Introduction
Hepatocellular carcinoma (HCC), which accounts for
~80% of adult primary liver cancer, is the most lethal form of
liver cancer worldwide (1), with the
prevalence and incidence rates rising worldwide annually (2). Early diagnosis of liver cancer is
challenging due to the limited amount of clinical studies and
accurate biomarkers (3). In
addition, the lack of specific symptoms in the early stages further
results in poor prognosis (3).
Nucleolar and spindle-associated protein 1 (NuSAP1)
is an important microtubule and chromatin-binding protein. It has
been demonstrated to serve a role in microtubule crosslink during
mitosis (4–6), regulate the function of kinetochore
microtubules (7), and dominate
chromosome oscillation (8).
Furthermore, NuSAP1 regulates spindle assembly, chromosome
segregation and cytokinesis (4,9).
Previous studies have reported that NuSAP1 is involved in
malignancies, including melanoma (10,11),
breast cancer (12,13), glioblastoma (14), HCC (15), prostate cancer (16), glioblastoma multiforme (17), astrocytoma (18) and meningioma (19). Cui et al (20) revealed that NuSAP1 may act as one of
the core modules in the suppression of HCC by silymarin. In
addition, it has been reported that, during cell division, NuSAP1
acted as a co-factor for the E3 SUMO-protein ligase RanBP2-Ran
GTPase-activating protein 1-SUMO-conjugating enzyme UBC9 complex
and participated in accurate chromosome segregation (21). The above studies indicated that
NuSAP1 serves a role in cancer development. To further understand
the role of NuSAP1 in liver cancer, the current study investigated
the effects of the downregulation of NuSAP1 expression in liver
cancer cell lines. Functional studies confirmed the biological
significance of the NuSAP1 gene in liver cancer.
Materials and methods
Patient and tissue samples
Tumorous and adjacent non-tumorous human liver
specimens were collected from 47 patients who underwent surgery for
HCC at the Nantong Third People's Hospital Affiliated to Nantong
University (Nantong, China) between September 2015 and July 2016.
All patients in this study were diagnosed with primary HCC. No
preoperative radiotherapy or chemotherapy was administered to these
patients. Histopathological diagnosis of all patients with HCC was
completed by the Department of Pathology. The pathological stage of
HCC was determined according to the International Union Against
Cancer Tumor-Node-Metastasis (TNM) Classification (22). The baseline characteristics of the
participants were summarized in a previous study (23). All tissues used in the current study
were frozen immediately following dissection and stored in liquid
nitrogen.
Databases
The expression of NuSAP1 in patients with HCC was
analyzed using the multiple cancer microarray datasets available
from Oncomine (www.oncomine.org) (24). The Cancer Genome Atlas expression
information for NuSAP1 was obtained using UALCAN (www.ualcan.path.uab.edu) (25). A 120-month follow-up study was
performed, using HCC patient information available from the
Kaplan-Meier plotter database (www.kmplot.com) (26),
to verify the association between NuSAP1 expression and prognosis
in patients with HCC. Furthermore, a 80-month follow-up study was
also performed using OncoLnc (www.oncolnc.org) (27).
Cell lines
LO2, HepG2, Huh-7 and SNU-423 cell lines were
obtained from the Cell Bank of Type Culture Collection of Chinese
Academy of Sciences (Shanghai, China). All cells were cultured in
Dulbecco's modified Eagle's medium (DMEM; Thermo Fisher Scientific,
Inc., Waltham, MA, USA) supplemented with 10% fetal bovine serum
(FBS; ScienCell Research Laboratories, Inc., San Diego, CA, USA)
and 1% Penicillin-Streptomycin (Thermo Fisher Scientific, Inc.) and
maintained at 37°C with 50 ml/l CO2.
Lentiviral infection
One day prior to infection, the HepG2 or Huh-7 cell
suspensions were seeded onto six-well plates, at a density of
1×105 cells/well. Two groups included, the siNuSAP1
group, transfected with NuSAP1-siRNA green fluorescent protein
(GFP) lentivirus, and the control group, transfected with a
scrambled siRNA GFP lentivirus. The following day, DMEM with 10%
v/v FBS (complete medium) was removed and replaced with Enhanced
Infection Solution (1 ml/well; Shanghai GeneChem Co., Ltd.,
Shanghai, China), and polybrene (Shanghai GeneChem Co., Ltd.) was
diluted to a final concentration of 5 µg/ml with Enhanced Infection
Solution. For each cell line, different multiplicity of infection
(MOI) values (20, 40, 60, 80, 100 and 120) were tested, and the
final MOI value used in the present study was determined when the
efficiency of infection reached 80% 3 days post-infection. HepG2
cells were infected at a MOI of 120. Huh-7 cells were infected at a
MOI of 60. After swirling the plate gently to mix the cells, the
plate was placed in an incubator with 50 ml/l CO2 at
37°C. After 8 h, the medium was removed and replaced by the
complete medium. Three days post-transfection, GFP expression was
observed in five randomly-selected fields using a fluorescence
microscope (magnification, ×200). Cells with transfection
efficiency >80% on day 3 were selected for subsequent analysis.
Lentiviruses were purchased from Shanghai GeneChem Co., Ltd.
(Shanghai, China). The following target siRNA sequences were used:
Scrambled control, 5′-TTCTCCGAACGTGTCACGT-3′, siNuSAP4-1,
5′-ATAAGCGTTCACTGACCAA-3′, and siNuSAP1 6-1,
5′-CCTTAAAGCTCAAATTCTT-3′.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted using TRIzol®
reagent (Thermo Fisher Scientific, Inc.) from HCC tumor tissues,
adjacent tissues and liver cancer cell lines. The RNA purity and
concentration were measured using a NanoDrop™ 2000
spectrophotometer (Thermo Fisher Scientific, Inc.). The Prime
Script™ RT Reagent kit (Takara Biotechnology Co., Ltd., Dalian,
China) was used for the RT of RNA, according to the manufacturer's
protocol. The reactions were performed using a Bio-Rad Real-Time
PCR Detection System (Bio-Rad Laboratories, Inc., Hercules, CA,
USA). SYBR® Green Master Mix (Vazyme, Piscataway, NJ,
USA) was used for the qPCR. The following primers were used: NuSAP1
forward, 5′-TCATTTCCTTTTCTTGCCTCA-3′ and reverse,
5′-CCCTCAAGTACAGTGACCTGC-3′; glioma-associated oncogene (GLI1)
forward, 5′-AACCCTTGGAAGGTGATATGTC-3′ and reverse,
5′-TTCATACACAGATTCAGGCTCA-3′; DNA methyltransferase (DNMT1)
forward, 5′-CGGCAGACCATCAGGCATTCTAC-3′ and reverse,
5′-CACACCTCACAGACGCCACATC-3′; β-actin forward,
5′-GGACTTCGAGCAAGAGATGG-3′ and reverse, 5′-AGGAAGGAAGGCTGGAAGA-3′.
The following thermocycling conditions were used: Initial
denaturation at 95°C for 5 min; 40 cycles of 10 sec at 95°C and 30
sec at 60°C. Data were analyzed using the 2−ΔΔCq method
(28).
Western blotting
Tissues and cells were lysed in
radioimmunoprecipitation assay buffer (Beyotime Institute of
Biotechnology, Shanghai, China). The supernatants of centrifuged
lysates (14,000 × g for 5 min at 4°C) were diluted in 5X Laemmli
SDS sample buffer (Beyotime Institute of Biotechnology) at a 4:1
ratio. Total protein was quantified using a bicinchoninic acid
assay (Beyotime Institute of Biotechnology) and then boiled at 95°C
for 5 min. Proteins (35 µg/lane) were separated by SDS-PAGE on 10%
gels. The separated proteins were transferred onto nitrocellulose
membranes (Pall Life Sciences, Port Washington, NY, USA), and
incubated with primary antibodies against NuSAP1 (cat. no.
