Introduction
Lung cancer is one of the most common types of
cancer with high incidence and mortality rates (1). Lung cancer accounts for 1.6 million
mortalities each year and is one of the most common causes of
cancer-associated mortality worldwide (2,3). Lung
cancer is the fourth leading cause of mortality in China (2,3).
Non-small cell lung cancer (NSCLC) is a subtype of lung cancer,
which accounts for approximately 85% of all lung cancer cases
(4). NSCLC can be classified into
several subtypes which include, squamous cell carcinoma,
adenocarcinoma, large-cell carcinoma and bronchioloalveolar
carcinoma (5). The main treatment
options for patients with NSCLC include surgery, radiotherapy and
chemotherapy (4). Surgery is
regarded as the most efficient treatment, however clinical
application is limited to patients with advanced NSCLC, which
account for 70% of all new lung cancer cases (6). The development of new therapeutic
strategies based on chemotherapeutic agents is required to
potentially improve clinical outcomes.
Targeted therapies for NSCLC have made promising
progress (7,8). Gefitinib is a well-known targeted drug
for the treatment with NSCLC, which acts as an epidermal growth
factor receptor (EGFR)-tyrosine kinase inhibitor (TKI) (9). Gefitinib competitively inhibits ATP
binding at the ATP intracellular domain of EGFR, to prevent the
autophosphorylation and activation of downstream signaling
pathways, leading to the inhibition of tumor cell proliferation,
metastasis, and angiogenesis (10).
Gefitinib is widely used as the standard first-line treatment for
patients with advanced NSCLC with active EGFR mutations (11,12).
Although gefitinib can improve the progression-free survival and
overall survival of patients with NSCLC, issues regarding drug
resistance, toxicity and limited applicability need to be addressed
(13).
Natural products, which include Chinese herbal
medicine extracts, have gained increasing attention in tumor
therapy due to high efficacy and low toxicity. Combined with
chemotherapeutic agents or targeted therapies, natural products may
enhance the antitumor efficacy whilst reducing side effects
associated with traditional therapeutic strategies (14–16).
Ginsenoside Rg3 is a steroidal saponin isolated from a traditional
Chinese herbal medicine, Panax ginseng, with anticancer
activity (17). Several studies
revealed that ginsenoside Rg3 could enhance lung cancer sensitivity
to chemotherapy (18,19). The aim of the current study was to
investigate whether ginsenoside Rg3 enhances gefitinib efficiency
in altering NSCLC cell proliferation, apoptosis, and migration by
using two NSCLC cell lines with different sensitivities to
gefitinib.
Materials and methods
Cell culture and reagents
NSCLC cell lines A549 and H1299 were purchased from
Chinese Academy of Sciences (Shanghai, China). Cells were cultured
in RPMI-1640 medium (HyClone; GE Healthcare Life Sciences, Logan,
UT, USA) supplemented with 10% fetal bovine serum (FBS; Invitrogen;
Thermo Fisher Scientific, Inc., Waltham, MA, USA), 100 U/ml
penicillin and 100 µg/ml streptomycin and maintained at 37°C in a
5% CO2-humidified incubator. Gefitinib was purchased
from Selleck Chemicals (cat. no. S1025; Houston, TX, USA) and
dissolved in DMSO (Sigma-Aldrich; Merck KGaA, Darmstadt, Germany)
at a concentration of 100 mM. Ginsenoside Rg3 was purchased from
Biopurify Phytochemicals Ltd. (Chengdu, China) and dissolved in
DMSO at a concentration of 50 mg/ml.
Cell proliferation assay
Cell viability was measured using MTT assay (cat.
no. C0009; Beyotime Institute of Biotechnology, Haimen, China).
A549 or H1299 cells were seeded in 96-well plates at a density of
1×104 cells/well and cultured overnight. Following
incubation, cells were treated with 0, 5, 10 or 20 µM gefitinib
with or without ginsenoside Rg3 (12.5 or 25 µg/ml) at 37°C for 24,
48 or 72 h. After washing twice with PBS, MTT solution was added to
each well to a final concentration of 0.5 mg/ml and further
incubated for 4 h. The MTT solution was removed and 100 µl DMSO was
added to each well to dissolve the formazan crystals. Cell
viability was determined by measuring the absorbance at a
wavelength of 570 nm using a microplate reader (Thermo Fisher
Scientific, Inc.). The cell viability rates were normalized to the
DMSO-treated control group.
Flow cytometric analysis of
apoptosis
A549 or H1299 cells were seeded in six-well plates
at a density of 5×105 cells/well and cultured overnight.
Following incubation, cells were treated with 10 µM gefitinib with
or without 12.5 µg/ml ginsenoside Rg3 at 37°C for 48 h. Following a
48-h incubation, cells were harvested, washed twice with PBS and
subsequently stained using the Annexin V-fluorescein isothiocyanate
(FITC)/propidium iodide (PI) Double Staining kit (Nanjing KeyGen
Biotech Co., Ltd., Nanjing, China), according to the manufacturer's
protocol. Apoptotic cells were measured using a flow cytometer (BD
Biosciences, Franklin Lakes, CA, USA) and the data were analyzed
using FlowJo software (version 7.6.1; FlowJo LLC, Ashland, OR,
USA).
Transwell migration assay
Cells were pre-treated with 10 µM gefitinib with or
without 12.5 µg/ml ginsenoside Rg3 at 37°C for 24 h, cells were
collected and re-suspended in culture medium without FBS at a
density of 5×105 cells/ml. Using Transwell chambers with
a pore size of 8 µm (Costar; Corning Inc., Corning, NY, USA), a
total of 1×105 A549 or H1299 cells in RPMI-1640 medium
without FBS were plated in the upper chamber, and 600 µl RPMI-1640
medium supplemented with 20% FBS was plated in the lower chamber.
