Introduction
Henoch-Schönlein purpura (HSP) is the most common
type of systemic small-vessel vasculitis in pediatric patients, and
is associated with multiple and complex pathogenic factors, as well
as diverse pathological damage (1).
It is a systemic disorder characterized by leukocytoclastic
vasculitis involving the capillaries and deposition of
immunoglobulin (Ig)A immune complexes (2). Of all pediatric patients with HSP,
>90% are <10 years old (3,4). Between
weeks 4 and 6 of the initial disease presentation, ~40% of
pediatric patients with HSP progress to HSP nephritis (HSPN), which
is one of the major manifestations and the primary cause of
mortality associated with HSP (2,5,6). To date, however, the exact
pathophysiology of HSPN has remained largely elusive and requires
further investigation.
Long non-coding RNAs (lncRNAs) are a class of RNA
with a length of >200 nucleotides and no coding function. In
recent years, as the rapid development of technologies has
facilitated the analysis of the ‘transcriptome’ the study of
lncRNAs has enabled the discovery of comprehensive genetic
information in the human genome. A large amount of evidence has
suggested that lncRNAs regulate protein-coding genes at the
transcriptional and post-transcriptional levels, and exert
transcription control (7,8). Numerous studies have provided novel
insight into different expression profiles of lncRNAs in a number
of human kidney diseases, including acute kidney rejection,
diabetic nephropathy, membranous nephropathy, chronic kidney
disease and lupus nephritis (9–15). In
parallel with their role in disease pathogenesis, lncRNAs may also
serve as a potential biomarker of disease status and aid in the
diagnosis, prognosis and clinical management of disease (16,17).
However, the expression patterns and functions of lncRNAs in HSPN
have largely remained to be elucidated.
In the present study, high-throughput sequencing was
applied to identify 820 lncRNAs and 3,557 mRNAs that are
significantly aberrantly expressed in the peripheral blood of
pediatric patients with HSPN. Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) provided results that were
consistent with those obtained by data analysis of the gene
expression profiles, thereby verifying them. Gene Ontology (GO) and
Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway analyses
were then performed to elucidate the roles and pathways of the
differentially expressed RNAs. These results indicated that the
aberrantly expressed RNAs may have important roles in the
development of HSPN through promoting serum proteins generation and
regulating the apoptosis pathway, and that knowing the differently
expressed RNAs might provide useful biomarkers for HSPN therapy and
diagnosis.
Patients and methods
Patients and sample collection
A total of 6 pediatric patients with HSPN (4 males
and 2 females; mean age, 12.17±1.72 years) were recruited at the
Affiliated Hospital of Liaoning University of Traditional Chinese
Medicine (Shenyang, China) between November 2016 and March 2017.
The diagnostic criteria for HSPN were according to those outlined
at the Congress of the Chinese Pediatric Society in 2000 (18). Patients with other coexisting renal
pathologies were excluded. None of the patients of the present
study was diagnosed with any other complications. Furthermore, none
of the subjects had taken any hormonal or immunosuppressive drugs
for ≥6 months prior to the commencement of the study. The clinical
characteristics of the patients with HSPN are presented in Table I. The mean urine red blood cell
count, 24-h urine protein quantity and serum IgA were 387.48±590.34
p/µl, 0.28±0.15 g/24 h and 3.00±1.21 g/l, respectively. A total of
4 age matched healthy subjects (1 male and 3 females; mean age,
11.25±2.99 years) were selected as healthy controls (HC). Blood
samples were obtained from the 6 children with HSPN and 4 healthy
volunteers.
 | Table I.Clinical characteristics of 6 HSPN
cases. |
Table I.
Clinical characteristics of 6 HSPN
cases.
| Patient ID | S64 | S87 | S105 | S111 | S173 | S303 |
|---|
| Age (years) | 14 | 13 | 10 | 14 | 11 | 11 |
| Sex | F | F | M | M | M | M |
| Purpura | + | + | + | + | + | + |
| Abdominal pain | − | + | − | − | − | + |
| Arthralgia | − | + | + | − | + | + |
| Period between the
onset of HSPN | 12 | 10 | 34 | 16 | 60 | 20 |
| and initiation of
therapy (months) |
The present study was approved by the Ethics
Committee of the Institutional Ethics Board of the Affiliated
Hospital of Liaoning University of Traditional Chinese Medicine
(approval no. 2016CS(KT)-002-01). Written informed consent was
obtained from the parents of all of the pediatric subjects enrolled
in the present study.
RNA extraction and library
construction
Peripheral blood was obtained from the subjects of
the groups and collected in tubes containing EDTA as an
anticoagulant. Peripheral blood total RNA was isolated using a
total RNA isolation kit purchased from BioTeke Corp. (Beijing,
China), according to the manufacturer's protocols. From the RNA (1
µg) the ribosomal (r)RNA was removed using a Ribo-Zero rRNA Removal
kit (Illumina, Inc., San Diego, CA, USA), according to the
manufacturer's protocols. RNA libraries were constructed using
rRNA-depleted RNAs with a TruSeq Stranded Total RNA Library Prep
kit (Illumina, Inc.), according to the manufacturer's protocols.
Libraries were controlled for quality and were quantified using a
BioAnalyzer 2100 system (Agilent Technologies, Inc., Santa Clara,
CA, USA).
High-throughput sequencing and
computational analysis
A DNA library (420 µl of a 10 pM library) was
denatured as single-stranded DNA molecules, captured on Illumina
flow cells, amplified in situ as clusters and finally
sequenced for 150 cycles on an Illumina Hiseq Sequencer, according
to the manufacturer's protocols. High-throughput sequencing was
performed by Shanghai Cloud-Seq Biotech, Inc., (Shanghai, China).
In brief, paired-end reads were harvested from the Illumina HiSeq
2000 sequencer (Illumina, Inc.), and were quality controlled via
their Q score (>Q30). Following 3′ adaptor-trimming and removal
of low-quality reads with cutadapt software (v1.9.3) (19), the high-quality, trimmed reads were
aligned to a reference genome (UCSC HG19) guided by the Ensembl GFF
gene annotation file with hisat2 software (v2.0.4; http://ccb.jhu.edu/software/hisat2/index.shtml). Next,
cuffdiff software (v2.2.1, part of cufflinks; http://cufflinks.cbcb.umd.edu/) was used to
obtain the gene fragment counts per kilobase of exon per million
fragments mapped (FPKM) as the expression profiles of lncRNA and
mRNA, and the fold change (FC) and P-value were calculated based on
the FPKM, from which differentially expressed lncRNAs and mRNAs
were identified.
GO term analysis and pathway
analysis
The GO (www.geneontology.org) and KEGG (www.genome.ad.jp/kegg) databases were utilized to
analyze biological functions and signaling pathways based on the
differentially expressed lncRNAs and mRNAs. Fisher's exact test was
used to determine whether there was more overlap between the gene
list and the GO annotation list than that expected to occur by
chance. The P-value denoted the significance of the GO term
enrichment and was the basis of the significance level, with
P<0.05 set as the threshold. Pathway analysis was used to
investigate the major pathways of the aberrantly expressed genes
according to the KEGG database. The threshold for the false
discovery rate was set at 0.05, and P<0.05 was considered to
indicate a statistically significant difference.
RT-qPCR validation
Total RNA was extracted from the peripheral blood
using TRIzol reagent (Invitrogen; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA), according to the manufacturer's protocols. In
brief, 1 µg total RNA from each sample was used for the synthesis
of first strand complementary DNA using a SuperScript™
III First-Strand Synthesis System kit (Invitrogen; Thermo Fisher
Scientific, Inc.), according to the manufacturer's instructions.
qPCR was performed on an Applied Biosystems ViiA™ 7
Real-time PCR System using the SYBR-Green method (both Thermo
Fisher Scientific, Inc.), according to the manufacturer's protocol.
