Introduction
Spinal cord injury (SCI) has become a common disease
with the development of modern transportation and the mining
industry and affects ~2.5 million individuals worldwide (1–3). Primary
SCI combined with secondary SCI seriously threatens the health and
quality of life of patients (3).
Primary SCI consists of irreversible tissue damages immediately
following external forces (3).
Secondary SCI refers to the cracking, progressive and
self-destructive process associated with various factors, including
reactive oxygen species and inflammation, based on primary injury
(4). The SCI is of fundamental
importance to elucidate the immunological mechanisms of secondary
SCI to prevent and reverse secondary SCI (5).
A previous study on the mechanisms of post-SCI
pathological injury have largely focused on primary and secondary
injuries. Primary trauma of the spinal cord leads to irreversible
primary injury (6). In contrast,
cascade reactions of primary injury lead to reversible secondary
injury, with injury levels that are even more serious than those of
primary injury (7). It has
previously been demonstrated that extensive nerve cell apoptosis
and necrosis occur following secondary injury (8). The apoptotic nerve cells include
astrocytes, microglia cells and oligodendrocytes. Furthermore,
nerve cell apoptosis deconstructs the microenvironment of axon
regeneration (9). A previous study
has suggested that the inflammatory response triggered by secondary
injury following central nerve injury is a key cause of further
injury (9). Brain-derived
neurotrophic factor (BDNF) is produced through proteolysis of
precursor molecules (10). It is a
neurotrophic factor belonging to the nerve growth factor family
(10). BDNF and its receptor are
extensively expressed in the nervous system (11). They are the most prevalent neurokines
in vivo (12). Changes in
endogenous BDNF following central lesion have been intensively
studied since the discovery of BDNF (12). BDNF is a neurotrophic factor that
greatly affects neuron growth and development. In addition, it can
improve the physiological status of neurons under pathological
status (12). BDNF has important
biological roles in inflammation and apoptosis (13); therefore, it is an indispensable
neurotrophic factor for maintaining normal nervous system function
(14). Furthermore, BDNF is a member
of the neurotrophic factor family that is mainly secreted by neuron
or glial cells. It can bind with its specific receptor, tyrosine
kinase receptor B (TrkB) on adjacent membranes in the paracrine or
autocrine manner to exert its function (11). Furthermore, it serves a vital role in
neuron survival and proliferation as well as synaptic plasticity
(14). Furthermore, it can promote
axonal regeneration and repair following central lesion, and
accelerate limb functional recovery (14).
Receptor tropomyosin-receptor-kinase B (TrkB) plays
a key role in the antidepressant functions of antidepressants
(15). Blocking of BDNF-TrkB
signaling inhibits the effects of neurological function recovery
after treadmill training in spinal cord injury rat (15). BDNF-TrkB is suppressed by
interleukin-1β via p38 mitogen-activated protein kinase (MAPK)
(16). The aim of the present study
was to investigate the pro-inflammation effects of BDNF signaling
in promoting inflammation following spinal cord injury (SCI) in
rats.
Materials and methods
Animals and surgical procedures
All animal studies were approved by the Committee of
Ethics on Animal Experiments of The First People's Hospital of
China Three Gorges University (Yichang, China). A total of 20 Adult
male Sprague Dawley (SD) rats (weight, 250–300 g; age, 8–10 weeks)
were purchased from the Animal Experiment Centre of Whuan
University (Wuhan, China) and randomly assigned into the following
two experimental groups: Sham-surgery group (n=10) and SCI model
group (n=10). All SD rats were bred in China Three Gorges
University at a constant temperature (23±3°C), 50±5% humidity and a
12-h light/dark cycle, with free access to food and water. The
sham-surgery group received sham surgery of laminectomies only.
Rats were anesthetized using 35 mg/kg pentobarbital sodium
(Sigma-Aldrich; Merck KGaA, Darmstadt, Germany). The T12 spinal
laminectomy of the SCI model group exposed the spinal cord and an
impactor with a diameter of 2.5 mm was used to perform a
moderate-intensity weight-drop following anesthesia.
Histological preparation
All rats (n=10) were sacrificed via decollation
under 35 mg/kg pentobarbital sodium anesthesia and spinal cord
tissue samples were collected and washed with PBS. Tissue was fixed
for 24–48 h at room temperature using 4% paraformaldehyde and
dehydrated using gradient ethanol. Fixed tissue was cut into 8
µm-thick sagittal sections, which were stained using hematoxylin
and eosin staining at room temperature for 10 min and examined
under a confocal Eclipse 5Oi microscope (magnification, ×100; Nikon
Corporation, Tokyo, Japan).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
A Trizol reagent was used to extract RNA from
AGE1.HN cells following SCI induction and spinal cord tissue
samples (Invitrogen; Thermo Fisher Scientific, Inc., Waltham, MA,
USA). RNA was reverse transcribed into cDNA using the First Strand
cDNA Synthesis kit (GeneCopoeia, Inc., Rockville, MD, USA). qPCR
was performed using the ABI 7300HT real-time PCR system (Applied
Biosystems; Thermo Fisher Scientific, Inc.) and the SYBR green
supermix (Bio-Rad Laboratories, Inc., Hercules, CA, USA) The
thermocycling conditions were as follows: 95°C for 10 min, followed
by 40 cycles at 95°C for 30 sec, 58°C for 45 sec and 72°C for 30
sec. Relative gene expression was assessed using the
2−∆∆Cq method (17).
Evaluation of Basso, Beattie and
Bresnahan (BBB) test
All rats (n=3/per group) were subjected to the BBB
test for the evaluation of locomotor recovery after thee induction
of SCI, using a locomotor rating scale of 0 (no observable
hind-limb movements) to 21 (normal locomotion). This was performed
in accordance with a previous study (18).
Evaluation of the water content of
spinal cord
The wet weight of spinal cord tissue samples was
recorded, then spinal cord tissue samples were dried at 70–80°C for
48 h and the dry weight was recorded. Spinal cord water content (%)
was calculated as follows: (Wet weight-dry weight)/wet weight
×100.
ELISA assays
All rats (n=3) were sacrificed, and sensorimotor
cortical tissues were removed and incubated with
radioimmunprecipitation assay buffer (Beyotime Institute of
Biotechnology, Haimen, China) on ice for 30 min. Then, miscible
liquids were centrifuged at 13,200 × g at 4°C for 10 min. The
protein concentration of soluble materials was determined using a
bicinchoninic acid protein assay kit in accordance with the
manufacturer's protocol (Beyotime Institute of Biotechnology). A
total of 10 µg protein was used to measure interleukin (IL)-1β
(cat. no. H002), IL-6 (cat. no. H007), IL-18 (cat. no. H015), tumor
necrosis factor (TNF)-α (cat. no. H052), inducible nitric oxide
synthase (iNOS; cat. no. A014-1-1) and cyclooxygenase (COX)-2 (cat.
no. H200) levels using ELISA kits (Nanjing Jiancheng Bioengineering
Institute, Nanjing, China).