Following incubation for 24 h, the migratory cells were fixed with
4% paraformaldehyde at room temperature for 15 min, washed twice
with PBS and stained with 0.05% crystal violet at room temperature
for 10 min. The unmigrated cells were removed with cotton swabs and
the stained migrated cells were counted using a light microscope
(magnification, ×100).
Western blot analysis
A549 or H1299 cells in 2 ml RPMI-1640 medium were
seeded in six-well plates at a density of 5×105
cells/well and cultured overnight. Subsequently, cells were treated
with 10 µM gefitinib with or without 12.5 µg/ml ginsenoside Rg3 at
37°C for 48 h. Following treatment for 48 h, total protein was
extracted from cells using radioimmunoprecipitation assay buffer
(Beyotime Institute of Biotechnology). Total protein was quantified
using bicinchoninic acid assay (Beyotime Institute of
Biotechnology) and 40 µg protein/lane was separated via SDS-PAGE on
a 10% gel. The separated proteins were transferred onto 0.45 mm
Immobilon-P polyvinylidene difluoride membranes (EMD Millipore,
Billerica, MA, USA). The membranes were blocked in 5% fat-free milk
for 1 h at room temperature and incubated overnight at 4°C with
primary antibodies against cleaved-caspase-3 (1:1,000; cat. no.
9661; CST Biological Reagents Co., Ltd., Shanghai, China), snail
family transcriptional repressor 1 (SNAIL; 1:500; cat. no.
sc-393172), snail family transcriptional repressor 2 (SLUG; 1:500;
cat. no. sc-166476), Bcl-2-associated X (Bax; 1:200; cat. no.
sc-49; all Santa Cruz Biotechnology, Inc., Dallas, TX, USA),
E-cadherin (1:1,000; cat. no. ab15148), B-cell lymphoma 2 (Bcl-2;
1:1,000; cat. no. ab194583), and GAPDH (1:5,000; cat. no. ab181602:
All Abcam, Cambridge, UK). Following the primary incubation,
membranes were incubated with horseradish peroxidase-labelled
secondary antibodies, including Peroxidase AffiniPrue Goat