The thermocycling conditions were as follows: A denaturation step
at 95°C for 10 min, followed by 40 cycles of 95°C for 10 sec and
60°C for 60 sec. The primer sequences are presented in Table II. Relative gene expression levels
were quantified using the 2−∆∆Cq method, in which
β-actin (ACTB) was used as an internal control (20).
| Table II.Primers used for reverse
transcription-quantitative polymerase chain reaction. |
Table II.
Primers used for reverse
transcription-quantitative polymerase chain reaction.
| Gene
name/direction | Sequence (5′ to
3′) | Product length
(bp) |
|---|
|
ENSG00000267121 |
|
|
| F |
GAGGAAGACCCTGGAAGGAG | 203 |
| R |
GTCCCAAGCTTCAGTCATCC |
|
|
ENSG00000252310 |
|
|
| F |
GGTCCGAGTGTTGTGGGTTA | 50 |
| R |
GGGGGAGACAATGTTAAATCAA |
|
| uc001kfc.1 |
|
|
| F |
AAAATTAGCCAGGCATGGTG | 209 |
| R |
TCTCTCACGGCTCTTGTGTG |
|
| uc010qna.2 |
|
|
| F |
GGGTCTTCCTCATGGCACTA | 202 |
| R |
CAGGCCTTCCAAGTTCTGAG |
|
|
ENST00000378432 |
|
|
| F |
CCTTTTCTCCATGGCATTTG | 202 |
| R |
TCCTGCATTCATTCATTCCA |
|
|
ENST00000571370 |
|
|
| F |
GGTTGTTTCATTCCGCAGTT | 204 |
| R |
TTTCTGGGACGATGAAAAGG |
|
| ACTB |
|
|
| F |
GGCCTCCAAGGAGTAAGACC | 73 |
| R |
AGGGGAGATTCAGTGTGGTG |
|
Statistical analysis
Statistical analysis and graphic presentation were
performed with SPSS version 19.0 software packages (IBM Corp.,
Armonk, NY, USA). Measurement data are expressed as the mean ±
standard deviation. The differences in expression levels of tested
lncRNAs and mRNAs between two groups were assessed using Student's
t-test. P<0.05 was considered to indicate a statistical
significant difference. FC≥1.5 indicated upregulation and <0.67
indicated downregulation. were considered to indicate significant
differences. Otherwise, the non-parametric Mann-Whitney U test was
used to analyze the data. Fisher's exact test was used for GO
analysis and KEGG pathway analysis. P<0.05 was considered to
indicate a statistically significant difference.
Results
Subject characteristics
The baseline demographic and clinical data of the
subjects in the HSPN group and the HC group are summarized in
Table III. The pediatric patients
with HSPN had a significantly higher urine red blood cell count,
24-h urine protein quantity and serum IgA than the HC group.
| Table III.Baseline demographic and clinical
data of subjects. |
Table III.
Baseline demographic and clinical
data of subjects.
| Parameter | HSPN group
(n=6) | HC group (n=4) | Normal range |
|---|
| Sex (male/female),
n | 4:2 | 1:3 |
|
| Age (years, mean ±
SD) | 12.17±1.72 | 11.25±2.99 |
|
| Serum IgA (g/l,
mean ± SD) | 3.00±1.21 | 1.39±0.55 | 0.63–1.79 |
| Urine red blood
cell count (p/µl, mean ± SD) | 387.48±590.34 | 3.60±8.10 | 0-25.00 |
| 24-h urine protein
quantity (g/24 h, mean ± SD) | 0.280±0.150 | 0.050±0.007 | 0–0.150 |
Differentially expressed lncRNAs
A total of 820 lncRNAs were aberrantly expressed
between the HSPN group and the HC group. High-throughput sequencing
revealed that 34 lncRNAs were upregulated and 786 lncRNAs were
downregulated. The top 25 differentially expressed lncRNAs
according to their FC values in the HSPN vs. HC group are presented
in Table IV. Hierarchical
clustering was performed according to the lncRNA expression levels
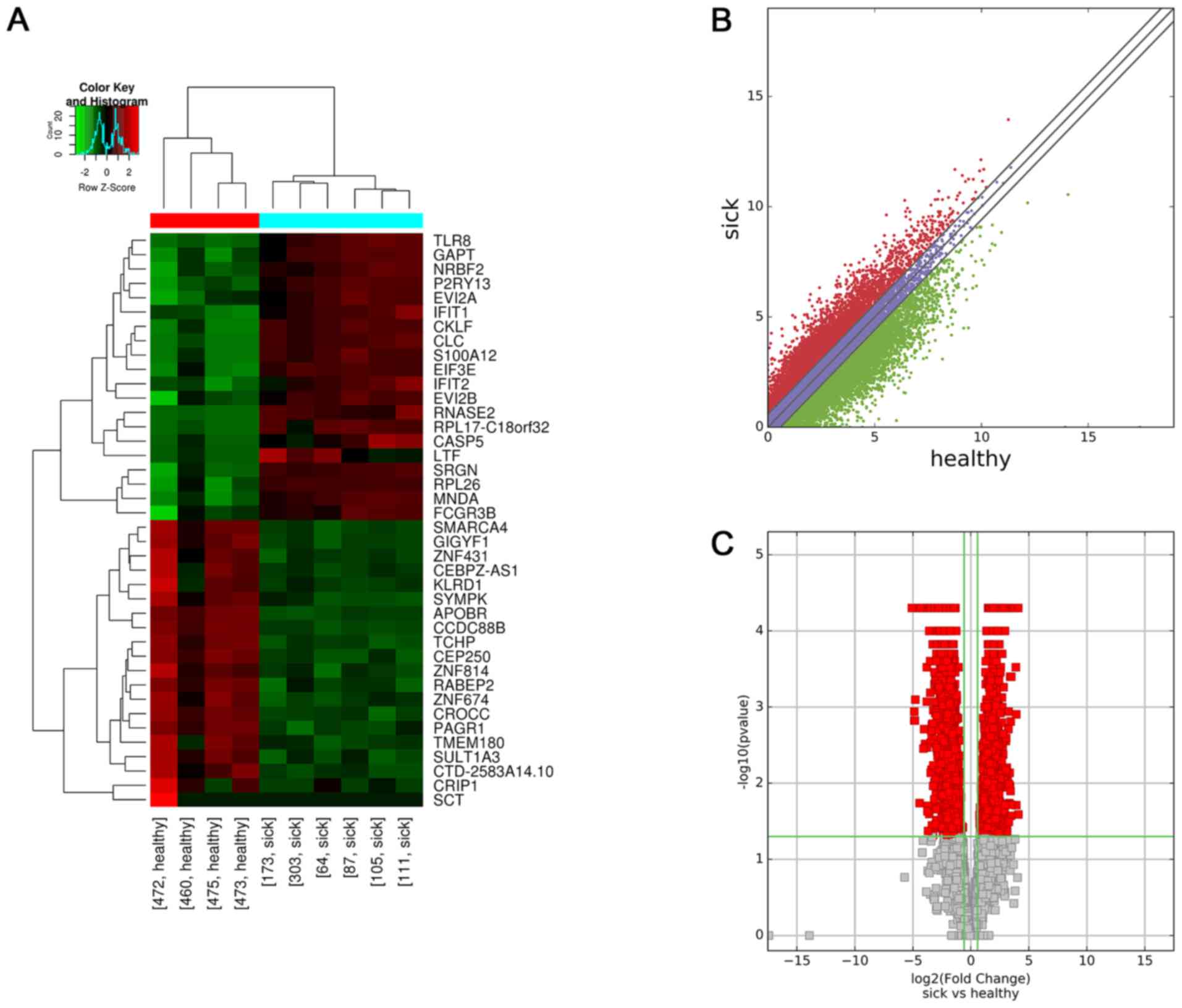
in these 10 samples (Fig. 1A).