In vitro model and transfection
The nerve cell line AGE1.HN was purchased from the
Type Culture Collection of the Chinese Academy of Sciences
(Shanghai, China) and cultured in Dulbecco's modified Eagle's
medium medium (Gibco; Thermo Fisher Scientific, Inc.) supplemented
with 10% fetal bovine serum (Gibco; Thermo Fisher Scientific,
Inc.), 100 U/ml penicillin and 100 µg/ml streptomycin (HyClone; GE
Healthcare Life Sciences, Logan, UT, USA), and maintained at 37°C
in a humidified atmosphere with 5% CO2. BDNF-pcDNA3.1
(forward, 5′-AGAAAAGCCAAFFAGTGAA-3′ and reverse,
5′-AAAAGGGGAAGATAGTGGATTTATGTT-3′) and negative-pcDNA3.1 negative
mimic plasmids (forward, 5′-CCCCCCCCCCCCCCCCCC-3′ and reverse,
5′-CCCCCCCCCCCCCCCCCC-3′) were constructed by Sangon Biotech Co.,
Ltd., (Shanghai, China). Cells were transfected with 100 ng BDNF
plasmid or negative mimics using Lipofectamine 2000 (Invitrogen;
Thermo Fisher Scientific, Inc.), according to the manufacturer's
protocol. Following 48 h transfection, cells were induced with 100
ng/ml lipopolysaccharide (LPS; Beyotime Institute of Biotechnology)
for 4 h at 37°C. Cells were treated with 10 µM ANA-12
(MedChemExpress, Monmouth Junction, NJ, USA), a TrkB inhibitor, or
5 nM TA-02 (MedChemExpress, Monmouth Junction, NJ, USA), a p-38
inhibitor for 44 h at 37°C, and cells were inducted with 100 ng/ml
LPS for 4 h at 37°C, respectively and then were induced by 100
ng/ml LPS for 4 h at 37°C. Negative group, cell was transfected
with negative mimics.
Western blotting
Total protein was extracted from sensorimotor
cortical tissues and protein concentration was determined as
detailed above. Equal amounts of protein (50 µg) were separated by
12% SDS-PAGE followed by transferring onto polyvinylidene
difluoride membranes. Polyvinylidene difluoride membranes were
blocked with 5% skimmed milk in 0.01% TBS-Tween 20 for 1 h at 37°C
and then incubated with anti-BDNF (sc-20981; 1:200; Santa Cruz
Biotechnology, Inc., Dallas, TX, USA), anti-phosphorylated (p)-TrkB
(4619; 1:500; Cell Signaling Technology, Inc., Danvers, MA, USA),
anti-p-p38 (sc-17852-R; 1:4,000; Santa Cruz Biotechnology, Inc.),
and anti-GAPDH (sc-51631; 1:5,000; Santa Cruz Biotechnology, Inc.)
overnight at 4°C. Membranes were washed with TBST for 15 min and
incubated with horseradish peroxidase conjugated goat anti-rabbit
immunoglobulin G (cat. no. sc-2004; Santa Cruz Biotechnology, Inc.)
for 1 h at 37°C. Membranes were analyzed using an
electrochemiluminescence (ECL) reagent and measured using Image Lab
3.0 (Bio-Rad Laboratories, Inc.).
Immunocytochemistry
Cells after the induction of SCI were washed with
PBS and fixed with 4% paraformaldehyde in PBS (pH 7.4) for 15 min
at room temperature. Cells were then blocked with 5% bovine serum
albumin (Beyotime Institute of Biotechnolgy) in PBS containing 0.5%
Triton X-100 for 1 h at room temperature, then incubated with
anti-TrkB antibody (cat. no. sc-12; 1:100; Santa Cruz
Biotechnology, Inc.), anti-BDNF antibody (cat. no. sc-546; 1:100;
Santa Cruz Biotechnology, Inc.) at 4°C overnight. Following washing
for 30 min using PBS-Tween 20 (PBST), cells were incubated with
goat anti-rabbit IgG-CFL 555 (cat. no. sc-362272; 1:1,000; Santa
Cruz Biotechnology, Inc.) for 1.5 h at room temperature. Cells were
then stained using a DAPI assay for 15 min at room temperature and
washed with PBST for 30 min at room temperature. Cells were
visualized under fluorescence microscopy (magnification, ×20; Nikon
Corporation).
Statistical analysis
Data are presented as the mean + standard error of
the mean. Data were analyzed using Student's t-test or one-way
analysis of variance followed by Tukey's test. P<0.05 was
considered to indicate a statistically significant difference.
Results
Expression of BDNF in SCI rats
To explore the function of BDNF on inflammation in
SCI rats, the expression of BDNF in SCI rats was analyzed. HE
staining demonstrated that spinal cord cells seemed to be dead in
SCI rats, compared with sham control group (Fig. 1A). BBB score was inhibited and water
content of the spinal cord was increased in SCI rats, compared with
the sham control group (Fig. 1B and
C). The levels of IL-1β, IL-6, IL-18 and TNF-α in SCI rats were
also increased in SCI rats compared with those in sham-surgery rats
(Fig. 1D-G). Additionally, iNOS and
COX-2 levels were also increased in SCI rats, compared with sham
control group (Fig. 1H and I). The
expression of BDNF in SCI rats was further analyzed, which revealed
that the expression of BDNF was reduced in SCI rats, compared with
the sham control group (Fig. 1J and
K). Therefore, these results demonstrated that BDNF may be
associated with inflammation of SCI.
