Introduction
Intracranial aneurysms, also referred to as brain
aneurysms, are pathological dilatations of the cerebral artery
caused by weakness in the wall of a cerebral artery or vein
(1). Intracranial aneurysms affect
~3.2% of the whole population, and the numbers may increase in the
near future due to the popularization of imaging techniques in the
diagnosis (2,3), which may increase the diagnosis rate of
this disease. The existence of intracranial aneurysms may not
significantly affect a patient's normal life; however, rupture of
an intracranial aneurysm can cause life-threatening subarachnoid
hemorrhages, which leads to high fatality and morbidity rates among
those patients (4). At present, the
treatment and prevention of intracranial aneurysms are still
challenged due to their unknown pathogenesis.
Vascular smooth muscle cells (VSMCs) serve pivotal
roles in maintaining vascular plasticity and the transition of
VSMCs from normal conditions to matrix remodeling, and the
pro-inflammatory phenotype is the primary pathological change in
the development of an intracranial aneurysm (5). Transforming growth factor (TGF)-β
signaling is proved to participate in the regulation of VSMC
proliferation (6), which indicates
the potential involvement of TGF-β signaling in the pathogenesis of
intracranial aneurysm. TGF-β signaling under certain conditions
achieves its biological roles through interactions with different
long non-coding (lnc)RNAs (7), which
serve critical roles in human diseases (8). Recent studies have demonstrated that
hypoxia inducible factor 1α-antisense RNA 1 (HIF1A-AS1) may
participate in the formation of thoracic aortic aneurysms by
regulating the proliferation and apoptosis of VSMCs (9,10). In
the present study, the results suggested that HIF1A-AS1 may
participate in the formation of intracranial aneurysms by
upregulating TGF-β1 and inhibiting VSMC proliferation.
Patients and methods
Subjects and specimens
A total of 110 individuals were recruited to the
current study. Blood specimens of 56 patients with intracranial
aneurysms (30 males and 26 females; age range, 23–69 years old,
with a mean age of 45.1±6.2 years old) were obtained from the
specimen library of Second Affiliated Hospital of Zhejiang
University (Hangzhou, China). These patients were diagnosed and
treated in Second Affiliated Hospital of Zhejiang University
between January 2015 and January 2018. The inclusion criteria were
as follows: Patients with complete medical records and willingness
to be involved. The exclusion criteria were as follows: Patients
complicated with other diseases or those that received any
treatment (such as intravenous injection) prior to blood extraction
were excluded from the present study. Blood samples were maintained
at room temperature for 30 min and underwent centrifugation at
1,000 × g for 20 min at room temperature to collect serum
(supernatant). At the same time, blood specimens of 54 healthy
subjects (29 males and 25 females; age range, 24–67 years old, with
a mean age of 44.7±5.9 years old) who received routine physical
examinations were also included to serve as the control group.
There were significant differences between the groups with regard
to age and sex. The ethic committee of Second Affiliated Hospital
of Zhejiang University approved the present study and all
participants provided their written, informed consent.
RNA extraction and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
TRIzol reagent (Thermo Fisher Scientific., Inc.,
Waltham, MA, USA) was used to extract total RNA. All procedures
were performed in strict accordance with the instructions of the
kits used. cDNA was synthesized through RT using a SuperScript IV
reverse transcriptase kit (Thermo Fisher Scientific, Inc.) with the
following thermal conditions: 5 min at 25°C, 20 min at 52°C and 5
min at 80°C. SYBR Green Real-Time PCR Master Mix (Thermo Fisher
Scientific, Inc.) was used to prepare all reaction systems. The
following primers were used in PCR reactions: Human lncRNA
HIF1A-AS1 forward, 5′-AATGTGTTCCTTGCTCTT-3′ and reverse,
5′-GTATGTCTCAGTTATCTTCCT-3′; and human GAPDH forward,
5′-CCCACTCCTCCACCTTTGAC-3′ and reverse, 5′-ATGAGGTCCACCACCCTGTT-3′;
TGF-β1 mRNA primers were purchased from SinoBiological Inc. (Wayne,
PA, USA; cat. no. HP100717). PCR reaction conditions were as
follows: 1 min at 95°C, followed by 40 cycles of 20 sec at 95°C and
55 sec at 62°C. Data were normalized using the 2−∆∆Cq
method (11).
