceived and designed the current study.
YY, HJ, CZ, BL, XuX, XiX, NW, YX, JL and YS performed the
experiments. DJ, LX, LH, YH, DL and YX performed the analyses. XC,
DJ, LX and HH prepared all figures and tables. XC wrote the
manuscript. All the authors reviewed the manuscript and agreed to
its publication.
Ethics approval and consent to
participate
The protocol of the current study was approved by
the Ethics Committee of Ningbo First Hospital (Ningbo, China) and
all methods were performed in accordance with the relevant
guidelines and regulations.
Patient consent for publication
Written informed consent was obtained from all
patients.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
CHD
|
coronary heart disease
|
|
DAC
|
5-aza-2′-deoxycytidine
|
|
GWAS
|
genome-wide association study
|
|
PCR
|
polymerase chain reaction
|
|
HAEC
|
human aortic endothelial cells
|
|
HPCASMC
|
human primary coronary artery smooth
muscle cells
|
References
|
1
|
Lettre G, Palmer CD, Young T, Ejebe KG,
Allayee H, Benjamin EJ, Bennett F, Bowden DW, Chakravarti A,
Dreisbach A, et al: Genome-wide association study of coronary heart
disease and its risk factors in 8,090 African Americans: The NHLBI
CARe Project. PLoS Genet. 7:e10013002011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Katzmarzyk PT, Perusse L, Rice T, Gagnon
J, Skinner JS, Wilmore JH, Leon AS, Rao DC and Bouchard C: Familial
resemblance for coronary heart disease risk: The HERITAGE family
study. Ethn Dis. 10:138–147. 2000.PubMed/NCBI
|
|
3
|
Loscalzo J and Handy DE: Epigenetic
modifications: Basic mechanisms and role in cardiovascular disease
(2013 Grover Conference series). Pulm Circ. 4:169–174. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Zhuang J, Peng W, Li H, Wang W, Wei Y, Li
W and Xu Y: Methylation of p15INK4b and expression of ANRIL on
chromosome 9p21 are associated with coronary artery disease. PLoS
One. 7:e471932012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Baccarelli A, Rienstra M and Benjamin EJ:
Cardiovascular epigenetics: Basic concepts and results from animal
and human studies. Circ Cardiovasc Genet. 3:567–573. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Turunen MP, Aavik E and Ylä-Herttuala S:
Epigenetics and atherosclerosis. Biochim Biophys Acta.
1790:886–891. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Feinberg AP: Phenotypic plasticity and the
epigenetics of human disease. Nature. 447:433–440. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Friso S, Lotto V, Choi SW, Girelli D,
Pinotti M, Guarini P, Udali S, Pattini P, Pizzolo F, Martinelli N,
et al: Promoter methylation in coagulation F7 gene influences
plasma FVII concentrations and relates to coronary artery disease.
J Med Genet. 49:192–199. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Guay SP, Brisson D, Munger J, Lamarche B,
Gaudet D and Bouchard L: ABCA1 gene promoter DNA methylation is
associated with HDL particle profile and coronary artery disease in
familial hypercholesterolemia. Epigenetics. 7:464–472. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Dayeh TA, Olsson AH, Volkov P, Almgren P,
Ronn T and Ling C: Identification of CpG-SNPs associated with type
2 diabetes and differential DNA methylation in human pancreatic
islets. Diabetologia. 56:1036–1046. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Yang M, Sun JZ, Sun YL, You W, Dai J and
Li GS: Association between leptin gene promoter methylation and
type 2 diabetes mellitus. Zhonghua Yi Xue Yi Chuan Xue Za Zhi.
29:474–477. 2012.(In Chinese). PubMed/NCBI
|
|
12
|
Samani NJ, Erdmann J, Hall AS,
Hengstenberg C, Mangino M, Mayer B, Dixon RJ, Meitinger T, Braund
P, Wichmann HE, et al: Genomewide association analysis of coronary
artery disease. N Eng J Med. 357:443–453. 2007. View Article : Google Scholar
|
|
13
|
Wellcome Trust Case Control Consortium, .
Genome-wide association study of 14,000 cases of seven common
diseases and 3,000 shared controls. Nature. 447:661–678. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Pilbrow AP, Folkersen L, Pearson JF, Brown
CM, McNoe L, Wang NM, Sweet WE, Tang WH, Black MA, Troughton RW, et
al: The chromosome 9p21.3 coronary heart disease risk allele is
associated with altered gene expression in normal heart and
vascular tissues. PLoS One. 7:e395742012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Soto JL, Cabrera CM, Serrano S and
López-Nevot MA: Mutation analysis of genes that control the G1/S
cell cycle in melanoma: TP53, CDKN1A, CDKN2A, and CDKN2B. BMC
Cancer. 5:362005. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Herman JG and Baylin SB: Gene silencing in
cancer in association with promoter hypermethylation. N Eng J Med.
349:2042–2054. 2003. View Article : Google Scholar
|
|
17
|
Motterle A, Pu X, Wood H, Xiao Q, Gor S,
Ng FL, Chan K, Cross F, Shohreh B, Poston RN, et al: Functional
analyses of coronary artery disease associated variation on
chromosome 9p21 in vascular smooth muscle cells. Hum Mol Genet.
