Introduction
Osteoarthritis (OA) is characterized by gradual
degeneration of articular cartilage due to various causes of
injury, hyperplasia at the edge of joint, formation of osteophytes,
and synovial inflammatory response (1). OA is the most common cause of joint
disease, especially in the elderly, and its clinical manifestations
are joint pain and dysfunction (2,3).
Although there are many causes of OA in humans, including changes
in subchondral bone, ligamentous laxity and synovial fluid
inflammation, articular cartilage injury is still an important
pathological factor leading to OA, and studied most frequently
(3,4). Reasons for articular cartilage injury
mainly include biomechanical and biochemical factors, which also
interact with each other. The structure of articular cartilage is
different from other tissues, and is almost impossible to repair
when it is damaged due to degeneration or trauma, ultimately
developing to pathological changes in OA (5,6).
Therefore, there is a lack of effective clinical treatment means of
OA currently. The early conservative treatment is mainly based on
oral administration of drugs and physical therapy, but they can
only relieve the symptoms, but cannot delay the progression of
disease. OA in the late stage can only be treated with surgery to
relieve joint pain and improve function. As the incidence of OA is
very common, a large number of medical resources are consumed each
year. Therefore, prevention and intervention of the disease are
particularly important.
The number of mesenchymal stem cells (MSCs) in
articular cartilage is very small, accounting for only 3–4% of that
of articular chondrocytes. When OA occurs in human, the
differentiation of MSCs in articular cartilage, despite its active
proliferation, into osteoblasts or cartilages is very limited. The
importance of using tissue engineering technique to overexpress the
target gene, promoting the proliferation and differentiation of
MSCs into cartilages when OA occurs, maintaining the dynamic
balance between cartilage tissue injury and repair, and restoring
the self-regulation and self-repair capacities of articular
chondrocytes in OA is inestimable in the treatment of OA (7,8). At
present, the treatment method is generally culturing cytokines
in vitro first to make tissue engineering components, and
then implanting them into the body, but the stability is poor with
higher cost, so it is not suitable to be applied directly (9). Therefore, introducing the target gene
into cells combined with transgenic technology can repair the
degraded or damaged cartilage through the high expression of target
gene in cells to promote the proliferation and differentiation of
MSCs into cartilages, so the research on these target genes has
important application prospect and value in the treatment of human
OA.
Micro ribonucleic acid (miRNA) is a class of
non-coding single-stranded RNA with approximately 18–22 nucleotides
in length, which negatively regulates the post-transcriptional
expression of target gene by binding specific sequences to
messenger RNA (mRNA). In recent years, studies have shown that
miRNA is not only involved in the regulation of articular cartilage
development, but also closely related to the occurrence and
development of OA (10–11).
The regulatory effect of miR-101 on differentiation
of MSCs in articular cartilage was studied, investigating whether
miR-101 can regulate the differentiation of MSCs into chondrocytes,
aiming elucidation of the interaction mechanism of miRNA and
protein-coding genes during the differentiation process of MSCs
induced by osteogenic microenvironment, and to lay a foundation for
the regulation of biological behavior of chondrocytes using miRNA
in clinic, which is expected to provide valuable clues for the
treatment of OA.
Materials and methods
Isolation and culture of MSCs
A total of 60 healthy Sprague-Dawley (SD) rats
(male=30; female=30) aged 12 weeks (meanweight=400 g) were selected
and sacrificed under anesthesia. The rats were housed in a
temperature controlled room (21±2°C) on a 12:12-h light/dark cycle
(lights on at 06:00). All rats had free access to water and food.
This study was approved by the Animal Ethics Committee of Jilin
University Animal Center (Changchun, China). Bilateral lower
extremities of rats were isolated, the epiphyseal ends were cut
off, and the bone marrow in marrow cavity was poured out. MSCs were
gently blown and beaten using phosphate-buffered saline (PBS) to be
prepared into single-cell suspension, and centrifuged at 1,050 × g
for 10 min. After the supernatant was discarded, cells were
resuspended in F12/Dulbecco's modified Eagle medium (DMEM),
inoculated into a 25 cm2 culture flask, and cultured
under 5% CO2 at 37°C. The solution was replaced once every 2 days.
The cell morphology and growth status were observed under an
inverted microscope (AZ100; Nikon Corporation, Tokyo, Japan). When
cells were fused, they were digested using 0.25% trypsin, followed
by passage at a ratio of 1:2.
Cell transfection and treatment
BMSCs at passage 3 were pre-cultured in a 24-well
plate for 24 h, miR-101 mimics and inhibitor were synthesized and
transfected to MSC cells to analyze biological function of miR-101.
