Introduction
Ischemic stroke or cerebral ischemia, one of the
most common cerebrovascular diseases, is mainly characterized by
the softening necrosis of brain tissue caused by abnormal blood
circulation, high incidence, sudden onset and easy relapse
(1). According to the reports,
ischemic stroke has become the third leading cause of death and
permanent disability in the United States, with up to 30% mortality
and higher disability rate (2,3). A large
number of studies have indicated that ischemic stroke gives rise to
complex cellular biochemical events, eventually causing apoptosis
of cells and the death of neurons in ischemic regions (4,5). By the
sudden onset of ischemic stroke and rapid development of brain
injury in ischemic region restrictions, it is difficult to
accurately find effective therapies to cure the patients with
cerebral ischemia in a short time (6,7).
Therefore, identifying the biomarkers and quickly diagnosing the
pathogenesis of cerebral ischemia is urgently needed to prevent the
onset of stroke. Recently, a large number of studies have
demonstrated that microRNAs (miRNAs) can be identified as
biomarkers of various diseases, and their expressive abilities play
an important role in clinical applications (8–12). miRNA
as one of the small non-coding RNAs, with ~19–25 nucleotides in
length, has been found to negatively regulate the
post-transcriptional gene expression by inhibiting protein
translation or cutting off the mRNAs of the target genes (13–16). To
date, the number of miRNAs in humans has reached >1,000 species
(17). The target genes of mature
miRNAs are distinguished by the base-pairing interactions between
different nucleotides in the seed and untranslated regions. For a
single miRNA, there are multiple evolutionarily conserved target
genes and several times non-conserved target genes (18). Presumably, ~30% of all genes in human
could be regulated by miRNAs (19).
Although the function of miRNAs on the pathogenesis
of ischemic brain injury has been investigated (20,21), it
is still a challenge to accurately predict the miRNA targets.
Furthermore, the achieved specificity is <50% and poor
consistency is shown among the most advanced algorithms (22). The prediction program of target genes
is mainly based on sequence complementarity, evolutionary
conservation, free energy, and target site accessibility (23–26).
Although evolutionary conservation contributes to enhance the
signal-to-noise ratios, the conservative approach is limited
because of the existence of non-conservation functional target
sites. Especially for the mammalian genomes, the performance of
conservation-based methods is significantly decreased due to the
short evolutionary time (27).
Similarly, the prediction program based on free energy or target
site accessibility has a dependence on the secondary structure
prediction tools, resulting in many shortcomings. The limitations
of all these prediction algorithms indicate a lack of genome-wide
functional data on investigating the effect of miRNA regulation
in vivo. The development of transcriptomic analysis and
proteomic profiling methods has assuaged the requirements for
genome-wide functional data to some extent (28–30).
Especially for mixed prediction methods, the expression profile of
mRNA obtained by microarray sequencing, miRNA and overexpression,
has been proven to be a promising predication method to illuminate
the effect of specific miRNA regulation on genome-wide, and this
approach does not rely on evolutionary conservation (31).
The impact of miRNA regulation on the cerebral
ischemia has been investigated in many studies (32,33).
However, very few reports are available on predicting the target
genes of miRNAs by the screening of differentially expressed (DE)
genes. Simultaneously, TargetScore, as a novel prediction
algorithm, has a high accuracy in estimating known target genes,
and it is used to identify the optimal target genes. In the present
study, the selected miRNAs correlated with cerebral ischemia are
hsa-miR-124, hsa-miR-221 and hsa-miR-223, and were studied by DE
genes and TargetScan analysis (34–38). In
addition, the probabilistic scoring method, named TargetScore, was
adopted to evaluate the consistency of the predicted value with the
true value.
Materials and methods
Overview
A new probabilistic method with high accuracy was
adopted to analyze the miRNA target prediction problem, which was
accomplished by combination of the miRNA-overexpression data and
the sequence-based scores obtained by other prediction methods.
Each score obtained could be considered as an independent
observation variable to be entered into the Variational
Bayesian-Gaussian Mixture Model (VB-GMM). The maximum likelihood
method was chosen to avoid overfitting. In particular, due to the
given expression fold-change resulted from the miRNA transfection,
the downregulated target genes that had few or position fold-change
because of the off-target effects were identified by using a
three-component VB-GMM (39). The
optimization of VB-GMM parameters was performed by using
Variational Bayesian-Expectation Maximization (VB-EM) algorithm.
Ultimately, the mixture component obtained from the largest
absolute methods of the observed negative fold-change or sequence
score was related to miRNA targets, which could be represented as
‘target component’. Any other component was considered as
‘background component’. Therefore, the inference result acquired
from the posterior distribution of the target component for the
observed variables was equivalent to the inference result of
miRNA-mRNA interactions. The values of TargetScore as the
sigmoid-transformed fold-change were calculated by weighting the
average posterior value of target components for all of the
features.
TargetScore
TargetScore can be considered as a comprehensive
probabilistic score of a gene becoming the target of a miRNA.
Briefly, the TargetScore value is defined as follows:
TargetScore=σ(-logFC)((1K+1∑x∈{xf,x1,…,xLp(t|x)))
where σ(-logFC) is calculated using the
formula:
σ(-logFC)=11+exp(logFC)
and p(t|x) is the posterior inferred from the
calculation method of VB-EM. The value of logFC in equations
1 and 2 is an actual value obtained from the experimental database.
The higher the TargetScore value, the more accurate the prediction
results.
miRNA-overexpression data
collection
Gene Expression Omnibus (GEO) database (www.ncbi.nlm.nih.gov/geo/) as a public microarray
data repository, was used to collect the miRNA-overexpression data
in the present study. To date, the GEO is the largest compendium of
miRNA-overexpression data. In order to automatically process data,
a pipeline written with R was developed by using the getGEO
function of the GEOquery R/Bioconductor package (40). The logFC value for treatment
(miRNA transfected) versus control (mock) was calculated in each
dataset. For mRNAs responding with multiple probes in a single
experiment, the average of the logFC values was adopted. The
mRNAs without logFC in both vectors that related to the same
miRNA transfection (investigated in different experiments) were
deleted, and the remaining missing values were filled into one
vector using the non-missing values in the other. For the same
miRNA transfection with >2 logFC, the mRNA deficiency in
all of those vectors was deleted and the remaining missing values
were interpolated making use of knn.impute from R package (41). Eventually, a representative
logFC vector in each miRNA was selected, which possessed the
highest Pearson's correlation with the binary vector of the
verified target genes. Moreover, if there was no valid validation
target, the average of multiple logFCs would be used as a
further method of selection parameters. In the present study, data
with no. GSE22255 were entered, and 40 data were exported. All data
were divided into two groups on average; one was the treatment
group with ischemic stroke, named IS, and the other was the normal
group without ischemic stroke, named control.
