Introduction
Bronchial asthma is a clinically common respiratory
chronic inflammatory disease, which mainly involves inflammatory
cells and structural cells (such as eosinophils, mast cells,
lymphocytes, neutrophils, smooth muscle cells and airway epithelial
cells), as well as relevant cellular components (1,2). Without
effective control, chronic inflammation induced by asthma will lead
to airway hyperresponsiveness, which further causes reversible
airflow limitation, as well as recurrent gasp for breath,
tachypnea, chest tightness or cough (3). Clinical studies show that the number of
asthma patients around the world is as high as 300 million, and the
prevalence of the disease varies from 1 to 18% across different
countries (4,5). In China, the number of asthma patients
is more than 30 million, and the mortality caused by asthma is the
highest in the world (6,7). Of note, childhood asthma attracts more
and more concern in medical practice, because this disease affects
children and is easy to cause death (8). At present, long-term inhalation of
corticosteroids is the main clinical treatment for children with
asthma, and it greatly alleviates the illness of the children
(9). However, long-term use of
corticosteroids causes hormone resistance or dependence, greatly
affects the growth and development of children, and induces related
complications, such as necrosis of femoral head (9). Therefore, it is of important clinical
value to explore the molecular mechanism of childhood asthma, to
discover potential drug targets and to find effective treatment
strategies. Existing studies have shown that chronic airway
inflammation, airway hyperresponsiveness, and airway remodeling are
the three pathological stages of asthma in children. Among these
stages, airway remodeling phase is irreversible, and directly
related to the prognosis of affected children (10). Airway smooth muscle cells play an
important role in this process (11), but the molecular mechanisms are still
unclear. It is reported that proteins in extracellular matrix play
important roles in airway remodeling induced by asthma, including
type I collagen (COL-I) and fibronectin (FN). The secretion of
COL-I and FN leads to thickening of basilar membrane and
subcutaneous fibrosis of the airway, eventually causing airway
remodeling (12).
microRNA (miRNA or miR) molecules are a class of
small non-encoding RNA molecules with 18–25 nucleotides. They form
silencing complexes by binding with the 3′-untranslated region
(UTR) of target genes to inhibit translation and reduce expression
of the target proteins (13,14). It is shown that miRNA molecules exist
widely in tissue cells and body fluids, participate in nearly all
pathophysiological processes, and have important clinical values
(15). miRNA molecules regulate the
biological functions of immune cells, smooth muscle cells and
epithelial cells through a variety of target genes (16). For example, miR-425 regulates the
differentiation of Th17 cells by targeting the Foxo1 gene and
participates in intestinal inflammation (17). miR-135a promotes the inflammatory
response of rat vascular smooth muscle cells by targeting Foxo1
gene (18). In addition, the
expression of miR-221 in fibrotic airway epithelial cells is
obviously upregulated and it can target the expression of ATF6
(19). However, the molecular
mechanisms by which miRNA molecules regulates the occurrence of
asthma are not clear yet.
miR-378 was first discovered in tumor tissues and
cells, and it plays important roles in biological processes of
tumor cells, such as proliferation, apoptosis, and drug resistance
(20,21). Depending on the types of tumors,
miR-378 can play a role in promoting cancer or inhibiting cancer
(20,21). It is reported that miR-378 is closely
related to the differentiation, hypertrophy and metabolism of
cardiomyocytes, and is involved in the pathological changes of the
heart, suggesting that miR-378 may participate in the regulation of
biological functions of myocytes (22). As a main component in respiratory
tract remodeling, airway smooth muscle cells participate in the
initiation and development of airway remodeling. As structural
cells, airway smooth muscle cells not only directly participate in
the thickening of airway wall through its own hypertrophy and
proliferation, but also promote airway remodeling by phenotypic
transformation, changes in migration function, and secretion of
inflammatory factors and extracellular matrix (23,24). In
the present study, we determine the expression of miR-378 in
children with asthma, investigate its effect on the biological
functions of smooth muscle cells, and try to provide experimental
basis for understanding the pathogenesis of childhood asthma.
Patients and methods
Patients
A total of 23 asthmatic children who underwent
biopsy by bronchoscopy at our hospital between January 2014 and
January 2017 were included in the present study. In addition, 15
healthy children were included into control group. Sputum samples
were obtained from all patients and healthy children. Peripheral
blood (5 ml) was collected from patients and healthy subjects. All
patients had mild to moderate asthma and this was their first
treatment for asthma. None of the patients had used hormones within
four weeks before admission to our hospital. All procedures were
approved by the Ethics Committee of Maternity and Child Health Care
Hospital of Zibo City (Zibo, China). Written informed consents were
obtained from all patients or their families.
Cells
Lung tissues were obtained from non-asthmatic
patients undergoing lobectomy, and washed with phosphate-buffered
saline (PBS) thoroughly. Smooth muscle layer was separated from
middle bronchoalveolar membrane, cut into small pieces (about 1
mm3), placed on the bottom of culture flasks containing DMEM medium
supplemented with 20% fetal bovine serum, and cultured under 37°C
and 5% CO2. Medium was replaced every two days, and smooth muscle
cells that migrated out of the tissues were monitored under a light
microscope. When reaching 70–80% confluency, smooth muscle cells
were passaged at a ratio of 1:3 and DMEM supplemented with 10%
fetal bovine serum was used for cell culture.
To transfect smooth muscle cells with miR-378
mimics, the cells (2×105) in logarithmic growth were seeded onto
24-well plates one day before transfection, and cultured in
antibiotics-free DMEM medium supplemented with 10% fetal bovine
serum until reaching 70% confluency. In the first vial, 1.25 µl
miR-negative control (NC), miR-378 mimics or miR-378 inhibitor (20
pmol/µl; Hanbio Biotechnology Co., Ltd., Shanghai, China) was mixed
with 50 µl Opti Mem medium (Thermo Fisher Scientific, Inc.,
Waltham, MA, USA). In the second vial, 1 µl Lipofectamine 2000
(Thermo Fisher Scientific, Inc.) was mixed with 50 µl Opti Mem
medium. After standing still for 5 min, the two vials were combined
for additional waiting at room temperature for 20 min. Then, the
mixtures were added onto cells in respective groups. Six hours
later, the medium was replaced with DMEM containing 10% fetal
bovine serum. After cultivation for 48 h, the cells were collected
for further assays.
To test the effect of serum from asthmatic children
on smooth muscle cells, the cells (2×105) in logarithmic growth
phase were seeded onto 24-well plates, and cultured in DMEM
supplemented with 10% fetal bovine serum. On the next day, 250 µl
serum from healthy children (NC group) or asthmatic children
(asthma serum group) was mixed with 250 µl DMEM, and then added
onto smooth muscle cells. The cells were cultured for 7 days,
during which the medium was replaced every two days.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Sputum samples (0.2 ml) were mixed with saline at a
ratio of 1:10, and centrifuged at 12,000 × g and 4°C for 10 min to
collect sediments. Then, the sediments were lysed using 1 ml TRIzol
reagent following the manufacturer's manual (Thermo Fisher
Scientific, Inc.). Total RNA was extracted using phenol chloroform
method. The concentration and quality of RNA was measured using
ultraviolet spectrophotometry (Nanodrop ND2000; Thermo Fisher
Scientific, Inc., Wilmington, DE, USA). Then, cDNA was obtained by
reverse transcription from 1 µg RNA and stored at −20°C. Reverse
transcription of miRNA was carried out using miScript II RT kit
(Qiagen GmbH, Hilden, Germany) following the manufacturer's
manual.