Scatter and volcano plots were used to assess variations in lncRNA
expression between the two groups (Fig.
1B and C, respectively).
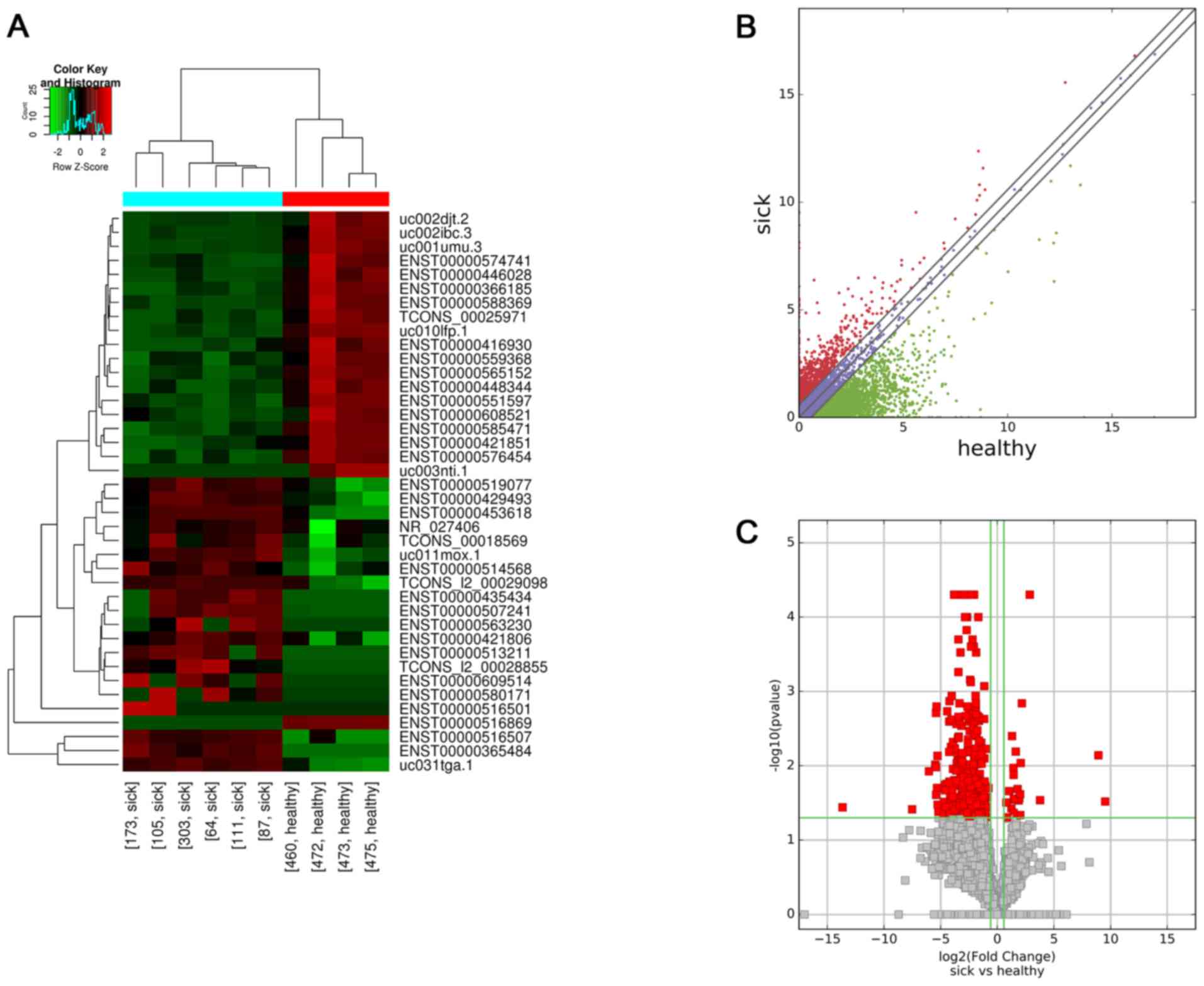 | Figure 1.Profiles of differentially expressed
lncRNAs between the two groups. (A) Hierarchical clustering
analysis of 40 lncRNAs that were aberrantly expressed between
children with HSPN (sick173, sick105, sick303, sick64, sick111 and
sick87) and HC (healthy460, healthy472, healthy473 and healthy475).
Red and green indicate high and low relative expression,
respectively. P<0.05 was considered to indicate a statistical
significant difference. FC≥1.5 indicated upregulation and <0.67
indicated downregulation. (B) Scatter plot of aberrant expression
of lncRNAs between the HSPN and HC libraries. Red indicates
upregulated lncRNAs [log2(HSPN/HC)>0], blue indicates
lncRNAs with no change in expression between HSPN and HC libraries,
green indicates downregulated lncRNAs
[log2(HSPN/HC)<0]. (C) Volcano plot of aberrant
lncRNA expression. The vertical green lines delineate a 1.5-fold
upregulation and 0.67-fold downregulation of lncRNAs, and the
horizontal line represents P=0.05. Red plots on the right side
represent 34 upregulated lncRNAs with FC≥1.5 and corrected
P<0.05. Red plots on the left side represent 786 down-regulated
lncRNAs with FC<0.67 and corrected P<0.05. lncRNA, long
non-coding RNA; HSPN, Henoch-Schönlein purpura nephritis; HC,
healthy controls; FC, fold change; NR, nitrate reductase. |
| Table IV.Top 25 aberrantly expressed lncRNAs
according to the FC values between the two groups. |
Table IV.
Top 25 aberrantly expressed lncRNAs
according to the FC values between the two groups.
| lncRNA ID | FC value | P-value | Class | Database |
|---|
| Upregulated |
|
|
|
|
|
ENSG00000202354 | 736.95 | 0.030 | Intergenic | Ensembl |
|
ENSG00000252310 | 487.59 | 0.007 | Intergenic | Ensembl |
| Downregulated |
|
|
|
|
|
ENSG00000259001 | −12971.66 | 0.036 | Bidirectional | Ensembl |
|
HLA-B | −182.99 | 0.039 | Sense | UCSC_knowngene |
|
ENSG00000257621 | −65.40 | 0.012 | Intronic | Ensembl |
|
ENSG00000262380 | −44.54 | 0.010 | Sense | Ensembl |
|
BC047651 | −42.38 | 0.010 | Sense | UCSC_knowngene |
|
ENSG00000267121 | −42.31 | 0.002 | Intergenic | Ensembl |
|
ENSG00000230105 | −42.22 | 0.025 | Intergenic | Ensembl |
|
DQ598910 | −41.06 | 0.002 | Sense | UCSC_knowngene |
|
ENSG00000236535 | −39.99 | 0.020 | Sense | Ensembl |
|
ENSG00000262879 | −38.70 | 0.007 | Intergenic | Ensembl |
|
BC044596 | −38.50 | 0.034 | Sense | UCSC_knowngene |
|
ENSG00000231485 | −36.49 | 0.041 | Intergenic | Ensembl |
|
ENSG00000260060 | −26.54 | 0.045 | Intergenic | Ensembl |
|
ENSG00000232593 | −26.14 | 0.031 | Intergenic | Ensembl |
|
ENSG00000246016 | −24.65 | 0.018 | Intergenic | Ensembl |
|
MGC72080 | −24.52 | 0.021 | Intergenic | UCSC_knowngene |
|
XLOC_012288 | −23.80 | 0.036 | Intergenic | TCONS |
|
ENSG00000247556 | −21.99 | 0.018 | Intergenic | Ensembl |
|
ENSG00000267672 | −21.24 | 0.002 | Sense | Ensembl |
|
ENSG00000227195 | −19.36 | 0.035 | Intergenic | Ensembl |
|
AX747098 | −19.05 | 0.018 | Sense | UCSC_knowngene |
|
AK098491 | −18.85 | 0.007 | Sense | UCSC_knowngene |
|
ENSG00000253430 | −18.81 | 0.003 | Sense | Ensembl |
Differentially expressed mRNAs
A total of 3,557 mRNAs that were aberrantly
expressed between the two groups were identified using the same
criteria as those for the lncRNAs. A total of 1,232 upregulated and
2,325 downregulated differentially expressed mRNAs were identified.