12024-I-AP; 1:2,000) and β-actin (cat. no. HRP-60008; 1:2,000; both
ProteinTech Group, Inc., Chicago, IL, USA) overnight at 4°C.
Following washing with tris-buffered saline and Polysorbate 20, the
membranes were incubated with horseradish peroxidase-conjugated
secondary antibody (cat. no. A00098; 1:2,000; GenScript Co., Ltd.,
Nanjing, China) for 2 h at room temperature. The protein-antibody
complexes were visualized using the SuperSignal West Femto Maximum
Sensitivity substrate (Thermo Fisher Scientific, Inc.) and exposed
to a medical X-ray film (Denville Scientific Inc., Holliston, MA,
USA). Proteins were quantified using the Multi Gauge software
(version 3.0; Fujifilm Dimatix Inc, Santa Clara, CA, USA).
Immunohistochemistry (IHC)
Tissues were fixed in 10% formalin for 12 h at 4°C
and embedded in paraffin. Paraffin-embedded tissue samples
(6-µm-thick) were cut. All sections were heated in roaster at 65°C
for 1 h, and then deparaffinized in dimethylbenzene and rehydrated
in a descending alcohol series at room temperature. Antigen
retrieval was performed by boiling the tissue sections in citrate
buffer (pH 6.0) in a microwave for 30 min. Endogenous peroxidase
activity was quenched by treatment with methanol and 3% hydrogen
peroxide for 15 min at room temperature and background staining was
blocked for 1 h at room temperature with 10% non-immune goat serum
(Thermo Fisher Scientific, Inc.). Tissue sections were incubated
with primary antibody against NuSAP1 (cat. no. 12024-I-AP; 1:200;
ProteinTech Group, Inc.) overnight at 4°C. Following washing with
PBS, the tissue sections were incubated with a horseradish
peroxidase-conjugated secondary antibody (Shanghai Long Island
Biotec Co., Ltd, Shanghai, China) for 30 min at room temperature.
Tissue sections were subsequently washed with PBS and stained with
3,3′-diaminobenzidine (DAB; Maixin-Bio, Guangzhou, China) for 60
sec. Tissue sections were counterstained with Hematoxylin for 15
sec at room temperature for nuclear staining. Sections without the
primary antibody served as negative controls. Histological sections
of fixed tissue samples were observed using a light microscope
(magnification, ×200; Leica Microsystems GmbH, Wetzlar,
Germany).
Cell proliferation assay
A total of 100 µl of cell suspension (3,000
cells/well) were seeded into each well of a 96-well plate. The
plate was subsequently pre-incubated for 12 h in a humidified
incubator at 37°C with 5% CO2. This time point was
defined as day 1. Subsequently, the plate was incubated for 0, 24,
48 or 72 h in the incubator, followed by the addition of 10 µl of
Cell Counting kit-8 (CCK-8) solution (Dojindo Molecular
Technologies, Inc., Kumamoto, Japan) to each well of the plate, and
further incubation for 1.5 h. The absorbance was measured at a
wavelength of 450 nm using a microplate reader (Thermo Fisher
Scientific, Inc.).
Cell cycle analysis
One day prior to transfection, HepG2 or Huh-7 cells
were seeded into six-well plates. Three days post-transfection,
cells were washed with PBS, collected and fixed in 70% ice-cold
ethanol and directly stored in 70% ethanol overnight at −20°C.
Prior to analysis, the cells were incubated in 500 µl sample buffer
[50 µg/ml propidium iodide (Merck KGaA, Darmstadt, Germany) and
0.25 mg/ml of RNase A (Takara Bio, Inc., Otsu, Japan)] for 30 min
at room temperature. The proportion of cells in each phase was
detected using a BD FACSCalibur Flow Cytometer (BD Biosciences, San
Jose, CA, USA) and analyzed using FlowJo software (FlowJo, LLC,
Ashland, OR, USA).
Apoptosis analysis
One day prior to the transfection, HepG2 or Huh-7
cells were seeded into six-well plates. Three days
post-transfection, cells were harvested and washed with PBS. Cell
apoptosis was assessed using the Annexin V-Allophycocyanin
Apoptosis Detection kit I (BD Biosciences). Cells were centrifuged
(200 × g for 5 min at room temperature) and resuspended in 500 µl
of binding buffer to the density of 1×106 cells/ml.
Subsequently, 100-µl cell suspension was incubated with 5 µl
7-aminoactinomycin D and 5 µl Annexin V-fluorescein isothiocyanate
for 15 min at room temperature, in the dark. Apoptotic cells were
detected using a BD FACSCalibur Flow cytometry (BD Biosciences) and
analyzed using FlowJo software.
Transwell invasion assay
Transwell filters were coated with Matrigel (3.9
µg/µl; 60–80 µl) on the upper surface of the polycarbonic membrane
(diameter, 6.5 mm; pore size, 8 µm). The Matrigel solidified
following incubation at 37°C for 6 h. Cells (1×105)
suspended in 200 µl serum-free DMEM were added into the upper
compartment of the chamber. Simultaneously, 600 µl complete medium
was added into the bottom compartment of the chamber. The medium
was removed from the upper chamber following incubation for 48 h at
37°C with 5% CO2. The Matrigel was cleaned with a cotton
swab. The cells migrated from the Matrigel into the pores of the
filter were fixed with 100% methanol, stained with crystal violet
(Beyotime Institute of Biotechnology) for 30 min at room
temperature, washed with PBS and dried at room temperature. The
cells invading through the Matrigel were observed using an inverted
microscope (magnification, ×200). Each assay was repeated three
times.
Statistical analysis
GraphPad Prism was used for data analysis (version
6.0; GraphPad Software, Inc., La Jolla, CA, USA). Student's t-test
and two-way analysis of variance followed by Tukey's post hoc test
were used in the current study. Significance of NuSAP1 expression
in association with clinicopathological parameters was calculated
using the Chi-square test. A paired t-test was used to analyze the
differences of NuSAP1 expression levels between HCC tissues and
corresponding adjacent normal tissues. P<0.05 was considered to
indicate a statistically significant difference. All data are
presented as the mean ± standard error of the mean. All the
experiments were repeated at least three times.
Results
NuSAP1 is overexpressed in HCC tissues
and HepG2 and Huh-7 cell lines
The expression of NuSAP1 in HCC was analyzed using
the multiple cancer microarray datasets available from Oncomine and
it was determined that the mRNA expression levels of NuSAP1 in HCC
tissues were markedly increased compared with those in the normal
tissues (Fig. 1A). The Cancer Genome
Atlas expression information for NuSAP1 was obtained using UALCAN
(Fig. 1B). To improve the
understanding of the clinical relevance of NuSAP1 expression in
HCC, the association between NuSAP1 expression and
clinicopathological parameters was examined (Table I). According to the median value of
relative NuSAP1 mRNA expression in HCC tissues, 47 HCC patients
were subsequently divided into NuSAP1-high (n=24) and NuSAP1-low
groups (n=23). The statistical analysis revealed that a high level
of NuSAP1 expression in HCC was significantly associated with the
TNM stage (P=0.011) and lymph node metastasis (P=0.019). However,
NuSAP1 expression was not associated with the other
clinicopathological features, including age (P=0.77), sex
(P=0.371), serum level of α-fetoprotein (P=0.494), hepatitis B
virus infection (P=0.286) and tumor size (P=0.248). The current
data indicate that elevated NuSAP1 expression may serve a role in
HCC progression, and may be a valuable biomarker for this disease.