Then three groups were established to study the potential relevance
between miR-101 and chondrogenic differentiation: miR-NC group
(negative control), miR-101 mimics (BMSCs transfected by miR-101
mimics) and mimics + inhibitor (BMSCs transfected by miR-101 mimics
and inhibitor). The above material were purchased from Guangzhou
RiboBio Co., Ltd. (Guangzhou, China), and transfected by using
Lipofectamine RNAiMAX (Life Technologies; Thermo Fisher Scientific,
Inc., Waltham, MA, USA) according to the manufacturer's protocol.
At 36 h after transfection, the chondrogenic differentiation of
bone marrow-derived MSCs into chondrocytes was induced, and
cartilage micelles were constructed via centrifugation at 2,500 × g
for 5 min.
Induced chondrogenic differentiation
of bone marrow-derived MSCs into chondrocytes
The cell suspension was prepared and the cell
concentration was adjusted into 1×109/l. Then cell
suspension was transferred into a centrifuge tube for
centrifugation at 2,500 × g for 5 min, and the supernatant was
discarded. Cartilage-inducing liquid (containing 50 mg/l ascorbic
acid, 1% fetal bovine serum, 100 nmol/l dexamethasone, 100 mg/l
sodium pyruvate, 1.0% indometacin, 40 mg/l L-proline, and
low-glucose DMEM) was added, cell mass was not dispersed, and cells
were cultured under 5% CO2 at 37°C. The cartilage-inducing liquid
was replaced once every 3 days, and cartilage mass was formed after
induction for 3 weeks.
Cell proliferation
At 21 days after induction of chondrogenic
differentiation, cells were harvested and inoculated into 96-well
plates at a density of 2×103 cells for 48 h, MTT
solution [5 mg/ml; Multisciences (Lianke) Biotech Co., Ltd.,
Hangzhou, China] was appended to each well after 4 h incubation.
Then, 150 µl of dimethyl sulfoxide (DMSO) was added to each well
for solubilizing the formazan. After 30 min, the absorbance was
measured by a microplate reader (Bio-Rad Laboratories, Inc.,
Hercules, CA, USA) set at 490 nm.
Alcian blue staining and
quantification
At 21 days after induction of chondrogenic
differentiation, the medium was discarded. Cells were washed twice
with PBS, and 0.5 ml alcian blue dye solution prepared by 0.1 mol/l
hydrochloric acid was added into each well. After the mixture was
placed at room temperature overnight, the dye solution was
discarded. Then cells were washed with double distilled water
several times, photographed under the inverted microscope (AZ100;
Nikon Corporation), added with 6 mol/l guanidine hydrochloride, and
placed at room temperature for 2 h. After the washing solution was
removed, the optical density was measured by a microplate reader
(Bio-Rad Laboratories, Inc., Hercules, CA, USA) set at 630 nm.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was procured by TRIzol reagent in
accordance with the manufacturer's protocol. SYBR-Green qPCR assay
was used to measure the level of collagen II, aggrecan, SRY-related
high mobility group-box gene9 (Sox9) and Runt-related transcription
factor 2 (Runx2) expression and endogenous controlled by GAPDH.
TaqMan miRNA assays (Applied Biosystems; Thermo Fisher Scientific,
Inc.) was used to measure the level of miR-101 expression
normalized to miRNA U6. Primers used in PCR reaction were listed as
follows: Sox9 forward, 5′-ATGGAAATCACGGAAGAGCGTC-3′; reverse,
5′-GTGCTGAAGGGCTACGACTGGA-3′. Runx2 forward,
5′-ACCAGCAGCACTCCATATCTCTAC-3′; reverse, 5′-CTTCCATCAGCGTCAACACCATC
−3′. MiR-101 forward, 5′-GCGGGCGTAGTGATAA-3′; reverse,
5′-GTGCAGGTCCGAGGT-3′. GAPDH forward, 5′-ACCTCAACTACATGGTCTAC-3′;
reverse, 5′-TTGTCATTGAGAGCAATCC-3′. With U6 as endogenous control,
the relative expression level of miR-101 was calculated by
2−ΔΔCq method (12).