DE genes
The screening of DE genes was conducted by the use
of limma that is an R/Bioconductor software package. It was
provided by limma for an integrated solution of analyzing data in
the gene expression experiments. In this study, two groups of
samples, including control and IS group, were processed using the
limma software package. Furthermore, the differential expression of
genes was calculated and clearly displayed, and the expression
genes with large differences were screened out. Using limma
software package, t-test and F-test were performed on the gene
expression matrix that was formed by the differential expression
genes, limFit function was used for the data to linearly fit,
eBayes statistics were carried out and FDR corrected P-values were
calculated (<0.05). The extracted target genes after inspection
in linear model should meet the the absolute value of
logFC≥0.05 and P<0.05.
Results
Obtaining DE genes
Forty samples derived from the GEO database were
divided into ‘control group’ and ‘IS group’. In total, 54,675 genes
were obtained by entering the database no. GSE22255. For the
obtained genes, the t- and F-statistics were verified by the limma
package. In addition, the linear fitting of data and eBayes
statistics were performed using the limFit function. Ultimately,
901 expression genes with large difference were screened out, and
the information of logFC and P-values of all genes were
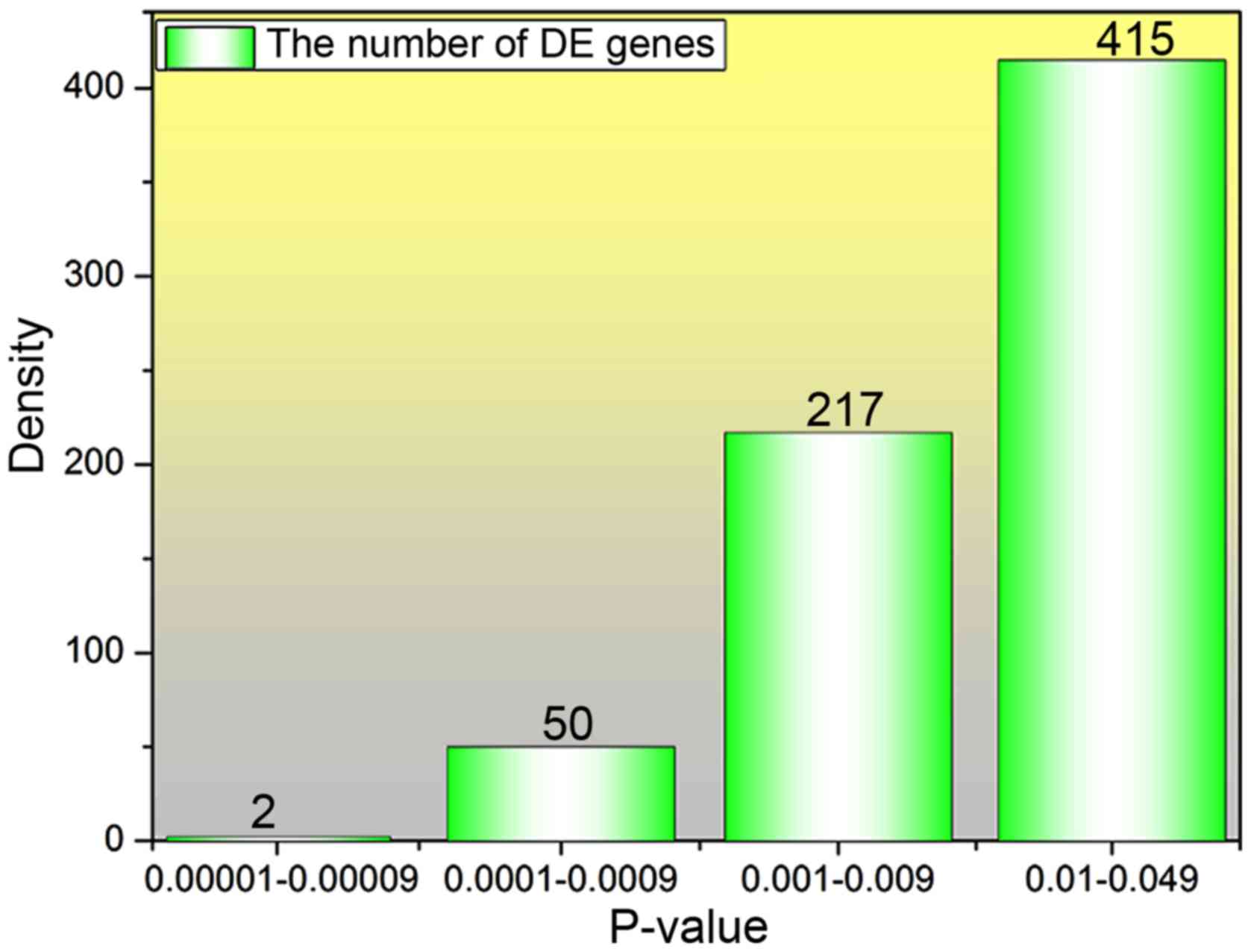