The expression of miR-378 was determined by miScript
SYBR-Green PCR kit (Qiagen GmbH), using U6 as internal reference.
The sequences of miR-378 primers were forward,
5′-CTCCTGACTCCAGGTCCTGTGT-3′ and reverse,
5′-ACTGGACTTGGAGTCAGAAGGC-3′; and the sequences of U6 primers were
forward, 5′-CTCGCTTCGGCAGCACA-3′ and reverse,
5′-AACGCTTCACGAATTTGCGT-3′. The reaction system (30 µl) contained
10 µl RT-qPCR-Mix, 1 µl upstream primer, 1 µl downstream primer, 5
µl cDNA and 13 µl ddH2O. The reaction protocol was: Initial
denaturation at 95°C for 5 min; 40 cycles of denaturation at 95°C
for 30 sec and annealing at 60°C for 30 sec (iQ5; Bio-Rad
Laboratories, Inc., Hercules, CA, USA). The 2−ΔΔCt
method was used to calculate the relative expression of miR-378
against U6. Each sample was tested in triplicate.
Immunohistochemistry
Smooth muscle cells (1×105) were seeded onto cell
slides. When reaching 90% confluency, medium was discarded, and the
cells were washed with PBS twice. After fixing with 4% formaldehyde
at room temperature for 10 min, the cells were washed with PBS
twice again. Then, the cells were incubated with α-smooth muscle
actin (α-SMA) antibody (1:5 dilution) at s4°C overnight. On the
next day, the cells were mixed with 3% H2O2 to block the activity
of peroxidase. Then, horseradish peroxidase-labelled secondary
antibody was added and the cells were incubated at room temperature
for 2 h to develop color.
CCK-8 assay
Cells were seeded at a density of 2,000/well in
96-well plates. At 0, 24, 48 and 72 h, 20 µl CCK-8 (5 g/l) was
added onto the cells. After incubation at 37°C for 2 h, the
absorbance of each well was measured at 490 nm for plotting cell
proliferation curves. Each group was tested in 3 replicate wells
and the values were averaged.
Flow cytometry
At 24 h after transfection, smooth muscle cells
(1×106) in each group were washed with pre-cooled PBS twice and
subjected to flow cytometry using Cell Cycle Assay kit (BD
Biosciences, Franklin Lakes, NJ, USA) following the manufacturer's
manual to detect cell cycle. The data were analyzed using ModFit
software (Verity Software House, Topsham, ME, USA).
At 24 h after being treated with serum from healthy
children or asthmatic children, or at 24 h after transfection with
miR-378 mimics, smooth muscle cells (1×106) in each group were
washed with pre-cooled PBS twice and subjected to flow cytometry
using Annexin V FITC Apoptosis DTEC kit I (BD Biosciences)
following the manufacturer's manual to detect cell apoptosis. Cells
with Annexin V-positive values were early apoptotic cells, those
with PI-positive values were necrotic cells, and those with double
positive values were late apoptotic cells.
Western blot analysis
Before lysis, tissues were ground into powder, and
cells were trypsinized and collected. Then, tissue samples or cells
were lysed with precooled Radio-Immunoprecipitation Assay (RIPA)
lysis buffer (600 µl; 50 mM Tris-base, 1 mM EDTA, 150 mM NaCl, 0.1%
sodium dodecyl sulfate, 1% TritonX-100, 1% sodium deoxycholate;
Beyotime Institute of Biotechnology, Shanghai, China) for 30 min on
ice. The mixture was centrifuged at 12,000 rpm and 4°C for 10 min.
The supernatant was used to determine protein concentration by
bicinchoninic acid (BCA) protein concentration determination kit
(RTP7102; Real-Times Biotechnology Co., Ltd., Beijing, China). The
samples were then mixed with 5× sodium dodecyl sulfate loading
buffer before denaturation in boiling water bath for 10 min.
Afterwards, the samples (5 µl) were subjected to 10% sodium dodecyl
sulfate-polyacrylamide gel electrophoresis at 100 V. The resolved
proteins were transferred to polyvinylidene difluoride membranes on
ice (250 mA, 1 h) and blocked with 50 g/l skimmed milk at room
temperature for 1 h. Then, the membranes were incubated with mouse
anti-human COL-I (1:1,000; ab90395; Abcam, Cambridge, UK), FN
(1:1,000; MAB1918; R&D Systems, Minneapolis, MN, USA) or GAPDH
(1:5,000; Beyotime Institute of Biotechnology) monoclonal primary
antibodies at 4°C overnight. After extensive washing with PBS with
Tween-20 for 3 times of 15 min, the membranes were incubated with
goat anti-mouse horseradish peroxidase-conjugated secondary
antibody (1:5,000; Santa Cruz, Dallas, TX, USA) for 1 h at room
temperature before washing with PBS with Tween-20 for 3 times of 15
min. Then, the membrane was developed with enhanced
chemiluminescence detection kit (Sigma-Aldrich; Merck KGaA,
Darmstadt, Germany) for imaging. Image lab v3.0 software (Bio-Rad
Laboratories, Inc.) was used to acquire and analyze imaging
signals. The relative contents of target proteins were expressed
against GAPDH.
Bioinformatics
On www.targetscan.org website, miR-378 was searched to
analyze potential target genes. Then, the target genes were
submitted on www.davidncifcrf.gov website for the analysis of
potential signaling pathways that involved the target genes.
Statistical analysis
The results were analyzed using SPSS 18.0
statistical software (SPSS Inc., Chicago, IL, USA). The data were
expressed as means ± standard deviations. Data were tested for
normality. Multigroup measurement data were analyzed using one-way
ANOVA. In case of homogeneity of variance, Least Significant
Difference and Student-Newman-Keuls methods were used; in case of
heterogeneity of variance, Tamhane's T2 or Dunnett's T3 method was
used. Comparison between two groups was carried out using Student's
t-test. P<0.05 was considered to indicate a statistically
significant difference.