Western blot analysis
At 21 days after induction of chondrogenic
differentiation, cells were collected and lysed using
radioimmunoprecipitation assay (RIPA) buffer. The total protein was
extracted, and its concentration was determined according to
instructions of the bicinchoninic acid (BCA) protein concentration
kit. The same amount of total protein (30 ug proteins per lane) was
separated via sodium dodecyl sulfate-polyacrylamide gel
electropheresis (10% SDS-PAGE), transferred onto a nitrocellulose
membrane, sealed with 5% skim milk and incubated with rabbit
anti-rat Sox9 and Runx2 primary antibodies (1:1,000) at 4°C
overnight. Primary mouse monoclonal Runx2 antibody (dilution,
1:500; cat. no. ab76956); rabbit monoclonal Sox9 antibody
(dilution, 1:500; cat. no. ab185966); rabbit polyclonal GAPDH
antibody (dilution, 1:500; cat. no. ab37168) and secondary goat
anti-rabbit (HRP) IgG antibody (dilution, 1:2,000; cat. no. ab6721)
were all purchased from Abcam (Cambridge, MA, USA). After the
membrane was fully washed with Tris-buffered saline with Tween-20
(TBST), anti-rabbit secondary antibody (coupled by horseradish
peroxidase) was added for incubation at room temperature for 2 h,
followed by development via enhanced chemiluminescence (ECL) (Merck
Millipore, Billerica, MA, USA), expo-sure in gel imaging system,
fixation and observation of results. With β-actin as an internal
reference, the relative changes in protein expression were
detected. Image J software (Version 1.38; National Institutes of
Health, Bethesda, MA, USA) was used for protein quantification.
Immunohistochemical staining
At 21 days after induction of chondrogenic
differentiation, the medium was discarded. Cells were washed twice
with PBS, and fixed with 4% paraformaldehyde for 30 min.
Immunohistochemical staining was performed for Sox9 and Runx2
according to protocol of the kit, and they were photographed under
the microscope.
Statistical analysis
Statistical Product and Service Solutions (SPSS)
19.0 software (IBM Corp., Armonk, NY, USA) was used for statistical
analysis. All quantitative data are expressed as mean ± standard
deviation. Comparison between groups was done using one-way ANOVA
test followed by post hoc test (least significant difference).
P<0.05 was considered to indicate a statistically significant
difference.
Results
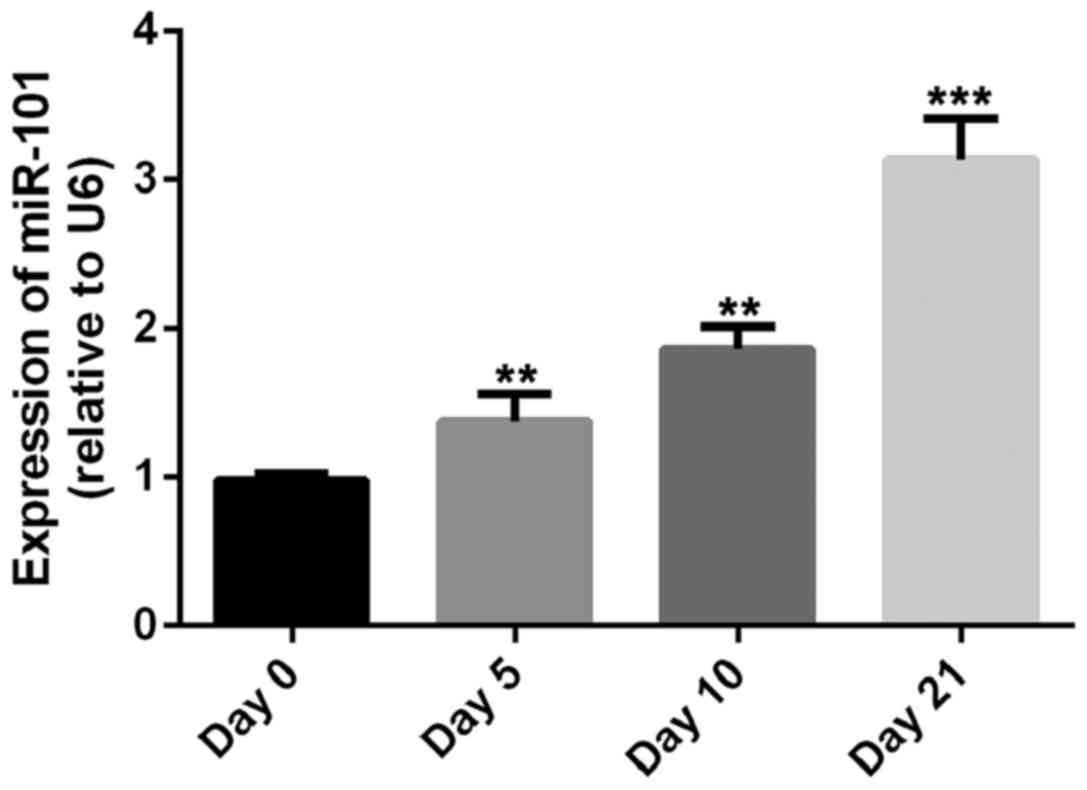
miR-101 is upregulated during
chondrogenic differentiation of MSCs
The present study demonstrated significant
upregulation of miR-101 expression during chondrogenic
differentiation (13). In our
preliminary experiment, the expression of miR-101 was detected
after the induction of chondrogenic differentiation of MSCs by
RT-qPCR, consistent with the literature, the expression level of
miR-101 was increased during the chondrogenic differentiation of
MSCs (Fig. 1). The results indicated
that miR-101 may have a regulating effect on the chondrogenic
differentiation of MSCs (Fig.