Cell culture and transfection
Human VSMCs (cat. no. C0075C; Cascade Biologics;
Thermo Fisher Scientific, Inc.) were cultured with Medium 231
containing smooth muscle growth supplements (both Cascade
Biologics; Thermo Fisher Scientific, Inc.) in an incubator (37°C,
5% CO2). An EcoRI-EcoRI fragment containing full
HIF1A-AS1 cDNA was amplified through PCR and was inserted into
pIRSE2 vector (Clontech Laboratories, Inc., Mountainview, CA, USA).
Vectors were first mixed with Lipofectamine 2000 reagent (cat. no.
11668-019, Invitrogen; Thermo Fisher Scientific, Inc.) to construct
vector-reagent complexes. Following this, cells were incubated at
37°C in an atmosphere containing 5% CO2 for 5 h.
Subsequently, cells were immediately washed with fresh culture
medium to avoid cytotoxicity. Cells without transfection were used
as control cells (C). Cells transfected with empty vector were used
as negative control cells (NC). In the analysis of the effects of
HIF1A-AS1 on TGF-β1, cells were treated with exogenous TGF-β1
(Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) at 10 and 20 ng/ml
for 6, 12 and 18 h prior to use. In the analysis of the effects of
HIF1A-AS1 overexpression and TGF-β1 signaling on VSMC
proliferation, cells were treated with TGF-β1 (10 ng/ml) for 12 h
or TGF-β1 inhibitor LY-364947 (LY, cat. no. 616451; Merck KGaA) for
12 h prior to use. Cells were harvested 24 h after transfection for
subsequent experiments.
Cell proliferation assay
Following transfection and confirmation of
overexpression (overexpression rate, <200%), cells were
collected to prepare cell suspensions using a cell density of
4×104 cells per 1 ml. Cell suspension (100 µl) was added
into each well of a 96-well plate. The plate was incubated in an
incubator (37°C, 5% CO2). Subsequently, 10 µl CCK-8
solution (Sigma-Aldrich; Merck KGaA) was added into each well at
24, 48, 72 and 96 h following the beginning of cell culture. Cells
were cultured in an incubator (37°C, 5% CO2) for a
further 4 h. A Fisherbrand accuSkan GO UV/Vis Microplate
Spectrophotometer (Thermo Fisher Scientific, Inc.) was used to
measure the optical density values at 450 nm. Cell proliferation
was normalized to C (100%).
Western blot analysis
Radioimmunoprecipitation assay lysis solution
(Thermo Fisher Scientific, Inc.) was used to extract total protein
from in vitro cultured cells transfected with HIF1A-AS1
expression vectors, control cells and negative control cells,
according to the manufacturer's instructions. Protein
concentrations were measured using a BCA assay. Protein samples
were mixed with 6X loading buffer with a ratio of 1:5. Following
denaturing at 95°C, SDS-PAGE (10% gels) was performed with 30 µg
protein loaded per lane. Proteins were transferred on
polyvinylidene difluoride (PVDF) membranes (Thermo Fisher
Scientific, Inc.) and blocked in 5% skimmed milk at room
temperature for 2 h. Membranes were incubated with rabbit
anti-human primary antibodies of TGF-β1 (1:2,000; cat. no. ab92486)
and GAPDH antibody (1:2,000; cat. no. ab181602; both Abcam,
Cambridge, MA, USA) at 4°C overnight. The following day, PVDF
membranes were incubated with goat anti-rabbit IgG-horseradish
peroxidase secondary antibody (1:1,000; cat. no. MBS435036;
MyBioSource, Inc., San Diego, CA, USA) at room temperature for 2 h.
Pierce ECL Western Blotting Substrate (Thermo Fisher Scientific.,
Inc.) was then dropped onto membranes to develop signals. Signal
normalization was performed using ImageJ 1.48 software (National
Institutes of Health, Bethesda, MD, USA).
Statistical analysis
GraphPad Prism 6 software (GraphPad Software, Inc.,
La Jolla, CA, USA) was used for all statistical analyses. All data
were recorded as the mean ± standard deviation and the unpaired
t-test and one-way analysis of variance followed by the Least
Significant Difference post hoc test were used for comparisons
between two groups and among multiple groups, respectively.