To investigate whether the NuSAP1 expression was altered in HCC
tissues, RT-qPCR and western blotting were used to detect the
NuSAP1 levels in 47 paired tumor and adjacent tissues. The mRNA and
protein levels of NuSAP1 in the tumor tissues were significantly
increased compared with the adjacent tissues (Fig. 1C and D). Furthermore, the expression
level of NuSAP1 was detected by IHC in 47 paired tumor and adjacent
tissues (Fig. 1E). The results
revealed that the expression of NuSAP1 was increased in HCC tissues
compared with adjacent tissues. To reveal the potential role of
NuSAP1 in HCC, NuSAP1 expression was examined in four liver cancer
cell lines: HepG2, Huh-7 and SNU-423 as well as the normal human
hepatic cell line LO2 (Fig. 1F). The
expression of NuSAP1 was significantly higher in HepG2 and Huh-7
cells compared with LO2 cells. There were no significant changes in
NuSAP1 expression level in the SNU-423 cell line compared with LO2.
Therefore, the HepG2 and Huh-7 cell lines were selected for future
investigation.
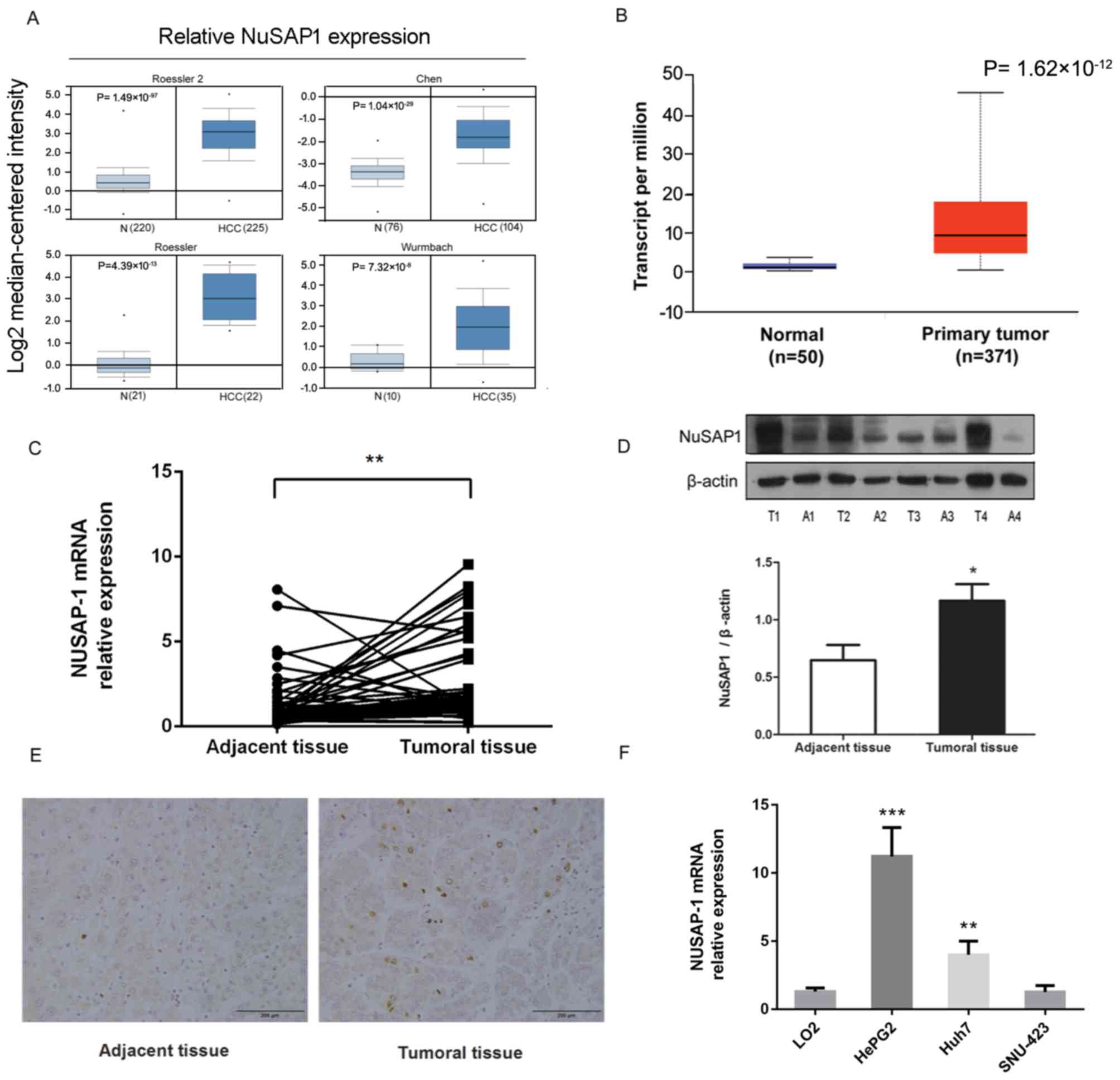 | Figure 1.NuSAP1 is overexpressed in HCC
tissues and HepG2 and Huh-7 cell lines. (A) NuSAP1 is overexpressed
in human HCC tissues, as compared with that in normal tissues, in
multiple cancer microarray datasets available from Oncomine. (B)
NuSAP1 is overexpressed in human HCC tissues compared with normal
tissues, in datasets available from The Cancer Genome Atlas. (C)
mRNA expression levels of NuSAP1 in HCC (n=47) and paired adjacent
tissue samples (n=47) were determined by reverse
transcription-quantitative polymerase chain reaction. (D) Protein
expression levels of NuSAP1 in HCC (n=47) and paired adjacent
tissue samples (n=47) were determined by western blotting. Gray
values of the western blot were quantified. Data were normalized to
β-actin. (E) Expression level of NuSAP1 in HCC and paired adjacent
tissues determined by immunohistochemistry. Scale bar, 200 µm. (F)
Relative NuSAP1 mRNA levels in LO2, HepG2, Huh-7 and SNU-423 cell
lines were determined by reverse transcription-quantitative
polymerase chain reaction. β-actin was used as the internal
control. *P<0.05 vs. with adjacent tissue. **P<0.01 and
***P<0.001 vs. LO2 cell line. HCC, hepatocellular carcinoma;
NuSAP1, nucleolar and spindle-associated protein 1; T, tumor
tissue; A, adjacent tissue. |
 | Table I.Association between NuSAP1 expression
and the clinicopathological characteristics of the 47 patients with
HCC. |
Table I.
Association between NuSAP1 expression
and the clinicopathological characteristics of the 47 patients with
HCC.
