Introduction
The prevalence of diabetes mellitus (DM) is
increasing each year. The International Diabetes Federation has
estimated that 415 million people have been diagnosed with DM
worldwide and anticipates an increase of up to 640 million by 2040
(1). Diabetic neuropathy (DN) is one
of the most common diabetic complications and is characterized by
complex changes in functional and sensorimotor parameters (2). The pathophysiology of DN involves a
complex cascade of specific interrelated mechanisms (3,4).
Oxidative stress is an important contributor to the development of
DN; this is due to the dramatic increase in the level of free
radicals generated in patients with diabetes (5,6).
Resveratrol (RSV), a naturally-occurring polyphenol
identified in grapes and red wine, has been demonstrated to exert
potent anti-diabetic, anti-oxidative and anti-inflammatory
properties (7). RSV was reported to
inhibit the apoptosis of pancreatic β-cells and significantly
decrease the expression of an inhibitor (nuclear
factor-kappa-B-inhibitor alpha) of nuclear factor (NF)-κB and NF-κB
p65 in NF-κB signaling (8). In
diabetic rats, RSV reduced the levels of malondialdehyde, xanthine
oxidase, nitric oxide and DNA fragmentation in the sciatic nerve,
and increased glutathione levels in the brain (9). Recent studies have demonstrated that
the most beneficial effects of RSV were dependent on sirtuin 1
(SIRT1) activation, through increasing intracellular cyclic
adenosine monophosphate (cAMP) and inhibiting cAMP-dependent
phosphodiesterase (10,11).
Mesenchymal cells from the umbilical cord possess
stem cell-like characteristics, including self-duplication and
differentiation (12). Human
umbilical mesenchymal stem cells (MSCs) may be induced to
differentiate into neuron-like cells in vitro (13). Our previous research demonstrated
that MSCs could improve hyperglycemia and the number of β-cells in
the pancreatic islets of diabetic mice and rats (14–16).
Clinical trials have demonstrated that MSCs may improve
hyperglycemia in patients with diabetes, reduce the dosage of
insulin and oral anti-diabetic drugs, and also decrease the
incidence of diabetic complications (17). Furthermore, additional researchers
have identified that MSCs may facilitate recovery from spinal cord
lesions by releasing brain natriuretic peptide and other vasoactive
factors, which reduce edema, decrease intracranial pressure, and
improve cerebral perfusion (18,19). Kim
et al (20) revealed that the
levels of human neurotrophin-3, vascular endothelial growth factor
receptor-3 and basic fibroblast growth factor in the stem cell
group were higher compared with the PBS group. The mechanism
underlying the promotive effect on the regeneration of the injured
spinal column following the administration of MSCs likely involved
the release of cytokines or growth factors from undifferentiated
stem cells, rather than the differentiation of these cells into
neuronal or glial cells (20).
Recent findings have demonstrated that RSV may
promote the self-renewal and differentiation of MSCs by regulating
SIRT1 signaling, which is associated with cell self-renewal,
senescence, apoptosis and neural differentiation (21). In addition, RSV may regulate cell
growth, osteoblastic differentiation and the expression of
osteogenic genes through estrogen receptor/mitogen-activated
protein kinase/nitric oxide synthase/cyclic guanosine monophosphate
signaling in MSCs cultures (22,23).
Previous studies have investigated the therapeutic effects of RSV
or MSCs on type 1 (T1)DM and DN, although the combined effect of
RSV + MSCs on T1DM and DN, and the mechanism of action, remains
unknown at present. The aim of the present study was to investigate
the combined therapeutic effect and mechanisms of RSV and MSCs on
DN in a mouse model.
Materials and methods
Ethics statement
The present study was approved by the Institutional
Animal Ethical Committee (Qingdao, China) and the Ethics Committee
of the Affiliated Hospital of Qingdao University (Qingdao,
China).
MSC preparation
MSCs were prepared from an umbilical cord obtained
from a healthy mother (28 years old) at The Affiliated Hospital of
Qingdao University on March 10, 2017 and fully informed consent was
obtained 2 weeks prior to delivery. The preparation of MSCs was
performed in the laminar flow laboratory using a previously
described method (19). The MSCs
were cultured for 4 passages to prepare final cell products that
were sterile and all qualified for examination, including aerobes,
mycoplasma, hepatitis B virus, hepatitis C virus, human
immunodeficiency virus, Epstein-Barr virus, cytomegalovirus,
syphilis and endotoxin testing. Cells were stained with CD-PE and
CD-FITC (from the Human MSC Analysis kit; 562245; BD Biosciences,
San Jose, CA, USA) and then analyzed using flow cytometry with a
FACSCalibur flow cytometer (BD Biosciences). These cells highly
expressed cluster of differentiation (CD) 90 (89.37%), CD105
(82.26%), CD73 (90.63%), and CD146 (65%) but not CD34 (0.23%), CD45
(0.02%) and Human Leukocyte Antigen-D Related (0.03%). The
differentiation of MSCs was verified by adipocyte and osteogenic
differentiation.
Osteogenic and adipogenic differentiation
experiments of MSCs were performed as follows: MSCs were seeded in
6-well plates at 1×105 cells/ml per well. When they
reached 80% confluence, the cells were induced using osteogenic
medium, containing 10 mmol/l β-sodium glycerophosphate (G9422), 50
µg/ml ascorbic acid (BP461) and 10 nmol/l dexamethasone (D4902;
each, Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) in standard
medium (containing DMEM/F12, 10% fetal bovine serum (10099-141;
Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA), 100 mg/ml
streptomycin and 100 U/ml penicillin) and adipogenic medium,
including 1 µmol/l hexadecadrol, 10 µmol/l insulin, 200 µmol/l
indomethacin, and 0.5 mmol/l isobutyl-methylxanthine (D8893, I2643,
I7378 and I5879, Sigma-Aldrich; Merck KGaA) for 3 weeks. Following
this, Alizarin Red staining and Oil Red O staining were used to
identify the osteogenic and adipogenic potential, respectively.