The top 25 differentially expressed mRNAs are listed in Table V. Hierarchical clustering was
performed according to the mRNA expression levels in all 10 samples
(Fig. 2A). Scatter and volcano plots
generated from these differentially expressed mRNAs exhibited a
clear segregation between the HSPN and HC groups (Fig. 2B and C, respectively).
| Table V.Top 25 differentially expressed mRNAs
according to the FC values between the two groups. |
Table V.
Top 25 differentially expressed mRNAs
according to the FC values between the two groups.
| Gene name | Ensembl ID | FC value | P-value | Chromosome |
|---|
| Upregulated |
|
RPL17-C18orf32 |
ENSG00000215472 | 17.16 | 0.026 |
chr18:47008027-47018906 |
|
MNDA |
ENSG00000163563 | 16.94 | <0.001 |
chr1:158801106-158819296 |
|
IFIT1 |
ENSG00000185745 | 15.09 | 0.001 |
chr10:90973325-91174314 |
| Downregulated |
|
ZNF431 |
ENSG00000196705 | −33.88 | <0.001 |
chr19:21324826-21373034 |
|
APOBR |
ENSG00000184730 | −29.51 | 0.001 |
chr16:28467692-28510291 |
|
CROCC |
ENSG00000058453 | −29.46 | 0.002 |
chr1:17066767-17299474 |
|
CTD-2583A14.10 |
ENSG00000268750 | −28.25 | 0.002 |
chr19:58281022-58427978 |
|
PAGR1 |
ENSG00000263136 | −27.74 | 0.001 |
chr16:29262828-30215631 |
|
KLRD1 |
ENSG00000134539 | −25.48 | <0.001 |
chr12:10378656-10469850 |
|
SYMPK |
ENSG00000125755 | −21.33 | <0.001 |
chr19:46318667-46366548 |
|
SCT |
ENSG00000070031 | −21.32 | 0.018 |
chr11:626430-627143 |
|
CEP250 |
ENSG00000126001 | −20.73 | <0.001 |
chr20:34042984-34099804 |
|
ZNF674 |
ENSG00000251192 | −20.15 | <0.001 |
chrX:46357161-46404892 |
|
GIGYF1 |
ENSG00000146830 | −19.36 | <0.001 |
chr7:100277129-100287071 |
|
ZNF814 |
ENSG00000204514 | −18.82 | <0.001 |
chr19:58281022-58427978 |
|
CEBPZ-AS1 |
ENSG00000218739 | −18.02 | <0.001 |
chr2:37394962-37551951 |
|
RABEP2 |
ENSG00000177548 | −17.18 | <0.001 |
chr16:28889725-28950667 |
|
SMARCA4 |
ENSG00000127616 | −17.17 | <0.001 |
chr19:11071597-11176071 |
|
CRIP1 |
ENSG00000257341 | −16.97 | 0.004 |
chr14:105952653-105965912 |
|
SULT1A3 |
ENSG00000261052 | −16.30 | 0.002 |
chr16:29262828-30215631 |
|
TMEM180 |
ENSG00000138111 | −15.97 | <0.001 |
chr10:104221148-104236802 |
|
CCDC88B |
ENSG00000168071 | −15.71 | <0.001 |
chr11:64107694-64125006 |
|
TCHP |
ENSG00000139437 | −15.38 | <0.001 |
chr12:110338068-110434194 |
|
CHCHD6 |
ENSG00000159685 | −15.37 | 0.003 |
chr3:126423062-126679249 |
|
BAIAP2 |
ENSG00000175866 | −15.31 | <0.001 |
chr17:79008947-79156964 |
Validation of sequencing data using
RT-qPCR
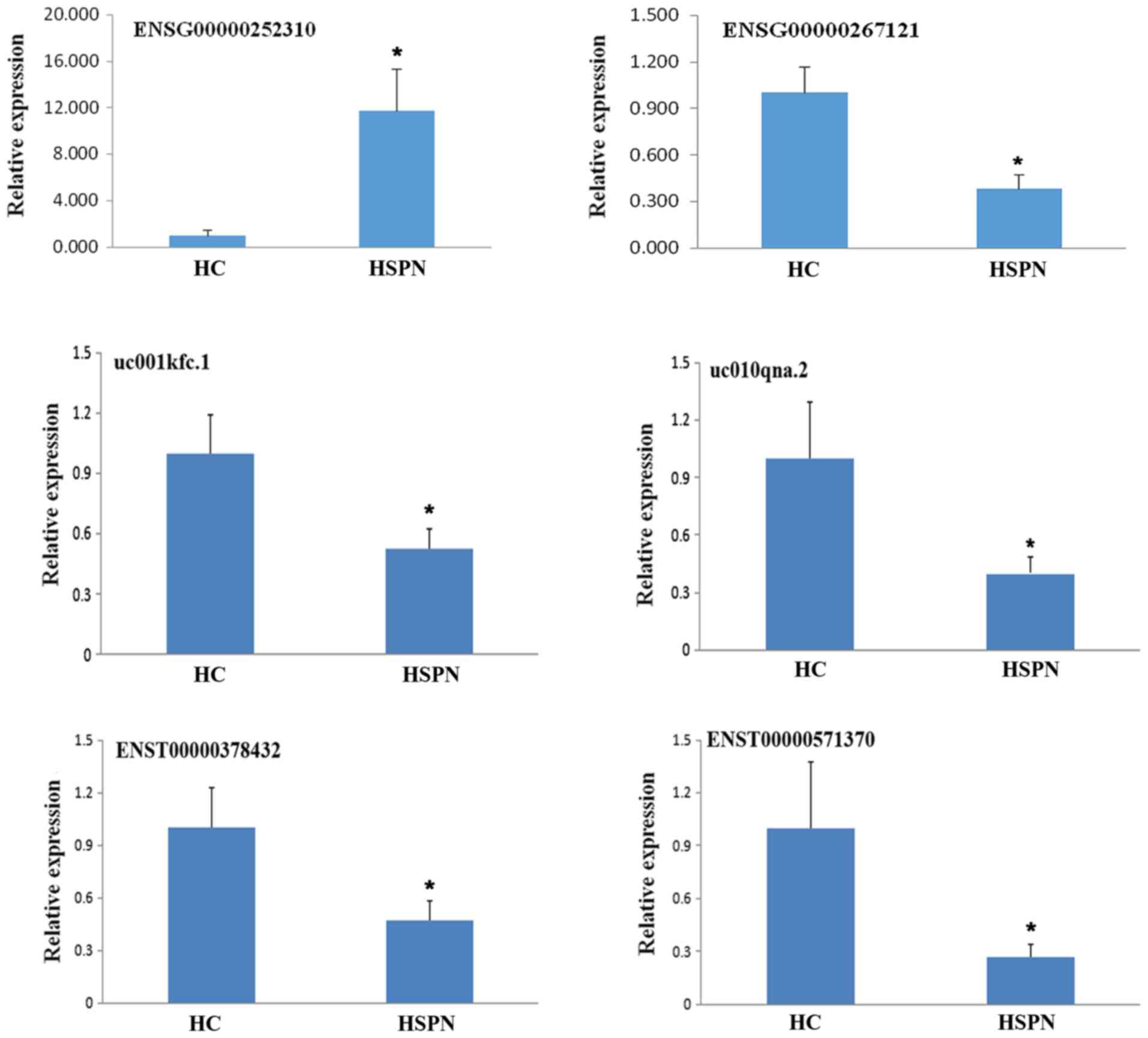
In order to validate the sequencing analysis, 6
differentially expressed lncRNAs were selected. RT-qPCR analysis
verified that the lncRNA ENSG00000252310 was upregulated, while
ENSG00000267121, uc001kfc.1, uc010qna.2, ENST00000378432 and
ENST00000571370 were downregulated in the 10 samples (Fig. 3). It was identified that the RT-qPCR
results were consistent with the high-throughput sequencing
data.