21:4021–4029. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
McPherson R, Pertsemlidis A, Kavaslar N,
Stewart A, Roberts R, Cox DR, Hinds DA, Pennacchio LA,
Tybjaerg-Hansen A, Folsom AR, et al: A common allele on chromosome
9 associated with coronary heart disease. Science. 316:1488–1491.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Helgadottir A, Thorleifsson G, Manolescu
A, Gretarsdottir S, Blondal T, Jonasdottir A, Jonasdottir A,
Sigurdsson A, Baker A, Palsson A, et al: A common variant on
chromosome 9p21 affects the risk of myocardial infarction. Science.
316:1491–1493. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Jarinova O, Stewart AF, Roberts R, Wells
G, Lau P, Naing T, Buerki C, McLean BW, Cook RC, Parker JS and
McPherson R: Functional analysis of the chromosome 9p21.3 coronary
artery disease risk locus. Arterioscler Thromb Vasc Biol.
29:1671–1677. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Lawton JS: Sex and gender differences in
coronary artery disease. Semin Thorac Cardiovasc Surg. 23:126–130.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Sbarouni E, Georgiadou P and Voudris V:
Gender-specific differences in biomarkers responses to acute
coronary syndromes and revascularization procedures. Biomarkers.
16:457–465. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Hu G, Jousilahti P, Qiao Q, Peltonen M,
Katoh S and Tuomilehto J: The gender-specific impact of diabetes
and myocardial infarction at baseline and during follow-up on
mortality from all causes and coronary heart disease. J Am Coll
Cardiol. 45:1413–1418. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Post WS, Goldschmidt-Clermont PJ, Wilhide
CC, Heldman AW, Sussman MS, Ouyang P, Milliken EE and Issa JP:
Methylation of the estrogen receptor gene is associated with aging
and atherosclerosis in the cardiovascular system. Cardiovasc Res.
43:985–991. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Barrett-Connor E and Bush TL: Estrogen and
coronary heart disease in women. JAMA. 265:1861–1867. 1991.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kannel WB, Hjortland MC, McNamara PM and
Gordon T: Menopause and risk of cardiovascular disease: The
Framingham study. Ann Intern Med. 85:447–452. 1976. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Hall E, Volkov P, Dayeh T, Esguerra JL,
Salö S, Eliasson L, Rönn T, Bacos K and Ling C: Sex differences in
the genome-wide DNA methylation pattern and impact on gene
expression, microRNA levels and insulin secretion in human
pancreatic islets. Genome Biol. 15:5222014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kadoglou NP, Kottas G, Lampropoulos S,
Vitta I and Liapis CD: Serum levels of fetuin-A, osteoprotegerin
and osteopontin in patients with coronary artery disease: Effects
of statin (HMGCoA-reductase inhibitor) therapy. Clin Drug Invest.
34:165–171. 2014. View Article : Google Scholar
|
|
29
|
Cross HR: Trimetazidine for stable angina
pectoris. Exp Opin Pharmacother. 2:857–875. 2001. View Article : Google Scholar
|
|
30
|
Ni W, Zhou Z, Liu T, Wang H, Deng J, Liu X
and Xing G: Gender-and lesion number-dependent difference in
‘atherogenic index of plasma’ in Chinese people with coronary heart
disease. Sci Rep. 7:132072017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zhou J, Huang Y, Huang RS, Wang F, Xu L,
Le Y, Yang X, Xu W, Huang X, Lian J and Duan S: A case-control
study provides evidence of association for a common SNP rs974819 in
PDGFD to coronary heart disease and suggests a sex-dependent
effect. Thromb Res. 130:602–606. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Tu Q, Hao J, Zhou X, Yan L, Dai H, Sun B,
Yang D, An S, Lv L, Jiao B, et al: CDKN2B deletion is essential for
pancreatic cancer development instead of unmeaningful co-deletion
due to juxtaposition to CDKN2A. Oncogene. 37:128–138. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Li Y, Chen M, Liu J, Li L, Yang X, Zhao J,
Wu M and Ye M: Upregulation of MicroRNA 18b contributes to the
development of colorectal cancer by inhibiting CDKN2B. Mol Cell
Biol. 37:2017. View Article : Google Scholar
|
|
34
|
Stepanenko AA and Dmitrenko VV: HEK293 in
cell biology and cancer research: Phenotype, karyotype,
tumorigenicity, and stress-induced genome-phenotype evolution.