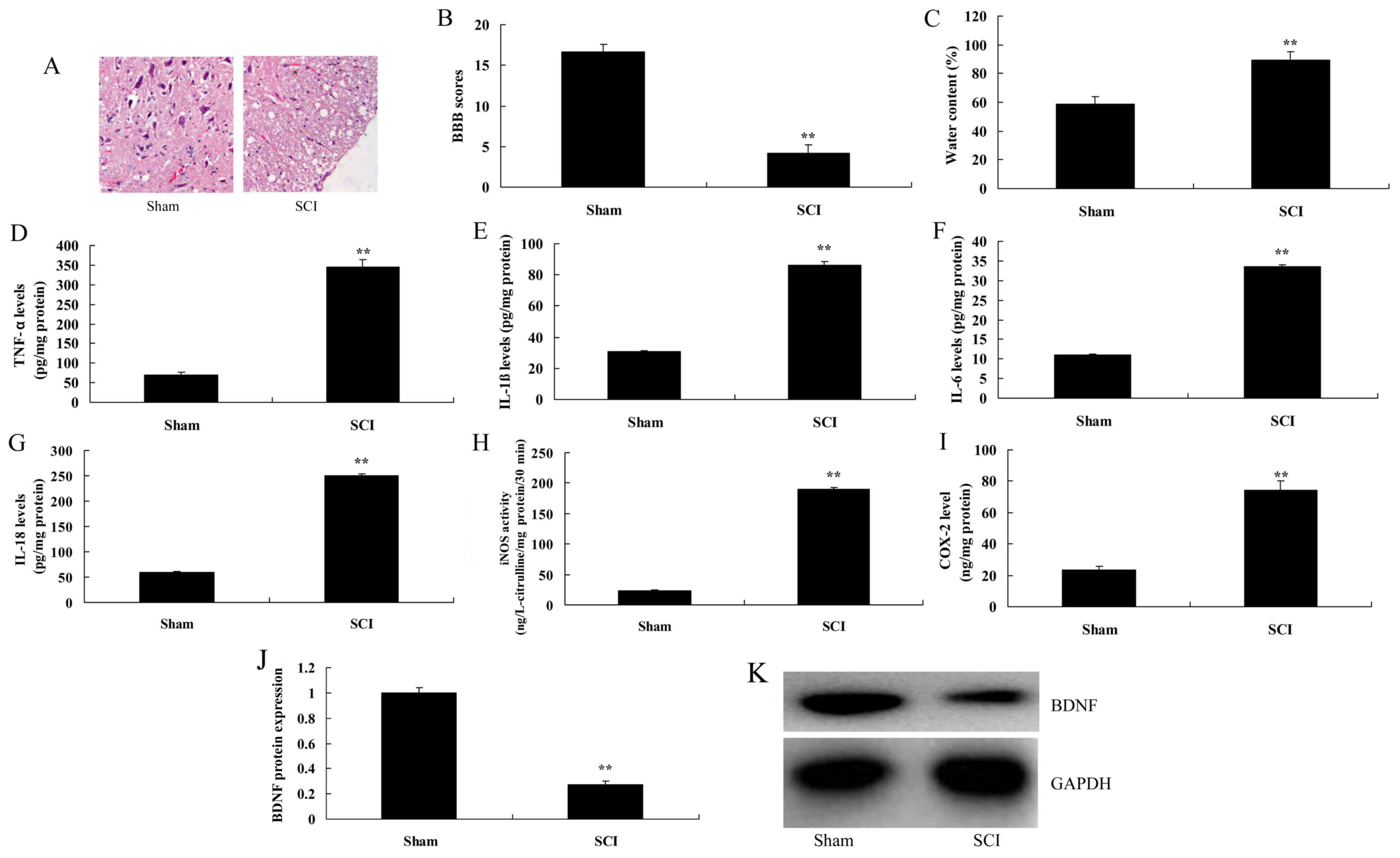 | Figure 1.Expression of BDNF in SCI rats. (A)
Histological evaluation (original magnification, ×100). (B) BBB
scores, (C) water content of spinal cord. (D) TNF-α, (E) IL-1β, (F)
IL-6, (G) IL-18, (H) iNOS and (I) COX2 levels. (J) BDNF protein
expression, as determined using statistical analysis. (K) Western
blot analysis of BDNF protein expression. Data are presented as the
mean ± standard error of the mean for three independent
experiments. **P<0.01 vs. sham. BBB, Basso, Beattie and
Bresnahan; SCI, spinal cord injury; TNF, tumor necrosis factor; IL,
interleukin; iNOS, inducible nitric oxide synthase; COX,
cyclooxygenase; BDNF, brain-derived neurotrophic factor. |
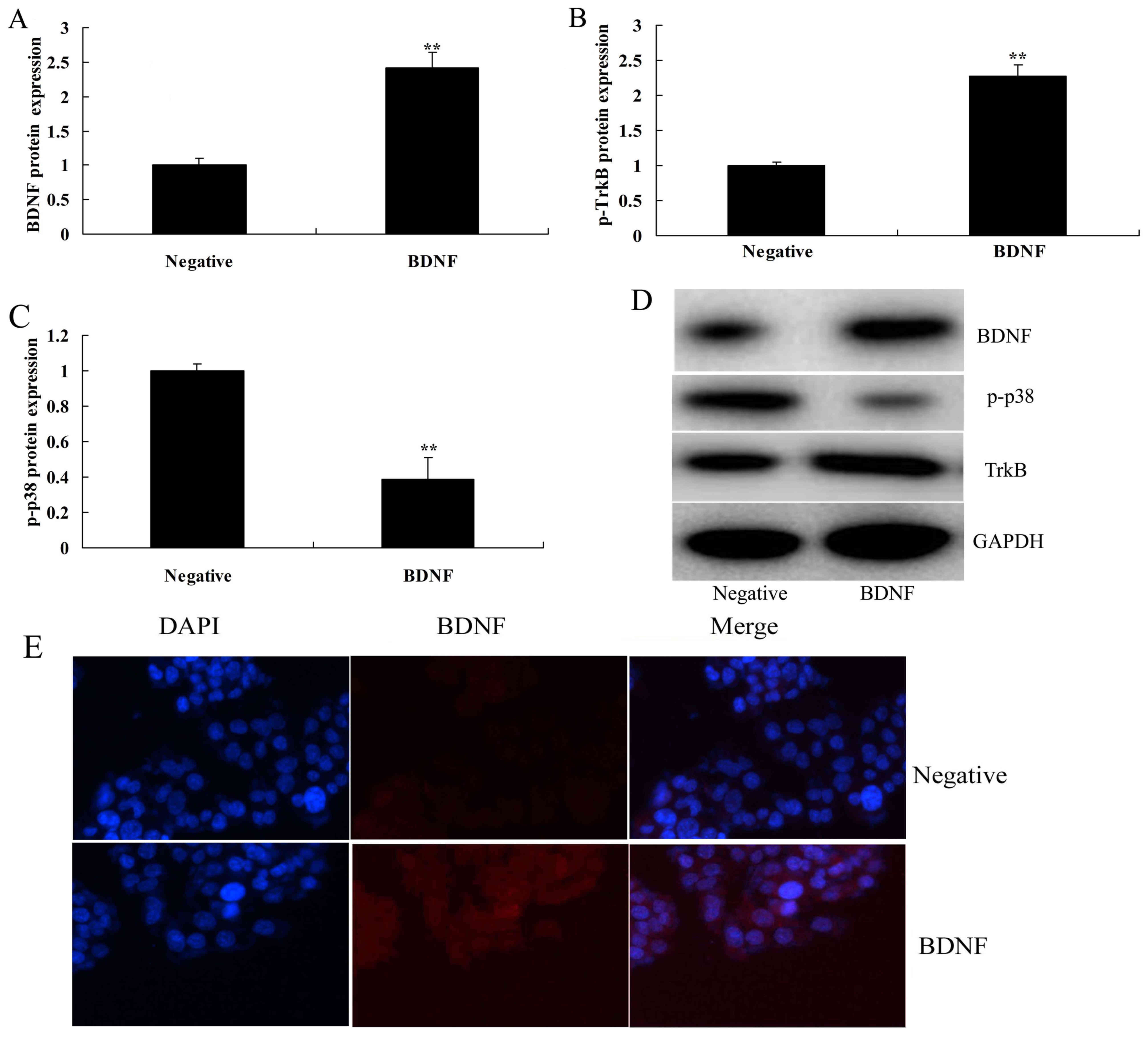
Overexpression of BDNF induces TrkB
and p-p-38 protein expressions in vitro
Next, the exact roles of BDNF in inflammation in
vitro were evaluated. As presented in Fig. 2A-D, BDNF expression plasmids
significantly increased the protein expression of BDNF and TrkB,
and suppressed that of p-p38 in an in vitro model, compared
with the negative group. Consistently, immunofluorescence indicated
that the BDNF expression plasmid markedly induced BDNF protein
expression in an in vitro model, in comparison with negative
group (Fig. 2E).