1).
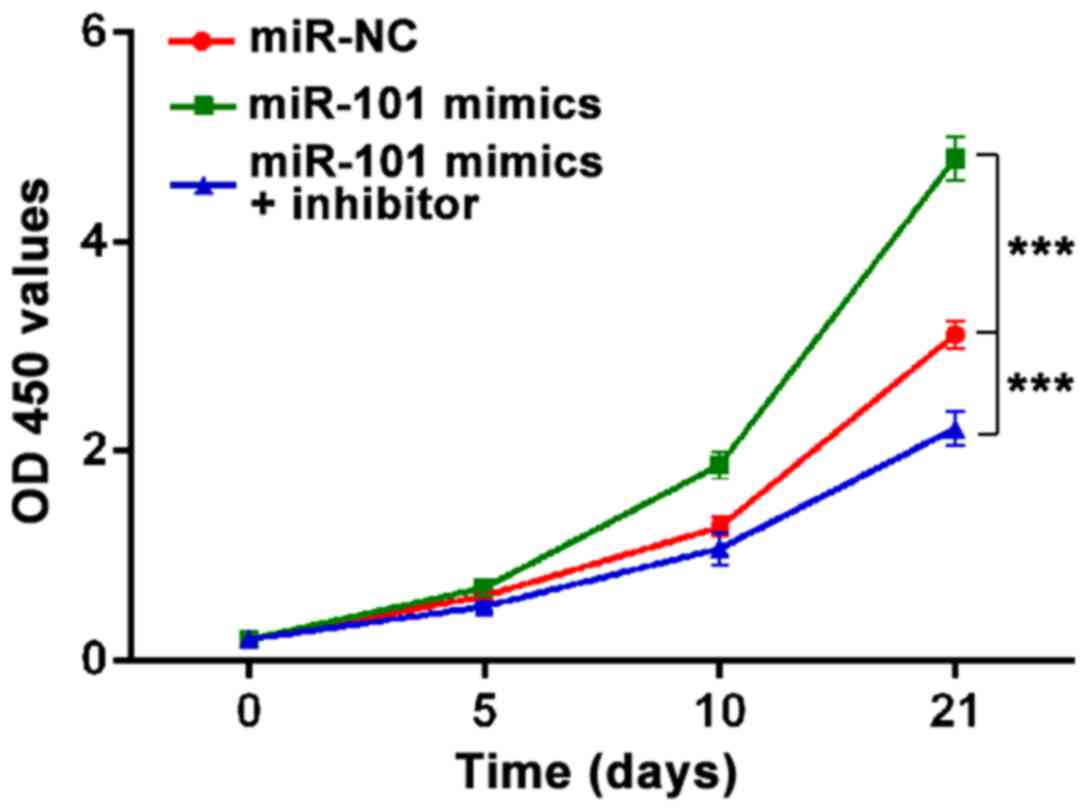
miR-101 promotes the cell viability of
MSCs
To examine the function of miR-101 on proliferation
of MSCs, we used MTT assay to detect the cell viability at 0, 5, 10
and 21 day of chondrogenic differentiation. The results from the
MTT assay revealed that miR-101 obviously enhanced MSC viability,
whereas downregulation of miR-101 by inhibitor markedly suppressed
MSC viability after four days incubation (Fig. 2).