Cells (1×104) were fixed with 95% ethanol for 10 min at
room temperature, washed with double distillated water three times
and stained with 0.1% Alizarin Red for 30 min at room temperature.
Samples were then washed with double distillated water 3 times and
photographed under a light microscope (magnification, ×40). For Oil
Red O staining, cells were fixed in ice cold 10% for formalin for
10 min, air dried for 40 min at room temperature and then placed in
absolute propylene glycol solution (W550132, Sigma-Aldrich; Merck
KGaA) for 5 min. Sections were then stained in pre-warmed oil red O
solution (O1516, Sigma-Aldrich; Merck KGaA) for 10 min at room
temperature and differentiated in 85% propylene glycol solution for
5 min, washed in water for 3 min and mounted with glycerin jelly.
All staining results were observed under a light microscope
(magnification, ×40).
Non-obese diabetic (NOD) mice
A total of 100 female NOD mice aged 6 weeks old,
weighing 16–20 g, were purchased from the Nanjing Laboratory Animal
Center (Nanjing, China). Mice were fed specific pathogen-free grade
mouse chow and water ad libitum. Animals were housed at a
temperature of 22–23°C and a humidity of 60%, with a 12 h
light/dark cycle. The fasting plasma glucose (FPG) of all mice were
examined at day 7 after mice were obtained. Notably, NOD mice can
spontaneously develop type 1 diabetes and are useful animal
diabetic models (24). Mice with an
FPG ≥16.6 mmol/l that occurred twice on different days that was
confirmed by pathological examination were considered as diabetic
mice. Cells that exhibited a significant decrease in islet β cells
or isletitis were considered diabetic for the purposes of the
present study.
Experimental grouping and treatment
design
Diabetic NOD mice were randomly divided into the
following five groups (n=10): A diabetic control group, MSCs group,
RSV group, MSCs + RSV group and insulin group. Mice that maintained
normal blood glucose levels throughout the experiment were
considered as normal controls (n=10). Due to the incidence of
diabetes (~80%) and the difference of onset time, 100 mice were
utilized to ensure that there were enough diabetic mice and normal
controls. Mice in the MSCs group were treated with an intravenous
infusion of MSCs (prepared from a healthy mother as aforementioned)
by vena caudalis from the third day following a diagnosis of
diabetes, and with a cell number of 1×106 MSCs per
mouse. Mice in the RSV group were treated with RSV (R5010,
Sigma-Aldrich; Merck KGaA) by intragastric administration from the
third day post-diagnosis with diabetes, and the dosage of RSV was
200 mg/kg per day administered for a total of 56 days. Mice in the
MSCs + RSV group were treated with the aforementioned MSC infusion
and RSV (200 mg/kg per day for 56 days). Mice in the insulin group
were treated with insulin glargine (Lantus; sanofi-aventis, Paris,
France) 1 day following the diagnosis of diabetes to maintain
normal blood glucose levels. Mice in the diabetic control group
were used as diabetic controls and did not receive any treatment.
At 8 weeks following diagnosis with diabetes, all mice were
sacrificed to acquire blood and tissues for analysis. Mice with two
consecutive blood glucose values >400 mg/dl were considered as
overt failure (25).
Observation and assessment
Any changes in the mice, including activity, food
and drink uptake and body weight, were recorded. Tail-vein blood
glucose levels were measured biweekly using OneTouch Horizon
glucose measurement strips (Johnson and Johnson Ltd., Milpitas, CA,
USA) and a glucometer. C-peptide was measured using ELISA (mouse
C-peptide Elisa kit; #90050; NeoScientific, Cambridge, MA, USA),
according to the manufacturer's protocol. The initial dosage of
insulin was based on the individual body weight and blood glucose
level of each mouse. Subsequent insulin dosage was adjusted based
on the level of blood glucose as well as body weight. Insulin
dosage levels were increased by 0.2 U for every 10–15 g increase in
body weight. Diabetic mice in the insulin group were treated with
daily injections of insulin glargine. For the MSCs, RSV and MSCs +
RSV groups, insulin treatment was administrated as described for
the insulin group if blood glucose was beyond glycemic control. All
insulin administrations were performed using subcutaneous
injection, and the dosage of insulin to maintain glycemic control
(FPG ≤8 mmol/l, fed blood glucose ≤10 mmol/l) for diabetic mice was
recorded.
Histological examination
All mice were sacrificed at 8 weeks following
diagnosis with diabetes, and tissues from the pancreas, heart,
liver, kidney and sciatic nerve were acquired. Tissue specimens
were fixed in 10% paraformaldehyde at room temperature for 24 h and
embedded in optimal cutting temperature compound (OCT,
Water-soluble mixture of polyethylene glycol and polyvinyl
alcohol). Tissue sections (6 µm-thick) were stained with
hematoxylin and eosin (H&E) at room temperature for 2 h and
were evaluated with light microscopy (magnification, ×400).