Results
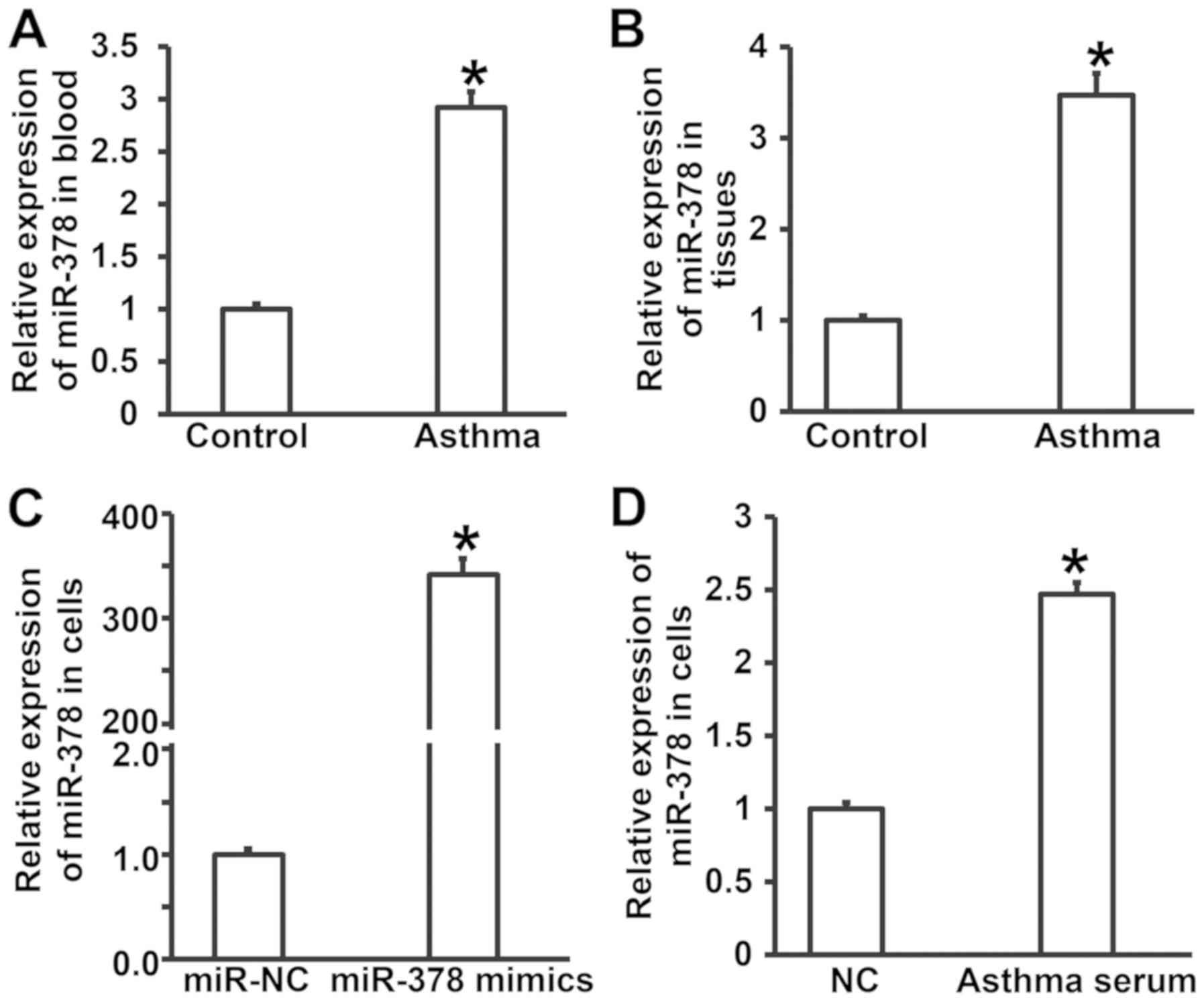
Expression of miR-378 in peripheral
blood and lung tissues from asthmatic children is increased
compared with that in healthy children
To measure the expression of miR-378 in peripheral
blood, lung tissues and smooth muscle cells, RT-qPCR was performed.
The data showed that the levels of miR-378 in peripheral blood and
tissues from asthmatic children were significantly higher than
those in control group (P<0.05) (Fig.
1A and B). Expression of miR-378 in smooth muscle cells
transfected with miR-378 mimics was significantly higher than that
in miR-NC group (P<0.05) (Fig.
1C). After incubating smooth muscle cells with serum from
asthmatic children for 7 days, the level of miR-378 in smooth
muscle cells was significantly higher than that of cells treated
with serum from healthy children (NC group) (P<0.05) (Fig. 1D). The results suggest that
expression of miR-378 in peripheral blood and lung tissues from
asthmatic children is increased compared with that in healthy
children.
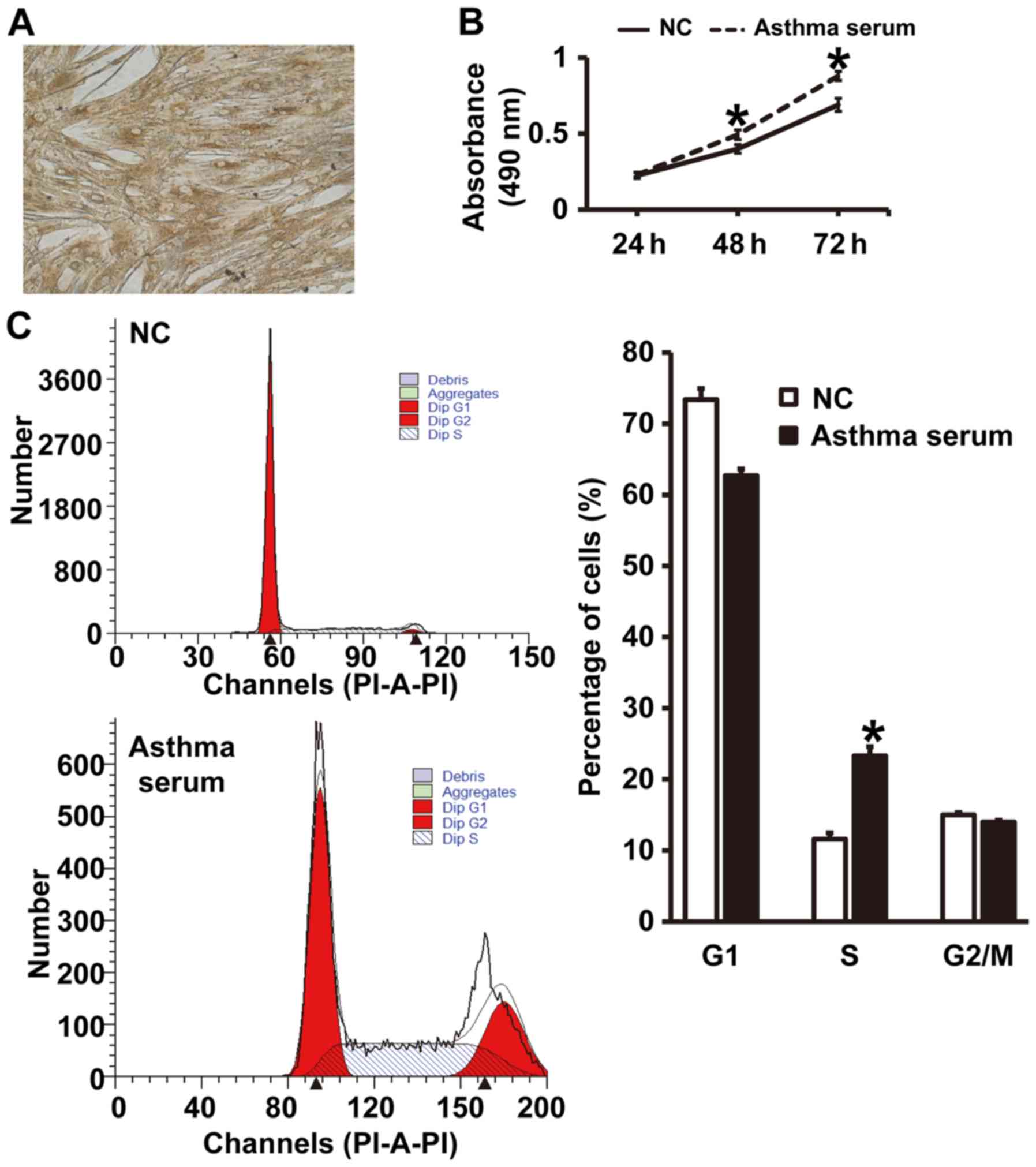
Serum from asthmatic children promotes
the proliferation of smooth muscle cells in vitro by affecting cell
cycle
To test the effect of serum from asthmatic children
on the proliferation and cell cycle of smooth muscle cells, CCK-8
assay and flow cytometry were carried out. The expression of α-SMA
in smooth muscle cells was identified by immunohistochemical
analysis (Fig. 2A), suggesting that
primary airway smooth muscle cells were successfully isolated.