Correlation analyses were performed using Pearson correlation
coefficient. P<0.05 was considered to indicate a statistically
significant difference.
Results
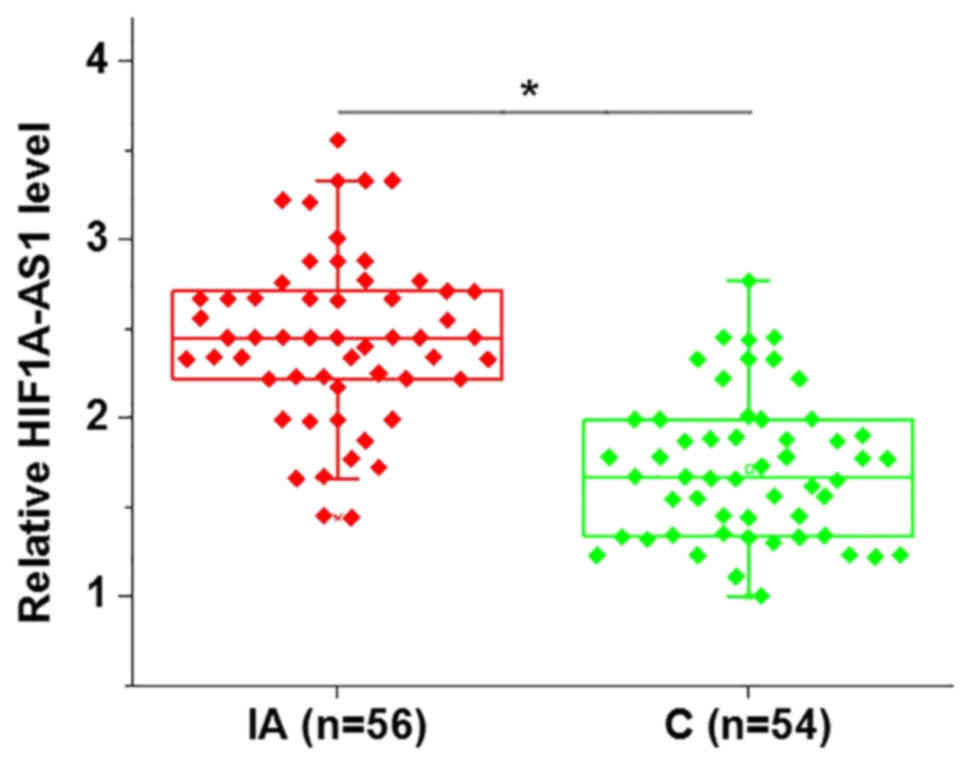
Expression levels of lncRNA HIF1A-AS1
in the blood are upregulated in patients with intracranial
aneurysms
In the present study, the expression levels of
lncRNA HIF1A-AS1 in the blood of patients with intracranial
aneurysms and healthy controls were detected by RT-qPCR. As
indicated in Fig. 1, the expression
levels of HIF1A-AS1 were identified to be significantly increased
in patients with intracranial aneurysms compared with healthy
controls.
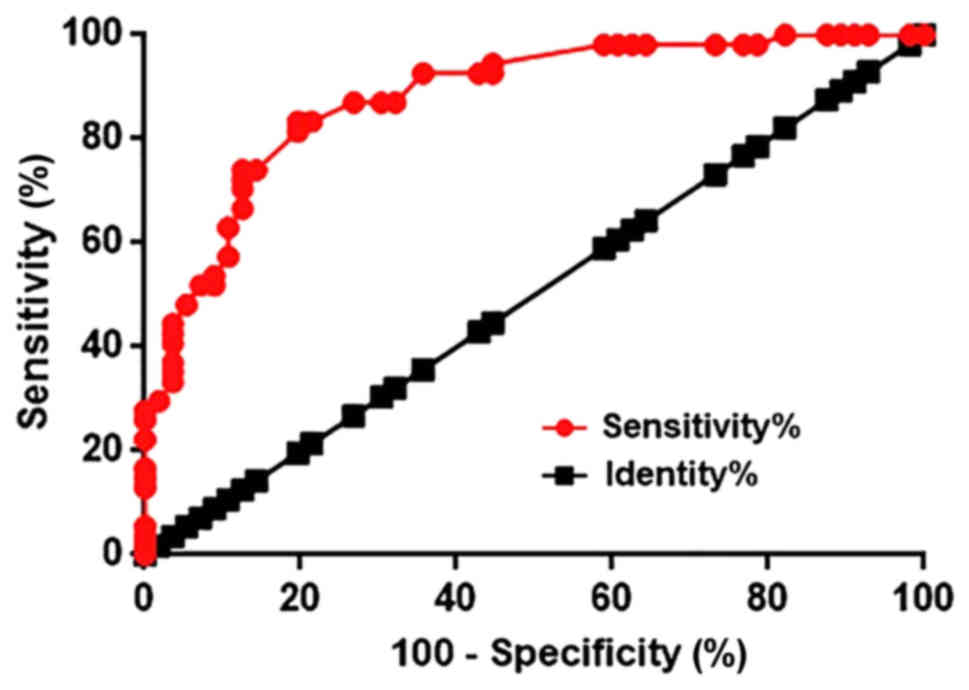
LncRNA HIF1A-AS1 in the blood may
serve as a potential biomarker for intracranial aneurysms
Receiver operating characteristic (ROC) curve
analysis was performed to evaluate the diagnostic value of
HIF1A-AS1 in the blood for intracranial aneurysms. As indicated in