Immunohistochemistry
Tissue sections were washed with phosphate buffered
saline (PBS) for 5 min, deactivated in H2O2
(10 min), washed with distilled water 3 times, fixed with EDTA and
blocked with 5% bovine serum albumin (B2064; Sigma-Aldrich; Merck
KGaA) at room temperature for 1 h. Tissue sections were incubated
with primary antibodies anti-insulin (#I2018), anti-glucagon
(G2654), anti-NF-κB (17–10060), anti-nerve growth factor (NGF;
N3279) and anti-myelin basic protein (MBP; AMAB91062; all
Sigma-Aldrich; Merck KGaA; all 1:300) at 25°C for 60 min. Following
washing with PBS, sections were incubated with the secondary
antibody (M8770; Sigma-Aldrich Merck KGaA; 1:300) at 25°C for 45
min, stained with 3,3′-diaminobenzidine (Dako REAL EnVision
Detection System; Agilent Technologies, Inc., Santa Clara, CA, USA)
for 30 sec, counterstained with hematoxylin for 12 min at room
temperature, dehydrated with a gradient alcohol series and mounted
with neutral gum under light microscopy (magnification, ×400).
Masson staining
Six odd number sections of each sciatic nerve were
stained with Masson's trichrome. Sections were treated with 10%
potassium dichromate and 10% trichloroacetic acid for 30 min,
nuclei were stained with hematoxylin for 20 min, differentiated
with hydrochloric acid and ethanol for 15 sec, returned to blue
coloration with weak ammonia for 15 sec, stained with Masson
solution (Cell Signaling Technology, Inc., Danvers, MA, USA) for 1
min, rinsed with 1% acetic acid, dehydrated with an increasing
ethanol series for 1 min each, cleared with xylene I and II for 10
min to make the sections transparent and finally sealed in resin.
All staining steps were performed at room temperature. Following
this, the sciatic fiber was observed under a light microscope at a
magnification of ×400.
Statistical analysis
All statistical analyses were performed using SPSS
version 17.0 software (SPSS Inc., Chicago, IL, USA). Results are
expressed as the mean ± standard deviation. Differences between six
groups were tested by one-way analysis of variance (ANOVA) and post
hoc analysis with Bonferroni correction for multiple comparisons
was applied. For parameters where repeated measurements were taken
over time (i.e., FPG and dosage of insulin), a two-way
repeated-measures ANOVA was performed. P<0.05 was considered to
indicate a statistically significant difference.
Results
Blood glucose, dosage of insulin and
C-peptide
There were no significant differences in FPG, fed
blood glucose and body weight of mice among the six groups (n=10,
per group) prior to therapy (data not shown). The diagnosis date of
diabetic mice was set as day 1 of the experiment. At day 56, there
were a total of 10, 7 (3 mice died of hyperglycemia, >33.3
mmol/l), 8 (2 mice died of hyperglycemia, >33.3 mmol/l), 9 (1
mice died of hyperglycemia, >33.3 mmol/l), 9 (1 mouse died of
head injury in fighting) and 10 surviving mice in the normal
control, diabetic control, insulin, MSCs, RSV and MSCs + RSV
groups, respectively. For the 6 mice that died of hyperglycemia, a
glucose level of <22.2 mmol/l was determined. In these
circumstances, mice received insulin therapy to attempt to improve
hyperglycemia. However, the following day glucose levels continued
to rise to >33.3 mmol/l, at which point, mice were sacrificed.
All mice in the experiment were female and 5 were placed in a
single cage. Although mice were monitored every day, an injury
occurred in one mouse as aforementioned. The injured mouse was
treated by a veterinarian, but due to subsequent hyperglycemia and
wound infection, this mouse succumbed. Following the administration
of the respective therapies (after Day 3), the body weight of mice
in the RSV, insulin and RSV + MSCs groups were increased compared
with the diabetic control group (P=0.032, P=0.019 and P=0.013,