GO and KEGG pathway enrichment
analysis
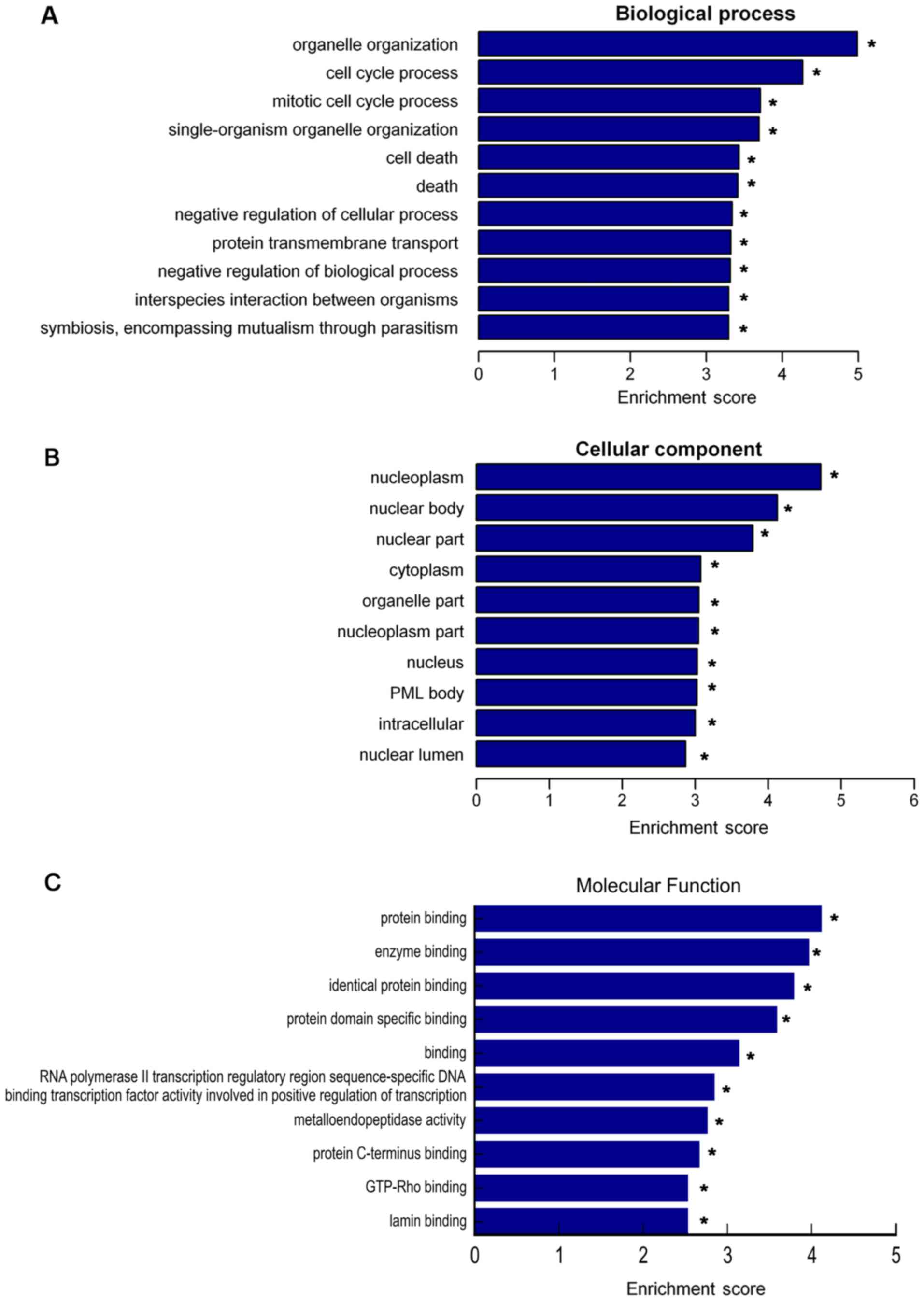
To investigate potential lncRNAs and the moieties
that they regulate, which are enriched in the GO terms biological
process, cellular component and molecular function, GO analysis was
performed for the differentially expressed lncRNAs (Fig. 4). The present study focused on GO
terms enriched among downregulated lncRNAs, and the top 10 terms in
the category biological process were as follows: i) Organelle
organization, ii) cell cycle process, iii) mitotic cell cycle
process, iv) single-organism organelle organization, v) cell death,
vi) death, vii) negative regulation of cellular process, viii)
protein transmembrane transport, ix) negative regulation of
biological process and x) interspecies interaction between
organisms (Fig. 4A). The top 10 GO
terms in the category cellular component were as follows: i)
Nucleoplasm, ii) nuclear body, iii) nuclear part, iv) cytoplasm, v)
organelle part, vi) nucleoplasm part, vii) nucleus, viii)
promyelocytic leukemia protein body, ix) intracellular and x)
nuclear lumen (Fig. 4B). The top 10
GO terms in the category molecular function were as follows: i)
Protein binding, ii) enzyme binding, iii) identical protein
binding, iv) protein domain specific binding, v) binding, vi) RNA
polymerase II transcription regulatory region sequence-specific DNA
binding transcription factor activity involved in positive
regulation of transcription, vii) metalloendopeptidase activity,
viii) protein C-terminus binding, ix) GTP-Rho binding and x) lamin
binding (Fig. 4C).