CCK-8 assay showed that the absorbance of smooth muscle cells
treated with serum from asthmatic children at 48 and 72 h was
significantly higher than that of cells in NC group (P<0.05)
(Fig. 2B). In addition, the
percentage of S-phase cells among all smooth muscle cells treated
with asthma serum was significantly higher than that in NC group
(P<0.05) (Fig. 2C). The results
indicate that serum from asthmatic children promotes the
proliferation of smooth muscle cells in vitro by affecting
cell cycle.
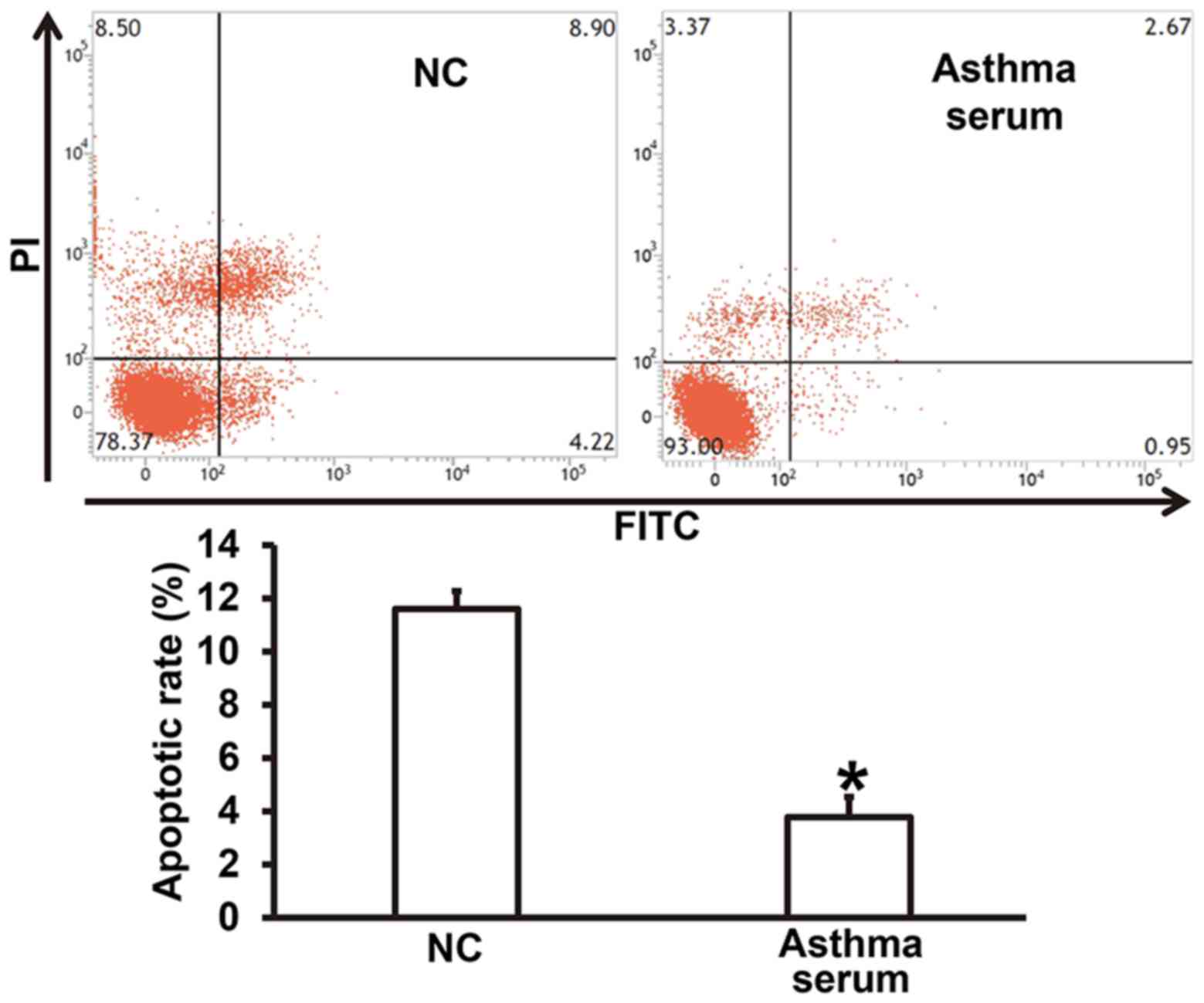
Serum from asthmatic children enhances
apoptosis resistance of smooth muscle cells
To examine how serum from asthmatic children affects
the apoptosis of smooth muscle cells, flow cytometry was used. The
data showed that treatment with serum from asthmatic children for 7
days reduced the apoptotic rate of smooth muscle cells compared
with NC group (P<0.05) (Fig. 3).