Fig. 2, the area under the curve was
0.8798, with a standard error of 0.03213 and a 95% confident
interval of 0.8168–0.9428. Therefore, upregulation of HIF1A-AS1 in
the blood effectively distinguished patients with intracranial
aneurysms from healthy controls.
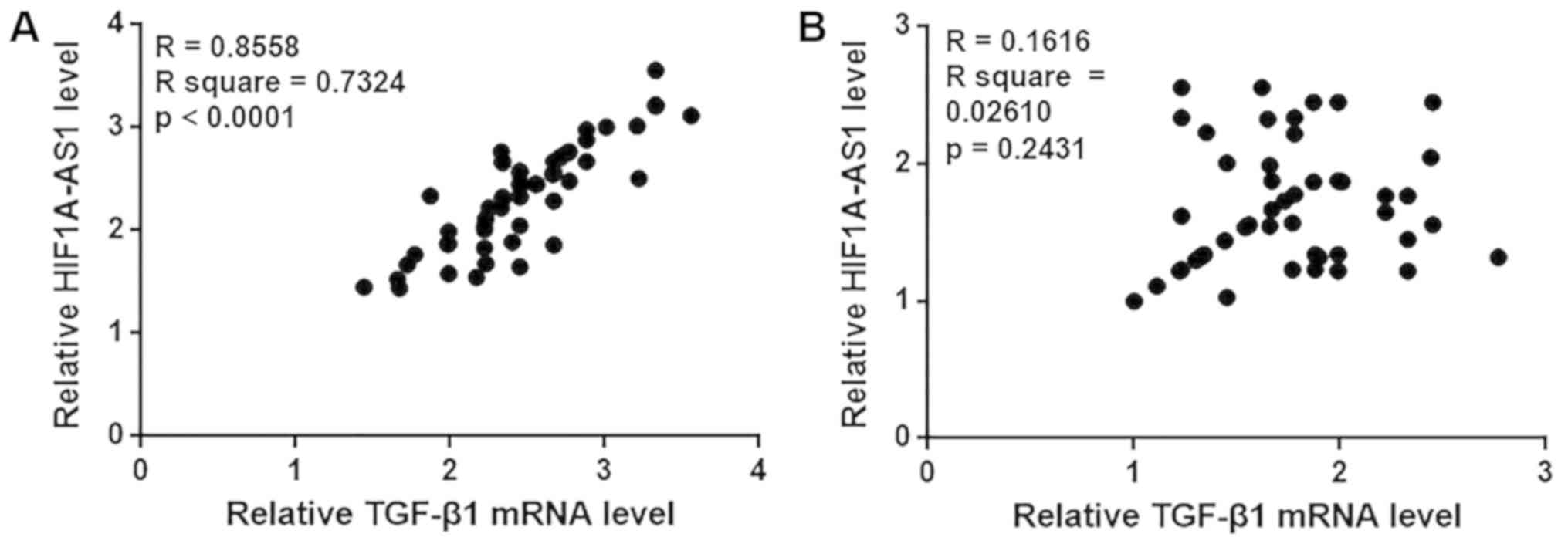
HIF1A-AS1 and TGF-β1 expression levels
are positively correlated in patients with intracranial aneurysms
but not in healthy controls
In the present study, the Pearson correlation
coefficient was used to analyze the correlation between HIF1A-AS1
and TGF-β1 expression in the blood. As demonstrated in Fig. 3A, a significant positive correlation
was identified between HIF1A-AS1 and TGF-β1 expression in the blood
of patients with intracranial aneurysms. By contrast, no
significant correlation between HIF1A-AS1 and TGF-β1 expression was
observed in the blood of healthy controls (Fig. 3B).