obtained. The density of P<0.05 was calculated (Fig. 1). P-value with a positive correlation
between the number of DE genes with larger difference and the
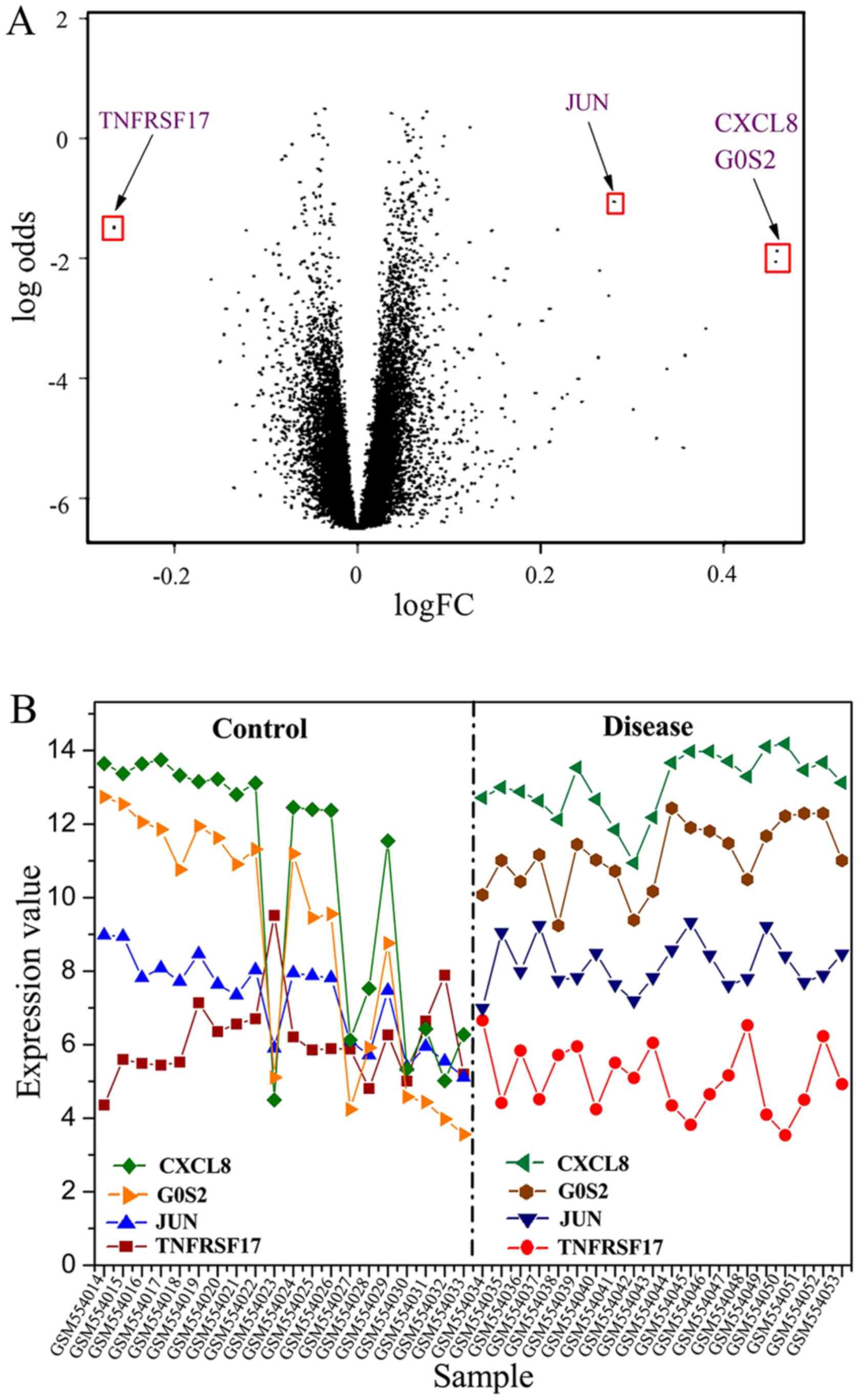
P-value can be seen from the results. Furthermore, a volcanic map
showing the DE results of all genes was plotted as shown in
Fig. 2A. A significant result can be
seen in Fig. 2A; that is, the DE
genes are reduced with the P-value decreased and the absolute value
of logFC increased. Several significant DE genes were found,
and their expression value in different samples was obtained and
plotted in Fig. 2B. A similar trend
for the four DE genes in IS group was presented, and the expression
value for the downregulated gene ‘TNFRSF17’ was found to be lower
than that of other upregulated genes. The expression of these
significant DE genes is shown in Table
I.
 | Table I.Relative values of the expression
levels of several differential genes. |
Table I.
Relative values of the expression
levels of several differential genes.
| DE gene | logFC | Ave Expr | t value | P-value |
|---|
| TNFRSF17 | −0.26587 | 2.458116 | −3.3729 | 0.001625 |
| JUN | 0.280499 | 3.007196 | 3.532038 | 0.00103 |
| CXCL8 | 0.458555 | 3.415384 | 3.221804 | 0.002486 |
| G0S2 | 0.456548 | 3.240999 | 3.150832 | 0.003025 |
Acquisition of TargetScore
miRNAs in human genome are well conserved and play
an important role in post-transcriptional regulation of gene