The result suggests that serum from asthmatic children enhances
apoptosis resistance of smooth muscle cells.
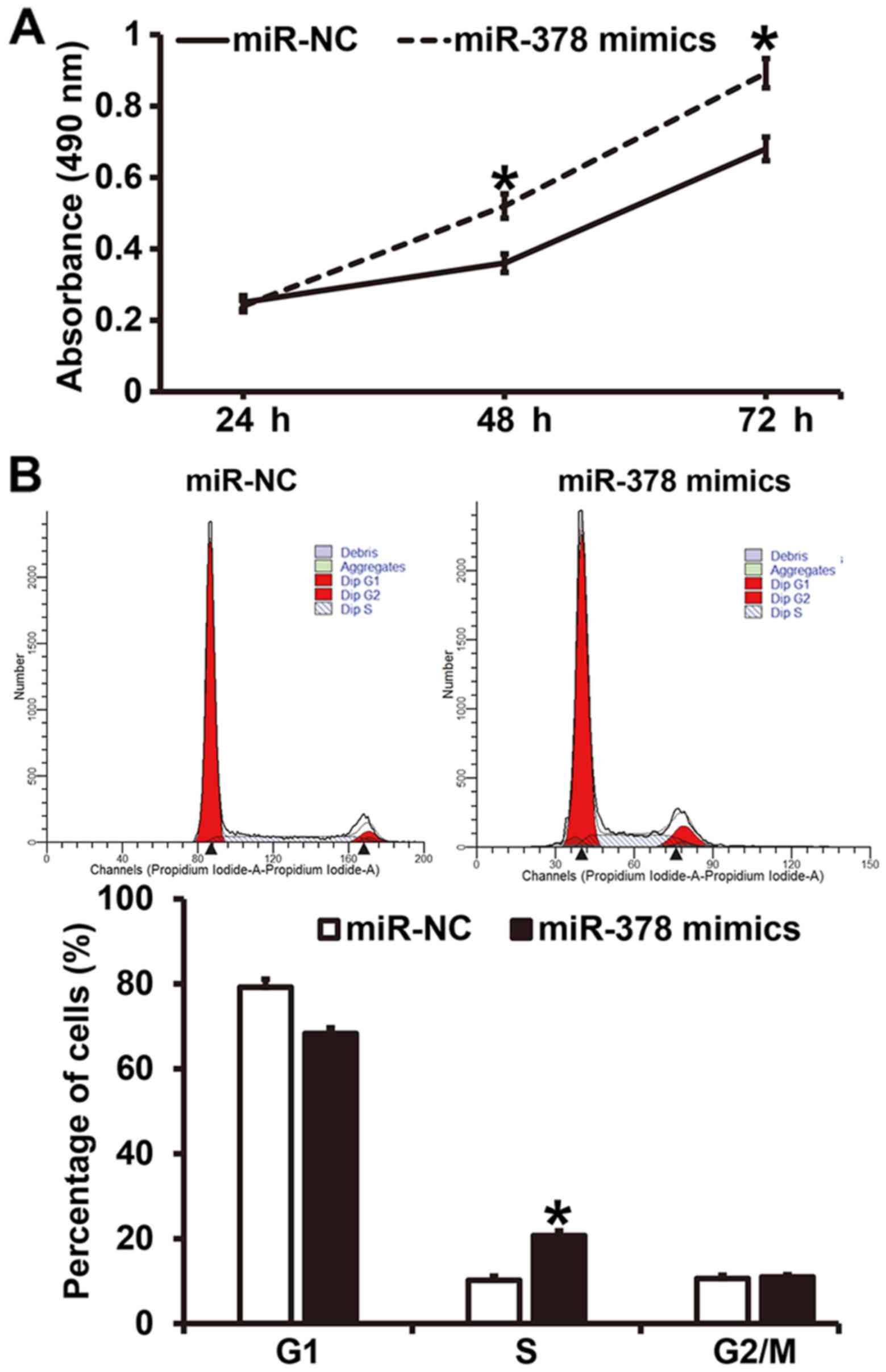
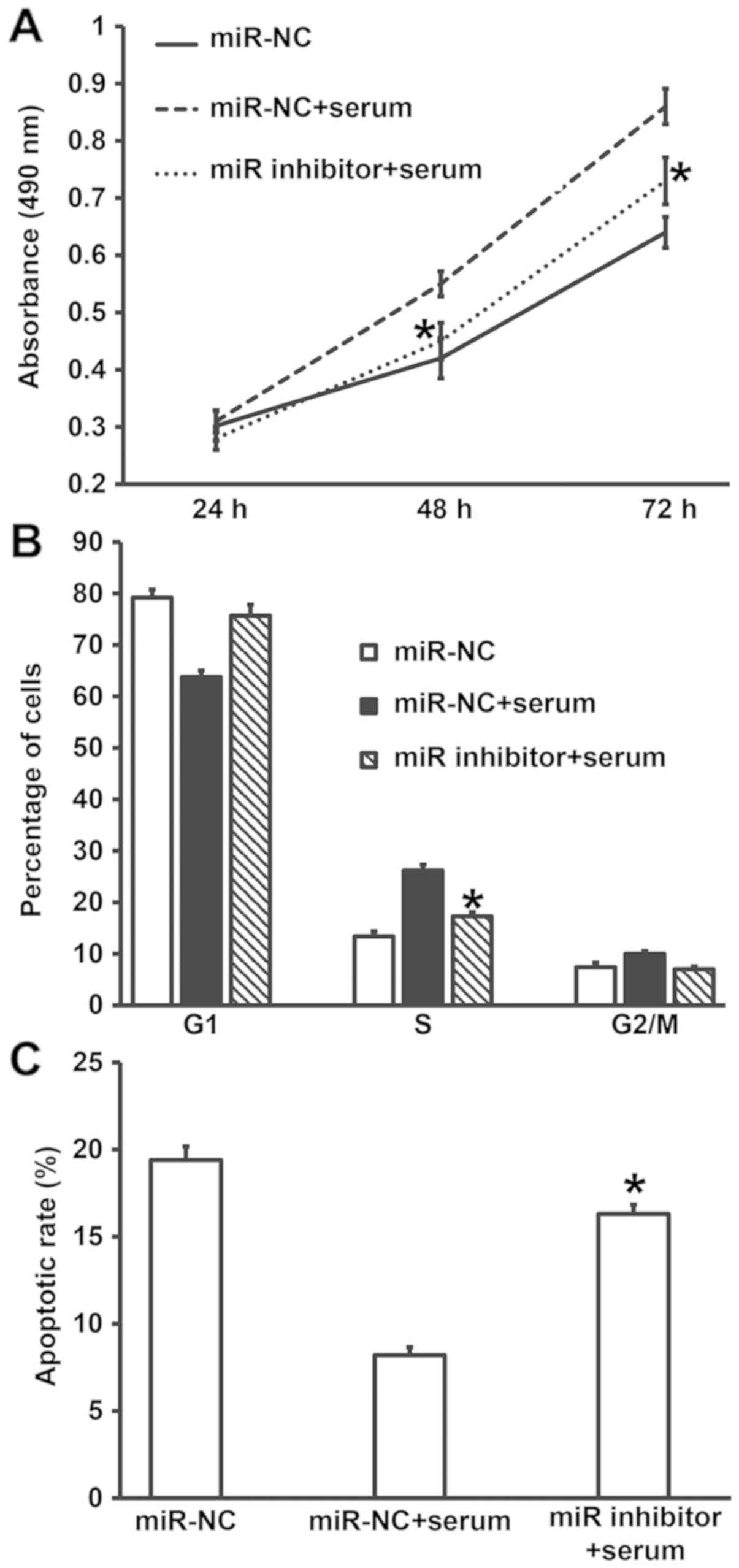
Overexpression of miR-378 increases
the proliferation of smooth muscle cells by affecting cell
cycle
To investigate how miR-378 affects the proliferation
and cell cycle of smooth muscle cells, CCK-8 assay and flow
cytometry were employed. CCK-8 assay showed that the absorbance of
smooth muscle cells transfected with miR-378 mimics at 48 and 72 h
was significantly higher than that of cells in miR-NC group
(P<0.05) (Fig. 4A). Similarly,
the percentage of S-phase cells among all smooth muscle cells
transfected with miR-378 mimics was significantly higher than that
in miR-NC group (P<0.05) (Fig.
4B). The results indicate that overexpression of miR-378
increases the proliferation of smooth muscle cells by affecting
cell cycle.