respectively), as illustrated in Fig.
1.
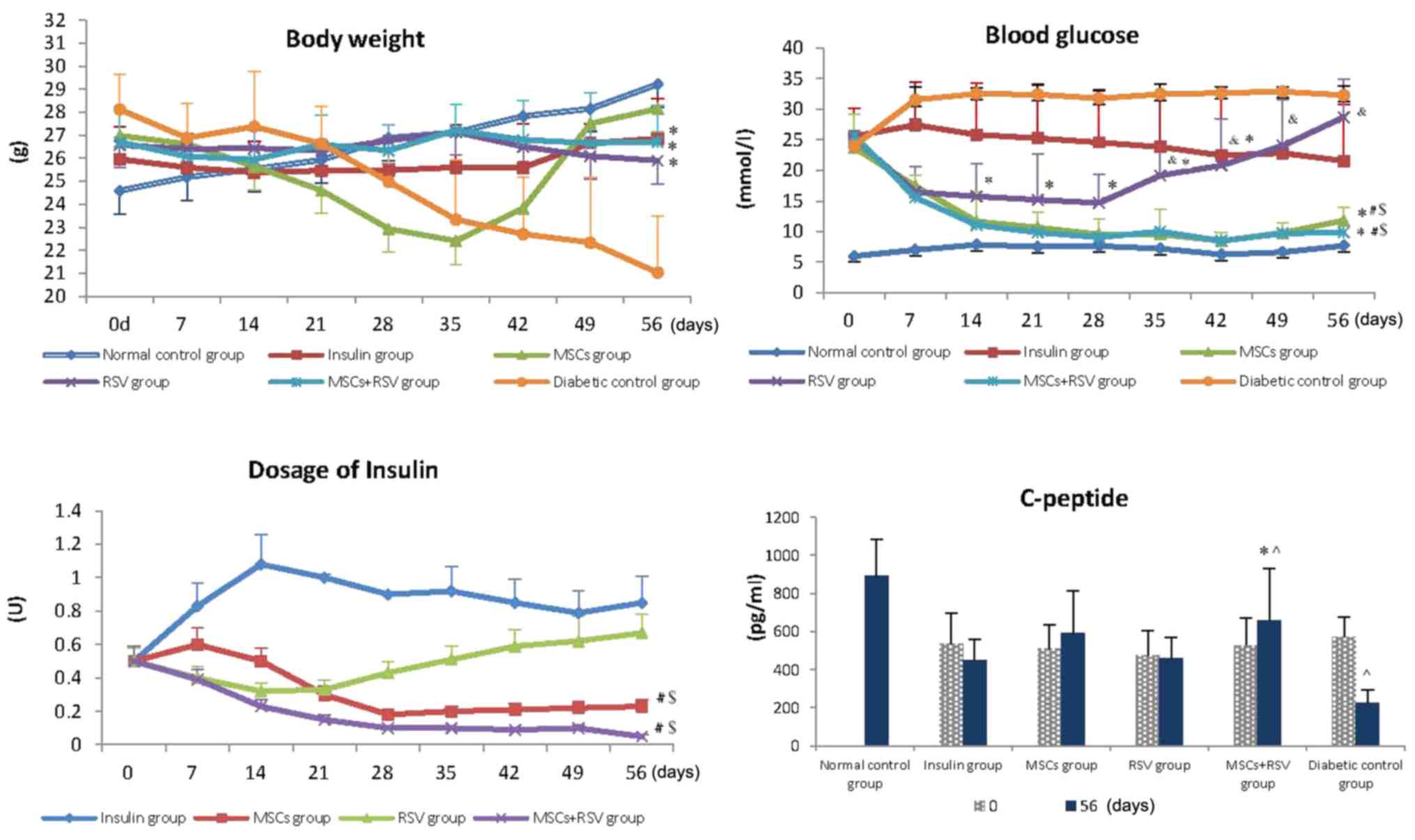 | Figure 1.Body weight, blood glucose, dosage of
insulin and C-peptide were examined among the six experimental
groups. Following the respective administration of therapies, the
body weight of mice in the RSV, insulin and RSV+ MSCs groups was
increased compared with the diabetic control group. Blood glucose
in the MSCs and RSV+ MSCs groups was decreased compared with the
diabetic control, insulin and RSV groups following therapy.
Furthermore, glucose levels in the MSCs and RSV+ MSCs groups
reached a normal level at day 14, which lasted until the end of the
experiment. The blood glucose in the RSV group was lower compared
with the diabetic control group, yet higher compared with the MSCs
group and MSCs + RSV group. The dosage of insulin required in the
MSCs group and RSV+ MSCs group was decreased compared with the
insulin group and RSV group. Levels of C-peptide in the MSCs + RSV
group were improved compared with the diabetic control group, yet
remained lower compared with those of the normal control group.
*P<0.05 vs. the diabetic control group; $P<0.05
vs. the insulin group; #P<0.05 vs. the RSV group;
&P<0.05 vs. the MSCs group and the MSCs + RSV
group; ^P<0.05 vs. the normal control group. MSCs,
mesenchymal stem cells; RSV, resveratrol. |
Compared with the diabetic control group, the blood
glucose of mice in the MSCs, RSV (until day 35) and MSCs + RSV
groups were decreased following therapy (P<0.05). Blood glucose
in the MSCs and MSCs + RSV group reached a normal level at day 14,
and this was maintained until the end of the experiment; no
significant differences were identified in the blood glucose levels
between the two groups (MSCs group vs. MSCs + RSV group). The blood
glucose in the RSV group was lower compared with the diabetic
control group, yet higher compared with the MSCs group and MSCs +
RSV group (P<0.05), and the lower glucose lasted for a total of
28 days. Subsequently, the blood glucose in the RSV group gradually
increased and reached a similar level to the diabetic control group
at day 56 (Fig. 1).
Insulin treatment was administered to the mice with
blood glucose levels of ≥10 mmol/l. As illustrated in Fig. 1, the dosage of insulin in the MSCs
+RSV group decreased from day 7 and the average dosage was 0.15
units during the experiment. The dosage of insulin in the MSCs
group decreased from day 14 and the average dosage was 0.27 units.
In addition, the dosage of insulin in the RSV group was reduced
slightly, followed by an increase until the end of experiment; the
average dosage was 0.46 units. The dose of insulin in the insulin
group was high, with an average dosage of 0.59 units. The dosages
of insulin in the MSCs and MSCs + RSV groups were significantly
reduced compared with the insulin and RSV group (P=0.012, P=0.029,
P=0.001 and P=0.016, respectively).
The present study also examined the levels of
C-peptide in mice on days 0 and 56. Concentrations of the C-peptide
were examined when the mouse was diagnosed as diabetic (0 days).
Mice that maintained normal blood glucose during the experiment
were considered normal controls, so only the concentrations of the
C-peptide in the normal control group at day 56 were examined.
There were no differences in levels of C-peptide in mice among the
insulin, MSCs, RSV, MSCs + RSV and diabetic control groups at the
beginning of experiment. However, at day 56, the level of C-peptide
in the diabetic control group was significantly decreased compared
with the normal control group (P=0.037). The level of C-peptide in
the MSCs + RSV group was significantly increased compared with the
diabetic control, yet it remained lower compared with normal
control group (P=0.041 and P=0.022), as illustrated in Fig. 1.