expression. In this study, the imput miRNAs were hsa-miR-124,
hsa-miR-221 and hsa-miR-223, which contained a set of samples and
expression profiles of 20,514 genes for the experimental data. The
average of all samples was taken to obtain the logFC average
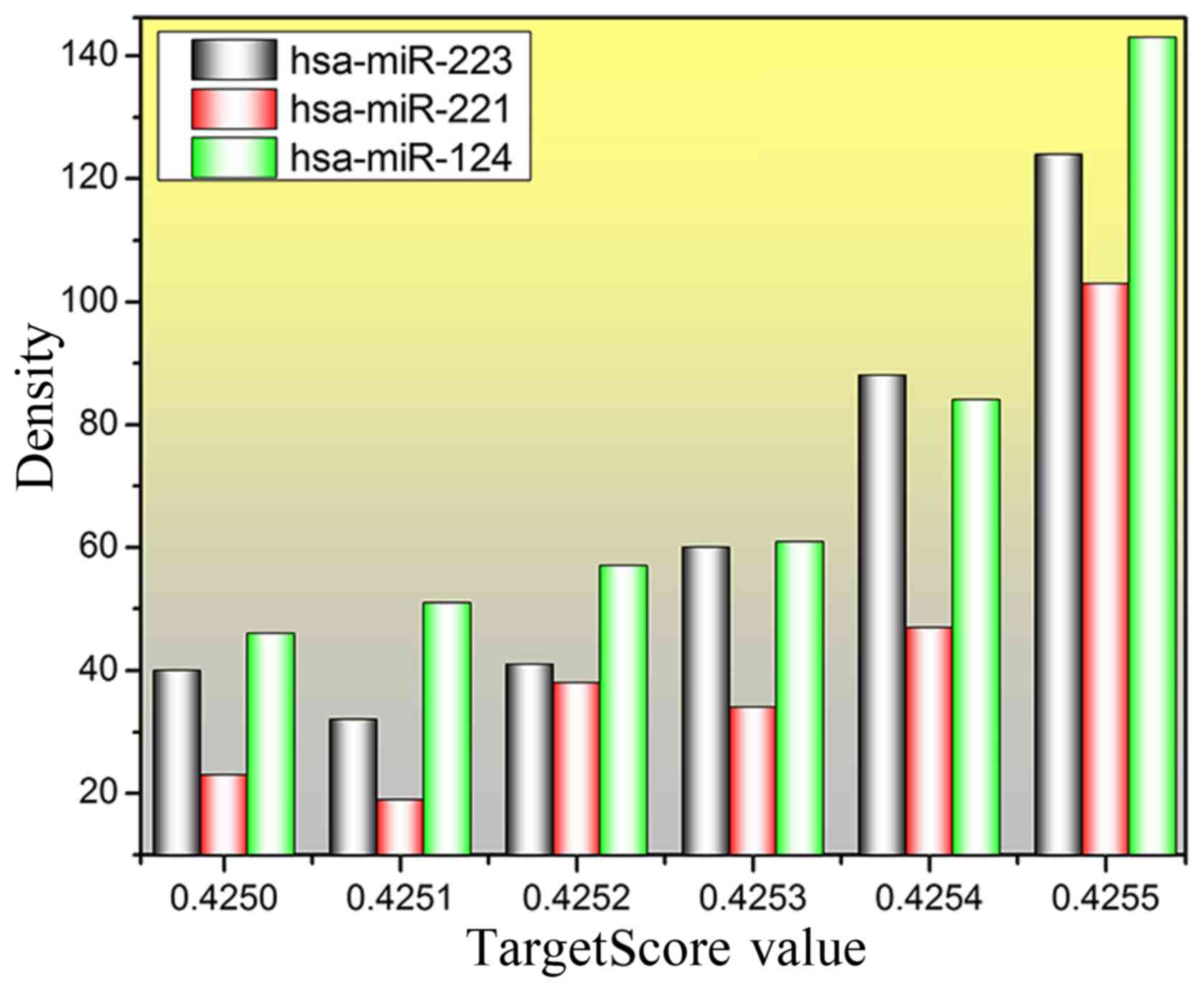
of each gene. Eventually, the TargetScore value of all genes was
constructed by importing the logFC value into equation 1. A
total of 442, 263 and 384 genes with TargetScore value >4.25
were predicted for hsa-miR-124, hsa-miR-221 and hsa-miR-223,
respectively. The distribution of TargetScore value and density for
the three miRNAs was plotted and is shown in Fig. 3. An obvious result can be found in
Fig. 3, that the number of predicted
genes is increasing, especially for the TargetScore value which is
>0.4254, and the number of genes at 0.4255 is >100.
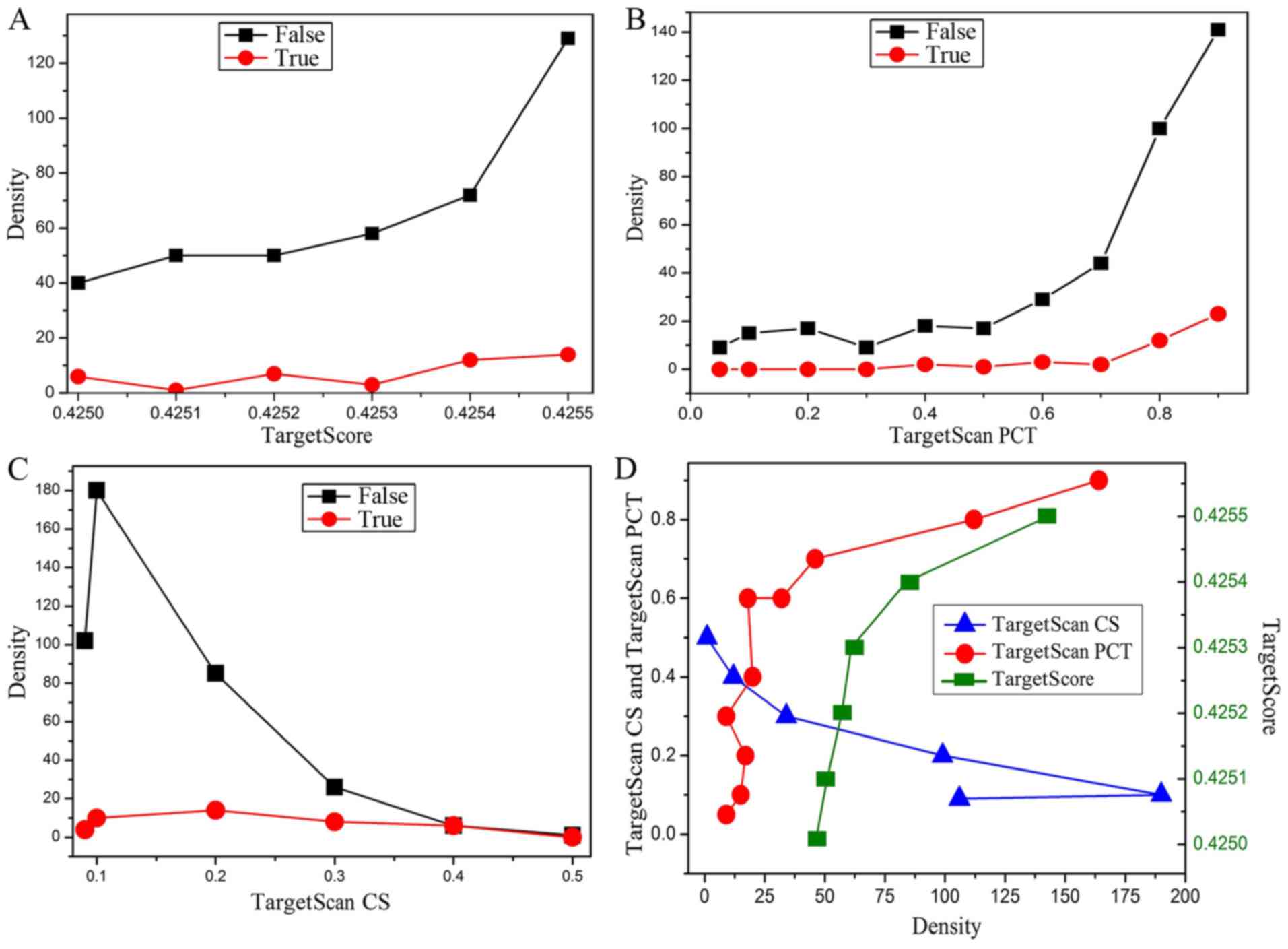
Screening optimal target genes
For all obtained genes, the value of the TargetScan
CS, TargetScan PCT derived from experimental data and TargetScore
value calculated by equation 1 were analyzed to determine the
optimal target genes. Density analysis of the TargetScan CS,
TargetScan PCT and TargetScore for the hsa-miR-124 in different
conditions was carried out, where ‘False’ indicates that the gene
is not reported in the literature, and ‘True’ indicates that the
gene has been reported in the literature (Fig. 4). It is worth mentioning that the
negative values of all data in Fig.