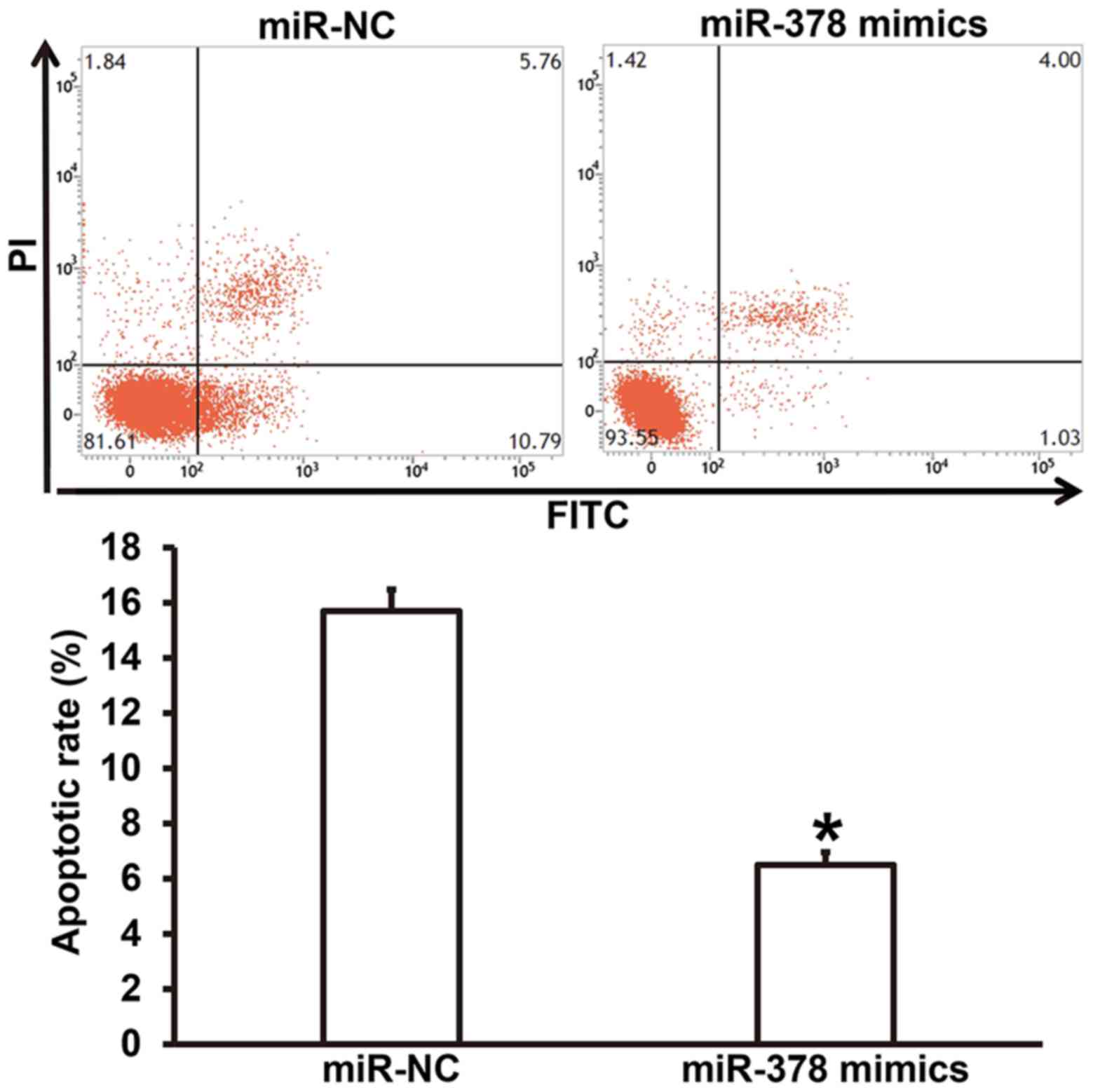
Overexpression of miR-378 upregulates
apoptosis resistance of smooth muscle cells
To test how miR-378 overexpression affects the
apoptosis of smooth muscle cells, flow cytometry was performed. The
data showed that the apoptotic rate of smooth muscle cells
transfected with miR-378 was significantly lower than that in
miR-NC group (P<0.05) (Fig. 5).
The result suggests that overexpression of miR-378
upregulatesapoptosis resistance of smooth muscle cells.
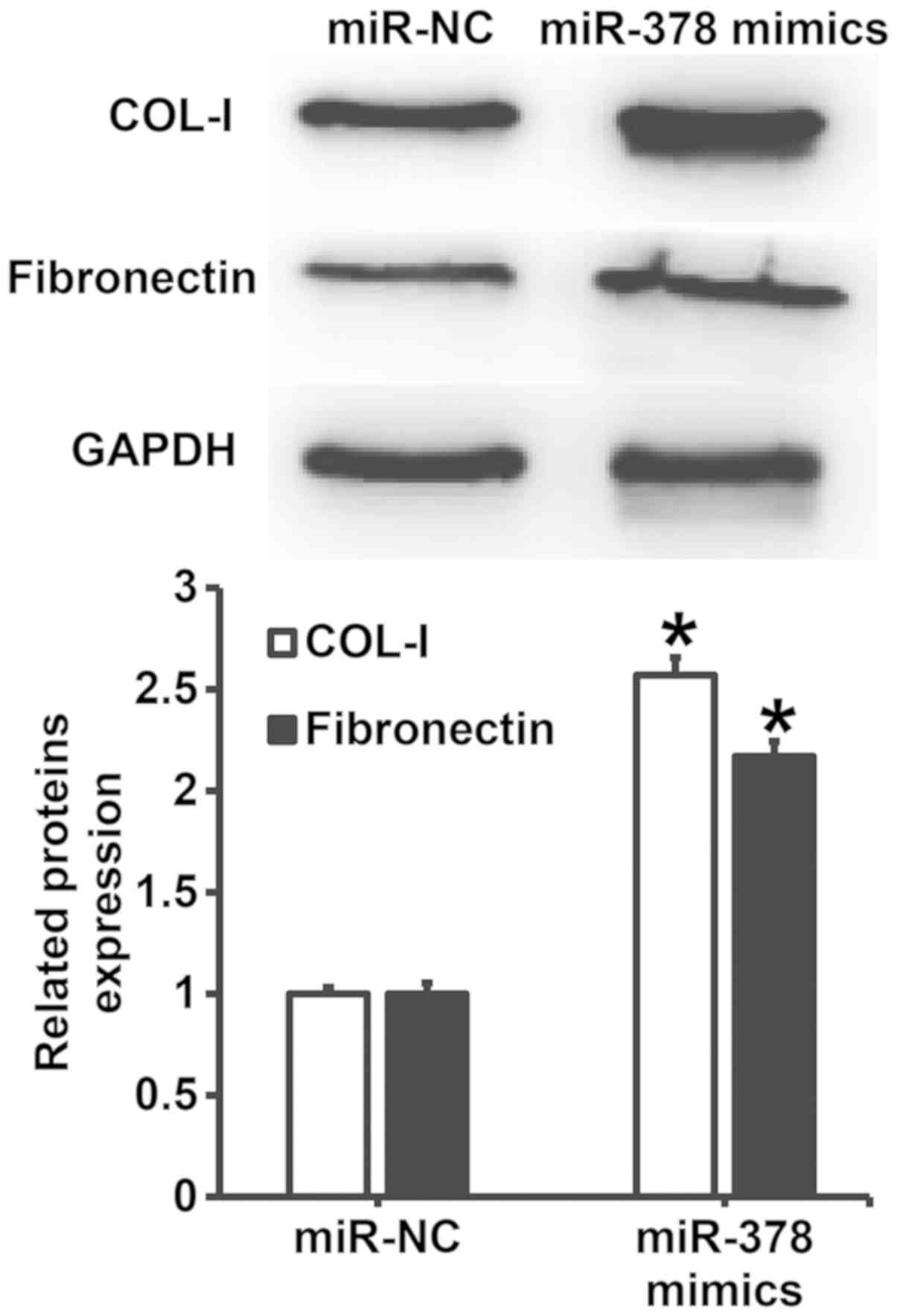
Overexpression of miR-378 increases
the expression of extracellular matrix-related proteins in smooth
muscle cells
To understand the effect of miR-378 overexpression
on the secretion of extracellular matrix-related proteins by smooth
muscle cells, western blotting was carried out. The data showed
that the expression of COL-I and FN proteins in smooth muscle cells
transfected with miR-378 was significantly higher than that in
miR-NC group (P<0.05) (Fig. 6).