|
|
| NuSAP1
expression |
|
|---|
|
|
|
|
|
|---|
|
Characteristics | Total, n | Low, n=23 | High, n=24 | P-value |
|---|
| Age, years |
|
|
|
|
|
≤56 | 28 | 13 | 15 | 0.77 |
|
>56 | 19 | 10 | 9 |
|
| Gender |
|
|
|
|
|
Female | 18 | 7 | 11 | 0.371 |
|
Male | 29 | 16 | 13 |
|
| Serum AFP,
ng/ml |
|
|
|
|
|
≤400 | 36 | 19 | 17 | 0.494 |
|
>400 | 11 | 4 | 7 |
|
| HBV infection |
|
|
|
|
|
Positive | 38 | 17 | 21 | 0.286 |
|
Negative | 9 | 6 | 3 |
|
| TNM stage,
I/II/III |
|
|
|
|
|
I+II | 32 | 20 | 12 | 0.011 |
|
III | 15 | 3 | 12 |
|
| Tumor size, cm |
|
|
|
|
| ≤5 | 24 | 14 | 10 | 0.248 |
|
>5 | 23 | 9 | 14 |
|
| Lymph node
metastasis |
|
|
|
|
|
Positive | 20 | 14 | 6 | 0.019 |
|
Negative | 27 | 9 | 18 |
|
High NuSAP1 expression is associated
with poor prognosis in HCC
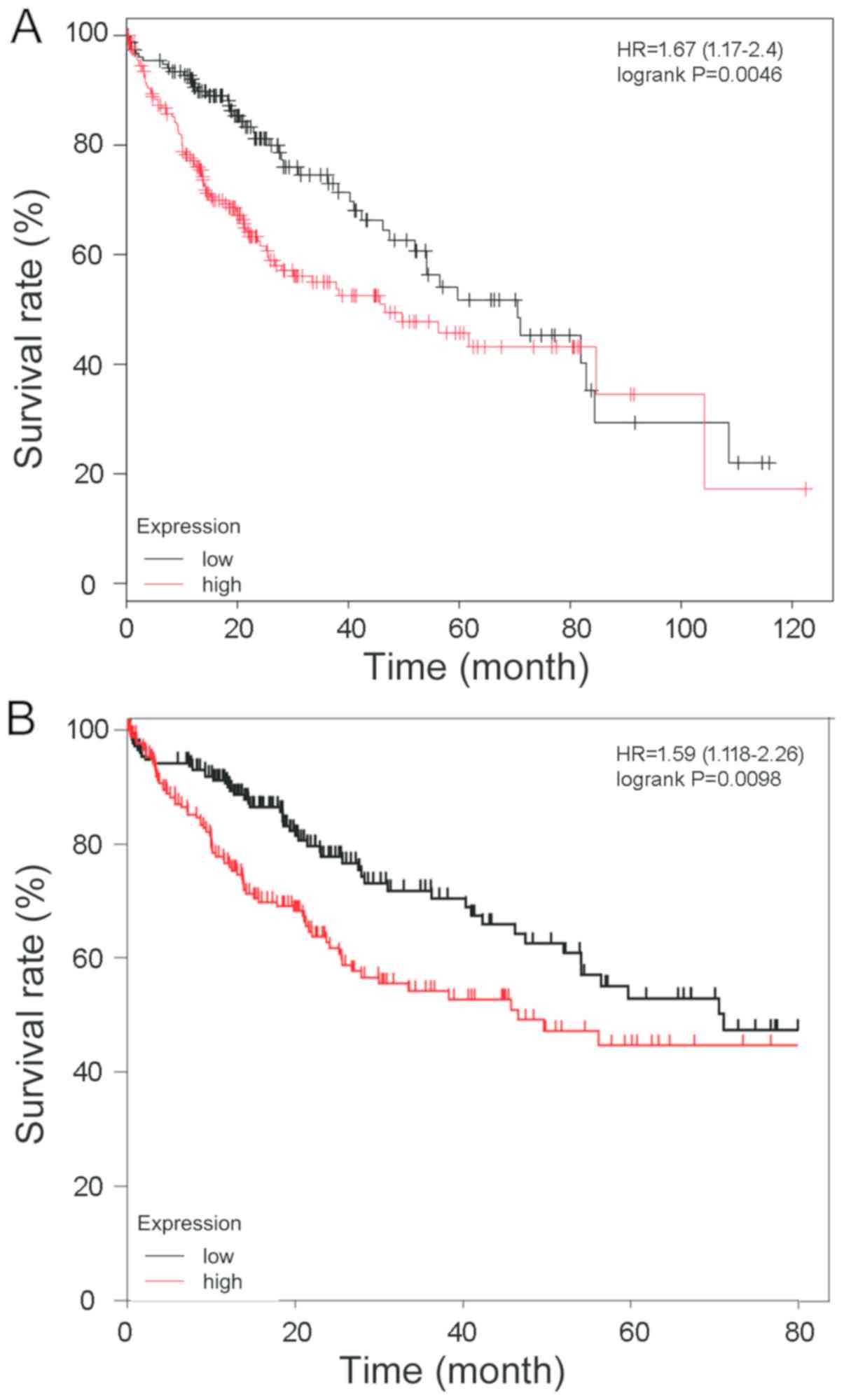
To verify the association between NuSAP1 expression
and prognosis in HCC, a 120-month follow-up study was performed
using HCC patient information available from the Kaplan-Meier
plotter database (Fig. 2A) and the
result was consistent with OncoLnc (Fig.
2B). The association between the NuSAP1 expression level and
prognosis was analyzed. Patients with a higher NuSAP1 expression
level exhibited a markedly shorter overall survival compared with
those with a lower NuSAP1 expression level.
NuSAP1 silencing inhibits the
proliferation and invasion of HepG2 and Huh7 cells
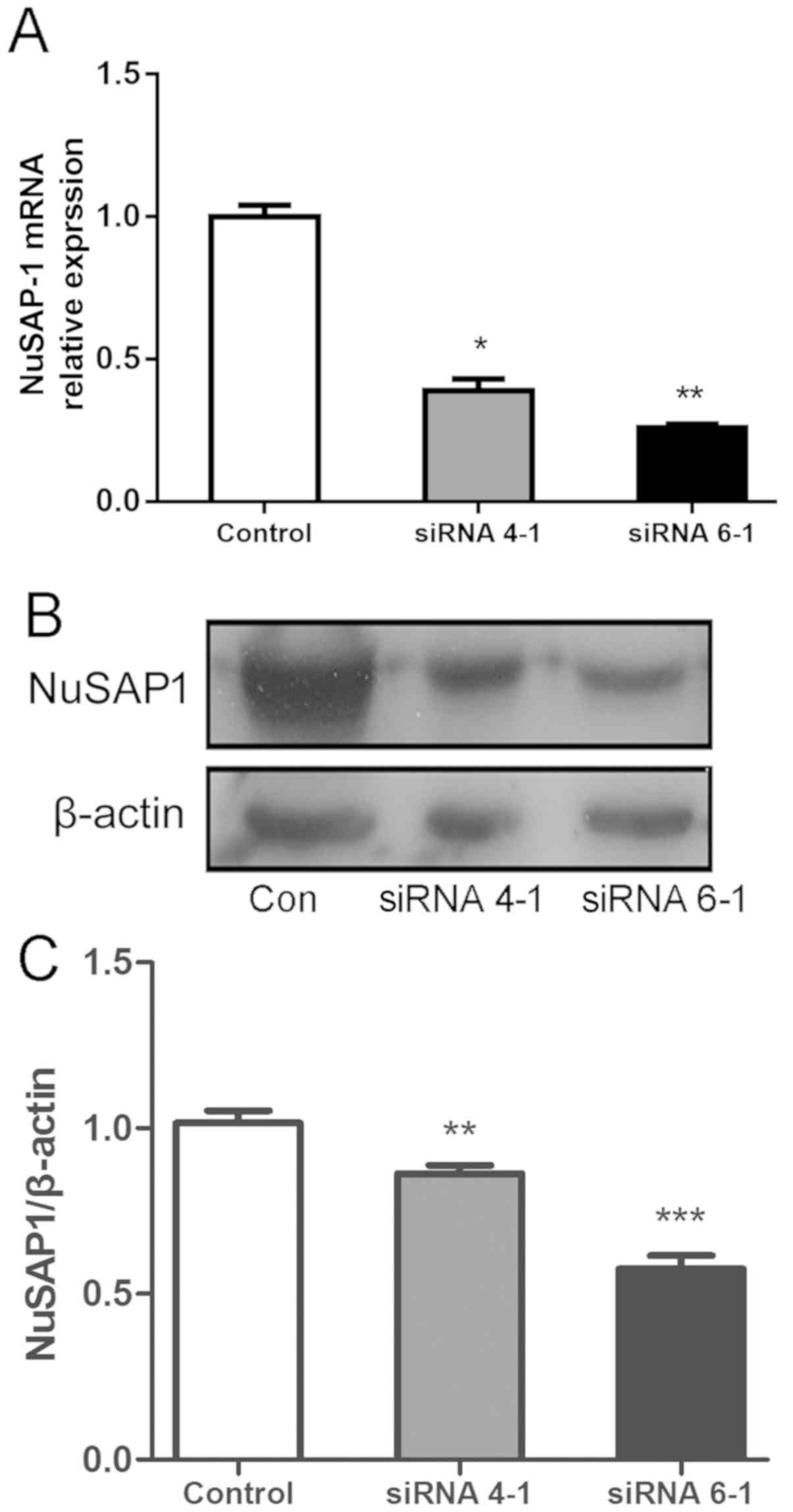
Following siRNA lentiviral transfection, RT-qPCR was
performed to determine the expression levels of NuSAP1 in different
groups. The results revealed that NuSAP1-siRNA (siRNA 4–1 and siRNA
6-1) decreased the expression of endogenous NuSAP1 mRNA in HepG2
cells (Fig. 3A). Simultaneously, the
protein expression of NuSAP1 in NuSAP1-siRNA-treated cells was
suppressed compared with cells transfected with the scrambled
control (Fig. 3B and C). The results
revealed that siRNA 6-1 exhibited a higher efficiency compared with
siRNA 4-1. Therefore, siRNA 6-1 was selected for further
investigation. In order to determine whether NuSAP1 exerted a tumor
facilitating function, the expression of NuSAP1 was suppressed
through a lentiviral vector system using siRNA 6-1 in HepG2 and
Huh-7 cells with high endogenous NuSAP1 levels. The NuSAP1
expression levels were determined by RT-qPCR and western blotting.