4 are treated as positive values to facilitate the analysis.
According to the density change of the curves in Fig. 4A-C, although the number of genes in
the ‘False’ curve is significantly higher than that of ‘True’
curve, the densities of the ‘False’ and ‘True’ curves have a
similar tendency. Therefore, the optimal target genes could be
identified by integrating the TargetScan CS (<-0.3 cut-off) and
TargetScore (>0.4254 cut-off). Besides, the absolute value of
TargetScan PCT (>0.8 cut-off) as an assistant reference was also
used to screen the target genes. Ultimately, 23 possible target
genes of miR-124 correlated with ischemic stroke were screened out,
and their related detection information is shown in Table II.
| Table II.Predicted target genes of miR-124 by
integrating the novel probability scoring method (TargetScore) and
TargetScan approach. |
Table II.
Predicted target genes of miR-124 by
integrating the novel probability scoring method (TargetScore) and
TargetScan approach.
| Target genes | TargetScan CS | TargetScan PCT | TargetScore |
|---|
| TMEM134 | −0.312 | −0.95 | 0.425554761 |
| ZCCHC24 | −0.43 | −0.96 | 0.425553511 |
| MDC1 | −0.506 | −0.95 | 0.42555199 |
| PTTG1IP | −0.419 | −0.94 | 0.425551244 |
| NEK9 | −0.305 | −0.9 | 0.425550074 |
| ALG2 | −0.376 | −0.93 | 0.425549367 |
| SLC16A1 | −0.436 | −0.96 | 0.425548265 |
| CTSH | −0.361 | −0.29 | 0.425540427 |
| SMARCAD1 | −0.387 | −0.96 | 0.425537318 |
| HEATR1 | −0.386 | −0.88 | 0.425536874 |
| PGRMC2 | −0.336 | −0.96 | 0.425536619 |
| MAGT1 | −0.465 | −0.96 | 0.425534569 |
| NID1 | −0.303 | −0.92 | 0.425533275 |
| RASSF1 | −0.338 | −0.18 | 0.425527887 |
| TARBP1 | −0.432 | −0.85 | 0.425525204 |
| CD164 | −0.393 | −0.96 | 0.425522658 |
| TYK2 | −0.328 | −0.13 | 0.425518604 |
| PQLC3 | −0.336 | −0.85 | 0.425516622 |
| ANXA11 | −0.433 | −0.94 | 0.425510468 |
| MYH9 | −0.402 | −0.96 | 0.425506438 |
| TMEM134 | −0.312 | −0.95 | 0.425554761 |
| ZCCHC24 | −0.43 | −0.96 | 0.425553511 |
| MDC1 | −0.506 | −0.95 | 0.42555199 |
Similarly, possible target genes of miR-223 and
miR-221 could be obtained by detecting the intersection of
TargetScan CS (context score <-0.3) and TargetScore
(>0.4254). However, the value of TargetScan PCT for the miR-221
and miR-223 could not be considered because of the absolute value
of TargetScan PCT is <0.8. Eventually, the number of predicted
target genes for miR-221 and miR-223 is 12 and 9, respectively. The
relevant information is listed in Tables III and IV.
| Table III.Predicted target genes of miR-221 by
integrating the novel probability scoring method (TargetScore) and
TargetScan approach. |
Table III.
Predicted target genes of miR-221 by
integrating the novel probability scoring method (TargetScore) and
TargetScan approach.
| Target genes | TargetScan CS | TargetScan PCT | TargetScore |
|---|
| NDST3 | −0.325 | −0.4 | 0.425553 |
| PHACTR4 | −0.409 | −0.61 | 0.425552 |
| GPBP1 | −0.305 | −0.58 | 0.425552 |
| PYROXD1 | −0.302 | −0.1 | 0.425551 |
| ARF4 | −0.329 | −0.24 | 0.425546 |
| MRPS7 | −0.303 | −0.09 | 0.425539 |
| NDUFA1 | −0.42 | −0.14 | 0.425539 |
| FOXN2 | −0.313 | −0.6 | 0.425536 |
| RFX7 | −0.356 | −0.55 | 0.425536 |
| ZNF652 | −0.395 | −0.61 | 0.425534 |
| CD164 | −0.343 | −0.12 | 0.425523 |
| RNF41 | −0.323 | −0.21 | 0.425516 |
| Table IV.Predicted target genes of miR-223 by
integrating the novel probability scoring method (TargetScore) and
TargetScan approach. |
Table IV.