Histological examination
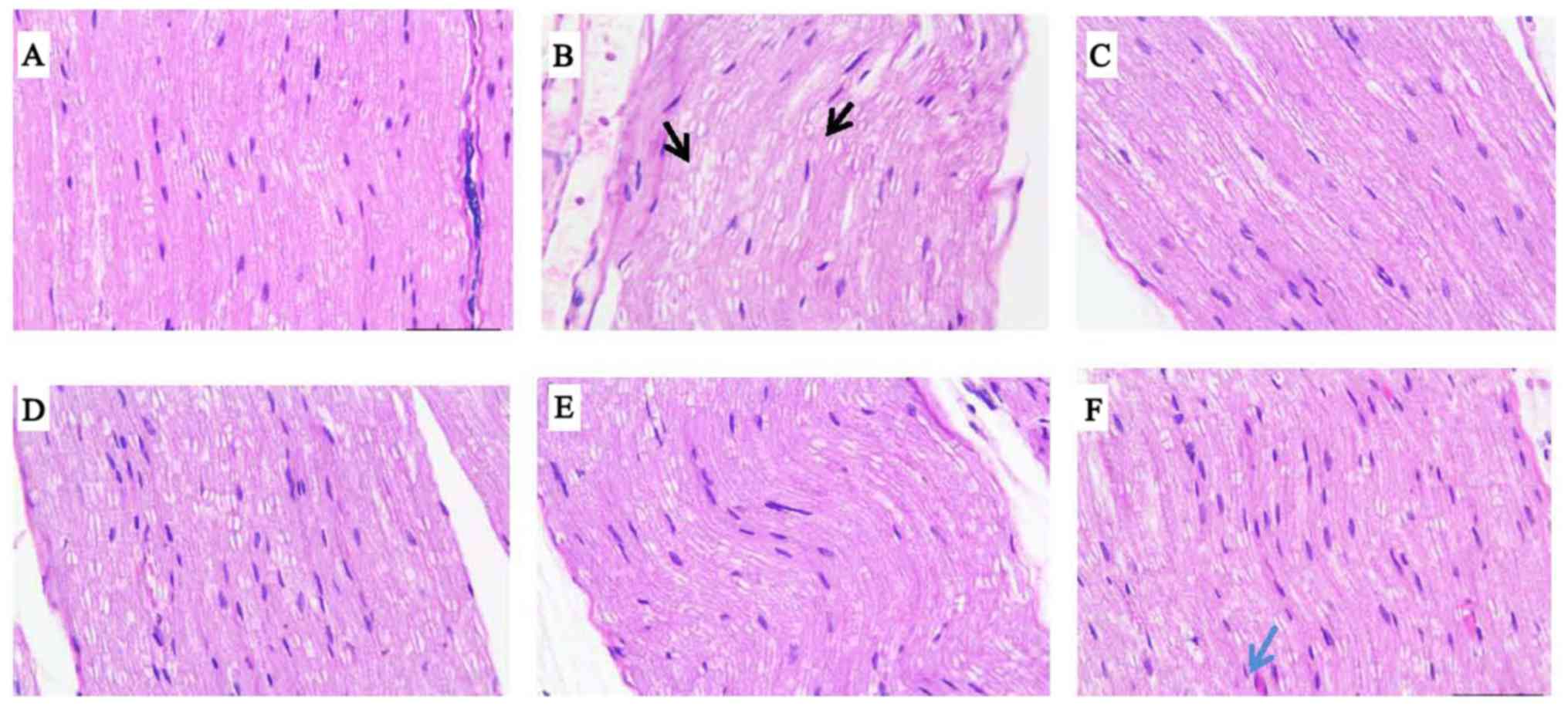
The sciatic nerves of mice were stained with H&E
for light microscopy. As illustrated in Fig. 2, sciatic nerve fibers were
degenerated and misarranged with large cracks and a high degree of
myelin vacuolization in the diabetic control and insulin groups. In
the RSV group, small cracks existed among the sciatic nerve fibers,
and visible myelin vacuolization and new capillaries were not
observed. In the MSCs group, sciatic nerve fibers were arranged
densely and regularly with occasional small cracks, myelin staining
was uniform with small air bubbles, and a small number of new
capillaries were identified under the microscope. In the MSCs + RSV
group, sciatic nerve fibers were densely aligned in a row, the
vacuolar degeneration of myelin was decreased and new capillaries
were observed under microscope, which were increased compared with
those in the MSCs group.
NF-κB, NGF and MBP
The present study examined the levels of the P56
subunit of NF-κB, NGF and MBP to evaluate the inflammation and
recovery of the sciatic nerve on day 56. The level of NF-κB in the
diabetic control group was significantly increased, while levels of
NGF and MBP were decreased compared with the normal control group
(P<0.05, Fig. 3A). Notably,
levels of NF-κB in the insulin group, RSV, MSCs and MSCs + RSV
groups were significantly decreased compared with the diabetic
control group (P=0.041, P=0.015, P=0.034 and P=0.01, respectively).
This indicated that MSCs, insulin and RSV all could improve nerve
inflammation in diabetic mice. Following therapy, levels of NGF in
the MSCs group and MSCs + RSV group were significantly increased
compared with the insulin, RSV and diabetic control groups, yet
were significantly decreased compared with the normal control group
(P<0.05). No statistically significant differences were
identified between the diabetic control, insulin and RSV groups.
Levels of MBP in the insulin group and diabetic control group were
lower than the normal control group, RSV group, MSCs group and MSCs
+ RSV group, as illustrated in Fig.
3C. No statistical differences were observed between the levels
of MBP in the insulin group and the diabetic control group. There
were no significant differences in levels of MBP between the normal
control group, MSCs group, RSV group and MSCs + RSV group. Taken
together, these data suggested that MSCs and RSV relieved
inflammation of the sciatic nerve, and that MSCs may secrete NGF to
promote recovery in the sciatic nerve.