Anti-Rabbit IgG (1:3,000; cat. no. 111-035-003) and Peroxidase
AffiniPrue Goat Anti-Mouse IgG (1:3,000; cat. no. 115-035-003; both
Jackson ImmunoResearch Europe, Ltd., Newmarket, UK) for 1 h at room
temperature. Protein bands were visualized using the enhanced
chemiluminescence detection reagents (Pierce™ ECL Western Blotting
Substrate; Thermo Fisher Scientific, Inc.), according to the
manufacturer's protocol, and blots were analyzed using ChemiDoc™
XRS+ System (Bio-Rad Laboratories, Inc., Hercules, CA, USA,
USA).
Statistical analysis
Data are presented as the mean ± standard deviation.
All statistical analyses were performed using GraphPad Prism
software (version 5.0; Graphpad Software, Inc., La Jolla, CA, USA).
Statistical analyses were performed using one-way analysis of
variance followed by Bonferroni correction. P<0.05 was
considered to indicate a statistically significant difference.
Results
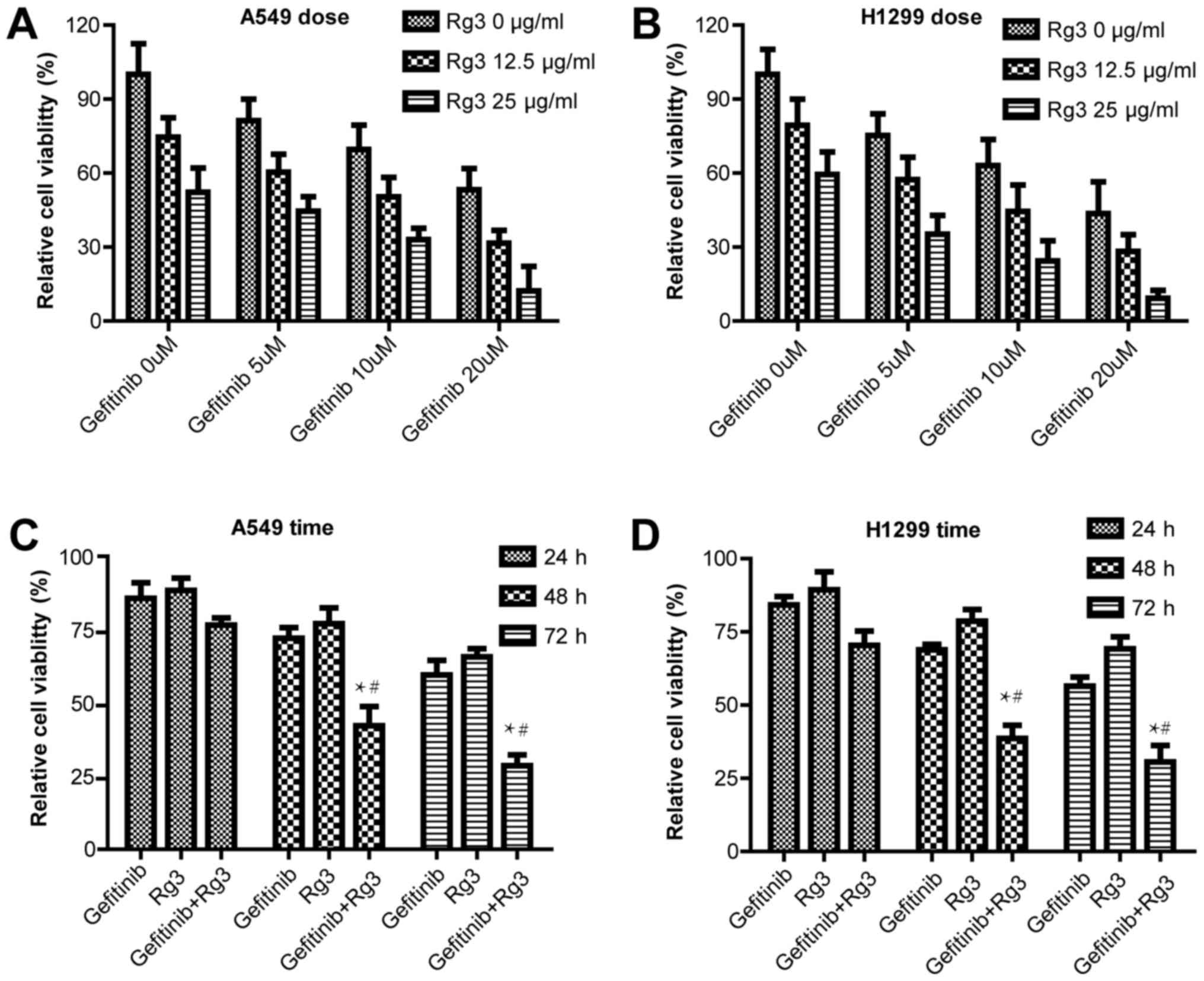
Effect of ginsenoside Rg3 on the
cytotoxic activity of gefitinib in NSCLC cell lines
MTT assay was used to examine cell proliferation in
A549 and H1299 cells treated with 0, 5, 10 or 20 µM gefitinib with
or without 12.5 or 25 µg/ml ginsenoside Rg3 for 48 h. The current
study demonstrated that in both NSCLC cell lines, gefitinib and
ginsenoside Rg3 inhibited cell proliferation in a dose-dependent
manner. The combined treatment with gefitinib and ginsenoside Rg3
further increased the cytotoxic effect compared with gefitinib or
ginsenoside Rg3 treatment alone (Fig. 1A
and B). In addition, A549 and H1299 cell viability was examined
following treatment with 10 µM gefitinib with or without 12.5 µg/ml
ginsenoside Rg3 for 24, 48 and 72 h. The current study demonstrated
the combined treatment with gefitinib and ginsenoside Rg3 increased
the cytotoxic effect compared with gefitinib or ginsenoside Rg3