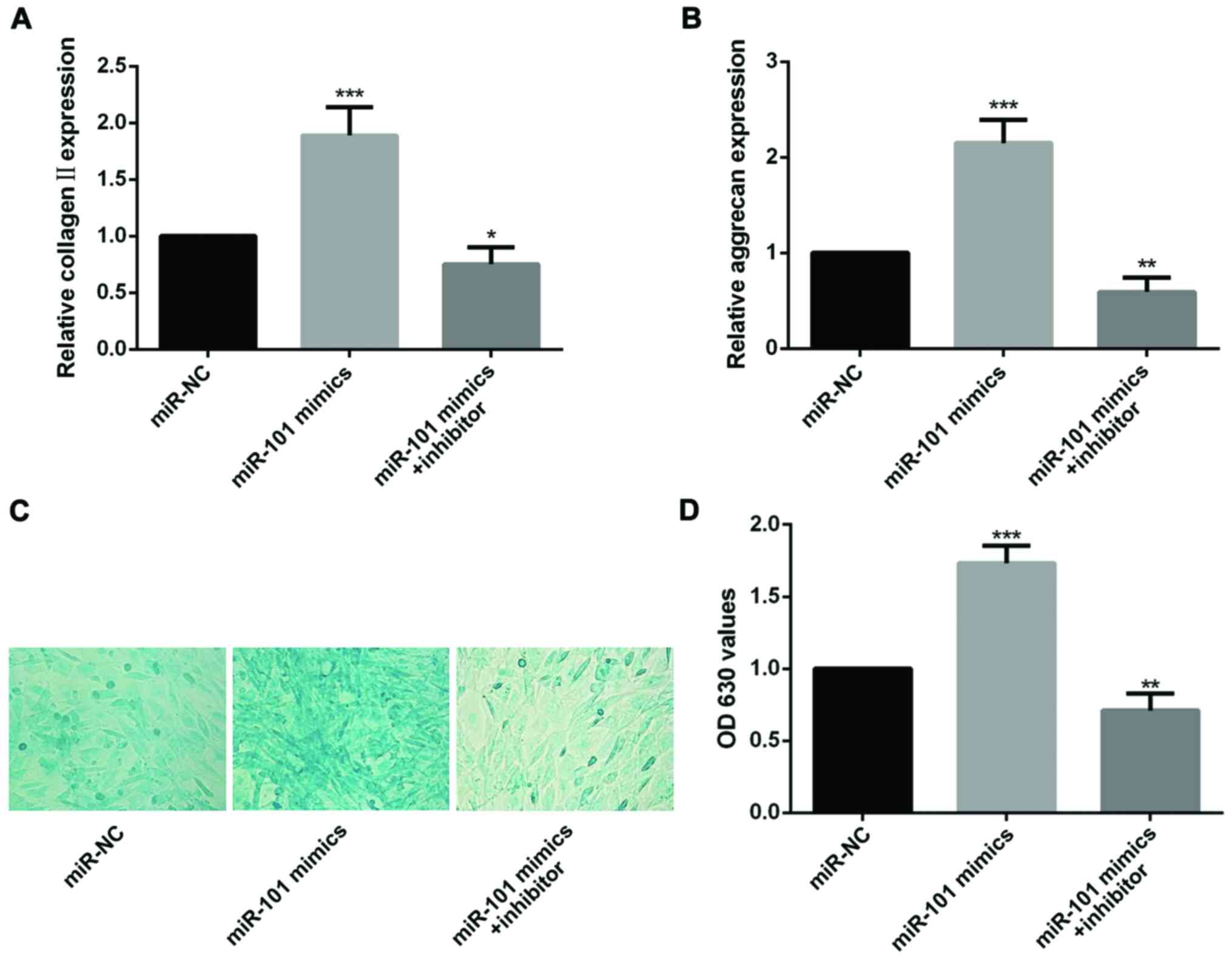
miR-101 increases the expression of
chondrogenic markers
Collagen II and aggrecan have been designed as
unique chondrogenic markers (14).
RT-qPCR was used to analyze the effect of miR-101 on expression of
chondrogenic markers. the results show that both expression level
of collagen II and aggrecan could be upregulated by intervened with
miR-101 mimics, and on the contrary, mRNA levels of two
chondrogenic markers were restrained by miR-101 inhibitor (Fig. 3A and B).
Alcian blue staining was a method to
detect the glycosaminoglycans in cells
After induction of chondrogenic differentiation for
21 days, alcian blue staining was performed, the intensity of the
cells in miR-101 mimics group was markedly increased by comparing
with the miR-NC group. Notably, the intensity was reduced by adding
miR-101 inhibitor (Fig. 3C and
D).
The above results suggest that miR-101 has a
positive effect during chondrogenic differentiation of MSCs.