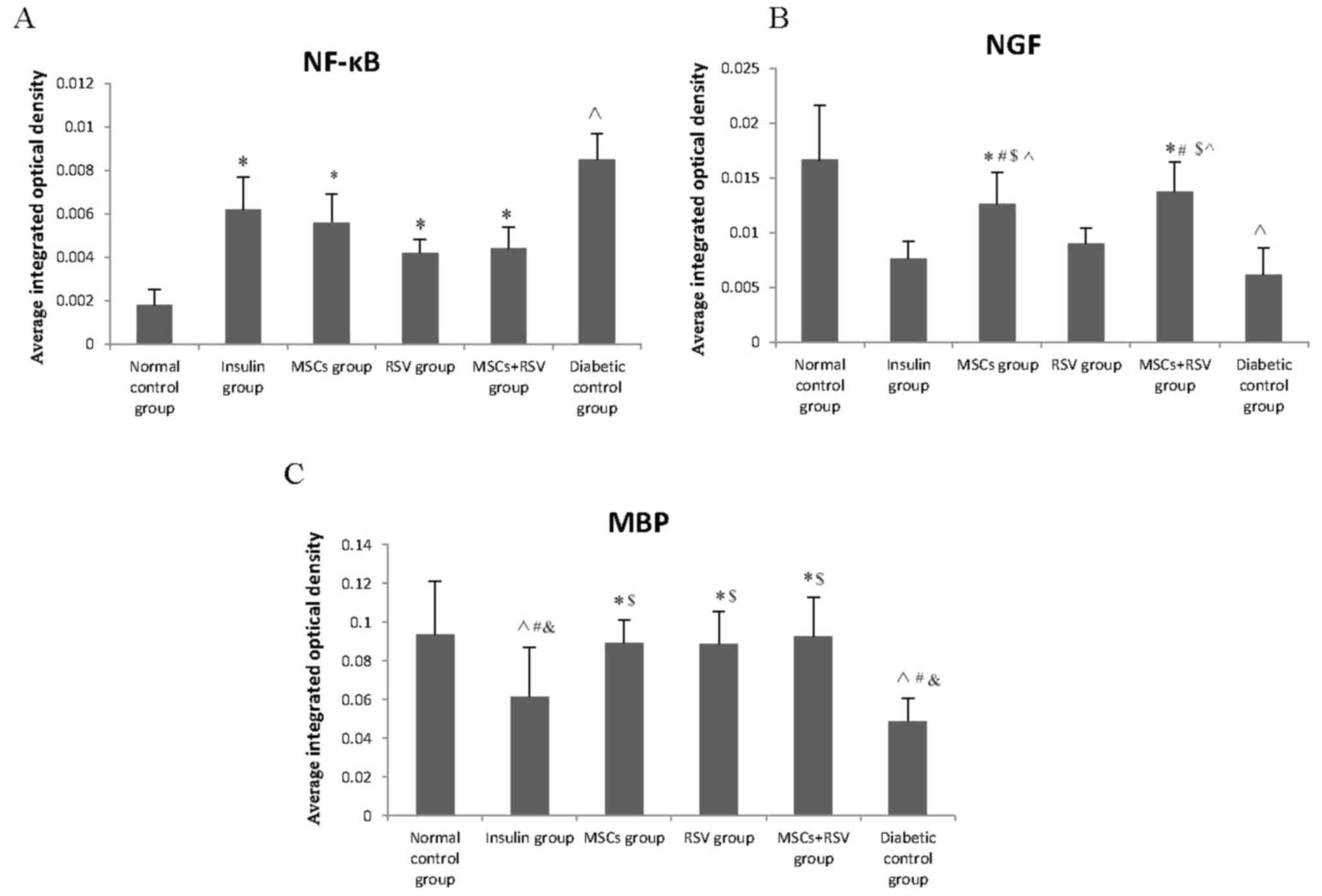 | Figure 3.Levels of NF-κB, NGF and MBP in the
sciatic nerve, assessed by immunohistochemistry. The level of NF-κB
in the diabetic control group was markedly increased, while levels
of NGF and MBP were decreased compared with the normal control
group). (A) Levels of NF-κB in the insulin, RSV, MSCs and MSCs +
RSV group were notably decreased compared with diabetic control
group, particularly in the RSV group and MSCs + RSV group. (B)
Levels of NGF in the MSC group and MSCs + RSV group were higher
compared with the insulin, RSV and diabetic control groups, yet
were significantly decreased compared with the normal control
group. (C) Levels of MBP in the MSCs group, RSV group and MSCs +
RSV group were higher compared with the insulin and diabetic
control groups. Levels of MBP in the insulin and diabetic control
groups were decreased compared with the other four groups.
^P<0.05 vs. the normal control group;
$P<0.05 vs. the insulin group; #P<0.05
vs. the RSV group; *P<0.05 vs. the diabetic control group;
&P<0.05 vs. the MSCs group and the MSCs + RSV
group. MSCs, mesenchymal stem cells; RSV, resveratrol; NGF, nerve
growth factor; MBP, myelin basic protein; NF-κB, nuclear
factor-κB. |
Masson staining
The sciatic nerve was examined using Masson staining
on day 56, as presented in Fig. 4A.
Sciatic nerve fibers were sparse and arranged irregularly, with an
increased number of visible cracks and uneven axonal myelin
staining; the diameters of the axons were markedly different in the
diabetic control group compared with the normal control group.
Compared with the diabetic control group, sciatic nerve fibers in
the insulin, MSCs, RSV and MSCs + RSV groups were improved
substantially and arranged regularly, although the myelin staining
was marginally inhomogeneous (Fig.