Predicted target genes of miR-223 by
integrating the novel probability scoring method (TargetScore) and
TargetScan approach.
| Target genes | TargetScan CS | TargetScan PCT | TargetScore |
|---|
| HAUS6 | −0.331 | −0.1 | 0.425554868 |
| C18orf54 | −0.471 | −0.23 | 0.425552879 |
| ZBTB42 | −0.379 | −0.08 | 0.425551922 |
| PKNOX1 | −0.427 | −0.47 | 0.425535322 |
| PARP1 | −0.363 | −0.47 | 0.425526427 |
| LAYN | −0.408 | −0.44 | 0.425526211 |
| FBXW2 | −0.385 | −0.19 | 0.425518779 |
| CYB5A | −0.314 | −0.2 | 0.425517504 |
| SLC39A14 | −0.354 | −0.19 | 0.425510965 |
Discussion
The development of most miRNA target prediction
techniques is transformed into a paradigm that changed from the
rule-based binary classification to the quantitative and
probabilistic approach in a more context dependent manner (42,43).
Under various experimental conditions, the number of miRNAs and
mRNAs expression profiling data is increasing, which to a large
extent promotes the transfer of the trend. By contrast, the large
expression profiles of miRNAs and mRNAs need to span different
tissues, cell lines or patients, and thus have certain limitations
for the application of stable miRNA targets. Furthermore, similar
general principles are adopted to develop the algorithm for most
miRNA target gene prediction methods, namely, hunting for the
targets in the 3′-UTR site of mRNAs and using degree of sequence
conservation to screen the probable miRNA targets (44,45).
However, there are some limitations that need to be overcome for
most algorithms. For example, if there are multiple miRNA targets
at the same location on a transcript, miRanda algorithm can only
detect a miRNA that has the highest scoring and lowest energy,
eventually leading to the appearance of false-negatives (13). In addition, for TargetScan algorithm,
although the false-positive predictions are reduced, the
applicability could be greatly restricted since the prediction is
limited to conservative miRNAs with less than one substitution
among the species (19). To our
knowledge, the earliest prediction method of transfection-based
miRNA target is the Sylamer, which is used to authenticate enriched
k-mer motifs (46). However, Sylamer
can not predict the specific targets since it does not detect the
distribution of fold-changes or sequence features. In this study,
the TargetScore, a Bayesian probabilistic scoring method was
introduced to predict the specific targets, taking into account
both the fold-change caused by miRNA overexpression and the
information based on the sequence. Different from previous target
prediction algorithms based on expression, the TargetScore approach
is an unsupervised algorithm, which means it does not demand
training data. And condition-specific miRNA targets could be
identified by using this novel algorithm. Moreover, TargetScore can
operate the whole genome to more accurately simulate the overall
likehood (47).
Ischemic stroke is an intricate pathology and
physiology process, which is regulated by many factors. It may
cause excitotoxicity in a few minutes, a strong inflammatory
response and apoptosis in a few hours and days from the stroke
onset, leading eventually to an irreversible neuronal damage of
brain tissue affected by the ischemic stroke. There is large number
of literature reports demonstrating that neuronal apoptosis is a
distinctive feature observed after ischemic stroke, and apoptosis
plays a key role in ischemic stroke (4,48).
Lately, miRNAs have been considered to be an important regulatory
factor of neuronal death caused by the ischemic stroke (49). They play an important role in
regulating secondary brain injury and functional outcome after
ischemic stroke, and serve as biomarkers, while opening up a new
field of ischemic disease treatment. However, their is little
research on miRNAs in ischemic stroke, and most studies have mainly
concentrated on profiling changes in the miRNA with ischemic
disease. Currently, the miRNA expression profiling analysis methods
have been used to identify the miRNA change in a rat middle
cerebral artery occlusion (MCAO) model and cerebral ischemia as
well as forebrain ischemia patients (21,50,51). It
is worth mentioning that a single miRNA as a new therapeutic target
can simultaneously control several related target genes, so the
miRNAs can be regarded as promising candidates for cerebral
ischemia therapeutics. It has been reported that the decrease of
miR-181a by intracerebroventricular infusion of its antagomir
significantly reduces the infarction area and defends the penumbra.
Therefore, it indicates that decreasing or blocking miR-181a
contributes in the protection of the brain from ischemic stroke
(52). Additionally, more
investigations have been reported on the effect of miRNAs
regulation on the ischemic stroke. For example, in rat brains after
middle cerebral artery occlusion, miR-29b is upregulated by
apoptosis regulator BCL2L2, while miR-21 selectively downregulates
Faslg (53,54).
In this study, the target genes of miRNAs with
ischemic stroke were investigated by combining different prediction
methods, especially for the TargetScore with higher accuracy in
identifying known targets (55).
miR-124, miR-221 and miR-223 have also been reported in other
literature as potential biomarkers (56). The results of Wang et al
(36) have revealed that miR-223 is
related to the acute cerebral ischemia, and plays an important role
in cerebral ischemia by upregulating growth factors, such as
insulin-like growth factor-1 gene. Chen et al believe that
miR-223 possesses the potential as a biomarker and treatment target
for cerebral ischemia (34). Zhu
et al have suggested that miR-124 is the most abundant miRNA
in brain, and the amount of miR-124 in brain obviously decreases
after ischemic stroke. Besides, as an endogenous regulator of Ku70,
it could be inversely upregulated by Ku70 expression, thereby
relieving brain damage and dysfunction induced by ischemic stroke
(35).
The research results on miRNAs in this study showed
that target genes with ischemic stroke obtained by TargetScore
value are downregulated, and the number of optimal target genes
identified by integrating the priori logFC and posterior
TargetScan CS, TargetScan PCT, and TargetScore is 23, 12 and 9 for
miR-124, miR-221 and miR-223, respectively. It is worth mentioning
that the values of TargetScan CS, TargetScan PCT were derived from
experimental data in the TargetScan site, and eventually introduced
into the equation 1 to get the TargetScore value. A tacit approval
for the TargetScore value has been proposed, that is, the higher
the value of TargetScore, the greater the accuracy of obtained
genes as a target of miRNA regulation. Moreover, miRNAs can control
the expression of proteins by regulating the transcription or
translation of target genes, which has been reported in literature.
The reliability of high accuracy of target genes predicted through
TargetScore values has been confirmed by Li et al (55). The predicted target genes in this
study can be used as a reference and a new method for future
investigation on the treatment and research of ischemic stroke.