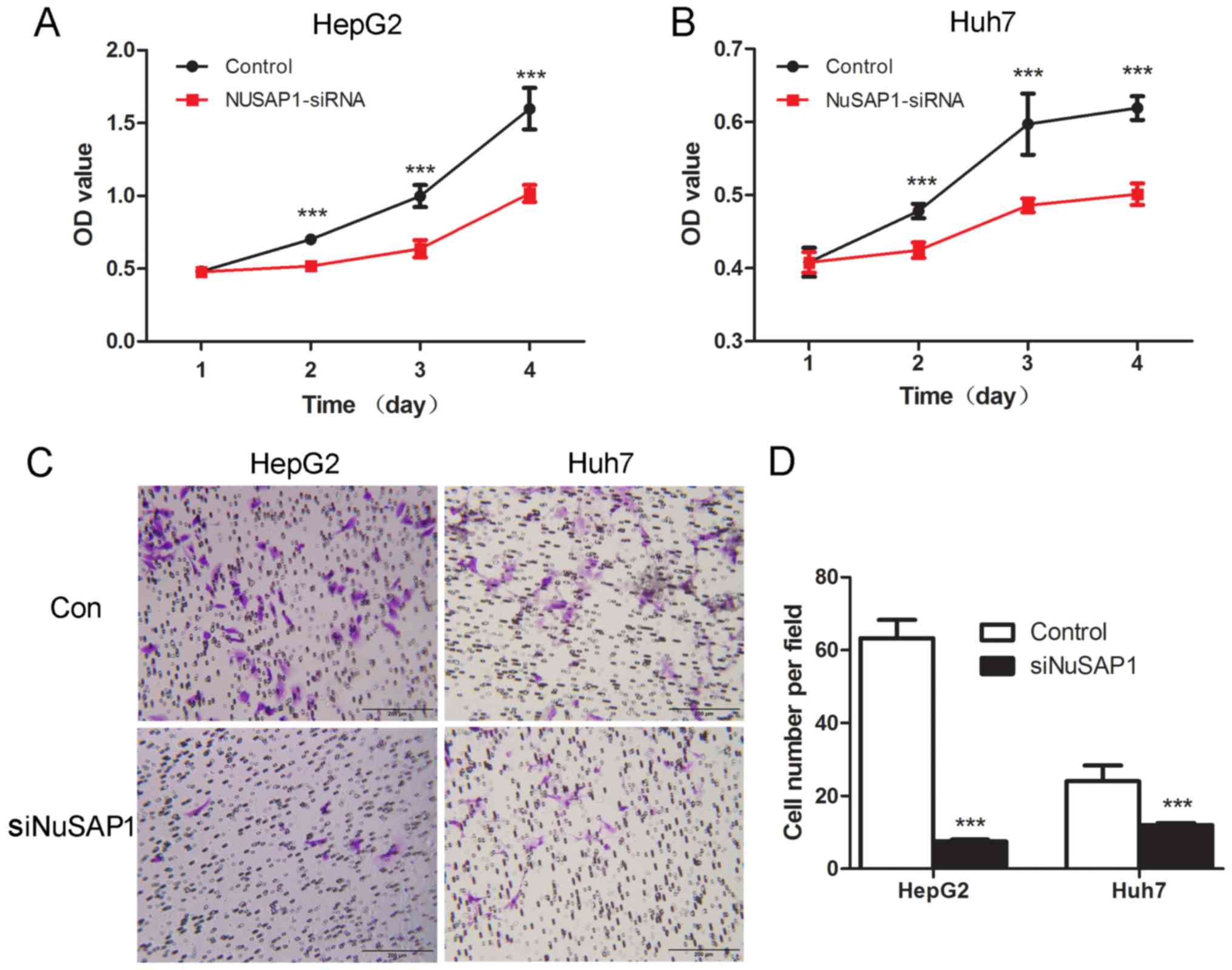
To investigate whether NuSAP1 served a role in cell growth in HepG2
and Huh-7 cells, cell proliferation was detected using the CCK-8,
and invasive potential was determined by the Transwell assay. The
results revealed that the downregulation of NuSAP1 markedly
suppressed cell proliferation (Fig. 4A
and B) and the invasive capability (Fig. 4C and D) of both HepG2 and Huh-7
cells.
NuSAP1 silencing induces cell cycle
progression and cell apoptosis
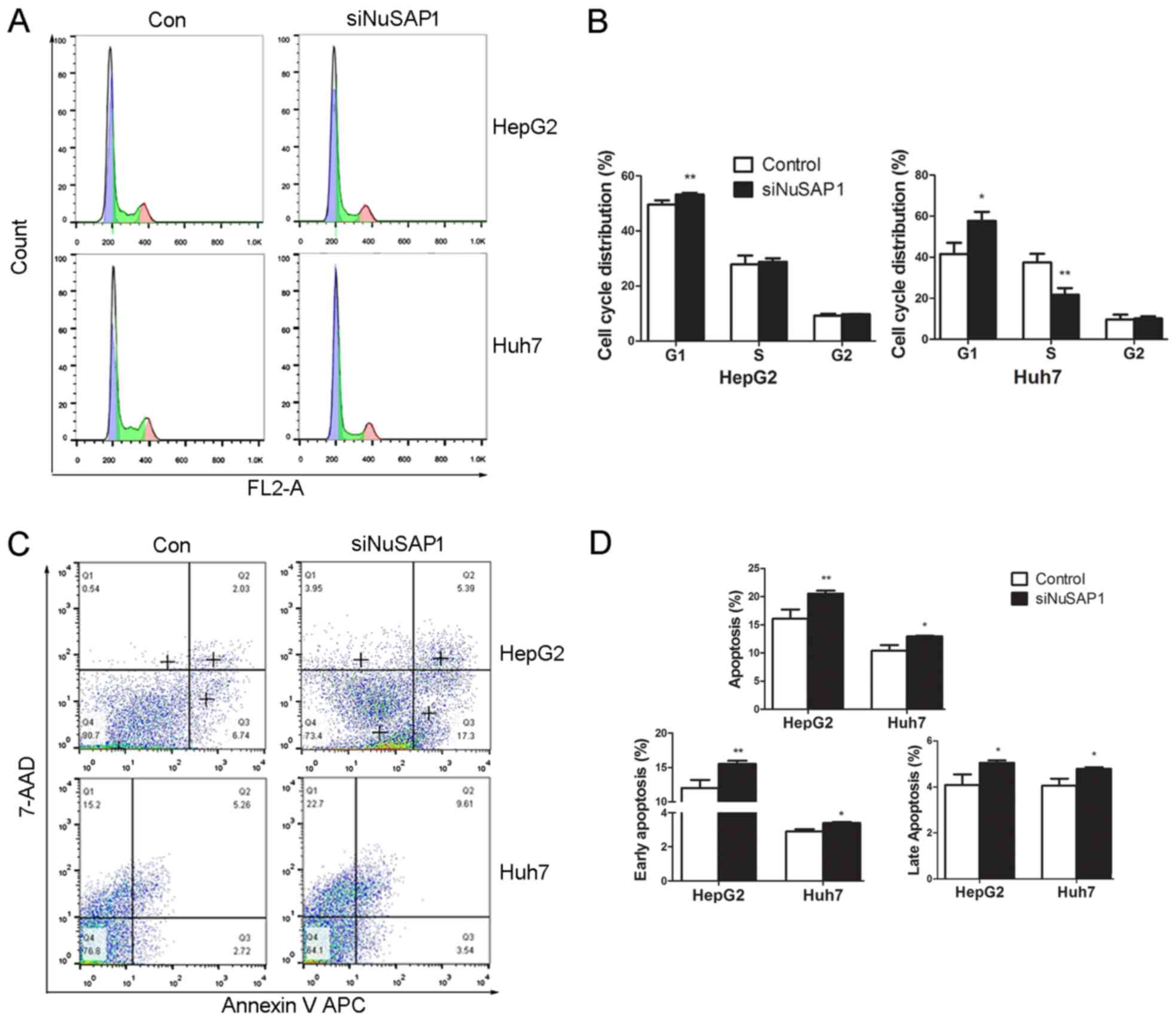
To investigate the growth-suppressing effect of
NuSAP1-siRNA on HepG2 and Huh-7 cells, the cell cycle distribution
was analyzed by flow cytometry. NuSAP1-siRNA significantly
increased the fraction of G1-phase cells in both HepG2 and Huh-7
cells compared with the control group, and decreased the proportion
of S-phase cells in the NuSAP1-siRNA group in Huh7 cells (Fig. 5A and B). The results indicated that
NuSAP1 silencing may induce cell cycle arrest at the G1 phase. The
current study further investigated whether NuSAP1 silencing was
associated with apoptosis in HepG2 and Huh-7 cells. The number of
cells with NuSAP1 gene silencing that exhibited apoptosis was
significantly higher compared with the control group (Fig. 5C and D). Lentiviral transfection
exhibited considerable infection efficiency toward cells, however
it has intrinsic cell toxicity (data not shown). The MOI value used
in the current experiment resulted in the most efficient knockdown
ratio of NuSAP1 at both mRNA and protein levels (data not shown).