treatment alone, and the difference was statistically significant
at 48 and 72 h (Fig. 1C and D).
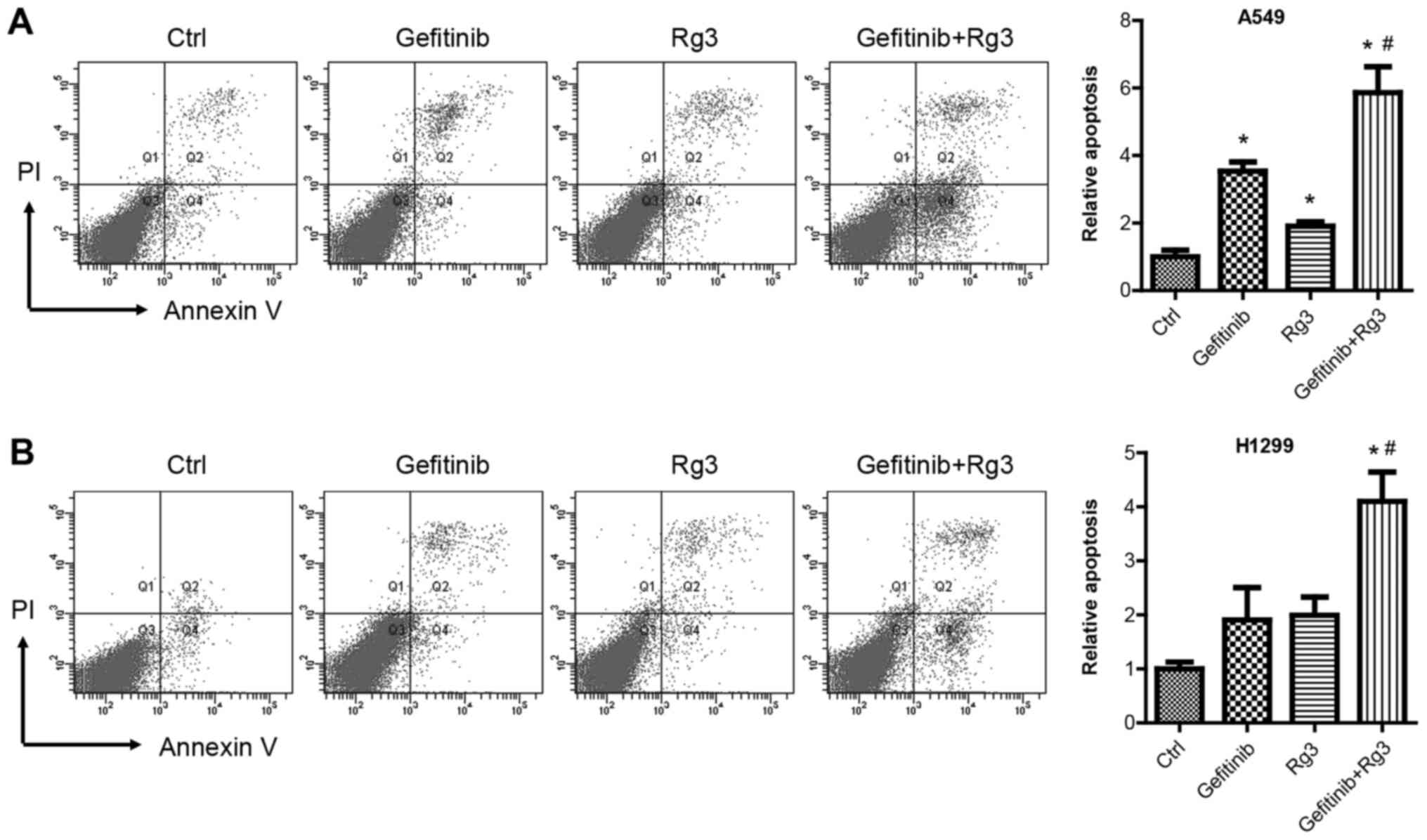
Effect of ginsenoside Rg3 on
gefitinib-induced apoptosis in NSCLC cancer cell lines
Flow cytometric analysis using Annexin V and PI
double staining was used to examine cell apoptosis in A549 and
H1299 cells treated with 10 µM gefitinib with or without 12.5 mg/ml
ginsenoside Rg3 for 48 h. The current study demonstrated that
gefitinib and Rg3 treatment significantly increased A549 cell
apoptosis compared with the control group. In addition, the
combined treatment with gefitinib + Rg3 significantly enhanced
gefitinib-induced apoptosis in A549 cells (Fig. 2A). Similarly, gefitinib or
ginsenoside Rg3 treatment alone increased H1299 cell apoptosis. The
combined treatment with gefitinib + Rg3 significantly enhanced cell
apoptosis compared with the control and gefitinib groups in H1299
cells (Fig. 2B).
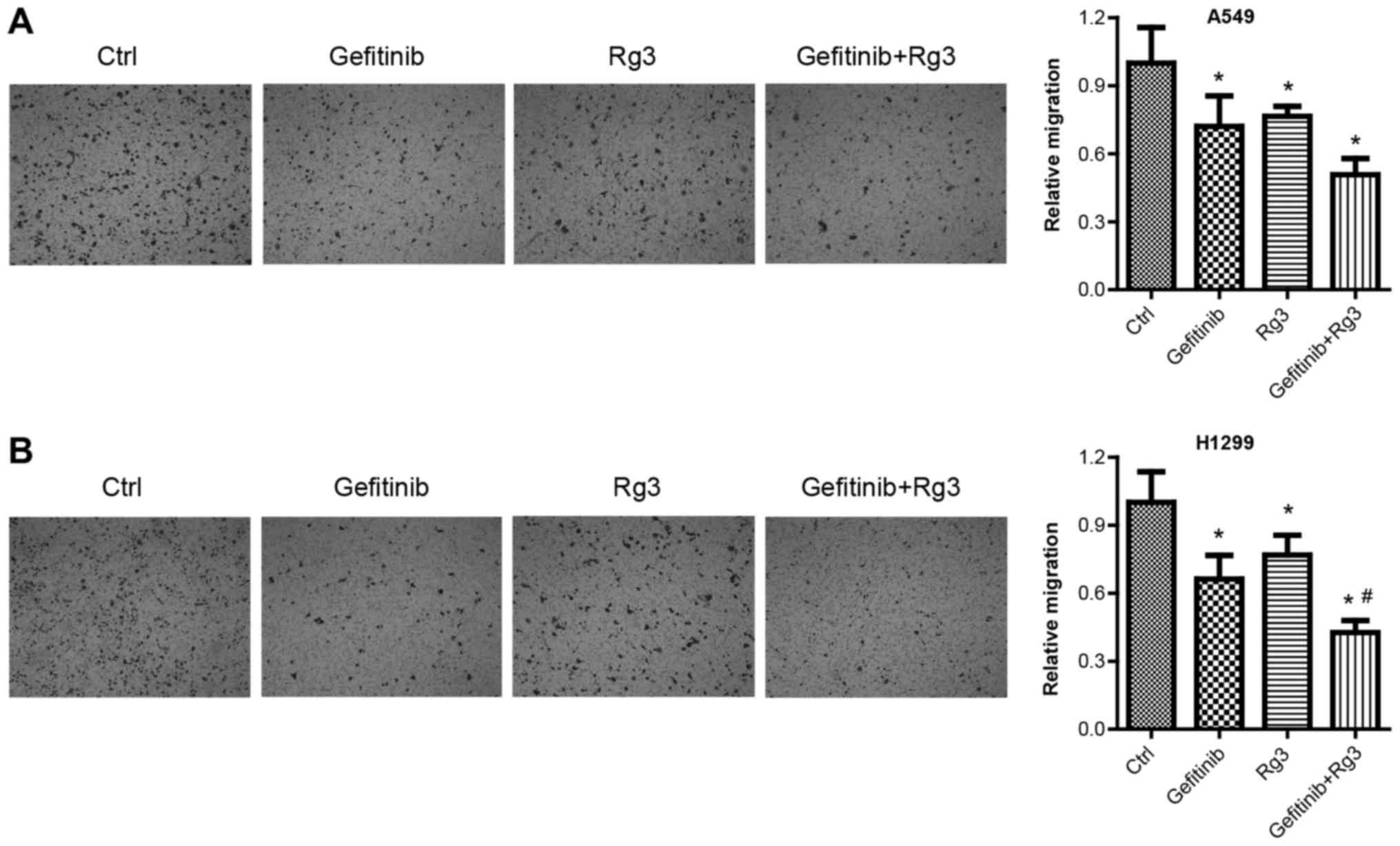
Ginsenoside Rg3 enhances the
inhibitory effect of gefitinib on NSCLC cell migration
Cell migration assays were used to examine migration
in A549 and H1299 cells treated with 10 µM gefitinib with or
without 12.5 µg/ml ginsenoside Rg3 for 24 h. Cell migration assays
were performed after a relatively short treatment time (24 h) to
exclude the influence of drug-induced cell death. The results from
the migration assays demonstrated that gefitinib, ginsenoside Rg3
and gefitinib + Rg3 treatment significantly decreased A549 cell
migration compared with the control group. (Fig. 3A). Similarly, the migration assays
demonstrated that gefitinib or ginsenoside Rg3 treatment alone
significantly decreased H1299 cell migration compared with the
control group. In addition, the combined treatment with gefitinib +
Rg3 significantly increased H1299 cell migration compared with the
control and gefitinib-treated group (Fig. 3B).