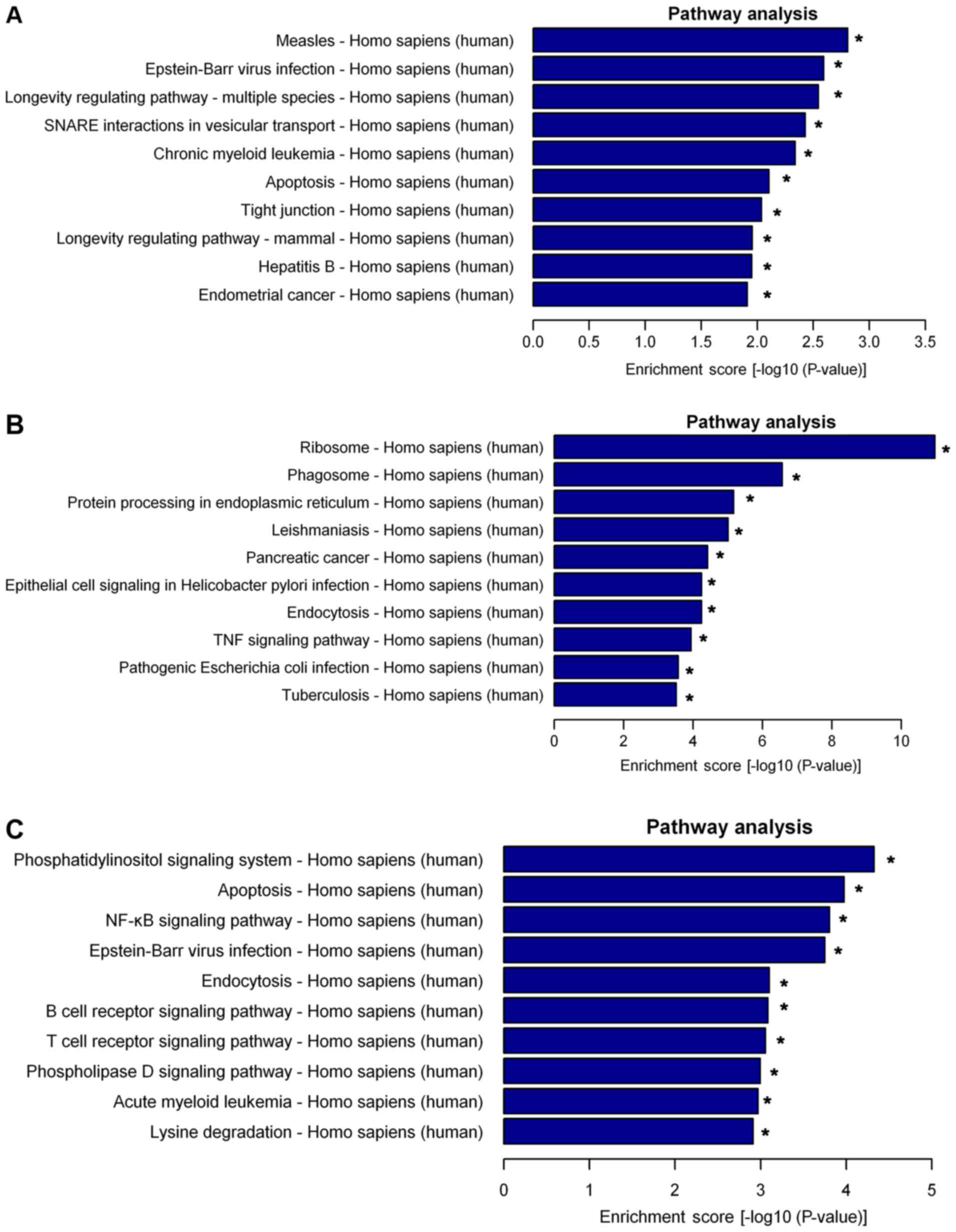
Pathway analysis indicated that 25 pathways were
significantly enriched among the differentially expressed lncRNAs.
The top 10 significantly enriched pathways were presented as
follows: i) Measles, ii) Epstein-Barr virus infection, iii)
longevity regulating pathway-multiple, iv) soluble N-ethylmaleimide
sensitive factor attachment protein receptor interactions in
vesicular transport, v) chronic myeloid leukemia, vi) apoptosis,
vii) tight junction, viii) longevity regulating pathway-mammal, ix)
hepatitis B and x) endometrial cancer (Fig. 5A). Furthermore, the differently
expressed mRNAs were also subjected to KEGG analysis, the top 10
enriched pathways for the upregulated mRNAs were presented as
follows: i) Ribosome, ii) phagosome, iii) protein processing in
endoplasmic reticulum, iv) leishmaniasis, v) pancreatic cancer, vi)
epithelial cell signaling in Helicobacter pylori infection,
vii) endocytosis, viii) tumor necrosis factor (TNF) signaling
pathway, ix) pathogenic Escherichia coli infection and x)
tuberculosis (Fig. 5B). The top 11
enriched pathways for the downregulated mRNAs were presented as
follows: i) Phosphatidylinositol signaling system, ii) apoptosis,
iii) nuclear factor (NF)-κB signaling pathway, iv) Epstein-Barr
virus infection, v) endocytosis, vi) B cell receptor signaling
pathway, vii) T cell receptor signaling pathway, viii) T cell
receptor signaling pathway, ix) phospholipase D signaling pathway,
x) acute myeloid leukemia and xi) lysine degradation (Fig. 5C).
Discussion
HSPN is a major public health problem that accounts
for 78.9% of secondary glomerulopathies in pediatric patients
(21). Etiologically, persistent
purpura or relapse, severe abdominal symptoms (abdominal pain,
gastrointestinal bleeding and severe bowel angina), arthritis and
being aged >10 years old are the most significant risk factors
for pediatric HSPN (22–24). Initially, lncRNAs were assumed to
simply be leaky transcription noise (25). However, an abundance of studies have
demonstrated that lncRNAs serve important roles in physiological
processes and the pathophysiology of numerous diseases,
participating in the regulation of DNA methylation, histone
modification, basal transcription, post-transcriptional processes,
directly binding proteins and protein function (26–30).
To the best of our knowledge, the present study was
the first to investigate the expression profile of lncRNAs and
mRNAs in patients with HSPN. Total RNA was extracted from the
peripheral blood of patients and HC, and differentially expressed
lncRNAs and mRNAs were identified using high-throughput sequencing,
followed by verification of certain RNAs by RT-qPCR analysis. In
the RT-qPCR experiments, ACTB was used as an endogenous control for
normalising the expression of lncRNAs and mRNAs according to
previous studies (31–34). Studies have demonstrated that ACTB is
suitable reference gene in gene expression studies of human
diseases when human peripheral blood as samples (35–37), and
may be more stably expressed in whole blood than in peripheral
blood mononuclear cells (38). In
the present study, it was identified that the RT-qPCR results were
consistent with the high-throughput sequencing data.
A total of 820 lncRNAs, including 34 upregulated and
786 downregulated ones, were identified to be differentially
expressed between the two groups. In addition, 3,557 differentially
expressed protein-coding mRNAs were identified from the same
samples, including 1,232 upregulated and 2,325 downregulated mRNAs.
The results demonstrated that the expression of lncRNAs and mRNAs
in patients with HSPN were quite different from that in HC.
Additionally, a greater number of lncRNAs and mRNAs were
significantly downregulated than upregulated in patients with HSPN.
GO and KEGG pathway analyses were used to investigate the possible
mechanistic roles and pathways of lncRNAs and mRNAs in HSPN.
An integrative method involving pathway analysis was
applied to identify possible functional associations between the
different RNA molecules. Based on the differentially expressed
mRNAs, a pathway analysis revealed via which biological functions
and mechanisms they may be involved in HSPN formation. HSPN is a
small-vessel form of autoimmune vasculitis caused by IgA1-mediated
inflammation (39,40). The results of the present study
suggested that a variety of KEGG pathways, including ribosomal
function, phagosome activity, protein processing in the endoplasmic
reticulum and endocytosis, are significantly enriched among the
upregulated mRNAs from patients with HSPN. The majority of these
pathways are involved in the generation of serum proteins,
including IgA. Among these associated pathways, epithelial cell
signaling in Helicobacter pylori infection, tumor necrosis
factor TNF signaling, pathogenic Escherichia coli infection,
the phosphatidylinositol signaling system, NF-κB signaling,
Epstein-Barr virus infection, the B cell receptor signaling pathway
and the T cell receptor signaling pathway exhibited significant
changes in upregulated and downregulated mRNAs. In line with this,
Shin et al (41) reported
that Helicobacter pylori infection may cause the serum
levels of IgA, C3 and cryoglobulins to increase, which promotes
immune complex formation and increases the risk of HSP occurrence.