Equal MOI was used for the scrambled control group and, therefore,
the conclusion was not affected. The above data suggested that the
silencing of NuSAP1 interrupted cell cycle progression and affected
cell survival.
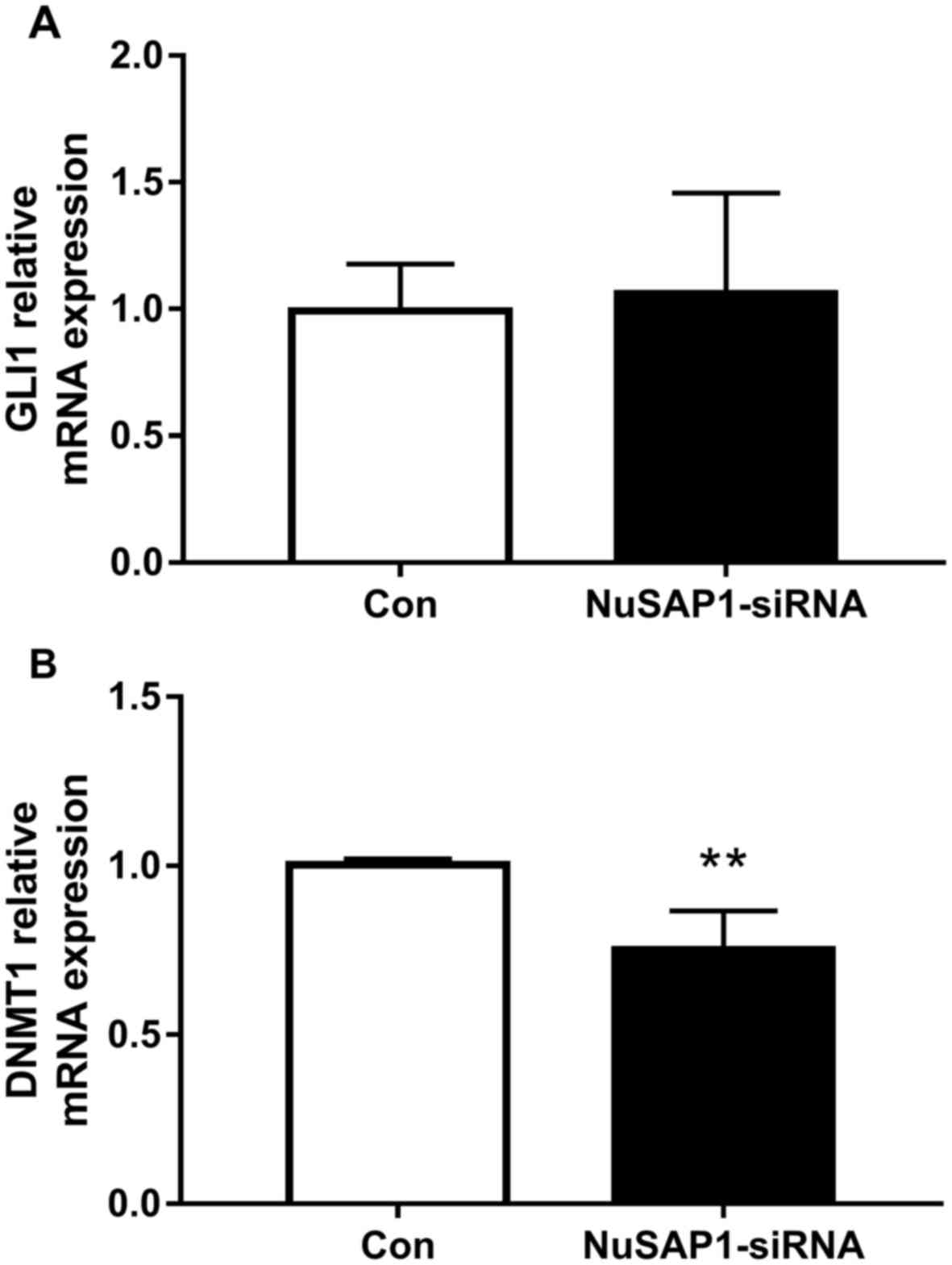
NuSAP1 silencing inhibits the DNMT1
but not the GLI 1 gene expression in liver cancer cells
A prioritized set of 841 enriched GLI1-binding
regions was previously defined by Peterson et al, (29), which included NuSAP1. GLI1 is
involved in the intracellular signal transduction controlled by the
Hedgehog family of secreted molecules (30). The current study further investigated
whether the Hedgehog (HH)/Gli pathway was associated with NuSAP1.
The results revealed no marked alteration in GLI1 mRNA expression
levels following NuSAP1 knockdown in the HepG2 cell line (Fig. 6A), which suggested that NuSAP1 may
not regulate the biological function of liver cancer through the
HH/Gli pathway. NuSAP1 influences the DNA damage response by
controlling breast cancer type 1 susceptibility protein (BRCA1)
expression levels, and global DNA methylation is affected by BRCA1
through the regulation of DNMT1 (31,32).
Thus, the expression of DNMT1 was also detected by RT-qPCR. The
results revealed that NuSAP1 silencing significantly inhibited the
mRNA level of DNMT1 in HepG2 cells compared with the control group
(Fig. 6B).
Discussion
NuSAP1 is a microtubule and chromatin-binding
protein that serves a role in spindle assembly (33). When NuSAP1 is overexpressed,
microtubules aggregate into bundles (4,6). NuSAP1
deficiency in mice causes early embryonic lethality (34). The upregulation of NuSAP1 has been
reported in a number of different human malignancies; however, few
studies have demonstrated the altered expression of NuSAP1 in liver
cancer (10–14,16,19).