Ginsenoside Rg3 enhances the effects
of gefitinib on the expression of migration- and
apoptosis-associated proteins in NSCLC cell lines
To investigate the effects of treatment with
gefitinib and ginsenoside Rg3 on apoptosis- and
migration-associated pathways, the protein expression levels of
apoptosis-associated proteins (Bax, cleaved-caspase-3 and Bcl-2)
and migration-associated proteins (E-cadherin, SNAIL and SLUG) were
determined by western blot analysis in A549 and H1299 cells treated
with 10 µM gefitinib with or without 12.5 µg/ml ginsenoside Rg3 for
24 h. Cell death causes the degradation of cellular proteins,
including pro-apoptotic proteins (20). Therefore, protein expression was
examined after a relatively short treatment time (24 h) to exclude
the influence of drug-induced cell death. The current study
demonstrated that in both NSCLC cell lines, the combined treatment
with gefitinib + Rg3 increased protein expression levels of
pro-apoptotic proteins Bax and cleaved caspase-3, whilst the
protein expression level of the anti-apoptotic protein Bcl-2 was
decreased compared with the control group. Furthermore, the
combined treatment with gefitinib and Rg3 increased the protein
expression level of anti-migration protein E-cadherin, whilst the
protein expression levels of two pro-migration factors SNAIL and
SLUG were decreased compared with the control group (Fig. 4A and B). These results suggest that
ginsenoside Rg3 may enhance gefitinib-induced apoptosis and
inhibition of migration in NSCLC cell lines.
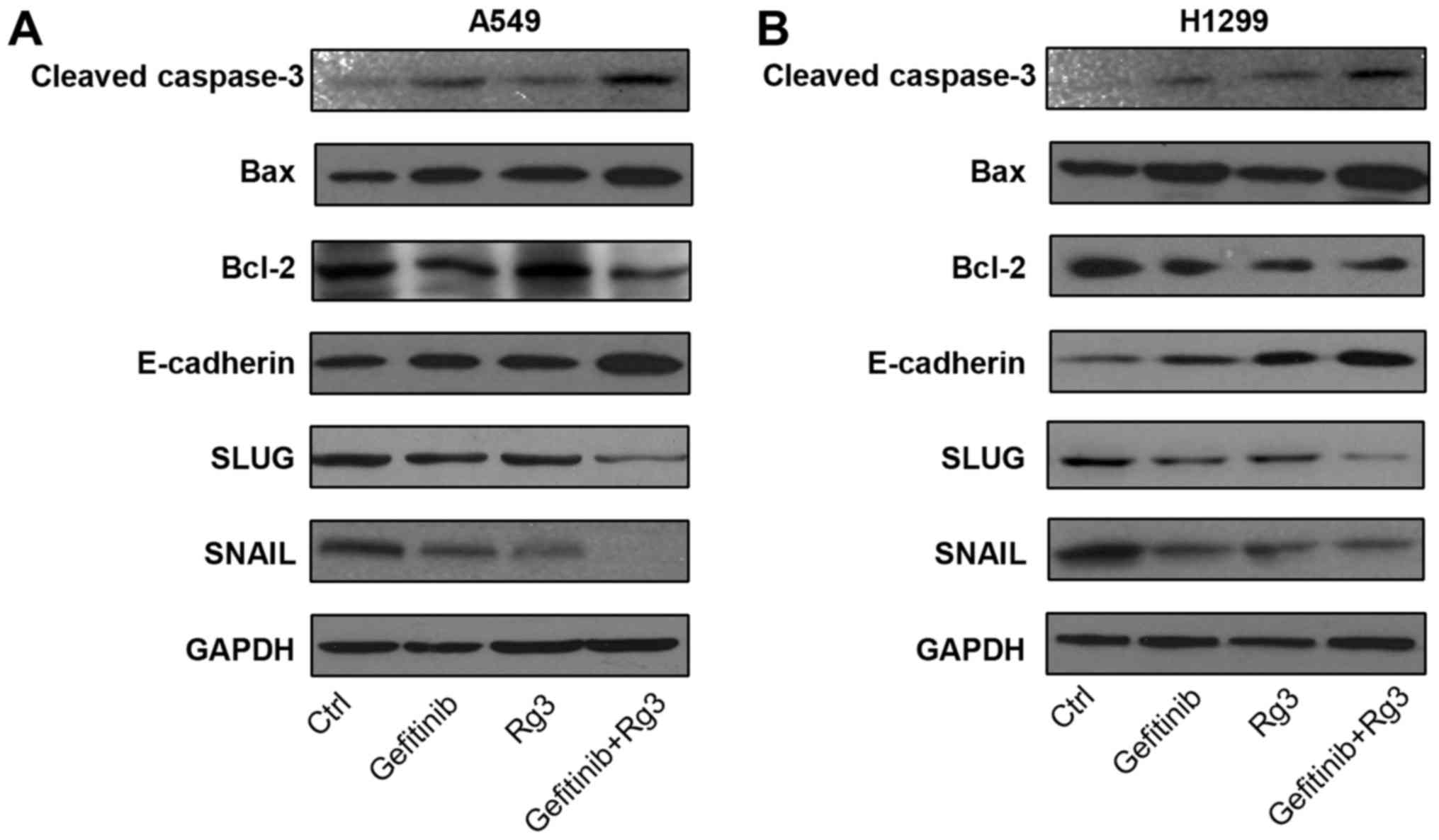 | Figure 4.Expression of migration and
apoptosis-associated proteins in lung cancer cell lines following
treatment with gefitinib with or without Rg3. Protein expression
levels of caspase-3, Bax, Bcl-2, E-cadherin, SLUG, SNAIL and GAPDH
were determined using western blot analysis in (A) A549 and (B)
H1299 cells treated with gefitinib (10 µM) with or without Rg3
(12.5 µg/ml) for 24 h. Rg3, ginsenoside Rg3; H1299 and A549,
non-small cell lung cancer cell lines. Bax, Bcl-2-associated X;
Bcl-2, B-cell lymphoma 2; SLUG, snail family transcriptional
repressor 2; SNAIL, snail family transcriptional repressor 1. |
Discussion
The current study demonstrated that ginsenoside Rg3
enhances gefitinib-induced tumor cytotoxicity and apoptosis, as
well as the inhibitory effect of gefitinib on cell migration,
thereby sensitizing NSCLC cells to gefitinib. The 5-year survival
rate for patients with NSCLC is approximately 15% (21,22). The
discovery and use of EGFR-TKIs, including gefitinib, has improved
prognosis in patients with advanced EGFR mutation-positive NSCLC
(6,23,24).
However, the majority of patients with NSCLC develop acquired
gefitinib resistance within 9–16 months, as a result of secondary
EGFR mutations (25,26). Second-generation EGFR-TKIs, including
afatinib, were developed to overcome acquired resistance to
first-generation inhibitors (27).
Third-generation EGFR-TKIs can target the constitutive activation
of EGFR mutations as well as resistant mutations (28,29).
Issues regarding high cost, side effects and limited applicability
of second- and third-generation EGFR-TKIs have prevented the
widespread clinical application of these drugs (27,30). To