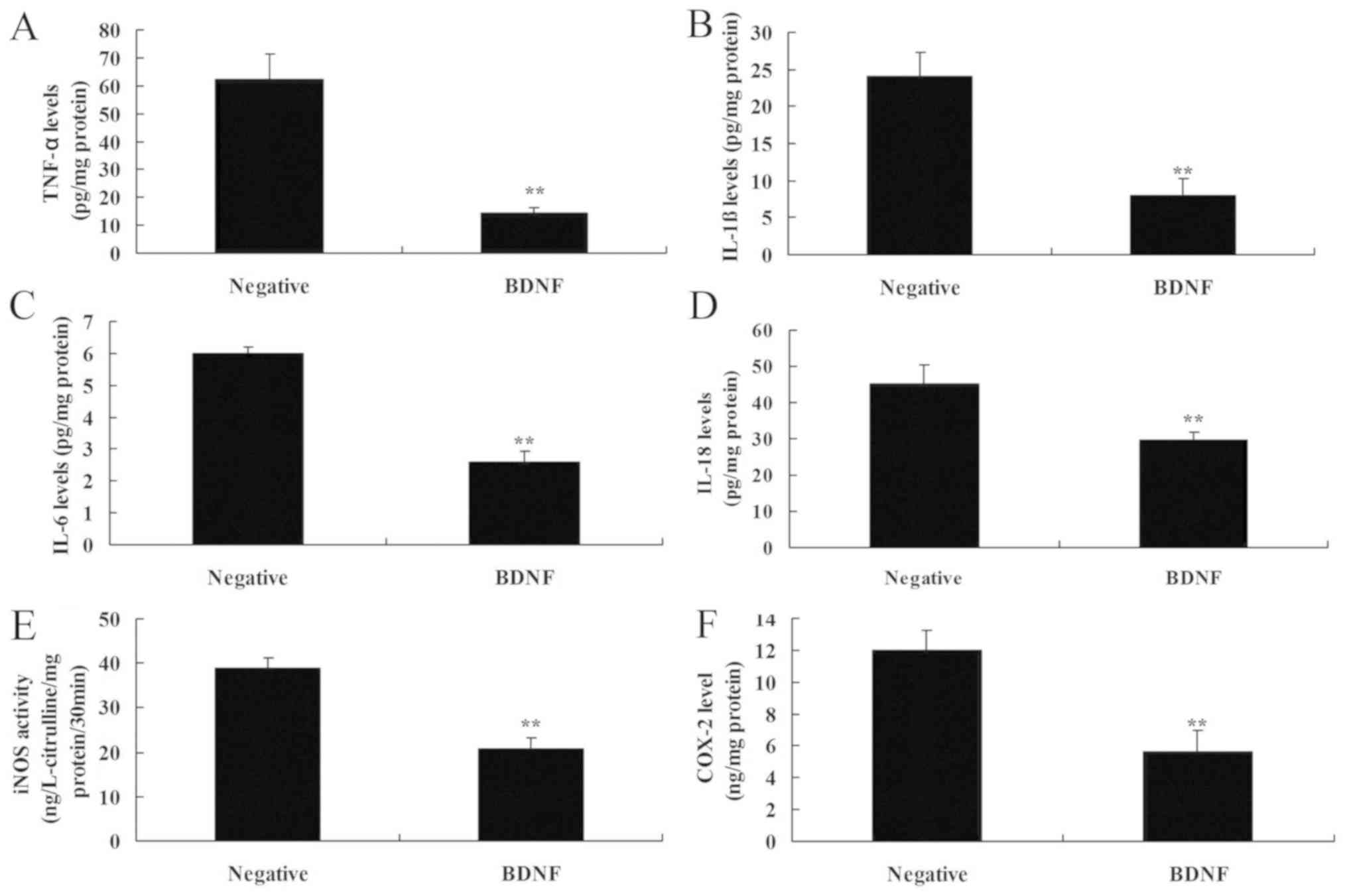
Overexpression of BDNF reduces
inflammation in vitro
Overexpression of BDNF reduced the levels of IL-1β,
IL-6, IL-18 and TNF-α in vitro, compared with the negative
group (Fig. 3A-D). In addition,
overexpression of BDNF decreased the levels of iNOS and COX-2
levels in vitro, compared with the negative group (Fig. 3E and F). These data indicated that
BDNF reduced inflammation in SCI via the TrkB/p38 mitogen-activated
protein kinase (MAPK) signaling pathway.
Inhibition of TrkB increases the
anti-inflammation effects of BDNF on TrkB and p-p-38 protein
expressions in vitro
The role of TrkB in the anti-inflammation effects of
BDNF on inflammation in SCI was next investigated. As presented in
Fig. 4, the TrkB inhibitor, ANA-12
suppressed the protein expression of TrkB, increased that of
p-p-38, promoted the levels of IL-1β, IL-6, IL-18 and TNF-α levels
and increased those of iNOS and COX-2 in vitro of SCI by
BDNF overexpression, compared with the BDNF overexpression group.
These findings suggest that overexpression of BDNF may be
associated with inflammation of SCI via TrkB.