Gene. 569:182–190. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Burnier JP, Borowski M, Furie BC and Furie
B: Gamma-carboxyglutamic acid. Mol Cell Biochem. 39:191–207. 1981.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Deyl Z, Macek K, Vancikova O and Adam M:
The presence of gamma-carboxyglutamic acid-containing protein in
atheromatous aortae. Biochim Biophys Acta. 581:307–315. 1979.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Xu YZ, Kanagaratham C, Jancik S and
Radzioch D: Promoter deletion analysis using a dual-luciferase
reporter system. Methods Mol Biol. 977:79–93. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Kojima Y, Downing K, Kundu R, Miller C,
Dewey F, Lancero H, Raaz U, Perisic L, Hedin U, Schadt E, et al:
Cyclin-dependent kinase inhibitor 2B regulates efferocytosis and
atherosclerosis. J Clin Invest. 124:1083–1097. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Svensson PA, Wahlstrand B, Olsson M,
Froguel P, Falchi M, Bergman RN, McTernan PG, Hedner T, Carlsson LM
and Jacobson P: CDKN2B expression and subcutaneous adipose tissue
expandability: Possible influence of the 9p21 atherosclerosis
locus. Biochem Biophys Res Commun. 446:1126–1131. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Zhong J, Chen X, Ye H, Wu N, Chen X and
Duan S: CDKN2A and CDKN2B methylation in coronary heart disease
cases and controls. Exp Ther Med. 14:6093–6098. 2017.PubMed/NCBI
|
|
42
|
Vaccarino V: Ischemic heart disease in
women: Many questions, few facts. Circ Cardiovasc Qual Outcomes.
3:111–115. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Pepine CJ, Balaban RS, Bonow RO, Diamond
GA, Johnson BD, Johnson PA, Mosca L, Nissen SE and Pohost GM:
National Heart, Lung and Blood Institute; American College of
Cardiology Foundation: Women's Ischemic Syndrome Evaluation:
Current status and future research directions: Report of the
National Heart, Lung and Blood Institute workshop: October 2-4,
2002: Section 1: Diagnosis of stable ischemia and ischemic heart
disease. Circulation. 109:e44–e46. 2004.PubMed/NCBI
|
|
44
|
Takasugi M, Hayakawa K, Arai D and Shiota
K: Age- and sex-dependent DNA hypomethylation controlled by growth
hormone in mouse liver. Mech Ageing Dev. 134:331–337. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Kwekel JC, Desai VG, Moland CL, Branham WS
and Fuscoe JC: Age and sex dependent changes in liver gene
expression during the life cycle of the rat. BMC Genomics.
11:6752010. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Jousilahti P, Vartiainen E, Tuomilehto J
and Puska P: Sex, age, cardiovascular risk factors, and coronary
heart disease: A prospective follow-up study of 14 786 middle-aged
men and women in Finland. Circulation. 99:1165–1172. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Jousilahti P, Vartiainen E, Tuomilehto J
and Puska P: Twenty-year dynamics of serum cholesterol levels in
the middle-aged population of eastern Finland. Ann Intern Med.
125:713–722. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Mendelsohn ME and Karas RH: Molecular and
cellular basis of cardiovascular gender differences. Science.
308:1583–1587. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Bredfeldt TG, Greathouse KL, Safe SH, Hung
MC, Bedford MT and Walker CL: Xenoestrogen-induced regulation of
EZH2 and histone methylation via estrogen receptor signaling to
PI3K/AKT. Mol Endocrinol. 24:993–1006. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Kulig E, Landefeld TD and Lloyd RV: The
effects of estrogen on prolactin gene methylation in normal and
neoplastic rat pituitary tissues. Am J Pathol. 140:207–214.
1992.PubMed/NCBI
|
|
51
|
Wiggins BS, Saseen JJ, Page RL II, Reed
BN, Sneed K, Kostis JB, Lanfear D, Virani S and Morris PB: American
Heart Association Clinical Pharmacology Committee of the Council on
Clinical Cardiology; Council on Hypertension; Council on Quality of
Care and Outcomes Research; and Council on Functional Genomics and
Translational Biology: Recommendations for management of clinically
significant drug-drug interactions with statins and select agents
used in patients with cardiovascular disease: A scientific
statement from the American Heart Association. Circulation.
134:e468–e495. 2016.PubMed/NCBI
|
|
52
|
Frishman WH: β-Adrenergic blockade in
cardiovascular disease. J Cardiovasc Pharmacol Ther. 18:310–319.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Horinaka S: Use of nicorandil in
cardiovascular disease and its optimization. Drugs. 71:1105–1119.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Ujhelyi MR, Ferguson RK and Vlasses PH:
Angiotensin-converting enzyme inhibitors: Mechanistic
controversies. Pharmacotherapy. 9:351–362. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Liu MZ, He FZ and Zhang W: Epigenetic
research progress of anti-tumor drugs. Yao Xue Xue Bao.
48:1629–1636. 2013.(In Chinese). PubMed/NCBI
|
|
56
|
Stone A, Valdés-Mora F, Gee JM, Farrow L,
McClelland RA, Fiegl H, Dutkowski C, McCloy RA, Sutherland RL,
Musgrove EA and Nicholson RI: Tamoxifen-induced epigenetic
silencing of oestrogen-regulated genes in anti-hormone resistant
breast cancer. PLoS One. 7:e404662012. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Baker EK, Johnstone RW, Zalcberg JR and
El-Osta A: Epigenetic changes to the MDR1 locus in response to
chemotherapeutic drugs. Oncogene. 24:8061–8075. 2005. View Article : Google Scholar : PubMed/NCBI
|