overcome the limitations associated with EGFR-TKIs in the treatment
with NSCLC, more efficient therapeutic strategies with fewer side
effects are required. The current study demonstrated that
ginsenoside Rg3 increased the cytotoxic activity of gefitinib in
NSCLC cell. These results suggest that the combined treatment with
gefitinib and ginsenoside Rg3 may be used for non-EGFR mutant
cancer at a lower dose, whilst reducing any potential side effects
associated with EGFR-TKIs.
Gefitinib is an effective treatment option for
patients with advanced EGFR mutation-positive NSCLC (9). However numerous patients with NSCLC do
not have gefitinib-sensitive mutations and therefore do not respond
to treatment with gefitinib (13).
In the present study, two NSCLC cell lines (H1299 and A549) with
wild-type EGFR (31) and different
sensitivities to gefitinib were selected and used to investigate
the enhanced efficacy and sensitivity of gefitinib with ginsenoside
Rg3. The current study demonstrated that both NSCLC cell lines
could be re-sensitized to treatment with gefitinib when combined
with ginsenoside Rg3. Furthermore, the different p53 states of A549
(p53-wildtype) and H1299 (p53-null) cells (31), suggests that p53 is unlikely to be
involved in the potential underlying process.
Ginsenoside Rg3 is a natural product extracted from
a traditional Chinese medicine, Panax ginseng (17). Several studies have suggested that
ginsenoside Rg3 may serve roles in the complex process of tumor
development, which includes proliferation, apoptosis, migration,
angiogenesis and tumor immunogenicity (18,19,32–36).
Unlike targeted drugs, ginsenoside Rg3 has multiple targets and
exhibits anticancer activity through a number of mechanisms, which
include targeting multiple tumor-associated signaling pathways as
well as regulating intracellular reactive oxygen species (34–36). The
potential mechanism of action of ginsenoside Rg3 is not dependent
on EGFR mutations (37), which
suggests that an enhanced therapeutic efficiency may be achieved
through the combined treatment with gefitinib. Consistent with
previous studies (34,35), the current study demonstrated that
ginsenoside Rg3 inhibited cell proliferation, induced cell
apoptosis and decreased NSCLC cell migration.
The current study demonstrated that ginsenoside Rg3
enhanced gefitinib-induced tumor cytotoxicity in NSCLC cells and
the inhibitory effect of gefitinib on NSCLC cell migration. Western
blot analysis demonstrated that the combined treatment with
gefitinib and ginsenoside Rg3 enhanced protein expression levels of
cell migration- and apoptosis-associated markers in NSCLC cell
lines which suggests that ginsenoside Rg3 may enhance gefitinib
efficacy in NSCLC. Several studies revealed that ginsenoside Rg3
reversed resistance to cisplatin in lung cancer (18,19,36). In
the current study, ginsenoside Rg3 enhanced gefitinib efficiency in
NSCLC cell proliferation, apoptosis, and migration. The mechanisms
of action of ginsenoside Rg3 may be distinct from gefitinib
(17), exerting a synchronous
inhibitory effect on EGFR. However, under different dose
combinations, gefitinib and ginsenoside Rg3 may exert a potential
synergistic anticancer effect.