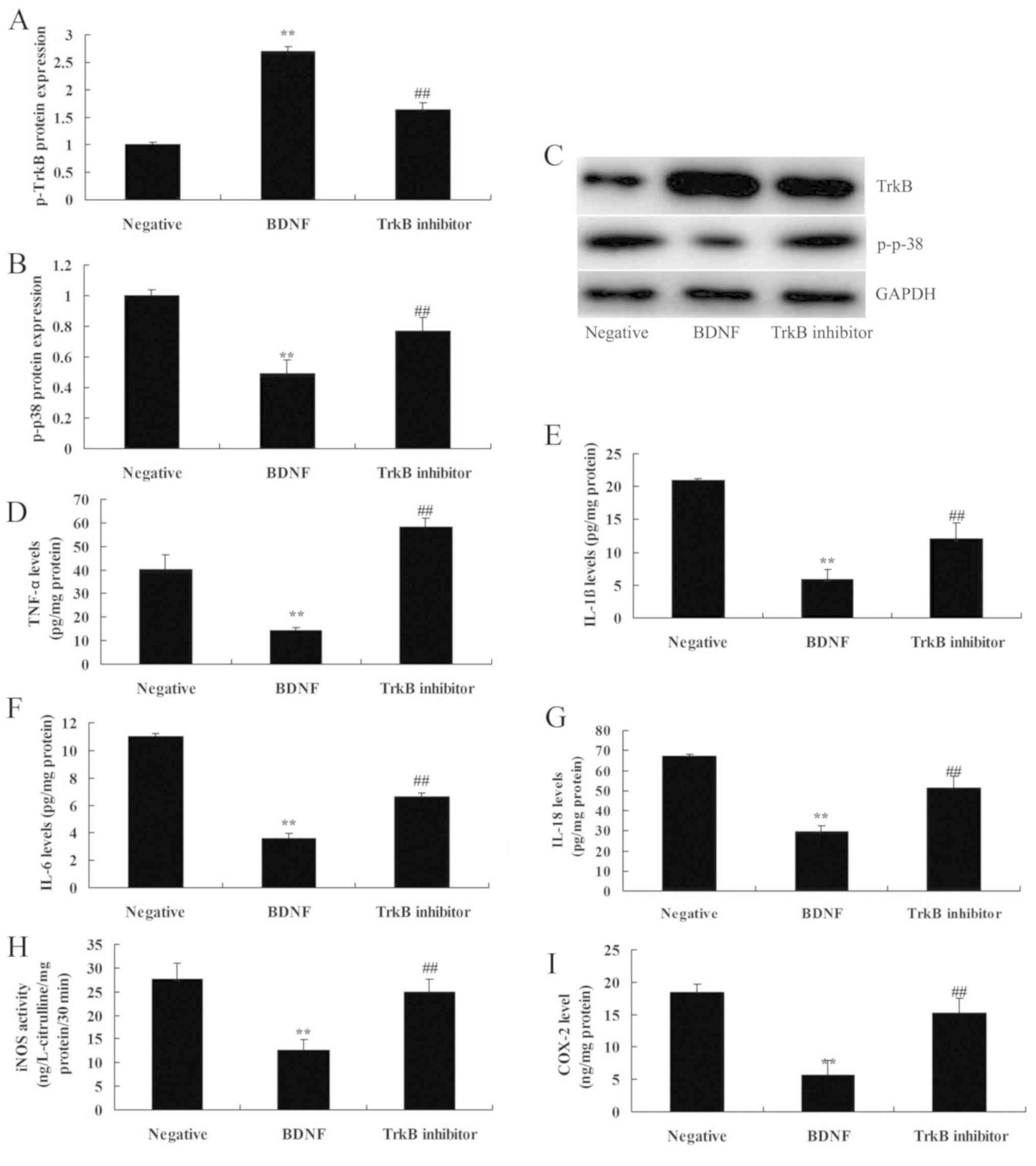 | Figure 4.Inhibition of p-TrkB increases the
anti-inflammation effects of BDNF on TrkB and p-p-38 protein
expressions in vitro. (A) TrkB and (B) p-p-38 protein
expression, as determined using statistical analysis, (C) Western
blot analysis of BDNF protein expression, (D) TNF-α, (E) IL-1β, (F)
IL-6, (G) IL-18, (H) iNOS and (I) COX2 levels. Negative, negative
mimics group; BDNF, Overexpression of BDNF group; TrkB
inhibitor, Overexpression of BDNF and TrkB inhibitor group.
Data are presented as the mean ± standard error of the mean for
three independent experiments. **P<0.01 compared with I group;
##P<0.01 compared with II group. TrkB, tyrosine
kinase receptor B; BDNF, brain-derived neurotrophic factor; TNF-α,
tumor necrosis factor-α; IL, interleukin; iNOS, inducible nitric
oxide synthase; COX2, cyclooxygenase-2. |
Inhibition of p38 increases the
anti-inflammation effects of BDNF on inflammation in vitro
Finally, the role of p38 was determined in
anti-inflammation effects of BDNF on inflammation in SCI. As
presented in Fig. 5, p-38 inhibitor,
TA-02, suppressed p-38 protein expression, reduced IL-1β, IL-6,
IL-18 and TNF-α levels and inhibited iNOS and COX-2 levels in an
in vitro model of SCI by BDNF overexpression, compared with
the BDNF overexpression group. Therefore, these data indicated that
BDNF attenuated inflammation in SCI by regulating the TrkB/p-38
pathway.
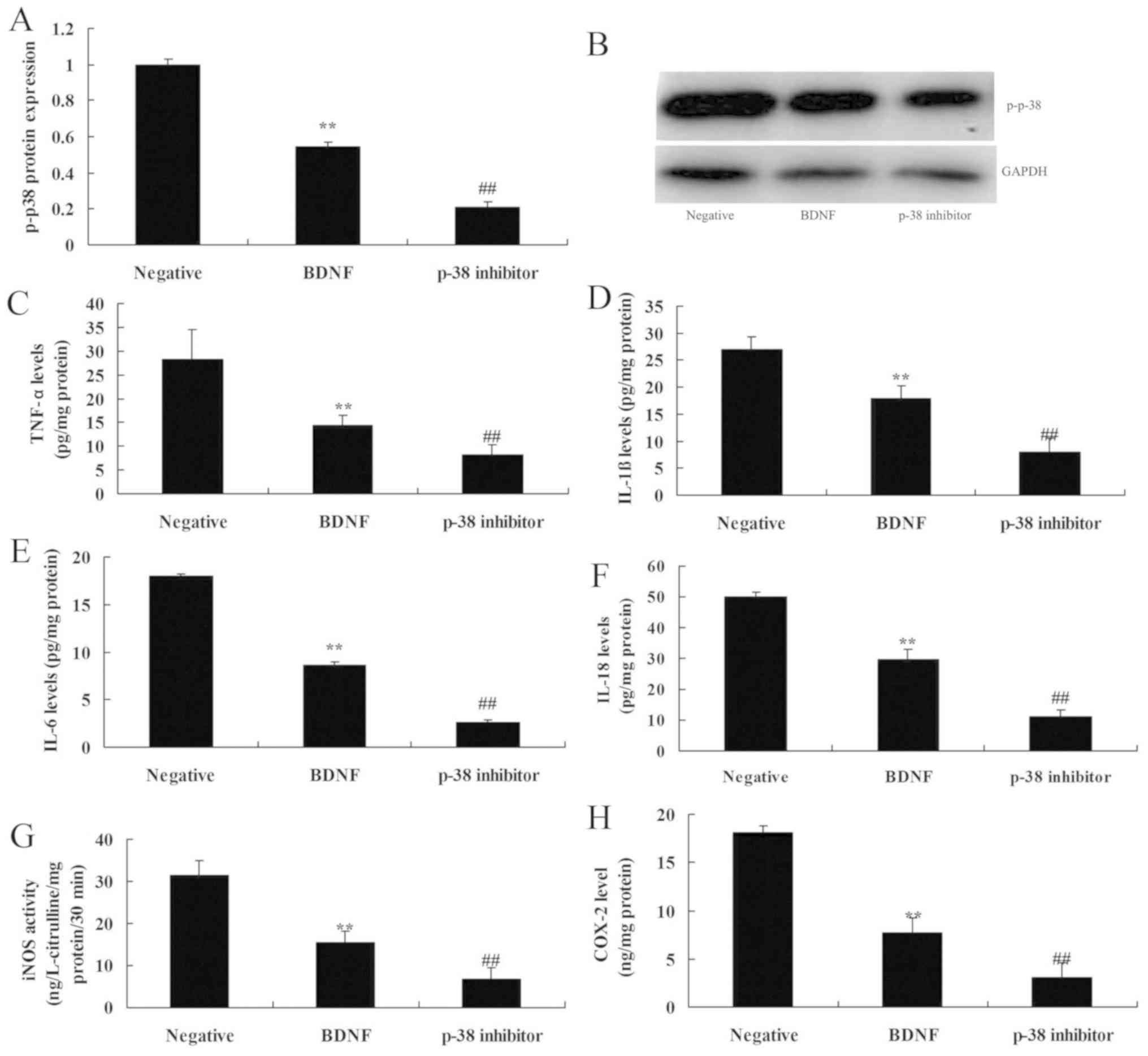 | Figure 5.Inhibition of p38 increases the
anti-inflammation effects of BDNF on inflammation in vitro.