In conclusion, new therapeutic strategies may be
discovered by verifying the novel target genes in a disease pathway
with the identification of disease-specific miRNAs. Therefore, the
ability to identify and validate the target genes of miRNAs is
imperative. Although not perfect, the calculation algorithms
combined with Bayesian and Gaussian mixture models and TargetScore
analyses can improve the accuracy of the identification of miRNA
targets. It is worth mentioning that although miRNA has great
potential as a promising candidate to treat ischemic stroke, a lot
of work still needs to be done in ascertaining the interaction
between the individual specific miRNA and target, understanding the
distribution region of miRNAs correlated with ischemic stroke to
promote the therapeutic potential of miRNAs.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
JW performed the experiments, analyzed the data, and
was a major contributor in writing the manuscript. BW and JZ made
substantial contribution to the conception and design of the study.
JW, BW and JZ performed statistical analysis. FJ was also involved
in the conception of the study, the drafting of the manuscript and
gave the final approval for publication. All authors read and
approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Lloyd-Jones D, Adams R, Carnethon M, De
Simone G, Ferguson TB, Flegal K, Ford E, Furie K, Go A, Greenlund
K, et al: American Heart Association Statistics Committee and
Stroke Statistics Subcommittee: Heart disease and stroke statistics
- 2009 update: A report from the American Heart Association
Statistics Committee and Stroke Statistics Subcommittee.
Circulation. 119:e1822009.
|
|
2
|
Kalache A and Aboderin I: Stroke: The
global burden. Health Policy Plan. 10:1–21. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Goldstein LB, Adams R, Becker K, Furberg
CD, Gorelick PB, Hademenos G, Hill M, Howard G, Howard VJ, Jacobs
B, et al: Primary prevention of ischemic stroke: A statement for
healthcare professionals from the Stroke Council of the American
Heart Association. Stroke. 32:280–299. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Love S: Apoptosis and brain ischaemia.
Prog Neuropsychopharmacol Biol Psychiatry. 27:267–282. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Yuan J: Neuroprotective strategies
targeting apoptotic and necrotic cell death for stroke. Apoptosis.
14:469–477. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Stapf C and Mohr JP: Ischemic stroke
therapy. Annu Rev Med. 53:453–475. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Schellinger PD, Kaste M and Hacke W: An
update on thrombolytic therapy for acute stroke. Curr Opin Neurol.
17:69–77. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Krützfeldt J and Stoffel M: MicroRNAs: A
new class of regulatory genes affecting metabolism. Cell Metab.
4:9–12. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Carè A, Catalucci D, Felicetti F, Bonci D,
Addario A, Gallo P, Bang ML, Segnalini P, Gu Y, Dalton ND, et al:
MicroRNA-133 controls cardiac hypertrophy. Nat Med. 13:613–618.
2007. View
Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhang J, Zhao H, Gao Y and Zhang W:
Secretory miRNAs as novel cancer biomarkers. Biochim Biophys Acta.
1826:32–43. 2012.PubMed/NCBI
|
|
11
|
Lionetti M, Musto P, Di Martino MT, Fabris
S, Agnelli L, Todoerti K, Tuana G, Mosca L, Gallo Cantafio ME,
Grieco V, et al: Biological and clinical relevance of miRNA
expression signatures in primary plasma cell leukemia. Clin Cancer
Res. 19:3130–3142. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Khoo SK, Neuman LA, Forsgren L, Petillo D
and Brundin P: Could miRNA expression changes be a reliable
clinical biomarker for Parkinson's disease? Neurodegener Dis Manag.
3:455–465. 2013. View Article : Google Scholar
|
|
13
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Bushati N and Cohen SM: microRNA
functions. Annu Rev Cell Dev Biol. 23:175–205. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Voinnet O: Origin, biogenesis, and
activity of plant microRNAs. Cell. 136:669–687. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Chuck G, Candela H and Hake S: Big impacts
by small RNAs in plant development. Curr Opin Plant Biol. 12:81–86.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Cortez MA, Bueso-Ramos C, Ferdin J,
Lopez-Berestein G, Sood AK and Calin GA: MicroRNAs in body fluids -
the mix of hormones and biomarkers. Nat Rev Clin Oncol. 8:467–477.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Bentwich I, Avniel A, Karov Y, Aharonov R,
Gilad S, Barad O, Barzilai A, Einat P, Einav U, Meiri E, et al:
Identification of hundreds of conserved and nonconserved human
microRNAs. Nat Genet. 37:766–770. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Lewis BP, Burge CB and Bartel DP:
Conserved seed pairing, often flanked by adenosines, indicates that
thousands of human genes are microRNA targets. Cell. 120:15–20.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Jeyaseelan K, Herath WB and Armugam A:
MicroRNAs as therapeutic targets in human diseases. Expert Opin
Ther Targets. 11:1119–1129. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Dharap A, Bowen K, Place R, Li LC and
Vemuganti R: Transient focal ischemia induces extensive temporal
changes in rat cerebral microRNAome. J Cereb Blood Flow Metab.
29:675–687. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Alexiou P, Maragkakis M, Papadopoulos GL,
Reczko M and Hatzigeorgiou AG: Lost in translation: An assessment
and perspective for computational microRNA target identification.
Bioinformatics. 25:3049–3055. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Lewis BP, Shih IH, Jones-Rhoades MW,
Bartel DP and Burge CB: Prediction of mammalian microRNA targets.
Cell. 115:787–798. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Enright AJ, John B, Gaul U, Tuschl T,
Sander C and Marks DS: MicroRNA targets in Drosophila. Genome Biol.
5:R12003. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Krek A, Grün D, Poy MN, Wolf R, Rosenberg
L, Epstein EJ, MacMenamin P, da Piedade I, Gunsalus KC, Stoffel M,
et al: Combinatorial microRNA target predictions. Nat Genet.
37:495–500. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kertesz M, Iovino N, Unnerstall U, Gaul U
and Segal E: The role of site accessibility in microRNA target
recognition. Nat Genet. 39:1278–1284. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Friedman RC, Farh KK, Burge CB and Bartel
DP: Most mammalian mRNAs are conserved targets of microRNAs. Genome
Res. 19:92–105. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Lim LP, Lau NC, Garrett-Engele P, Grimson
A, Schelter JM, Castle J, Bartel DP, Linsley PS and Johnson JM:
Microarray analysis shows that some microRNAs downregulate large
numbers of target mRNAs. Nature. 433:769–773. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Baek D, Villén J, Shin C, Camargo FD, Gygi
SP and Bartel DP: The impact of microRNAs on protein output.