The current study has several limitations. Two NSCLC
cell lines were used to investigate the antitumor effects of
ginsenoside Rg3 and gefitinib, therefore NSCLC cell lines with
different EGFR status should be used for further investigation. In
addition, the underlying mechanism of ginsenoside Rg3 and gefitinib
in NSCLC remains unknown and should be further investigated.
In conclusion, ginsenoside Rg3 may be able to
enhance the anticancer activity of gefitinib. Ginsenoside Rg3
enhanced gefitinib-induced cytotoxicity, apoptosis and migration
inhibition, making NSCLC cells more sensitive to gefitinib. These
results indicated the potential clinical application of the
combined treatment with gefitinib and ginsenoside Rg3 for patients
with NSCLC.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
YD, WW and JT designed the experiments and analyzed
the data. QS performed flow cytometric experiments. YD and WW wrote
the manuscript. JT supervised the work.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Wong MCS, Lao XQ, Ho KF, Goggins WB and
Tse SLA: Incidence and mortality of lung cancer: Global trends and
association with socioeconomic status. Sci Rep. 7:143002017.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ferlay J, Soerjomataram I, Dikshit R, Eser
S, Mathers C, Rebelo M, Parkin DM, Forman D and Bray F: Cancer
incidence and mortality worldwide: Sources, methods and major
patterns in GLOBOCAN 2012. In J Cancer. 136:E359–E386. 2015.
|
|
3
|
Zhou M, Wang H, Zhu J, Chen W, Wang L, Liu
S, Li Y, Wang L, Liu Y and Yin P: Cause-specific mortality for 240
causes in China during 1990–2013: A systematic subnational analysis
for the global burden of disease study 2013. Lancet. 387:251–272.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Zappa C and Mousa SA: Non-small cell lung
cancer: Current treatment and future advances. Transl Lung Cancer
Res. 5:288–300. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ramalingam SS, Owonikoko TK and Khuri FR:
Lung cancer: New biological insights and recent therapeutic
advances. CA Cancer J Clin. 61:91–112. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Prabhu VV and Devaraj N: Epidermal growth
factor receptor tyrosine kinase: A potential target in treatment of
non-small-cell lung carcinoma. J Environ Pathol Toxicol Oncol.
36:151–158. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Goss G, Tsai CM, Shepherd FA, Bazhenova L,
Lee JS, Chang GC, Crino L, Satouchi M, Chu Q, Hida T, et al:
Osimertinib for pretreated EGFR Thr790Met-positive advanced
non-small-cell lung cancer (AURA2): A multicentre, open-label,
single-arm, phase 2 study. Lancet Oncol. 17:1643–1652. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Solomon BJ, Mok T, Kim DW, Wu YL, Nakagawa
K, Mekhail T, Felip E, Cappuzzo F, Paolini J, Usari T, et al:
First-line crizotinib versus chemotherapy in ALK-positive lung
cancer. N Engl J Med. 371:2167–2177. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ghafoor Q, Baijal S, Taniere P, O'Sullivan
B, Evans M and Middleton G: Epidermal growth factor receptor (EGFR)
kinase inhibitors and non-small cell lung cancer (NSCLC)-advances
in molecular diagnostic techniques to facilitate targeted therapy.
Pathol Oncol Res. 24:723–731. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lynch TJ, Bell DW, Sordella R,
Gurubhagavatula S, Okimoto RA, Brannigan BW, Harris PL, Haserlat
SM, Supko JG, Haluska FG, et al: Activating mutations in the
epidermal growth factor receptor underlying responsiveness of
non-small-cell lung cancer to gefitinib. N Engl J Med.
350:2129–2139. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Maemondo M, Inoue A, Kobayashi K, Sugawara
S, Oizumi S, Isobe H, Gemma A, Harada M, Yoshizawa H, Kinoshita I,
et al: Gefitinib or chemotherapy for non-small-cell lung cancer
with mutated EGFR. N Engl J Med. 362:2380–2388. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Li Y, Wang Y, Niu K, Chen X, Xia L, Lu D,
Kong R, Chen Z, Duan Y and Sun J: Clinical benefit from EGFR-TKI
plus ginsenoside Rg3 in patients with advanced non-small cell lung
cancer harboring EGFR active mutation. Oncotarget. 7:70535–70545.
2016.PubMed/NCBI
|
|
13
|
Li ZX, Qu LY, Wen H, Zhong HS, Xu K, Qiu
XS and Wang EH: Mig-6 overcomes gefitinib resistance by inhibiting
EGFR/ERK pathway in non-small cell lung cancer cell lines. Int J
Clin Exp Pathol. 7:7304–7311. 2014.PubMed/NCBI
|
|
14
|
Zheng R, Jiang H, Li J, Liu X and Xu H:
Polyphyllin II restores sensitization of the resistance of PC-9/ZD
cells to gefitinib by a negative regulation of the PI3K/Akt/mTOR
signaling pathway. Curr Cancer Drug Target. 17:376–385. 2017.
View Article : Google Scholar
|
|
15
|
Song S, Du L, Jiang H, Zhu X, Li J and Xu
J: Paris saponin I sensitizes gastric cancer cell lines to
cisplatin via cell cycle arrest and apoptosis. Med Sci Monit.