Hirayama et al (42)
demonstrated that the percentage values of the CD3-gated β-chain of
the T cell receptor in patients with HSPN were significantly
increased compared with those in healthy individuals. Chen et
al (43) revealed that TNF-like
weak inducer of apoptosis, a member of the TNF family, was elevated
in patients with acute-stage HSP and may act as a regulator of
NF-κB activation and chemokine production in human dermal
microvascular endothelial cells, promoting leucocyte migration in
cutaneous vasculitis. These results indicated that aberrant IgA
circulating immune complexes, as well as elevated pro-inflammatory
cytokines and chemokines, are all associated with the pathogenesis
of HSPN.
Previous studies have demonstrated that apoptosis is
one of the most important factors for controlling inflammation in
inflammatory diseases such as HSP (44,45).
Among the top 10 enriched pathways of the aberrantly expressed
lncRNAs in the HSPN group, the present study focused on the
apoptotic pathway, which serves an important role in the
pathogenesis of HSPN. Yuan et al (46) suggested that IgA1 from patients with
HSP may induce apoptosis of human umbilical vein endothelial cells,
which may be associated with vascular endothelial injury in HSP.
Ozaltin et al (45) observed
a marked expression of Fas on peripheral blood neutrophils and
lymphocytes in patients with HSP in the acute and the resolution
phases. This suggested that increased removal of inflammatory cells
through apoptosis may contribute to the early control of the
inflammatory response and repair in this self-limited vasculitis.
The present study revealed that ENST00000378432, ENST00000571370,
uc001kfc.1 and uc010qna.2 expression was decreased in patients with
HSPN and healthy controls. These lncRNAs were associated with the
p53 signaling pathway and apoptosis-associated genes (AKT
serine/threonine kinase 2, tumor protein 53, phosphatase and tensin
homolog and FAS). Further studies on these differentially expressed
lncRNAs will be performed to establish their functions in
apoptosis.
The major limitation of the present study was the
sample size of the patients and controls. The present results
should be validated in larger cohorts. Interaction networks
analyses are also required to further investigate the associations
between ncRNAs, coding RNAs and proteins. Furthermore, lncRNAs and
mRNAs validated to be associated with HSPN by RT-qPCR should be
further investigated at the cellular level.
In conclusion, the results of the present study
demonstrated a significant difference in the expression of certain
lncRNAs and mRNAs between patients with HSPN and HCs. The results
indicated that lncRNAs are important regulators in HSPN
pathophysiological mechanisms. Functional research on these lncRNAs
may be a novel and interesting research field.
Acknowledgements
The authors of the present study would like to thank
Shanghai Cloud-Seq Biotech Laboratory, for their help in the
guidance of the experiments and analysis of the data of this
manuscript.
Funding
The present study was financially supported by the
2015 National Scientific Research Specific of Traditional Chinese
Medicine Industry (grant no. 201507001-03). The State
Administration of Traditional Chinese Medicine of the People's
Republic of China contributed to the conception of the study and
helped perform the collection of data.
Availability of data and materials
The datasets generated and analyzed during the
present study are available from the Gene Expression Omnibus
(GSE102114; www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE102114).
Authors' contributions
SP, JL, SW and JZ were responsible for the design,
supervision of the study and revision of the manuscript. JL and SW
acquired the data and helped perform the analysis with constructive
discussions. SP analyzed and interpreted the patient data regarding
the HSPN and was a major contributor in writing the manuscript. GY
and XD participated in the designing the study, drafting the
manuscript and critically revising it for intellectual content. JZ
agreed to being accountable for all aspects of the work to ensure
that questions associated with the accuracy or integrity of any
part of the study are appropriately investigated and resolved. All
authors read, discussed, revised and approved the final
manuscript.
Ethics statement and consent to
participate
The present study was approved by Ethics Committee
of the Institutional Ethics Board of the Affiliated Hospital of
Liaoning University of Traditional Chinese Medicine (approval no.
2016CS(KT)-002-01). Written informed consent was obtained by the
parents of all of the subjects enrolled in the study.
Patient consent for publication
Written informed consent was obtained by the parents
of all of the pediatric patients enrolled in the study for
publication of any associated data.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
HSPN
|
Henoch-Schönlein purpura nephritis
|
|
lncRNA
|
long non-coding RNA
|
|
GO
|
Gene Ontology
|
|
KEGG
|
Kyoto Encyclopedia of Genes and
Genomes
|
|
RT-qPCR
|
reverse transcription-quantitative
polymerase chain reaction
|
|
FPKM
|
fragment per kilobase of exon per
million fragments mapped
|
|
HC
|
healthy controls
|
|
ACTB
|
β-actin
|
References
|
1
|
Chen JY and Mao JH: Henoch-Schönlein
purpura nephritis in children: Incidence, pathogenesis and
management. World J Pediatr. 11:29–34. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kawasaki Y, Ono A, Ohara S, Suzuki Y,
Suyama K, Suzuki J and Hosoya M: Henoch-Schönlein purpura nephritis
in childhood: Pathogenesis, prognostic factors and treatment.
Fukushima J Med Sci. 59:15–26. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Gardner-Medwin JM, Dolezalova P, Cummins C
and Southwood TR: Incidence of Henoch-Schönlein purpura, Kawasaki
disease, and rare vasculitides in children of different ethnic
origins. Lancet. 360:1197–1202. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Yang YH, Hung CF, Hsu CR, Wang LC, Chuang
YH, Lin YT and Chiang BL: A nationwide survey on epidemiological
characteristics of childhood Henoch-Schönlein purpura in Taiwan.
Rheumatology (Oxford). 44:618–622. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Saulsbury FT: Clinical update:
Henoch-Schönlein purpura. Lancet. 369:976–978. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Sano H, Izumida M, Shimizu H and Ogawa Y:
Risk factors of renal involvement and significant proteinuria in
Henoch-Schönlein purpura. Eur J Pediatr. 161:196–201. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kung JT, Colognori D and Lee JT: Long
noncoding RNAs: Past, present, and future. Genetics. 193:651–669.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kapusta A and Feschotte C: Volatile
evolution of long noncoding RNA repertoires: Mechanisms and
biological implications. Trends Genet. 30:439–452. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yu TM, Palanisamy K, Sun KT, Day YJ, Shu
KH, Wang IK, Shyu WC, Chen P, Chen YL and Li CY: RANTES mediates
kidney ischemia reperfusion injury through a possible role of
HIF-1α and LncRNA PRINS. Sci Rep. 6:184242016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lin J, Zhang X, Xue C, Zhang H, Shashaty
MG, Gosai SJ, Meyer N, Grazioli A, Hinkle C, Caughey J, et al: The
long noncoding RNA landscape in hypoxic and inflammatory renal
epithelial injury. Am J Physiol Renal Physiol. 309:F901–F913. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Alvarez ML and DiStefano JK: Functional
characterization of the plasmacytoma variant translocation 1 gene
(PVT1) in diabetic nephropathy. PLoS One. 6:e186712011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Duan LJ, Ding M, Hou LJ, Cui YT, Li CJ and
Yu DM: Long noncoding RNA TUG1 alleviates extracellular matrix
accumulation via mediating microRNA-377 targeting of PPARγ in
diabetic nephropathy. Biochem Biophys Res Commun. 484:598–604.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Huang YS, Hsieh HY, Shih HM, Sytwu HK and
Wu CC: Urinary Xist is a potential biomarker for membranous
nephropathy. Biochem Biophys Res Commun. 452:415–421. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Arvaniti E, Moulos P, Vakrakou A,
Chatziantoniou C, Chadjichristos C, Kavvadas P, Charonis A and
Politis PK: Whole-transcriptome analysis of UUO mouse model of
renal fibrosis reveals new molecular players in kidney diseases.