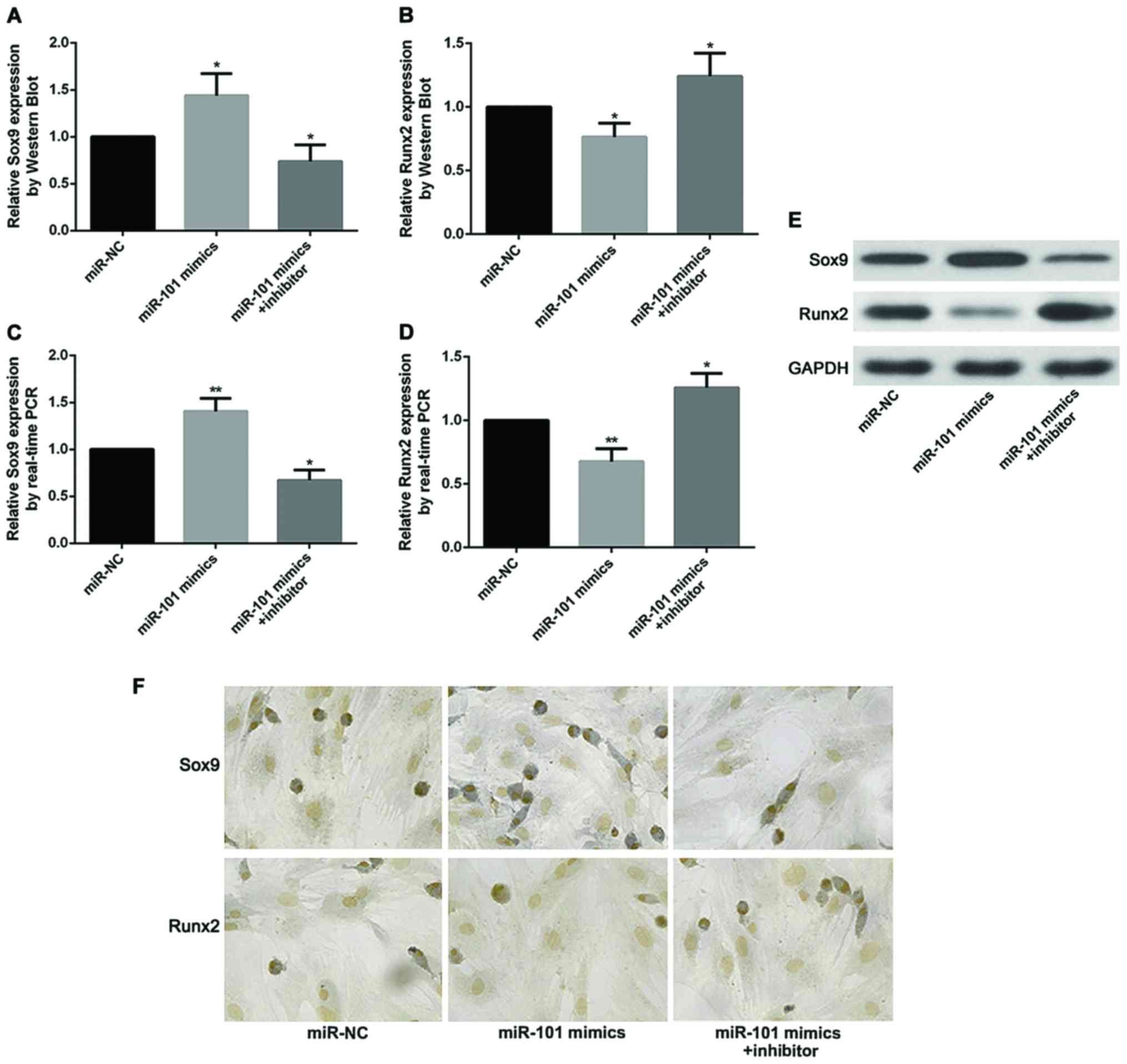
miR-101 increases the expression level
of Sox9 and decrease the expression level of Runx2
Recent studies have demonstrated that expression
levels of Sox9 and Runx2 directly impact the progress of OA. In the
pathogenesis of OA, Sox9 promotes the differentiation of
chondrocytes and has inhibitory effects on OA, while Runx2 mediates
the degradation of chondrocytes and promotes the OA. In our
research, expression of Sox9 was found substantially increased
after treatment with miR-101 mimics by comparing with the miR-NC
group. However, the effect of miR-101 has been counteracted by
addition of miR-101 inhibitor. As expected, the opposite trend was
found in the expression of Runx2 measuring by RT-qPCR, western blot
analysis and immunohistochemical staining (Fig. 4).
Discussion
OA is the most common chronic disease among all
types of degenerative joint diseases, which can affect tissues and
joints in the whole body (15).
There are not many treatment methods for OA, and the effects of
drugs and non-surgical treatment are not satisfactory, so joint
replacement surgery is a last choice for OA in the late stage.
Currently, MSCs used to treat various degenerative skeletal muscle
diseases provide a new therapeutic strategy for OA (8).