Therefore, to clarify the roles of NuSAP1 in liver cancer, the
expression and biological functions of this gene were investigated.
RT-qPCR and western blotting detected NuSAP1 protein expression in
HCC tumor and adjacent tissues. The results revealed that NuSAP1
was upregulated in HCC tumor tissues, which was in accordance with
a previous study (15). This result
indicated that NuSAP1 is closely associated with HCC and may be
used as a biomarker. Subsequently, the expression of NuSAP1 was
determined in different HCC cell lines by RT-qPCR. The results
revealed that NuSAP1 was highly expressed in HepG2 and Huh-7 cell
lines. Although there was no significant alteration in NuSAP1
expression levels in SNU-423, compared with the normal human
hepatic cell line LO2, SNU-423 may be a suitable cell line for
overexpression of NuSAP1 to validate the function of this gene in
future studies.
After years of optimization and improvements,
lentiviral packaging is currently a mature genetic engineering
technology applied in numerous studies (35,36). In
the present study, NuSAP1 gene expression was effectively
suppressed in the HepG2 and Huh-7 cell lines by lentivirus-mediated
siRNA. Proliferation is not only necessary for the development and
tissue homeostasis during adult life, it is also a hallmark of
malignancy (37). To identify genes
associated with the clinical outcome of breast cancer, NuSAP1 was
detected in four independent studies (38). Therefore, in addition to being a
reliable proliferation marker, NuSAP1 is a diagnostic marker for
breast cancer and, potentially, other tumor types (38). The current data revealed that cell
proliferation and invasion were significantly decreased following
silencing of NuSAP1, indicating that its overexpression may enhance
cell activity.
Cell cycle assays were performed to investigate the
mechanism through which NuSAP1 promotes cell proliferation. The
results revealed that the decrease in NuSAP1 expression in liver
cancer cells delayed cell cycle progression by arresting the cycle
at the G1 phase. It may be hypothesized that an upregulated NuSAP1
expression induces a dysregulated cell cycle, which leads to the
abnormal proliferation of tumor cells. Therefore, the accelerated
liver cancer cell proliferation may be inhibited due to the
decreased expression of NuSAP1.
A markedly increased apoptosis in NuSAP1-silenced
liver cancer cells was observed in the current study. The
mechanisms of apoptosis caused by the knockdown of NuSAP1 remain to
be elucidated. The present results indicated that NuSAP1 serves a
role in maintaining liver cancer cell growth. These data strongly
suggest that NuSAP1 may affects tumor cell proliferation and
apoptosis, which contributes to the pathogenesis of liver
cancer.
A previous study revealed that NUSAP1 promotes
invasion, migration and metastasis by modulating family with
sequence similarity 101 member B, a transforming growth factor b1
signaling effector involved in the epithelial to mesenchymal
transition (39). Another study
indicated a strong correlation between E2F1 and NuSAP expression in
human prostate cancer samples (16).
In the present study, to further investigate the underlying
molecular mechanism of NuSAP1 in liver cancer, the GLI1 and DNMT1
expression levels were measured in liver cancer cells using
RT-qPCR. It has been previously reported that NuSAP1 promoted the
aggressiveness of astrocytoma by activating the HH signaling
pathway (18). However, no
significant difference was observed in the GLI1 gene expression
level following NuSAP1 silencing in the current study. A previous
study revealed that NuSAP1 influenced the DNA damage response by
controlling breast cancer type 1 susceptibility protein (BRCA1)
expression levels (31). Global DNA
methylation is affected by BRCA1 through the regulation of DNMT1
(32). Therefore, determining
whether NuSAP1 may regulate the DNMT1 expression would be
noteworthy. The present results indicated that NuSAP1 silencing
inhibited the expression of the DNMT1 gene in liver cancer cells.
However, the underlying mechanism remains unclear and requires
further research. Overall, these results suggested that NuSAP1
silencing may have no effect on the HH signaling pathway. However,
NuSAP1 silencing inhibited the DNMT1 gene expression in liver
cancer cells.
In conclusion, the current study demonstrated that
NuSAP1 upregulation may be an early event in the processes of liver
cancer, therefore NuSAP1 may serve an important role in liver
cancer development. NuSAP1 may provide a new target for liver
cancer gene therapy. Although the underlying mechanisms require
further investigation, the current study provides an improved
understanding of the function of NuSAP1 in liver cancer and lays
the foundation for the development of novel liver cancer treatment
strategies.
Acknowledgements
Not applicable.
Funding
The present study was funded by grants from the
Health Bureau of Nantong City (grant nos. WKZL2018063, WKZL2018057
and WQ2016011), the Nantong Science and Technology Bureau (grant
nos. MS22015105 and GJZ16007), the Natural Science Foundation of
Jiangsu Province (grant no. BK20160420) and the National Science
Foundation for Young Scientists of China (grant no. 81601842).
Availability of data and materials
The datasets and databases used and/or analyzed
during the current study are available from the corresponding
author on reasonable request.
Authors' contributions
ZB and ZL contributed to the study design and
literature search. YW contributed to the western blotting,
immunohistochemistry, RT-qCPR, writing of the manuscript,
statistical analysis and figure creation. FX contributed to the
tissue collection and histopathological analysis. HL and LJ
performed the flow cytometry. XL and LC contributed to the data
collection.
Ethical approval and consent to
participate
The current study was performed in accordance with
the ethical guidelines of the 1975 Declaration of Helsinki. The
current study was reviewed and approved by the Ethics Committee of
the Nantong Third People's Hospital Affiliated to Nantong
University.
Patient consent for publication
Informed consent was obtained from each patient.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Villanueva A and Llovet JM: Liver cancer
in 2013: Mutational landscape of HCC-the end of the beginning. Nat
Rev Clin Oncol. 11:73–74. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
El-Serag HB: Epidemiology of viral
hepatitis and hepatocellular carcinoma. Gastroenterology.
142:1264–1273 e1261. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Tsuchiya N, Sawada Y, Endo I, Saito K,
Uemura Y and Nakatsura T: Biomarkers for the early diagnosis of
hepatocellular carcinoma. World J Gastroenterol. 21:10573–10583.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Raemaekers T, Ribbeck K, Beaudouin J,
Annaert W, Van Camp M, Stockmans I, Smets N, Bouillon R, Ellenberg
J and Carmeliet G: NuSAP, a novel microtubule-associated protein
involved in mitotic spindle organization. J Cell Biol.
162:1017–1029. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Verbakel W, Carmeliet G and Engelborghs Y:
SAP-like domain in nucleolar spindle associated protein mediates
mitotic chromosome loading as well as interphase chromatin
interaction. Biochem Biophys Res Commun. 411:732–737. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ribbeck K, Groen AC, Santarella R,
Bohnsack MT, Raemaekers T, Köcher T, Gentzel M, Görlich D, Wilm M,
Carmeliet G, et al: NuSAP, a mitotic RanGTP target that stabilizes
and cross-links microtubules. Mol Biol Cell. 17:2646–2660. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Li C, Zhang Y, Yang Q, Ye F, Sun SY, Chen
ES and Liou YC: NuSAP modulates the dynamics of kinetochore
microtubules by attenuating MCAK depolymerisation activity. Sci
Rep. 6:187732016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Li C, Xue C, Yang Q, Low BC and Liou YC:
NuSAP governs chromosome oscillation by facilitating the
Kid-generated polar ejection force. Nat Commun. 7:105972016.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Hussain S, Benavente SB, Nascimento E,
Dragoni I, Kurowski A, Gillich A, Humphreys P and Frye M: The
nucleolar RNA methyltransferase Misu (NSun2) is required for
mitotic spindle stability. J Cell Biol. 186:27–40. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ryu B, Kim DS, Deluca AM and Alani RM:
Comprehensive expression profiling of tumor cell lines identifies
molecular signatures of melanoma progression. PLoS One. 2:e5942007.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Bogunovic D, O'Neill DW, Belitskaya-Levy
I, Vacic V, Yu YL, Adams S, Darvishian F, Berman R, Shapiro R,
Pavlick AC, et al: Immune profile and mitotic index of metastatic
melanoma lesions enhance clinical staging in predicting patient
survival. Proc Natl Acad Sci USA. 106:20429–20434. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Chen DT, Nasir A, Culhane A, Venkataramu
C, Fulp W, Rubio R, Wang T, Agrawal D, McCarthy SM, Gruidl M, et
al: Proliferative genes dominate malignancy-risk gene signature in
histologically-normal breast tissue. Breast Cancer Res Treat.