Sci Rep. 6:262352016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wu Y, Zhang F, Ma J, Zhang X, Wu L, Qu B,
Xia S, Chen S, Tang Y and Shen N: Association of large intergenic
noncoding RNA expression with disease activity and organ damage in
systemic lupus erythematosus. Arthritis Res Ther. 17:1312015.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Bao X, Duan J, Yan Y, Ma X, Zhang Y, Wang
H, Ni D, Wu S, Peng C, Fan Y, et al: Upregulation of long noncoding
RNA PVT1 predicts unfavorable prognosis in patients with clear cell
renal cell carcinoma. Cancer Biomark. 21:55–63. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Leti F and DiStefano JK: Long noncoding
RNAs as diagnostic and therapeutic targets in type 2 diabetes and
related complications. Genes (Basel). 8(pii): E2072017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Subspecialty Group of Renal Disease,
Society of Pediatrics, Chinese Medical Association: Clinical
classification, diagnosis and treatment of glomerular diseases in
children. Chin J Pediatr. 39:746–749. 2001.
|
|
19
|
Martin M: Cutadapt removes adapter
sequences from high-throughput sequencing reads. EMBnet J. 17:2011.
View Article : Google Scholar
|
|
20
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Liu GL, Gao YF, Xia ZK, Fan ZM, Ren XG,
Mao S, He X and Sun T: The pathological analysis of 2551 children
patients with glomerulopathy. J Med Postgrad. 24:294–297. 2011.(In
Chinese).
|
|
22
|
Hennies I, Gimpel C, Gellermann J, Möller
K, Mayer B, Dittrich K, Büscher AK, Hansen M8, Aulbert W, Wühl E,
et al: Presentation of pediatric Henoch-Schönlein purpura nephritis
changes with age and renal histology depends on biopsy timing.
Pediatr Nephrol. 33:277–286. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chan H, Tang YL, Lv XH, Zhang GF, Wang M,
Yang HP and Li Q: Risk factors associated with renal involvement in
childhood Henoch-Schönlein purpura: A meta-analysis. PLoS One.
11:e01673462016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Bogdanović R: Henoch-Schönlein purpura
nephritis in children: Risk factors, prevention and treatment. Acta
Paediatr. 98:1882–1889. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Gibb EA, Vucic EA, Enfield KS, Stewart GL,
Lonergan KM, Kennett JY, Becker-Santos DD, MacAulay CE, Lam S,
Brown CJ and Lam WL: Human cancer long non-coding RNA
transcriptomes. PLoS One. 6:e259152011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Arab K, Park YJ, Lindroth AM, Schäfer A,
Oakes C, Weichenhan D, Lukanova A, Lundin E, Risch A, Meister M, et
al: Long noncoding RNA TARID directs demethylation and activation
of the tumor suppressor TCF21 via GADD45A. Mol Cell. 55:604–614.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wang Z, Zhang XJ, Ji YX, Zhang P, Deng KQ,
Gong J, Ren S, Wang X, Chen I, Wang H, et al: The long noncoding
RNA Chaer defines an epigenetic checkpoint in cardiac hypertrophy.
Nat Med. 22:1131–1139. 2016. View
Article : Google Scholar : PubMed/NCBI
|
|
28
|
Li SY and Susztak K: The long noncoding
RNA Tug1 connects metabolic changes with kidney disease in
podocytes. J Clin Invest. 126:4072–4075. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Wu G, Cai J, Han Y, Chen J, Huang ZP, Chen
C, Cai Y, Huang H, Yang Y, Liu Y, et al: LincRNA-p21 regulates
neointima formation, vascular smooth muscle cell proliferation,
apoptosis, and atherosclerosis by enhancing p53 activity.
Circulation. 130:1452–1465. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Hu G, Lou Z and Gupta M: The long
non-coding RNA GAS5 cooperates with the eukaryotic translation
initiation factor 4E to regulate c-Myc translation. PLoS One.
9:e1070162014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Bi HS, Yang XY, Yuan JH, Yang F, Xu D, Guo
YJ, Zhang L, Zhou CC, Wang F and Sun SH: H19 inhibits RNA
polymerase II-mediated transcription by disrupting the hnRNP
U-actin complex. Biochim Biophys Acta. 1830:4899–4906. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Song H, Sun W, Ye G, Ding X, Liu Z, Zhang
S, Xia T, Xiao B, Xi Y and Guo J: Long non-coding RNA expression
profile in human gastric cancer and its clinical significances. J
Transl Med. 11:2252013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Yu A, Wang Y, Yin J, Zhang J, Cao S, Cao J
and Shen Y: Microarray analysis of long non-coding RNA expression
profiles in monocytic myeloid-derived suppressor cells in
Echinococcus granulosus-infected mice. Parasit Vectors. 11:3272018.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Xiang Y, Zhang Y, Tang Y and Li Q: MALAT1
modulates TGF-β1-induced endothelial-to-mesenchymal transition
through downregulation of miR-145. Cell Physilo Biochem.
42:357–372. 2017. View Article : Google Scholar
|
|
35
|
Zsóri KS, Muszbek L, Csiki Z and Shemirani
AH: Validation of reference genes for the determination of platelet
transcript level in healthy individuals and in patients with the
history of myocardial infarction. Int J Mol Sci. 14:3456–3466.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Kozmus CE and Potočnik U: Reference genes
for real-time qPCR in leukocytes from asthmatic patients before and
after anti-asthma treatment. Gene. 570:71–77. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zhang X, Ding L and Sandford AJ: Selection
of regerence genes for gene expression studies in human neutrophils
by real-time PCR. BMC Mol Biol. 6:42005. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Wang J, Wang Y, Wang H, Hao X, Wu Y and
Guo J: Selection of reference genes for gene expression studies in
porcine whole blood and peripheral blood mononuclear cells under
polyinosinic: Polycytidylic acid stimulation. Asian-Australas J
Anim Sci. 27:471–478. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Park SJ, Suh JS, Lee JH, Lee JW, Kim SH,
Han KH and Shin JI: Advances in our understanding of the
pathogenesis of Henoch-Schönlein purpura and the implications for
improving its diagnosis. Expert Rev Clin Immunol. 9:1223–1238.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Davin JC: Henoch-Schonlein purpura
nephritis: Pathophysiology, treatment, and future strategy. Clin J
Am Soc Nephrol. 6:679–689. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Shin JI, Koh H and Lee JS:
Henoch-Schönlein purpura associated with helicobacter pylori
infection: The pathogenic roles of IgA, C3, and cryoglobulins?
Pediatr Dermatol. 26:768–769. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Hirayama K, Kobayashi M, Muro K, Yoh K,
Yamagata K and Koyama A: Specific T-cell receptor usage with
cytokinemia in Henoch-Schönlein purpura nephritis associated with
Staphylococcus aureus infection. J Intern Med. 249:289–295. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Chen T, Guo ZP, Li MM, Li JY, Jiao XY,
Zhang YH and Liu HJ: Tumour necrosis factor-like weak inducer of
apoptosis (TWEAK), an important mediator of endothelial
inflammation, is associated with the pathogenesis of
Henoch-Schonlein purpura. Clin Exp Immunol. 166:64–71. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Haslett C: Granulocyte apoptosis and
inflammatory disease. Br Med Bull. 53:669–683. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Ozaltin F, Besbas N, Uckan D, Tuncer M,
Topaloglu R, Ozen S, Saatci U and Bakkaloglu A: The role of
apoptosis in childhood Henoch-Schonlein purpura. Clin Rheumatol.
22:265–267. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Yuan P, Bo Y, Ming G, Wen-Jun F, Qin Z and
Bo H: Apoptosis of human umbilical vein endothelial cells (HUVEC)
induced by IgA1 isolated from Henoch-Schonlein purpura children.
Asian Pac J Allergy Immunol. 32:34–38. 2014.PubMed/NCBI
|