p-p-38 protein expression (A) using statistical analysis, (B)
Western blot analysis of BDNF protein expression, (C) TNF-α, (D)
IL-1β, (E) IL-6, (F) IL-18, (G) iNOS and (H) COX2 levels. Negative,
negative mimics group; BDNF, Overexpression of BDNF group; p-p38
inhibitor, Overexpression of BDNF and p-p38 inhibitor group. Data
are presented as the mean + standard error of the mean for three
independent experiments. **P<0.01 compared with I group;
##P<0.01 compared with II group. BDNF, brain-derived
neurotrophic factor; TNF-α, tumor necrosis factor-α; IL,
interleukin; iNOS, inducible nitric oxide synthase; COX2,
cyclooxygenase-2. |
Discussion
SCI refers to serious damage of the central nervous
system. In general, serious neurologic impairment may occur
following SCI (19). However, motor
dysfunction is a principal factor leading to the loss of ability of
daily activities (20). Therefore,
motor dysfunction repair of SCI patients has been a focus of
doctors and patients (21). However,
motor function training is an important method for motor function
rehabilitation (21). Previous
studies have demonstrated that persistent nursing intervention at
early stages can partially promote the rehabilitation of motor
functions (21,22). However, the underlying mechanisms
have not yet been illustrated and more fundamental research is
required. The results of the present study suggested that the
expression of BDNF was reduced in SCI rats.
IL-1β and IL-6 are cytokines with broad biological
effects. Various types of cell produce IL-1β and IL-6, such as
monocytes, B cells, T cells, fibroblasts and endothelial cells
(23). IL-1β and IL-6 may originate
from microglial cells and astrocytes, and be secreted by neurons at
early stages of SCI (24).
Neurogliocytes can regulate cell growth and functions in the
central nervous system and participate in the inflammatory response
and immunoreaction (25). It is
potentially an important factor for amplification of the
inflammatory response at damage regions and secondary immune injury
following primary injury (25). In
the present study, it was demonstrated that overexpression of BDNF
induces TrkB and p-p-38 protein expressions in vitro.
BDNF mainly functions to promote the survival of
neurons and regeneration of protuberance (26). Previous studies have suggested that
following SCI, expression levels of BDNF in mice would re-actively
increase (26). Following SCI, the
requirement of BDNF by dynamoneure is enhanced (26,27).
Endogenous BDNF originates from Schwann cells and muscular tissues
(28), and serves an important role
in remyelination of the axon (28).
Endogenous BDNF also participates in the regulation of
growth-associated protein-43 (28).
Neurotrophic factors are a type of polypeptide or
protein factor (29). They can
promote the growth of nerve cells, survival and differentiation.
Neurotrophic factors regulate the survival of neurons during
neuro-development and activate enzyme activities biochemically and
physiologically (27). Furthermore,
they can prevent the death of neurons following injury, and promote
the repair of neurons and axonal regeneration (27). Furthermore, they can also regulate
nervous system functions, including plasticity of synapses and
neurotransmitter transmission (25).
The present study demonstrates that overexpression of BDNF reduces
SCI inflammation in vitro. Chang et al (30) previously demonstrated that 7,8-DHF,
produces fast-onset antidepressant-like effects in rats exposed to
chronic mild stress via BDNF levels. The present results
demonstrated that DHF regulates BDNF to prevent SCI in a rat
model.
TrkB is expressed in neurons in the central nervous
system, including the spinal cord, cerebral cortex, epencephalon
and olfactory bulb (31). Following
SCI, TrkB expression in a high-zinc dietary group is markedly
higher than that in an appropriate zinc group (32). TrkB protein expression in a zinc
deficiency group is relatively low (32). High expression of TrkB is possibly
conducive to the increase of BDNF expression, which may reduce the
possibility of secondary injury following acute SCI (32). In addition, it may facilitate the
recovery of the spinal cord. Previous studies have demonstrated
that zinc released from synaptic vesicle has protective effects on
ischemic neuron injury (32). Zinc
deficiency may lead to death of nerve cells following SCI (32). In the present study, treatment with
DHF effectively induced SCI-inhibited TrkB signaling pathway in SCI
rats. Han et al (33)
suggested that 7,8-DHF prevents onset of psychosis in adult
offspring following maternal immune activation via BDNF-TrkB
signaling, and demonstrated that DHF reduced inflammation via
BDNF-TrkB signaling.
As one member of MAPK family, extracellular
signal-regulated kinase is widely prevalent in the central nervous
system (34). Following its
activation of phosphorylation, extracellular signal-regulated
kinase may cause proliferation and activity of microglial cells,
and also secrete cytokines and TNF, promote inflammatory responses
following injuries and eliminate necrotic tissues (34). Inhibition of TrkB or p38 increases
the anti-inflammation effects of BDNF on TrkB and p-p-38 protein
expression in vitro. Park et al (35) previously reported that 7,8-DHF
attenuates inflammation through the suppression of the nuclear
factor-κB and MAPK signaling pathways. BDNF-TrkB-p38 signaling is
an important signaling pathway of treatment with DHF for SCI. In
the present study, it was only analyzed whether 7,8-DHF regulates
BDNF-TrkB-p38 signaling.