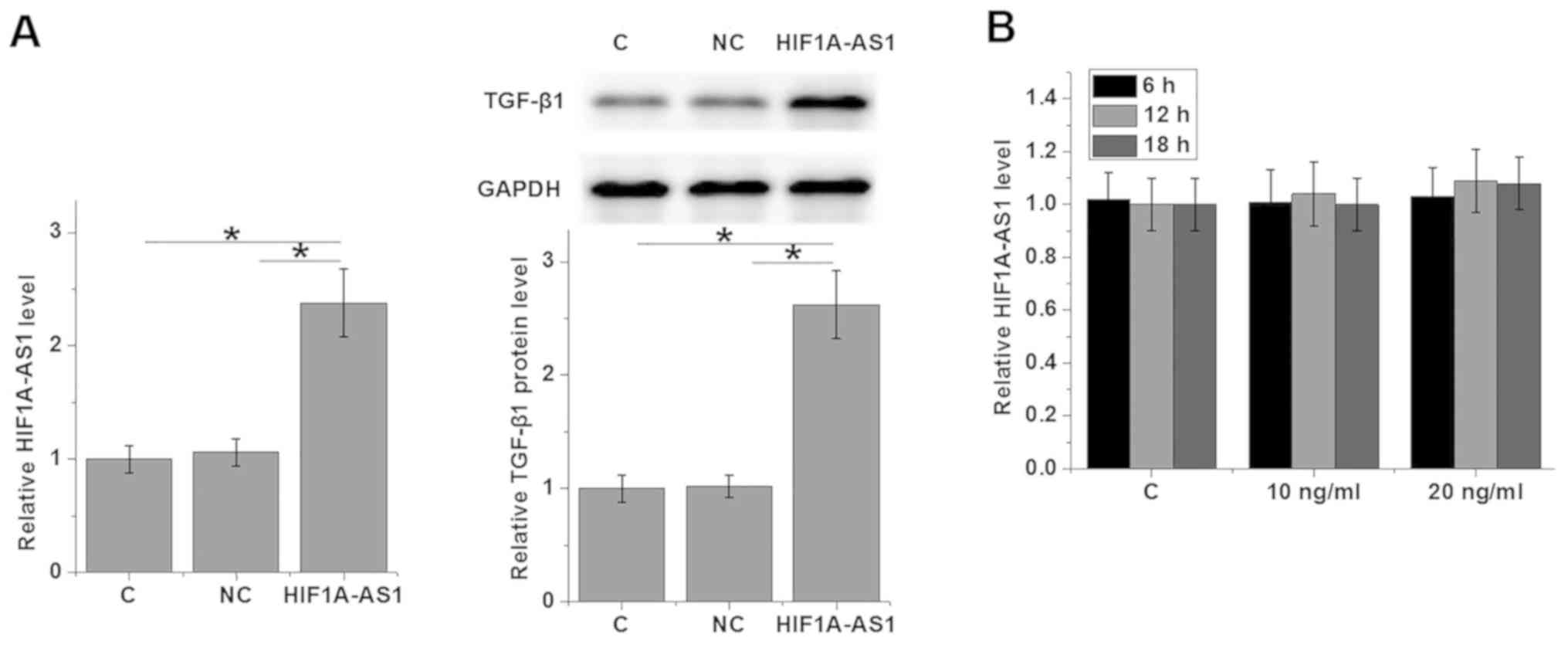
HIF1A-AS1 is likely an upstream
activator of TGF-β1 in human VSMCs
To further explore the interactions between
HIF1A-AS1 and TGF-β1, HIF1A-AS1 expression vector was transfected
in to VSMCs and the expression levels of TGF-β1 were detected by
western blot analysis. As revealed in Fig. 4A, VSMC cells transfected with
HIF1A-AS1 expression vectors (HIF1A-AS1) exhibited significantly
upregulated expression levels of TGF-β1 protein compared with C and
NC. By contrast, cells treated with exogenous TGF-β1 for 6, 12 and
18 h exhibited no significant changes in HIF1A-AS1 expression
(Fig. 4B).