22:3798–3803. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Jiang H, Zhao PJ, Su D, Feng J and Ma SL:
Paris saponin I induces apoptosis via increasing the Bax/Bcl-2
ratio and caspase-3 expression in gefitinib-resistant non-small
cell lung cancer in vitro and in vivo. Mol Med Rep.
9:2265–2272. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Sun M, Ye Y, Xiao L, Duan X, Zhang Y and
Zhang H: Anticancer effects of ginsenoside Rg3 (Review). Int J Mol
Med. 39:507–518. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Jiang Z, Yang Y, Yang Y, Zhang Y, Yue Z,
Pan Z and Ren X: Ginsenoside Rg3 attenuates cisplatin resistance in
lung cancer by downregulating PD-L1 and resuming immune. Biomed
Pharmacother. 96:378–383. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kim YJ, Choi WI, Jeon BN, Choi KC, Kim K,
Kim TJ, Ham J, Jang HJ, Kang KS and Ko H: Stereospecific effects of
ginsenoside 20-Rg3 inhibits TGF-beta1-induced
epithelial-mesenchymal transition and suppresses lung cancer
migration, invasion and anoikis resistance. Toxicology. 322:23–33.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Elmore S: Apoptosis: A review of
programmed cell death. Toxicol Pathol. 35:495–516. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Socinski MA, Evans T, Gettinger S, Hensing
TA, VanDam Sequist L, Ireland B and Stinchcombe TE: Treatment of
stage IV non-small cell lung cancer: Diagnosis and management of
lung cancer, 3rd ed: American college of chest physicians
evidence-based clinical practice guidelines. Chest. 143 Suppl
5:e341S–e368S. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2016. CA Cancer J Clin. 66:7–30. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Baselga J and Arteaga CL: Critical update
and emerging trends in epidermal growth factor receptor targeting
in cancer. J Clin Oncol. 23:2445–2459. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Bareschino MA, Schettino C, Rossi A,
Maione P, Sacco PC, Zeppa R and Gridelli C: Treatment of advanced
non small cell lung cancer. J Thorac Dis. 3:122–133.
2011.PubMed/NCBI
|
|
25
|
Su KY, Chen HY, Li KC, Kuo ML, Yang JC,
Chan WK, Ho BC, Chang GC, Shih JY, Yu SL and Yang PC: Pretreatment
epidermal growth factor receptor (EGFR) T790M mutation predicts
shorter EGFR tyrosine kinase inhibitor response duration in
patients with non-small-cell lung cancer. J Clin Oncol. 30:433–440.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Xu C, Zhou Q and Wu YL: Can EGFR-TKIs be
used in first line treatment for advanced non-small cell lung
cancer based on selection according to clinical factors?-A
literature-based meta-analysis. J Hematol Oncol. 5:622012.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Hirsh V: Next-Generation covalent
irreversible kinase inhibitors in NSCLC: Focus on afatinib.
BioDrugs. 29:167–183. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Tan CS, Gilligan D and Pacey S: Treatment
approaches for EGFR-inhibitor-resistant patients with
non-small-cell lung cancer. Lancet Oncol. 16:e447–e459. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Yver A: Osimertinib (AZD9291)-a
science-driven, collaborative approach to rapid drug design and
development. Ann Oncol. 27:1165–1170. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Engel J, Lategahn J and Rauh D: Hope and
disappointment: Covalent inhibitors to overcome drug resistance in
non-small cell lung cancer. ACS Med Chem Lett. 7:2–5. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Rho JK, Choi YJ, Ryoo BY, Na II, Yang SH,
Kim CH and Lee JC: p53 enhances gefitinib-induced growth inhibition
and apoptosis by regulation of Fas in non-small cell lung cancer.
Cancer Res. 67:1163–1169. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Kim JW, Jung SY, Kwon YH, Lee JH, Lee YM,
Lee BY and Kwon SM: Ginsenoside Rg3 attenuates tumor angiogenesis
via inhibiting bioactivities of endothelial progenitor cells.
Cancer Biol Ther. 13:504–515. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Son KJ, Choi KR, Lee SJ and Lee H:
Immunogenic cell death induced by ginsenoside Rg3: Significance in
dendritic cell-based anti-tumor immunotherapy. Immune Netw.
16:75–84. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Sun HY, Lee JH, Han YS, Yoon YM, Yun CW,
Kim JH, Song YS and Lee SH: Pivotal roles of ginsenoside Rg3 in
tumor apoptosis through regulation of reactive oxygen species.
Anticancer Res. 36:4647–4654. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Tian L, Shen D, Li X, Shan X, Wang X, Yan
Q and Liu J: Ginsenoside Rg3 inhibits epithelial-mesenchymal
transition (EMT) and invasion of lung cancer by down-regulating
FUT4. Oncotarget. 7:1619–1632. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Wang J, Tian L, Khan MN, Zhang L, Chen Q,
Zhao Y, Yan Q, Fu L and Liu J: Ginsenoside Rg3 sensitizes hypoxic
lung cancer cells to cisplatin via blocking of NF-κB mediated
epithelial-mesenchymal transition and stemness. Cancer Lett.
415:73–85. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Joo EJ, Chun J, Ha YW, Ko HJ, Xu MY and
Kim YS: Novel roles of ginsenoside Rg3 in apoptosis through
downregulation of epidermal growth factor receptor. Chem Biol
Interact. 233:25–34. 2015. View Article : Google Scholar : PubMed/NCBI
|