119:335–346. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Chen L, Yang L, Qiao F, Hu X, Li S, Yao L,
Yang XL and Shao ZM: High levels of nucleolar spindle-associated
protein and reduced levels of BRCA1 expression predict poor
prognosis in triple-negative breast cancer. PLoS One.
10:e01405722015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Marie SK, Okamoto OK, Uno M, Hasegawa AP,
Oba-Shinjo SM, Cohen T, Camargo AA, Kosoy A, Carlotti CG Jr, Toledo
S, et al: Maternal embryonic leucine zipper kinase transcript
abundance correlates with malignancy grade in human astrocytomas.
Int J Cancer. 122:807–815. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhang M, Yang D, Liu X, Liu Y, Liang J, He
H, Zhong K, Lin L, Tao G, Zhang C and Zhou J: Expression of Nusap1
in the surgical margins of hepatocellular carcinoma and its
association with early recurrence. Nan Fang Yi Ke Da Xue Xue Bao.
33:937–938, inside back cover, 2013 (In Chinese). PubMed/NCBI
|
|
16
|
Gulzar ZG, McKenney JK and Brooks JD:
Increased expression of NuSAP in recurrent prostate cancer is
mediated by E2F1. Oncogene. 32:70–77. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Xue YP, Ji XY, Yang L, Liu HR, Sheng YJ,
Dai XX, Xi YJ, Liu JC, Shi J, Xie T, et al: Experimental studies on
correlation between nucleolus spindle-related protein 1 and the
malignant progression and prognosis of human glioblastoma
multiforme. Zhonghua Yi Xue Za Zhi. 98:340–345. 2018.(In Chinese;
Abstract available in Chinese from the publisher). PubMed/NCBI
|
|
18
|
Wu X, Xu B, Yang C, Wang W, Zhong D, Zhao
Z, He L, Hu Y, Jiang L, Li J, et al: Nucleolar and spindle
associated protein 1 promotes the aggressiveness of astrocytoma by
activating the Hedgehog signaling pathway. J Exp Clin Cancer Res.
36:1272017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Stuart JE, Lusis EA, Scheck AC, Coons SW,
Lal A, Perry A and Gutmann DH: Identification of gene markers
associated with aggressive meningioma by filtering across multiple
sets of gene expression arrays. J Neuropathol Exp Neurol. 70:1–12.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Cui H, Li TL, Guo HF, Wang JL, Xue P,
Zhang Y, Fan JH, Li ZP and Gao YJ: Silymarin-mediated regulation of
the cell cycle and DNA damage response exerts antitumor activity in
human hepatocellular carcinoma. Oncol Lett. 15:885–892.
2018.PubMed/NCBI
|
|
21
|
Mills CA, Suzuki A, Arceci A, Mo JY,
Duncan A, Salmon ED and Emanuele MJ: Nucleolar and
spindle-associated protein 1 (NUSAP1) interacts with a SUMO E3
ligase complex during chromosome segregation. J Biol Chem.
292:17178–17189. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Vogel A, Cervantes A, Chau I, Daniele B,
Llovet J, Meyer T, Nault JC, Neumann U, Ricke J, Sangro B, et al:
Hepatocellular carcinoma: ESMO Clinical Practice Guidelines for
diagnosis, treatment and follow-up. Ann Oncol. 29:iv238–iv255.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ju LL, Chen L, Li JH, Wang YF, Lu RJ, Bian
ZL and Shao JG: Effect of NDC80 in human hepatocellular carcinoma.
World J Gastroenterol. 23:3675–3683. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Rhodes DR, Kalyana-Sundaram S, Mahavisno
V, Varambally R, Yu J, Briggs BB, Barrette TR, Anstet MJ,
Kincead-Beal C, Kulkarni P, et al: Oncomine 3.0: Genes, pathways,
and networks in a collection of 18,000 cancer gene expression
profiles. Neoplasia. 9:166–180. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Chandrashekar DS, Bashel B, Balasubramanya
SAH, Creighton CJ, Ponce-Rodriguez I, Chakravarthi BVSK and
Varambally S: UALCAN: A portal for facilitating tumor subgroup gene
expression and survival analyses. Neoplasia. 19:649–658. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Lánczky A, Nagy Á, Bottai G, Munkácsy G,
Szabó A, Santarpia L and Győrffy B: miRpower: A web-tool to
validate survival-associated miRNAs utilizing expression data from
2178 breast cancer patients. Breast Cancer Res Treat. 160:439–446.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Anaya J: OncoLnc: Linking TCGA survival
data to mRNAs, miRNAs, and lncRNAs. PeerJ Computer Science.
2:e672016. View Article : Google Scholar
|
|
28
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2 (-Delta Delta C (T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Peterson KA, Nishi Y, Ma W, Vedenko A,
Shokri L, Zhang X, McFarlane M, Baizabal JM, Junker JP, van
Oudenaarden A, et al: Neural-specific Sox2 input and differential
Gli-binding affinity provide context and positional information in
Shh-directed neural patterning. Genes Dev. 26:2802–2816. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Hui CC and Angers S: Gli proteins in
development and disease. Annu Rev Cell Dev Biol. 27:513–537. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kotian S, Banerjee T, Lockhart A, Huang K,
Catalyurek UV and Parvin JD: NUSAP1 influences the DNA damage
response by controlling BRCA1 protein levels. Cancer Biol Ther.
15:533–543. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Shukla V, Coumoul X, Lahusen T, Wang RH,
Xu X, Vassilopoulos A, Xiao C, Lee MH, Man YG, Ouchi M, et al:
BRCA1 affects global DNA methylation through regulation of DNMT1.
Cell Res. 20:1201–1215. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Iyer J, Moghe S, Furukawa M and Tsai MY:
What's Nu (SAP) in mitosis and cancer? Cell Signal. 23:991–998.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Vanden Bosch A, Raemaekers T, Denayer S,
Torrekens S, Smets N, Moermans K, Dewerchin M, Carmeliet P and
Carmeliet G: NuSAP is essential for chromatin-induced spindle
formation during early embryogenesis. J Cell Sci. 123:3244–3255.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Yu HT and Lu PR: Effects of lentiviral
short hairpin RNA silencing of Toll-like receptor 4 on the lens
epithelial cell line HLEC. Genet Mol Res. 15:gmr78972016.
View Article : Google Scholar
|
|
36
|
Zhang L, Liu HJ, Li TJ, Yang Y, Guo XL, Wu
MC, Rui YC and Wei LX: Lentiviral vector-mediated siRNA knockdown
of SR-PSOX inhibits foam cell formation in vitro. Acta Pharmacol
Sin. 29:847–852. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Hanahan D and Weinberg RA: Hallmarks of
cancer: The next generation. Cell. 144:646–674. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Lauss M, Kriegner A, Vierlinger K, Visne
I, Yildiz A, Dilaveroglu E and Noehammer C: Consensus genes of the
literature to predict breast cancer recurrence. Breast Cancer Res
Treat. 110:235–244. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Gordon CA, Gong X, Ganesh D and Brooks JD:
NUSAP1 promotes invasion and metastasis of prostate cancer.
Oncotarget. 8:29935–29950. 2017. View Article : Google Scholar : PubMed/NCBI
|