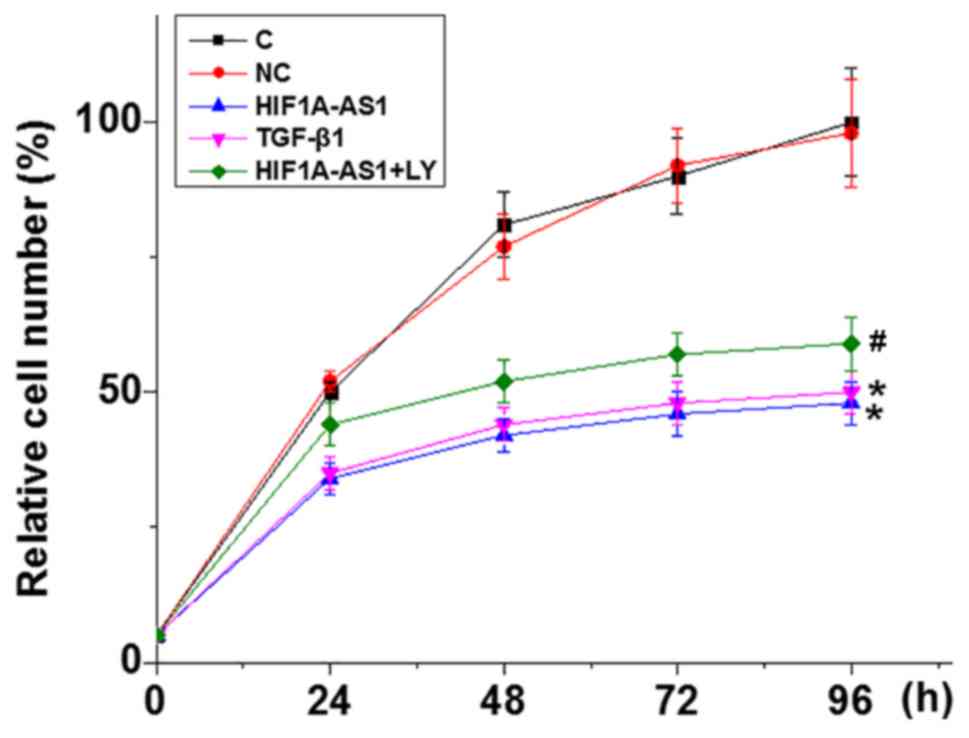
HIF1A-AS1 overexpression inhibits the
proliferation of VSMCs
The CCK-8 assay was performed to detect VSMC
proliferation following HIF1A-AS1 overexpression; the rate of
overexpression was >200% compared with control cells (data not
shown). As indicated in Fig. 5,
compared with C, VSMC proliferation was significantly inhibited
when cells were transfected with HIF1A-AS1 expression vector and 10
ng/ml TGF-β1. In addition, compared with VSMCs treated with
HIF1A-AS expression vector only, cells treated with HIF1A-AS
expression vector transfection and the TGF-β1 inhibitor LY
exhibited significantly increased cell proliferation.
Discussion
The present study reported that HIF1A-AS1 was
involved in in the pathogenesis of intracranial aneurysms. Notably,
the present study indicated that HIF1A-AS1 expression inhibited
VSMC proliferation and upregulated TGF-β1 expression.
Although the pathogenesis of aneurysms is still
unclear, previous research has indicated that the development and
progression of different types of aneurysms are usually accompanied
with changes in expression of a large set of non-coding RNAs
(12). In a recent study, Wang et
al (13) reported 4,129
differentially expressed lncRNAs between intracranial aneurysm
tissues and superficial temporal arteries, and those lncRNAs were
identified to likely be involved in intracranial aneurysms through
their roles in the regulating the immune and inflammatory response.
Upregulation of HIF1A-AS1 has been observed in thoracic aortic
aneurysms (9). In the present study,
significantly increased blood levels of HIF1A-AS1 in patients with
intracranial aneurysms were observed compared with in heathy
controls, indicating that upregulation of HIF1A-AS1 may also
participate in the pathogenesis of intracranial aneurysms.