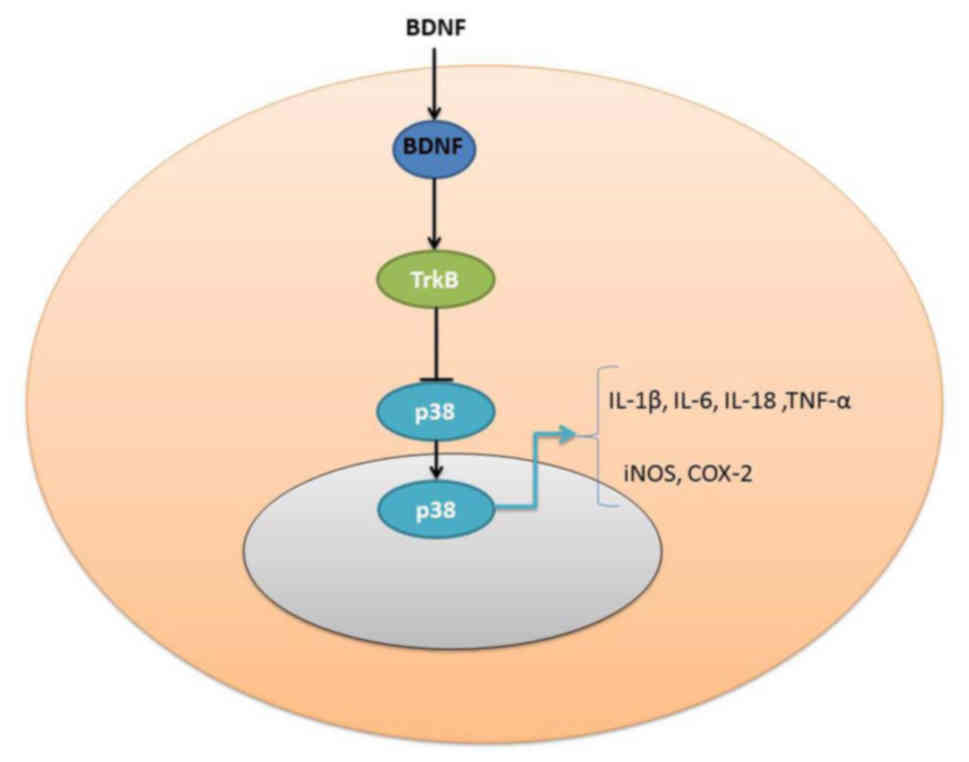
Taken together, the present results indicate that
the expression of BDNF was reduced in SCI rats and that the
overexpression of BDNF reduces SCI inflammation in vitro
through the induction of TrkB and p-p-38 protein expression.
Furthermore, the inhibition of TrkB increases the anti-inflammation
effects of BDNF on TrkB and p-p-38 protein expression in
vitro, which inhibited the effects of BDNF to reduce
inflammation in SCI model (Fig.
6).
Acknowledgements
Not applicable.
Funding
The present study was supported in part by grants
from the National Natural Science Foundation of China (grant no.
81401014), the Young Scientists Awards Foundation of Shandong
Province (grant no. BS2013YY049) and the China Postdoctoral Science
Foundation (grant nos. 2012M511036 and 2015T80725).
Availability of data and materials
The analyzed data sets generated during the study
are available from the corresponding author on reasonable
request.
Authors' contributions
JL designed the experiments of the present study; GD
and HH performed the experiments; JL and GD analyzed the data; and
JL wrote the manuscript.
Ethics approval and consent to
participate
All animal studies were approved by the Committee of
Ethics on Animal Experiments of The First People's Hospital of
Three Gorges University (Yichang, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
de Araujo AVL, Barbosa VRN, Galdino GS,
Fregni F, Massetti T, Fontes SL, de Oliveira Silva D, da Silva TD,
Monteiro CBM, Tonks J and Magalhães FH: Effects of high-frequency
transcranial magnetic stimulation on functional performance in
individuals with incomplete spinal cord injury: Study protocol for
a randomized controlled trial. Trials. 18:5222017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Nakashima A, Yamanaka-Tatematsu M, Fujita
N, Koizumi K, Shima T, Yoshida T, Nikaido T, Okamoto A, Yoshimori T
and Saito S: Impaired autophagy by soluble endoglin, under
physiological hypoxia in early pregnant period, is involved in poor
placentation in preeclampsia. Autophagy. 9:303–316. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Pons J, Huang Y, Arakawa-Hoyt J, Washko D,
Takagawa J, Ye J, Grossman W and Su H: VEGF improves survival of
mesenchymal stem cells in infarcted hearts. Biochem Biophys Res
Commun. 376:419–422. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Tsuzuki T, Okada H, Cho H, Tsuji S,
Nishigaki A, Yasuda K and Kanzaki H: Hypoxic stress simultaneously
stimulates vascular endothelial growth factor via hypoxia-inducible
factor-1α and inhibits stromal cell-derived factor-1 in human
endometrial stromal cells. Hum Reprod. 27:523–530. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ottosen LD, Hindkaer J, Husth M, Petersen
DE, Kirk J and Ingerslev HJ: Observations on intrauterine oxygen
tension measured by fibre-optic microsensors. Reprod Biomed Online.
13:380–385. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kuhn NZ and Tuan RS: Regulation of
stemness and stem cell niche of mesenchymal stem cells:
Implications in tumorigenesis and metastasis. J Cell Physiol.
222:268–277. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kato K: Stem cells in human normal
endometrium and endometrial cancer cells: Characterization of side
population cells. Kaohsiung J Med Sci. 28:63–71. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Schwab KE, Hutchinson P and Gargett CE:
Identification of surface markers for prospective isolation of
human endometrial stromal colony-forming cells. Hum Reprod.
23:934–943. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Miernik K and Karasinski J: Porcine uterus
contains a population of mesenchymal stem cells. Reproduction.
143:203–209. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhang L, Xiong W, Xiong Y, Liu H, Li N, Du
Y and Liu Y: Intracellular Wnt/beta-catenin signaling underlying
17beta-estradiol-induced matrix metalloproteinase 9 expression in
human endometriosis. Biol Reprod. 94:702016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Cosar E, Mamillapalli R, Ersoy GS, Cho S,
Seifer B and Taylor HS: Serum microRNAs as diagnostic markers of
endometriosis: A comprehensive array-based analysis. Fertil Steril.