In the process of cartilage formation, Sox9 gene
plays an important role in the cartilage differentiation and has a
protective effect on cartilage tissues. However, when the
degradation rate of cartilage tissues exceeds its synthesis rate,
it will develop into the pathological process of OA. In addition,
endochondral ossification also plays a crucial role in the
pathogenesis of OA (16). The
pathological process of OA actually begins from the transformation
from chondrocytes to mastocytes, which transforms cartilage tissues
from low-oxygen tissues without vessels to bone tissues with blood
supply through the expressions of type X collagen, vascular
endothelial growth factor, metalloproteinase 13 and other molecular
substances, accompanied by matrix degeneration, vascular
endothelial invasion, endochondral ossification at the center of
joint, and marginal osteophyte formation. Runx2 is an important
factor initiating this pathological process.
Sox9 gene is expressed in all pre-chondrocytes and
differentiated chondrocytes, excluding hypertrophic chondrocytes,
which not only has important positive regulatory and control
effects on the differentiation and maturation of cartilage, but
also is essential for the normal physiological development process
of cartilage growth plates and endochondral ossification (17,18). The
differentiation of MSCs into cartilage and the maintenance of
chondrocyte phenotype depend on the stable expression of Sox9 gene
(19–22). Runx2 is highly expressed in
pre-hypertrophic chondrocytes and hypertrophic chondrocytes in
mammals. Runx2 is involved in both endochondral ossification and
chondrocyte hypertrophy. Besides, Runx2 can be activated in many
signaling pathways on osteogenic differentiation, such as
transforming growth factor-β (TGF-β) (23), Wnt/β-catenin (24), Notch (25) and bone morphogenetic protein 2 (BMP2)
(26), activating a series of
downstream genes, and making cells differentiate into osteoblasts.
It is found from the above that during the pathogenesis of OA, Sox9
inhibits OA through promoting chondrocyte differentiation, while
Runx2 mediates the process of chondrocyte hypertrophy and controls
the degradation of chondrocytes, thus promoting OA.
In the present study, the expression of miR-101 on
differentiation of MSCs in articular cartilage was analyzed, and it
was found that the expression level of miR-101 was upregulated
during chondrogenic differentiation of MSCs. After induction of
chondrogenic differentiation for 21 days. MTT assay, alcian blue
staining, RT-qPCR, western blot analysis and immunohistochemical
staining were performed, and it was found that miR-101 could
promote the cell viability of MSCs and elevate the expression of
chondrogenic markers such as collagen II, aggrecan and
glycosaminoglycans. Furthermore, the expression level of Sox9 which
has promotion effect in chondrogenic differentiation was more
noticeable in MSCs in miR-101 group, while the expression level of
Runx2, which is a obstructive factor in chondrogenic
differentiation, was decreased by miR-101 intervention. All effects
of miR-101 on chondrogenic differentiation of MSCs could be
suppression by miR-101 inhibitor.
In conclusion, in the present study, miR-101 was
found upregulated during chondrogenic differentiation and play an
important role to deaccelerate chondrogenic differentiation of
MSCs, and its function depends on regulating the expression of Sox9
and Runx2, our results revealed that miR-101 could be a potential
therapeutic strategy for the treatment of OA.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
FG and MW designed the study and performed the
experiments, FG, CZ and SZ established the animal models, CP and SZ
collected the data, FG and CZ analyzed the data, FG and MW prepared
the manuscript. All authors read and approved the final study.
Ethics approval and consent to
participate
This study was approved by the Animal Ethics
Committee of Jilin University Animal Center (Changchun, China).
Patient consent for publication
Not applicable
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Goldring MB and Goldring SR: Articular
cartilage and subchondral bone in the pathogenesis of
osteoarthritis. Ann N Y Acad Sci. 1192:230–237. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Reynard LN and Loughlin J: Insights from
human genetic studies into the pathways involved in osteoarthritis.
Nat Rev Rheumatol. 9:573–583. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Blagojevic M, Jinks C, Jeffery A and
Jordan KP: Risk factors for onset of osteoarthritis of the knee in
older adults: A systematic review and meta-analysis. Osteoarthritis
Cartilage. 18:24–33. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Qi Y, Feng G and Yan W: Mesenchymal stem
cell-based treatment for cartilage defects in osteoarthritis. Mol
Biol Rep. 39:5683–5689. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Guermazi A, Niu J, Hayashi D, Roemer FW,
Englund M, Neogi T, Aliabadi P, McLennan CE and Felson DT:
Prevalence of abnormalities in knees detected by MRI in adults
without knee osteoarthritis: Population based observational study
(Framingham Osteoarthritis Study). BMJ. 29:e53392012. View Article : Google Scholar
|
|
6
|
Kao YJ, Ho J and Allen CR: Evaluation and
management of osteochondral lesions of the knee. Phys Sportsmed.