Currently, the diagnosis of intracranial aneurysms
predominantly relies on imaging techniques, such as computerized
tomography (14) and magnetic
resonance imaging (15). However,
only ~10% of patients with intracranial aneurysms present with
clinical symptoms before rupture (16), and early diagnosis, which is critical
for survival, is not common. The onset, development and progression
of human disease are usually accompanied with changes in certain
blood substances, and monitoring changes in those blood substances
may provide guidance for disease diagnosis (17). In the present study, blood HIF1A-AS1
was detected in all patients with intracranial aneurysms and
healthy controls. ROC curve analysis indicated that blood HIF1A-AS1
can be used to effectively distinguish patients with intracranial
aneurysms from healthy controls. Therefore, blood HIF1A-AS1 may
serve as a potential diagnostic marker for intracranial aneurysms.
However, altered expression of HIF1A-AS1 has been observed in
different diseases, such as thoracic aortic aneurysms (9) and non-small cell lung cancer (18). Therefore, multiple markers should be
used to improve the diagnostic specificity.
The present study observed a significant correlation
between HIF1A-AS1 and TGF-β1 expression in patients with
intracranial aneurysms but not in healthy controls, indicating the
existing of an intracranial aneurysm-specific interaction between
HIF1A-AS1 and TGF-β1. HIF1A-AS1 is likely an upstream activator of
TGF-β1 in VSMCs due to the following reasons: i) HIF1A-AS1
overexpression promoted TGF-β1; ii) exogenous TGF-β1 produced no
significant effects on HIF1A-AS1 expression; and iii) exogenous
TGF-β1 inhibitor reduced the effects of HIF1A-AS1 overexpression in
VSMCs proliferation. It is worth noting that a recent study
reported that the TGF-β signaling pathway promotes VSMCs
proliferation (19), whereas the
present study revealed that TGF-β1 inhibited the proliferation of
VSMCs. The inconsistency between the present data and this previous
study is possibly due to the different VSMCs used in the present
study
In conclusion, the present study suggested that
HIF1A-AS1 is upregulated in patients with intracranial aneurysms.
The findings indicate that HIF1A-AS1 may participate in the
pathogenesis of HIF1A-AS1 by upregulating TGF-β1 and inhibiting
VSMCs proliferation. However, the current study is limited by the
small sample size. Future studies with bigger sample size are
required to further confirm the conclusions of the current
study.
Acknowledgements
Not applicable.
Funding
The present study was funded by the National Natural
Science Foundation of China (grant nos. 81501016 and 81603487), the
Natural Science Foundation of Zhejiang Province (grant no.
LY14H09006) and the Science and Technology Project of Zhejiang
Province (grant no. 2014F81G2010024).
Availability of data and materials
The analyzed data sets generated during the study
are available from the corresponding author on reasonable
request.
Authors' contributions
JX and JG designed experiments. JX, YZ and LC
performed experiments. WC and YD prepared materials and analyzed
the data. JG interpreted data and drafted the experiment. All
authors approved the manuscript.
Ethics approval and consent to
participate
The ethic committee of Zhejiang Chinese Medical
University Hospital approved the present study and all participants
provided their written, informed consent.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Cebral JR, Duan X, Chung BJ, Putman C,
Aziz K and Robertson AM: Wall mechanical properties and
hemodynamics of unruptured intracranial aneurysms. AJNR Am J
Neuroradiol. 36:1695–1703. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Anderson JR, Thompson WL, Alkattan AK,
Diaz O, Klucznik R, Zhang YJ, Britz GW, Grossman RG and Karmonik C:
Three-dimensional printing of anatomically accurate, patient
specific intracranial aneurysm models. J Neurointerv Surg.
8:517–520. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Vlak MH, Algra A, Brandenburg R and Rinkel
GJ: Prevalence of unruptured intracranial aneurysms, with emphasis
on sex, age, comorbidity, country, and time period: A systematic
review and meta-analysis. Lancet Neurol. 10:626–636. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Korja M and Kaprio J: Controversies in
epidemiology of intracranial aneurysms and SAH. Nat Rev Neurol.