Nature. 455:64–71. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Selbach M, Schwanhäusser B, Thierfelder N,
Fang Z, Khanin R and Rajewsky N: Widespread changes in protein
synthesis induced by microRNAs. Nature. 455:58–63. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Arvey A, Larsson E, Sander C, Leslie CS
and Marks DS: Target mRNA abundance dilutes microRNA and siRNA
activity. Mol Syst Biol. 6:3632010. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Fasanaro P, Greco S, Ivan M, Capogrossi MC
and Martelli F: microRNA: Emerging therapeutic targets in acute
ischemic diseases. Pharmacol Ther. 125:92–104. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Yin KJ, Deng Z, Huang H, Hamblin M, Xie C,
Zhang J and Chen YE: miR-497 regulates neuronal death in mouse
brain after transient focal cerebral ischemia. Neurobiol Dis.
38:17–26. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Chen Y, Song Y, Huang J, Qu M, Zhang Y,
Geng J, Zhang Z, Liu J and Yang GY: Increased circulating exosomal
miRNA-223 is associated with acute ischemic stroke. Front Neurol.
8:572017. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Zhu F, Liu JL, Li JP, Xiao F, Zhang ZX and
Zhang L: MicroRNA-124 (miR-124) regulates Ku70 expression and is
correlated with neuronal death induced by ischemia/reperfusion. J
Mol Neurosci. 52:148–155. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Wang Y, Zhang Y, Huang J, Chen X, Gu X,
Wang Y, Zeng L and Yang GY: Increase of circulating miR-223 and
insulin-like growth factor-1 is associated with the pathogenesis of
acute ischemic stroke in patients. BMC Neurol. 14:772014.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Maitrias P, Metzinger-Le Meuth V, Massy
ZA, M'Baya-Moutoula E, Reix T, Caus T and Metzinger L: MicroRNA
deregulation in symptomatic carotid plaque. J Vasc Surg.
62:1245–1250. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Bazan HA, Hatfield SA, O'Malley CB, Brooks
AJ, Lightell D Jr and Woods TC: Acute loss of miR-221 and miR-222
in the atherosclerotic plaque shoulder accompanies plaque rupture.
Stroke. 46:3285–3287. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Khan AA, Betel D, Miller ML, Sander C,
Leslie CS and Marks DS: Transfection of small RNAs globally
perturbs gene regulation by endogenous microRNAs. Nat Biotechnol.
27:549–555. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Davis S and Meltzer PS: GEOquery: A bridge
between the Gene Expression Omnibus (GEO) and BioConductor.
Bioinformatics. 23:1846–1847. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Troyanskaya O, Cantor M, Sherlock G, Brown
P, Hastie T, Tibshirani R, Botstein D and Altman RB: Missing value
estimation methods for DNA microarrays. Bioinformatics. 17:520–525.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Huang JC, Morris QD and Frey BJ: Bayesian
inference of MicroRNA targets from sequence and expression data. J
Comput Biol. 14:550–563. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Le HS and Bar-Joseph Z: Inferring
interaction networks using the ibp applied to microrna target
prediction. Adv Neural Inf Process Syst. 2011:235–243. 2011.
|
|
44
|
Rajewsky N: microRNA target predictions in
animals. Nat Genet. 38 (Suppl):S8–S13. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Watanabe Y, Tomita M and Kanai A:
Computational methods for microRNA target prediction. Methods
Enzymol. 427:65–86. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
van Dongen S, Abreu-Goodger C and Enright
AJ: Detecting microRNA binding and siRNA off-target effects from
expression data. Nat Methods. 5:1023–1025. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Chen HX, Liu YS and Zhang XJ: TargetScore
used to reveal potential targets of miRNA203 and miRNA-146a in
psoriasis by integrating microRNA overexpression and microarray
data. Medicine (Baltimore). 97:e126712018. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
An YT, Zhu P, Zhong Y, Sheng YC, Zhao Z,
Min Y and Xia YY: A neuroprotective mechanism of YGY-E in cerebral
ischemic injury in rats. CNS Neurosci Ther. 18:14–20. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Yu H, Wu M, Zhao P, Huang Y, Wang W and
Yin W: Neuroprotective effects of viral overexpression of
microRNA-22 in rat and cell models of cerebral ischemia-reperfusion
injury. J Cell Biochem. 116:233–241. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Jeyaseelan K, Lim KY and Armugam A:
MicroRNA expression in the blood and brain of rats subjected to
transient focal ischemia by middle cerebral artery occlusion.
Stroke. 39:959–966. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Yuan Y, Wang JY, Xu LY, Cai R, Chen Z and
Luo BY: MicroRNA expression changes in the hippocampi of rats
subjected to global ischemia. J Clin Neurosci. 17:774–778. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Krützfeldt J, Kuwajima S, Braich R, Rajeev
KG, Pena J, Tuschl T, Manoharan M and Stoffel M: Specificity,
duplex degradation and subcellular localization of antagomirs.
Nucleic Acids Res. 35:2885–2892. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Buller B, Liu X, Wang X, Zhang RL, Zhang
L, Hozeska-Solgot A, Chopp M and Zhang ZG: MicroRNA-21 protects
neurons from ischemic death. FEBS J. 277:4299–4307. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Shi G, Liu Y, Liu T, Yan W, Liu X, Wang Y,
Shi J and Jia L: Upregulated miR-29b promotes neuronal cell death
by inhibiting Bcl2L2 after ischemic brain injury. Exp Brain Res.
216:225–230. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Li Y, Goldenberg A, Wong KC and Zhang Z: A
probabilistic approach to explore human miRNA targetome by
integrating miRNA-overexpression data and sequence information.
Bioinformatics. 30:621–628. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Tsai PC, Liao YC, Wang YS, Lin HF, Lin RT
and Juo SH: Serum microRNA-21 and microRNA-221 as potential
biomarkers for cerebrovascular disease. J Vasc Res. 50:346–354.
2013. View Article : Google Scholar : PubMed/NCBI
|