39:60–69. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Longo UG, Petrillo S, Franceschetti E,
Berton A, Maffulli N and Denaro V: Stem cells and gene therapy for
cartilage repair. Stem Cells Int. 2012:1683852012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Sampson S, Botto-van Bemden A and Aufiero
D: Stem cell therapies for treatment of cartilage and bone
disorders: Osteoarthritis, avascular necrosis, and non-union
fractures. PM R. 7 Suppl 4:S26–S32. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Houdek MT, Wyles CC, Martin JR and Sierra
RJ: Stem cell treatment for avascular necrosis of the femoral head:
Current perspectives. Stem Cells Cloning. 7:65–70. 2014.PubMed/NCBI
|
|
10
|
Wei ZJ, Liu J and Qin J: miR-138
suppressed the progression of osteoarthritis mainly through
targeting p65. Eur Rev Med Pharmacol Sci. 21:2177–2184.
2017.PubMed/NCBI
|
|
11
|
Le LT, Swingler TE, Crowe N, Vincent TL,
Barter MJ, Donell ST, Delany AM, Dalmay T, Young DA and Clark IM:
The microRNA-29 family in cartilage homeostasis and osteoarthritis.
J Mol Med (Berl). 94:583–596. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Han J, Yang T, Gao J, Wu J, Qiu X, Fan Q
and Ma B: Specific microRNA expression during chondrogenesis of
human mesenchymal stem cells. Int J Mol Med. 25:377–384.
2010.PubMed/NCBI
|
|
14
|
Lahiji A, Sohrabi A, Hungerford DS and
Frondoza CG: Chitosan supports the expression of extracellular
matrix proteins in human osteoblasts and chondrocytes. J Biomed
Mater Res. 51:586–595. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Cucchiarini M, de Girolamo L, Filardo G,
Oliveira JM, Orth P, Pape D and Reboul P: Basic science of
osteoarthritis. J Exp Orthop. 3:222016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kawaguchi H: Regulation of osteoarthritis
development by Wnt-beta-catenin signaling through the endochondral
ossification process. J Bone Miner Res. 24:8–11. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Guérit D, Philipot D, Chuchana P, Toupet
K, Brondello JM, Mathieu M, Jorgensen C and Noël D: Sox9-regulated
miRNA-574-3p inhibits chondrogenic differentiation of mesenchymal
stem cells. PLoS One. 8:e625822013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Cairns DM, Liu R, Sen M, Canner JP,
Schindeler A, Little DG and Zeng L: Interplay of Nkx3.2, Sox9 and
Pax3 regulates chondrogenic differentiation of muscle progenitor
cells. PLoS One. 7:e396422012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Cao L, Yang F, Liu G, Yu D, Li H, Fan Q,
Gan Y, Tang T and Dai K: The promotion of cartilage defect repair
using adenovirus mediated Sox9 gene transfer of rabbit bone marrow
mesenchymal stem cells. Biomaterials. 32:3910–3920. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Cucchiarini M, Orth P and Madry H: Direct
rAAV SOX9 administration for durable articular cartilage repair
with delayed terminal differentiation and hypertrophy in vivo. J
Mol Med (Berl). 91:625–636. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Hargus G, Kist R, Kramer J, Gerstel D,
Neitz A, Scherer G and Rohwedel J: Loss of Sox9 function results in
defective chondrocyte differentiation of mouse embryonic stem cells
in vitro. Int J Dev Biol. 52:323–332. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
de Crombrugghe B, Lefebvre V, Behringer
RR, Bi W, Murakami S and Huang W: Transcriptional mechanisms of
chondrocyte differentiation. Matrix Biol. 19:389–394. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ito Y and Miyazono K: RUNX transcription
factors as key targets of TGF-beta superfamily signaling. Curr Opin
Genet Dev. 13:43–47. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Marie PJ: Transcription factors
controlling osteoblastogenesis. Arch Biochem Biophys. 473:98–105.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Shen Q and Christakos S: The vitamin D
receptor, Runx2, and the Notch signaling pathway cooperate in the
transcriptional regulation of osteopontin. J Biol Chem.
280:40589–40598. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Hassan MQ, Tare RS, Lee SH, Mandeville M,
Morasso MI, Javed A, van Wijnen AJ, Stein JL, Stein GS and Lian JB:
BMP2 commitment to the osteogenic lineage involves activation of
Runx2 by DLX3 and a homeodomain transcriptional network. J Biol
Chem. 281:40515–40526. 2006. View Article : Google Scholar : PubMed/NCBI
|