4A). Notably, there were no significant differences in the
sciatic nerve between the MSCs + RSV and normal control groups
(P=0.932).
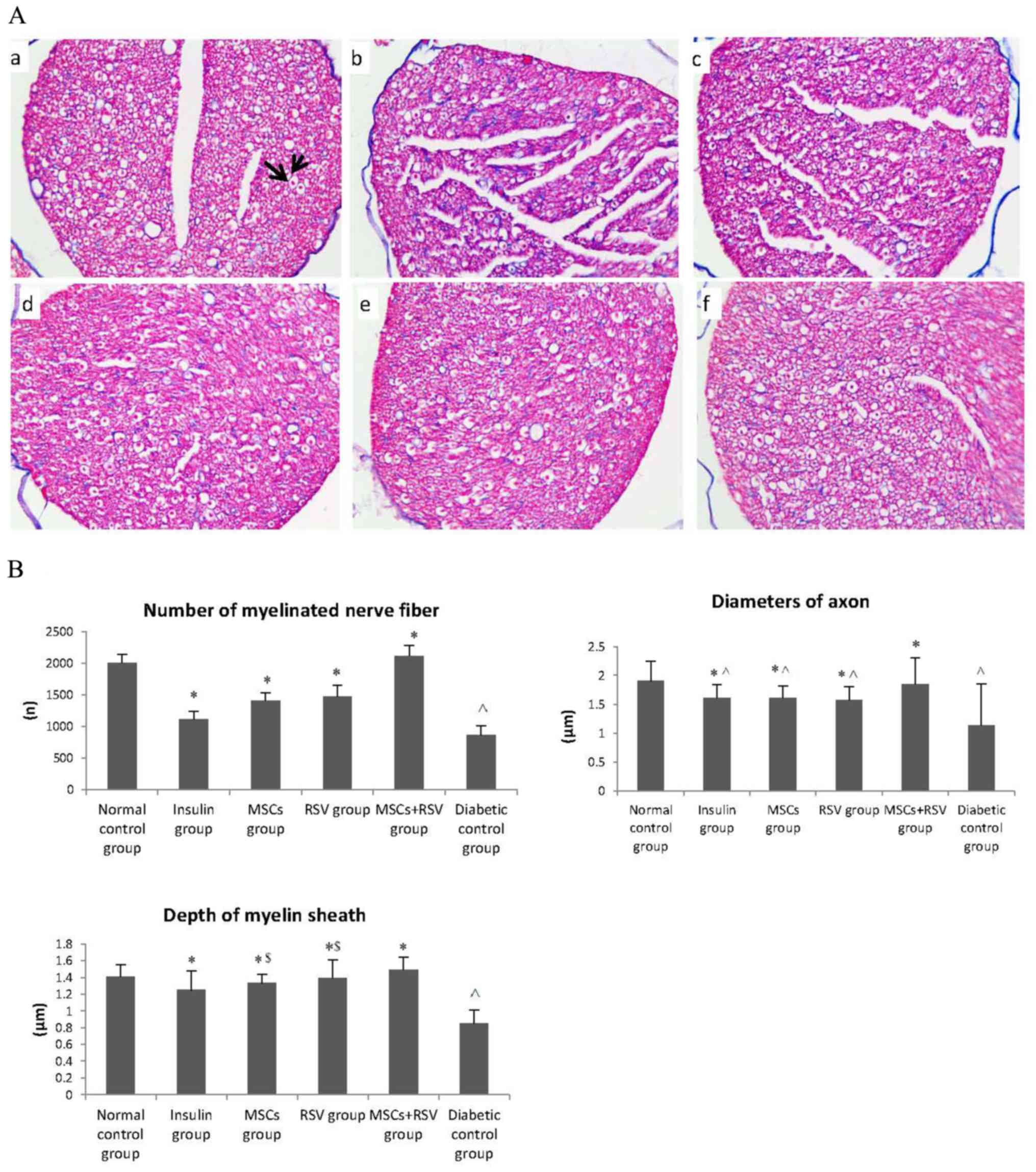 | Figure 4.Diameter of the axon, number of
myelinated nerve fibers and depth of the myelin sheath in the
sciatic nerve, assessed by Masson staining. (A) Sciatic nerve,
assessed by Masson staining. (a) Normal control group, where the
arrows illustrate the axon and myelin sheath; (b) diabetic control
group; (c) insulin group; (d) MSCs group; (e) RSV group; and (f)
MSCs + RSV group. (B) The diameter of the axon, number of
myelinated nerve fibers and depth of the myelin sheath in the
diabetic control group were decreased compared with the normal
control group. Compared with the diabetic control group, the
diameter of the axon, number of myelinated nerve fibers and depth
of the myelin sheath in the insulin, MSCs, RSV group and MSCs + RSV
group were improved, particularly in the MSCs + RSV group. The
depth of the myelin sheath in the MSCs and RSV groups was improved
compared with the insulin group. The axon diameters in the insulin,
MSCs and RSV groups were shorter when compared with the normal
control group. ^P<0.05 vs. the normal control group;
*P<0.05 vs. the diabetic control group; $P<0.05
vs. the insulin group. MSCs, mesenchymal stem cells; RSV,
resveratrol. |
The diameter of the axon, number of myelinated nerve
fibers and depth of the myelin sheath (examined under light
microscope by two pathologists from the Department of Pathology,
the Affiliated Hospital of Qingdao University) in the diabetic
control group were decreased compared with the normal control group
(P=0.035, P=0.029 and P=0.032 respectively; Fig. 4B). Compared with the diabetic control
group, the diameter of the axon, number of myelinated nerve fibers
and depth of the myelin sheath in the insulin, MSCs, RSV and MSCs +
RSV groups were significantly improved (P<0.05), most notably in
the MSC + RSV group. Furthermore, there were no statistically
significant differences in the diameter of the axon, number of
myelinated nerve fibers and depth of the myelin sheath between the
MSCs + RSV group and the normal control group (P=0.798, P=0.836 and
P=0.147, respectively; Fig. 4B).