12:50–55. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Starke RM, Chalouhi N, Ding D, Raper DM,
Mckisic MS, Owens GK, Hasan DM, Medel R and Dumont AS: Vascular
smooth muscle cells in cerebral aneurysm pathogenesis. Transl
Stroke Res. 5:338–346. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Tsai S, Hollenbeck ST, Ryer EJ, Edlin R,
Yamanouchi D, Kundi R, Wang C, Liu B and Kent KC: TGF-β through
Smad3 signaling stimulates vascular smooth muscle cell
proliferation and neointimal formation. American Journal of
Physiology-Heart and Circulatory Physiology. 297:H540–H549. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Tu X, Zhang Y, Zheng X, Deng J, Li H, Kang
Z, Cao Z, Huang Z, Ding Z, Dong L, et al: TGF-β-induced hepatocyte
lincRNA-p21 contributes to liver fibrosis in mice. Sci Rep.
7:29572017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Shi X, Sun M, Liu H, Yao Y and Song Y:
Long non-coding RNAs: A new frontier in the study of human
diseases. Cancer Lett. 339:159–166. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhao Y, Feng G, Wang Y, Yue Y and Zhao W:
Regulation of apoptosis by long non-coding RNA HIF1A-AS1 in VSMCs:
implications for TAA pathogenesis. Int J Clin Exp Pathol.
7:7643–7652. 2014.PubMed/NCBI
|
|
10
|
Wang S, Zhang X, Yuan Y, Tan M, Zhang L,
Xue X, Yan Y, Han L and Xu Z: BRG1 expression is increased in
thoracic aortic aneurysms and regulates proliferation and apoptosis
of vascular smooth muscle cells through the long non-coding RNA
HIF1A-AS1 in vitro. Eur J Cardiothorac Surg. 47:439–446. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta (CT)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Duggirala A, Delogu F, Angelini TG, Smith
T, Caputo M, Rajakaruna C and Emanueli C: Non coding RNAs in aortic
aneurysmal disease. Front Genet. 6:1252015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wang W, Li H, Yu L, Zhao Z, Wang H, Zhang
D, Zhang Y, Lan Q, Wang J and Zhao J: Aberrant expression of
lncRNAs and mRNAs in patients with intracranial aneurysm.
Oncotarget. 8:2477–2484. 2017.PubMed/NCBI
|
|
14
|
Shintai K, Matsubara N and Izumi T:
High-resolution cone beam CT for evaluation of vascular channel in
intracranial partial thrombosed aneurysm. Nagoya J Med Sci.
80:279–284. 2018.PubMed/NCBI
|
|
15
|
Vakil P, Ansari SA, Cantrell CG, Eddleman
CS, Dehkordi FH, Vranic J, Hurley MC, Batjer HH, Bendok BR and
Carroll TJ: Quantifying intracranial aneurysm wall permeability for
risk assessment using dynamic contrast-enhanced MRI: A pilot study.
AJNR Am J Neuroradiol. 36:953–959. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wiebers DO, Whisnant JP, Huston J III,
Meissner I, Brown RD Jr, Piepgras DG, Forbes GS, Thielen K, Nichols
D, O'Fallon WM, et al: International study of unruptured
intracranial aneurysms investigators. Unruptured intracranial
aneurysms: Natural history, clinical outcome, and risks of surgical
and endovascular treatment. Lancet. 362:103–110. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
De Sanctis P, Elmakky A, Farina A,
Caramelli E, Seracchioli R, Mabrouk M, Mignemi G, Venturoli S,
Villa G, Guerrini M, et al: Matrix metalloproteinase-3 mRNA: A
promising peripheral blood marker for diagnosis of endometriosis.
Gynecol Obstet Invest. 71:118–123. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Tantai J, Hu D, Yang Y and Geng J:
Combined identification of long non-coding RNA XIST and HIF1A-AS1
in serum as an effective screening for non-small cell lung cancer.
Int J Clin Exp Pathol. 8:7887–7895. 2015.PubMed/NCBI
|
|
19
|
DiRenzo DM, Chaudhary MA, Shi X, Franco
SR, Zent J, Wang K, Guo LW and Kent KC: A crosstalk between
TGF-β/Smad3 and Wnt/β-catenin pathways promotes vascular smooth
muscle cell proliferation. Cell Signal. 28:498–505. 2016.
View Article : Google Scholar : PubMed/NCBI
|