The result indicates that overexpression of miR-378 increases the
expression of extracellular matrix-related proteins in smooth
muscle cells.
Downregulation of miR-378 expression
reverses the promoting effect of asthma serum on the proliferation
and apoptosis resistance of smooth muscle cells
To examine whether miR-378 was the key factor by
which asthma serum promoted the proliferation of smooth muscle
cells, we cultured smooth muscle cells transfected with miR-NC or
miR-378 inhibitor together with serum from asthmatic children for 7
days. CCK-8 assay data showed that the absorbance of smooth muscle
cells transfected with miR-378 inhibitor and treated with serum
from asthmatic children at 48 and 72 h was significantly lower than
that of cells transfected with miR-NC and treated with asthma serum
(P<0.05), reaching a level similar to that of miR-NC group
(Fig. 7A). In addition, the
percentage of S-phase cells among all smooth muscle cells
transfected with miR-378 inhibitor and treated with asthma serum
was significantly lower than that in cells transfected with miR-NC
and treated with asthma serum (P<0.05), reaching a level similar
to that of miR-NC group (Fig. 7B).
Apoptosis analysis showed that the apoptotic rate of smooth muscle
cells transfected with miR-378 inhibitor and treated with asthma
serum was significantly higher than that of cells transfected with
miR-NC and treated with asthma serum (P<0.05), reaching a level
similar to that of miR-NC group (Fig.
7C). These results suggest that downregulation of miR-378
expression reverses the promoting effect of asthma serum on the
proliferation and apoptosis resistance of smooth muscle cells.
miR-378 may affect the proliferation,
apoptosis and motility of airway smooth muscle cells via its
downstream signaling pathways
To predict the downstream signaling pathways of
miR-378, bioinformatics was used.
KEGG PATHWAY Database analysis showed that miR-378
has 282 potential target genes, and it possibly participates in the
regulation of ErbB signaling pathway, Ras signaling pathway,
calcium signaling pathway, and MAPK signaling pathway via its
target genes (Table I). The result
indicates that miR-378 may affect the proliferation, apoptosis and
motility of airway smooth muscle cells via its downstream signaling
pathways.
 | Table I.Bioinformatics analysis of downstream
signaling pathways of miR-378. |
Table I.
Bioinformatics analysis of downstream
signaling pathways of miR-378.
| Ranks | Signaling pathways
predicted by KEGG |
|---|
| 1 | ErbB signaling
pathway |
| 2 | Ras signaling
pathway |
| 3 | Calcium signaling
pathway |
| 4 | Pancreatic
secretion |
| 5 | Phosphatidylinositol
signaling system |
| 6 | Estrogen signaling
pathway |
| 7 | MAPK signaling
pathway |
Discussion
In recent years, the incidence of asthma is
increasing year by year in children (25). Asthma is mainly characterized by
airway hyperresponsiveness, inflammation, and remodeling (26). Airway smooth muscle plays important
roles in these three stages, especially airway remodeling (27). miRNA molecules participate in almost
all pathophysiological processes of the body, but their roles and
mechanisms of action in asthma are rarely reported. Airway
inflammation is a key factor in the occurrence and development of
asthma, and airway remodeling is an inevitable result of persistent
airway inflammation (11). Under
repeated stimulation by various inflammatory factors, the repair
mechanism of airway structural cells is disordered, resulting in
pathological changes such as epithelial metaplasia, smooth
hypertrophy, and extracellular matrix precipitation. Furthermore,
airway contractile dysfunction, small airway spasm or stenosis, or
even irreversible constriction and airflow limitation occurs,
finally resulting in decreased pulmonary function in patients
(24). Under the stimulation by
inflammatory factors, smooth muscle cells are transformed from
contractile type to synthetic type, leading to significant
functional changes (28). Therefore,
it is necessary to use smooth muscle cells as target cells for the
treatment of asthma. In the present study, we have isolated and
cultured airway smooth muscle cells in vitro. Spindle-shape
smooth muscle cells are observed under a light microscope, and
immunohistochemistry shows positive expression of α-SMA, suggesting
that the isolated smooth muscle cells are suitable for subsequent
experiments.
miR-378 is a miRNA molecule that was initially found
in tumors. It is reported that miR-378 not only promotes the
occurrence and development of tumors, but also acts as a
tumor-suppressor gene, depending on the type of tumor (29). In addition, miR-378 has also been
found to have biological functions in the development of myocardium
and muscle. For example, miR-378 can regulate the hypertrophy of
cardiomyocytes through Ras signaling pathway (30). Moreover, miR-378 can inhibit the
regeneration of muscle cells (31).
These reports suggest that miR-378 can participate in the
regulation of biological functions of muscle cells. In the present
study, we discover that expression of miR-378 is significantly
increased in both lung tissues and peripheral blood from asthmatic
children. In addition, stimulation of primary smooth muscle cells
by serum from asthmatic children elevates the level of miR-378, and
promotes the proliferation and apoptosis resistance of the cells.
Of note, inhibition of miR-378 expression reduces the promoting
effect of asthma serum on the proliferation and apoptosis
resistance of the cells. Further analysis shows that upregulation
of miR-378 promotes the proliferation and apoptosis resistance of
smooth muscle cells, and elevates the expression of COL-I and FN
proteins. Bioinformatics analysis shows that there are more than
200 target genes of miR-378 that are involved in Ras, MAPK or
calcium signaling pathways. These results suggest that the
biological functions of miR-378 are associated with changes in
these signaling pathways. However, the detailed mechanism still
remains to be elucidated in our future studies. The limitation of
the study is the low sample number, due to difficulties in
collecting samples in asthmatic children and healthy children.
In conclusion, the present study demonstrates that
the expression of miR-378 in children with asthma is elevated,
aggravating airway remodeling by promoting the proliferation and
apoptosis resistance of airway smooth muscle cells. Therefore,
miR-378 could be a potential therapeutic target for asthma in
children. In the future, we will screen miR-378 pathway through
signaling pathway inhibitors or agonists, and then focus on the
target genes in this pathway.
Acknowledgements
The authors wish to thank the Department of
Pediatrics, Maternity and Child Health Care Hospital of Zibo City
and research team for their help and dedication.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
PL and SX collaborated to design the study. PL and
XL were responsible for performing experiments. PL and SX analyzed
the data. All authors collaborated to interpret results and develop
the manuscript. The final version of the manuscript has been read
and approved by all authors.
Ethics approval and consent to
participate
All procedures performed in the present study were
approved by the Ethics Committee of Maternity and Child Health Care
Hospital of Zibo City. Written informed consent was obtained from
all patients or their families.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Kothari PH, Qiu W, Croteau-Chonka DC,
Martinez FD, Liu AH, Lemanske RF Jr, Ober C, Krishnan JA, Nicolae
DL, Barnes KC, et al: Role of local CpG DNA methylation in
mediating the 17q21 asthma susceptibility gasdermin B (GSDMB)/ORMDL
sphingolipid biosynthesis regulator 3 (ORMDL3) expression
quantitative trait locus. J Allergy Clin Immunol. 141:2282–2286.e6.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Hyland ME, Lanario JW, Pooler J, Masoli M
and Jones RC: How patient participation was used to develop a
questionnaire that is fit for purpose for assessing quality of life
in severe asthma. Health Qual Life Outcomes. 16:242018. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Yan W: Toward better management for
asthma: From smart inhalers to injections to wearables, researchers
are finding new ways to improve asthma treatment. IEEE Pulse.