106:402–409. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Panda H, Pelakh L, Chuang TD, Luo X,
Bukulmez O and Chegini N: Endometrial miR-200c is altered during
transformation into cancerous states and targets the expression of
ZEBs, VEGFA, FLT1, IKKβ, KLF9, and FBLN5. Reprod Sci. 19:786–796.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Furuya M, Masuda H, Hara K, Uchida H, Sato
K, Sato S, Asada H, Maruyama T, Yoshimura Y, Katabuchi H, et al:
ZEB1 expression is a potential indicator of invasive endometriosis.
Acta Obstet Gynecol Scand. 96:1128–1135. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Sanchez AM, Vigano P, Quattrone F,
Pagliardini L, Papaleo E, Candiani M and Panina-Bordignon P: The
WNT/β-catenin signaling pathway and expression of survival
promoting genes in luteinized granulosa cells: Endometriosis as a
paradigm for a dysregulated apoptosis pathway. Fertil Steril.
101:1688–1696. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kim SH, Park JG, Lee J, Yang WS, Park GW,
Kim HG, Yi YS, Baek KS, Sung NY, Hossen MJ, et al: The dietary
flavonoid Kaempferol mediates anti-inflammatory responses via the
Src, Syk, IRAK1, and IRAK4 molecular targets. Mediators Inflamm.
2015:9041422015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhang JC, Yao W and Hashimoto K:
Brain-derived neurotrophic factor (BDNF)-TrkB signaling in
inflammation-related depression and potential therapeutic targets.
Curr Neuropharmacol. 14:721–731. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Basso DM, Beattie MS, Bresnahan JC,
Anderson DK, Faden AI, Gruner JA, Holford TR, Hsu CY, Noble LJ,
Nockels R, et al: MASCIS evaluation of open field locomotor scores:
Effects of experience and teamwork on reliability. Multicenter
Animal Spinal Cord Injury Study. J Neurotrauma. 13:343–359. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Cindolo L, Chiodini P, Brookman-May S, De
Cobelli O, May M, Squillacciotti S, De Nunzio C, Tubaro A, Coman I,
Feciche B, et al: Assessing the accuracy and generalizability of
the preoperative and postoperative karakiewicz nomograms for renal
cell carcinoma: Results from a multicentre European and US study.
BJU Int. 112:578–584. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Masuda H, Maruyama T, Gargett CE, Miyazaki
K, Matsuzaki Y, Okano H and Tanaka M: Endometrial side population
cells: Potential adult stem/progenitor cells in endometrium. Biol
Reprod. 93:842015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Siebels M, Rohrmann K, Oberneder R,
Stahler M, Haseke N, Beck J, Hofmann R, Kindler M, Kloepfer P and
Stief C: A clinical phase I/II trial with the monoclonal antibody
cG250 (RENCAREX(R)) and interferon-alpha-2a in metastatic renal
cell carcinoma patients. World J Urol. 29:121–126. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Opper B, Elsner P and Ziemer M: Cutaneous
metastasis of renal cell carcinoma. Am J Clin Dermatol. 7:271–272.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Liang W, Guan H, He X, Ke W, Xu L, Liu L,
Xiao H and Li Y: Down-regulation of SOSTDC1 promotes thyroid cancer
cell proliferation via regulating cyclin A2 and cyclin E2.
Oncotarget. 6:31780–31791. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Jhaveri KS, Elmi A, Hosseini-Nik H,
Hedgire S, Evans A, Jewett M and Harisinghani M: Predictive value
of chemical-shift MRI in distinguishing clear cell renal cell
carcinoma from non-clear cell renal cell carcinoma and minimal-fat
angiomyolipoma. AJR Am J Roentgenol. 205:W79–W86. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Rini BI, Plimack ER, Takagi T, Elson P,
Wood LS, Dreicer R, Gilligan T, Garcia J, Zhang Z, Kaouk J, et al:
A Phase II study of pazopanib in patients with localized renal cell
carcinoma to optimize preservation of renal parenchyma. J Urol.
194:297–303. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Guo R, Overman M, Chatterjee D, Rashid A,
Shroff S and Wang H, Katz MH, Fleming JB, Varadhachary GR,
Abbruzzese JL and Wang H: Aberrant expression of p53, p21, cyclin
D1, and Bcl2 and their clinicopathological correlation in ampullary
adenocarcinoma. Hum Pathol. 45:1015–1023. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Erdem H, Oktay M, Yildirim U, Uzunlar AK
and Kayikci MA: Expression of AEG-1 and p53 and their
clinicopathological significance in malignant lesions of renal cell
carcinomas: A microarray study. Pol J Pathol. 64:28–32. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Mishra SK and Crasta JA: An
immunohistochemical comparison of P53 and Bcl-2 as apoptotic and
MIB1 as proliferative markers in low-grade and high-grade ovarian
serous carcinomas. Int J Gynecol Cancer. 20:537–541. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Liu G, Wang T, Wang T, Song J and Zhou Z:
Effects of apoptosis-related proteins caspase-3, Bax and Bcl-2 on
cerebral ischemia rats. Biomed Rep. 1:861–867. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Chang HA, Wang YH, Tung CS, Yeh CB and Liu
YP: 7,8-Dihydroxyflavone, a tropomyosin-kinase related receptor B
agonist, produces fast-onset antidepressant-like effects in rats
exposed to chronic mild stress. Psychiatry Investig. 13:531–540.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Guo YB, Bao XJ, Xu SB, Zhang XD and Liu
HY: Honokiol induces cell cycle arrest and apoptosis via p53
activation in H4 human neuroglioma cells. Int J Clin Exp Med.
8:7168–7175. 2015.PubMed/NCBI
|
|
32
|
De Giorgi U, Yuan J, Moroni M, Veronese S,
Sartore-Bianchi A, Broggini M, Rosti G, Strebhardt K and Ruffini
PA: Germ cell tumors overexpress the candidate therapeutic target
cyclin B1 independently of p53 function. Int J Biol Markers.
30:e275–e281. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Han M, Zhang JC, Yao W, Yang C, Ishima T,
Ren Q, Ma M, Dong C, Huang XF and Hashimoto K: Intake of
7,8-dihydroxyflavone during juvenile and adolescent stages prevents
onset of psychosis in adult offspring after maternal immune
activation. Sci Rep. 6:360872016. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Zhuang Z, Ye G and Huang B: Kaempferol
alleviates the interleukin-1β-induced inflammation in rat
osteoarthritis chondrocytes via suppression of NF-kB. Med Sci
Monit. 23:3925–3931. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Park HY, Park C, Hwang HJ, Kim BW, Kim GY,
Kim CM, Kim ND and Choi YH: 7,8-Dihydroxyflavone attenuates the
release of pro-inflammatory mediators and cytokines in
lipopolysaccharide-stimulated BV2 microglial cells through the
suppression of the NF-κB and MAPK signaling pathways. Int J Mol
Med. 33:1027–1034. 2014. View Article : Google Scholar : PubMed/NCBI
|