9:28–33. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Davies HM: Living with asthma in
19th-century France: The doctor, Armand Trousseau and the patient,
Emile Pereire. J Med Biogr 967772017741763. 2018:
|
|
5
|
Frey SM, Jones MR, Goldstein N, Riekert K,
Fagnano M and Halterman JS: Knowledge of inhaled therapy and
responsibility for asthma management among young teens with
uncontrolled persistent asthma. Acad Pediatr. 18:317–323. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Bian R, Zhang Y, Yang Y, Yin Y, Zhao X,
Chen H and Yuan Y: White matter integrity disruptions correlate
with cognitive impairments in asthma. J Magn Reson Imaging.
21–Jan;2018.(Epub ahead of print). View Article : Google Scholar
|
|
7
|
Zhu D, Zhang C, Shen H and Ying S:
Breaking through restricting bottleneck for better asthma control.
J Transl Int Med. 5:192–193. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Chitamanni P, Chandrasekaran V and
Rajendiran S: Serum total magnesium level and its correlation with
symptom control in children with mild persistent asthma. Indian J
Pediatr. 85:420–425. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ménard S, Jbilou J and Lauzier S: Family
caregivers' reported nonadherence to the controller medication of
asthma in children in Casablanca (Morocco): Extent and associated
factors. J Asthma. 1–11. 2018. View Article : Google Scholar
|
|
10
|
Burgess JK, Ketheson A, Faiz A, Limbert
Rempel KA, Oliver BG, Ward JPT and Halayko AJ: Phenotype and
functional features of human telomerase reverse transcriptase
immortalized human airway smooth muscle cells from asthmatic and
non-asthmatic donors. Sci Rep. 8:8052018. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Panariti A, Baglole CJ, Sanchez V,
Eidelman DH, Hussain S, Olivenstein R, Martin JG and Hamid Q:
Interleukin-17A and vascular remodelling in severe asthma; lack of
evidence for a direct role. Clin Exp Allergy. 48:365–378. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
An G, Zhang X, Wang W, Huang Q, Li Y, Shan
S, Corrigan CJ, Wang W and Ying S: The effects of interleukin-33 on
airways collagen deposition and matrix metalloproteinase expression
in a murine surrogate of asthma. Immunology. 18–Feb;2018.(Epub
ahead of print). View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Chen J, Jiang Y, Zhou J, Liu S, Qin N, Du
J, Jin G, Hu Z, Ma H, Shen H and Dai J: Evaluation of CpG-SNPs in
miRNA promoters and risk of breast cancer. Gene. 651:1–8. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Staedel C, Tran TPA, Giraud J, Darfeuille
F, Di Giorgio A, Tourasse NJ, Salin F, Uriac P and Duca M:
Modulation of oncogenic miRNA biogenesis using functionalized
polyamines. Sci Rep. 8:16672018. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Long YJ, Liu XP, Chen SS, Zong DD, Chen Y
and Chen P: miR-34a is involved in CSE-induced apoptosis of human
pulmonary microvascular endothelial cells by targeting Notch-1
receptor protein. Respir Res. 19:212018. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Maemura T, Fukuyama S, Sugita Y, Lopes
TJS, Nakao T, Noda T and Kawaoka Y: Lung-derived exosomal
miR-483-3p regulates the innate immune response to influenza virus
infection. J Infect Dis. 217:1372–1382. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yang X, He Q, Guo Z, Xiong F, Li Y, Pan Y,
Gao C, Li L and He C: MicroRNA-425 facilitates pathogenic Th17 cell
differentiation by targeting forkhead box O1 (Foxo1) and is
associated with inflammatory bowel disease. Biochem Biophys Res
Commun. 496:352–358. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Lu X, Yin D, Zhou B and Li T: MiR-135a
promotes inflammatory responses of vascular smooth muscle cells
from db/db mice via downregulation of FOXO1. Int Heart J.
59:170–179. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Oglesby IK, Agrawal R, Mall MA, McElvaney
NG and Greene CM: miRNA-221 is elevated in cystic fibrosis airway
epithelial cells and regulates expression of ATF6. Mol Cell
Pediatr. 2:12015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ho CS, Noor SM and Nagoor NH: MiR-378 and
MiR-1827 regulate tumor invasion, migration and angiogenesis in
human lung adenocarcinoma by targeting RBX1 and CRKL, respectively.
J Cancer. 9:331–345. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Li W, Liu Y, Yang W, Han X, Li S, Liu H,
Gerweck LE, Fukumura D, Loeffler JS, Yang BB, et al: MicroRNA-378
enhances radiation response in ectopic and orthotopic implantation
models of glioblastoma. J Neurooncol. 136:63–71. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Proctor CJ and Goljanek-Whysall K: Using
computer simulation models to investigate the most promising
microRNAs to improve muscle regeneration during ageing. Sci Rep.
7:123142017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Mehta AK, Doherty T, Broide D and Croft M:
Tumor necrosis factor family member LIGHT acts with IL-1β and TGF-β
to promote airway remodeling during rhinovirus infection. Allergy.
73:1415–1424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Hsiao YH, Tseng CM, Su KC, Chen WC, Wu MT,
Wu YC, Chang SC, Lee YC, Kou YR and Perng DW: Glycopyrronium
bromide inhibits lung inflammation and small airway remodeling
induced by subchronic cigarette smoke exposure in mice. Respir
Physiol Neurobiol. 249:16–22. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Griffiths LJ, Lyons RA, Bandyopadhyay A,
Tingay KS, Walton S, Cortina-Borja M, Akbari A, Bedford H and
Dezateux C: Childhood asthma prevalence: Cross-sectional record
linkage study comparing parent-reported wheeze with general
practitioner-recorded asthma diagnoses from primary care electronic
health records in Wales. BMJ Open Respir Res. 5:e0002602018.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Flanigan C, Sheikh A, DunnGalvin A, Brew
BK, Almqvist C and Nwaru BI: Prenatal maternal psychosocial stress
and offspring's asthma and allergic disease: A systematic review
and meta-analysis. Clin Exp Allergy. 48:403–414. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Lee YT, Wu CT, Sun HL, Ko JL and Lue KH:
Fungal immunomodulatory protein-fve could modulate airway remodel
through by affect IL17 cytokine. J Microbiol Immunol Infect.
51:598–607. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Huang N, Liu K, Liu J, Gao X, Zeng Z,
Zhang Y and Chen J: Interleukin-37 alleviates airway inflammation
and remodeling in asthma via inhibiting the activation of NF-κB and
STAT3 signalings. Int Immunopharmacol. 55:198–204. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Li S, Yang F, Wang M, Cao W and Yang Z:
miR-378 functions as an onco-miRNA by targeting the
ST7L/Wnt/β-catenin pathway in cervical cancer. Int J Mol Med.
40:1047–1056. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Nagalingam RS, Sundaresan NR, Gupta MP,
Geenen DL, Solaro RJ and Gupta M: A cardiac-enriched microRNA,
miR-378, blocks cardiac hypertrophy by targeting Ras signaling. J
Biol Chem. 292:51232017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zeng P, Han W, Li C, Li H, Zhu D, Zhang Y
and Liu X: miR-378 attenuates muscle regeneration by delaying
satellite cell activation and differentiation in mice. Acta Biochim
Biophys Sin (Shanghai). 48:833–839. 2016. View Article : Google Scholar : PubMed/NCBI
|