Furthermore, myelin sheath depths in the MSCs group and RSV group
were significantly improved compared with the insulin group
(P=0.044 and P=0.031). In addition, the axon diameters in the
insulin, MSCs and RSV groups were significantly shorter when
compared with the normal control group (P=0.035, P=0.041 and
P=0.026 respectively; Fig. 4B).
Side effects
There were no symptoms of rejection or toxic effects
observed by macroscopic observation, laboratory or histological
examination among the six experimental groups.
Discussion
Previous studies revealed that MSCs or RSV may
improve hyperglycemia in patients with diabetes; however, there are
few reports regarding the combined effect and mechanisms of MSCs +
RSV on DN. The present study investigated the therapeutic effect of
MSCs + RSV on DN, and demonstrated that levels of blood glucose and
C-peptide in mice in the MSCs + RSV group were significantly
improved compared with the other experimental groups, and the
dosage of insulin required was the lowest among the five diabetic
groups.
In our previous research, MSCs were demonstrated to
increase the number of islet β-cells due to a paracrine effect;
however, this effect did not extend to cell differentiation.
Furthermore, MSCs were revealed to regulate regulatory T cells,
CD4+ T and CD8+ T cells in diabetic mice to
decrease immune attacks on islet cells (15,16). In
the present study, RSV was demonstrated to reduce hyperglycemia;
however, this reduction was more evident following treatment with
MSCs. An increasing amount of evidence has demonstrated that
oxidative stress and inflammatory activity serve a role in the
pathogenesis of diabetes (26,27). RSV
may regulate tissue function by inducing insulin receptor substrate
(IRS)-1 and IRS-2 phosphorylation to induce
phosphatidylinositol-4,5-bisphosphate 3-kinase and Akt
phosphorylation, and increasing endothelial nitric oxide synthase
phosphorylation (28,29). RSV may improve SIRT1 gene and protein
expression, which is suppressed in DM, thereby regulating Forkhead
box O3 deacetylation in vitro and in vivo
(29,30).
The present study also identified that the levels of
NGF in the MSCs and MSCs + RSV groups were significantly increased
compared with the RSV, insulin and diabetic control groups. The
levels of MBP in the RSV, MSCs and MSCs + RSV groups were increased
compared with the insulin and diabetic control groups. In addition,
the levels of NF-κB in the RSV and MSCs + RSV groups were
significantly decreased compared with the MSCs, insulin and
diabetic control groups. These data suggested that RSV may promote
the recovery of the sciatic nerve in diabetic mice by regulating
inflammation, while MSCs may increase the levels of neurotrophic
factors and promote the recovery of the sciatic nerve.
NGF is a small secreted protein that is important
for the growth, maintenance and survival of sympathetic and sensory
neurons (31). Reduced or absent
expression of NGF may lead to cholinergic degeneration and
cognitive impairment in rats, which indicates that NGF may serve as
a therapeutic agent to protect or restore axonal branching and
elongation (32–34). Growing evidence has suggested that
numerous growth factors have dual neurotrophic and angiogenic
effects, including vascular endothelial growth factor, insulin-like
growth factor, NGF, brain derived neurotrophic factor and
fibroblast growth factor-2, which are deficient in DN (35,36). It
has been demonstrated that human MSCs are able to produce >84
trophic factors in conditioned medium and cell lysates (20). Treatment with MSCs partially restored
nerve function in the sciatic nerve via the secretion of vascular
endothelial growth factor, glial cell-derived neurotrophic factor
and NGF and modulation of the immune response, rather than
engraftment and differentiation of MSCs to the injured site
(20,36).
MBP is an important protein synthesized by Schwann
cells, which serves a key role in myelination. Decreased expression
of the MBP has been observed in the sciatic nerve of diabetic rats
(37). Lower expression levels of
MBP signifies the decreased myelination of nerve fibers (38). IN a previous study, following
treatment with RSV/MSCs in diabetic mice, an improvement in the
expression of myelin proteins in the sciatic nerve was observed
(39). Additionally, in the present
study, the diameter of the axon, number of myelinated nerve fibers
and depth of the myelin sheath in the insulin, MSCs, RSV and MSCs +
RSV groups were improved. Furthermore, the therapeutic effect in
the MSCs + RSV group was the greatest, and there were no
statistically significant differences between the MSCs + RSV group
and the normal control group.
In conclusion, the present study investigated the
combined effect of RSV and MSCs on type 1 DN. A combination of RSV
and MSCs was revealed to relieve hyperglycemia and improve DN by
paracrine mechanisms or immune regulation, and the combined
administration of RSV and MSCs led to the most marked improvement
in DN. This indicated that combination of RSV and MSCs may be a
novel therapeutic method of treating T1DM and DN. However, the
specific mechanisms of action of RSV and MSCs in DN require further
clarification; further investigations should focus on the possible
mechanisms of action and potential side effects, due to the complex
nature and pathophysiology of DN. Furthermore, the present study
did not trace and check the MSC cells in vivo. Subsequently,
in further research MSC cells will be investigated in
vivo.
Acknowledgements
The authors would like to acknowledge the patient
who donated her umbillical cord to the present study.
Funding
The present study was supported by the Science and
Technology Development Plan of Shandong Province (grant no.
2015GGB01204) and People's Livelihood Science and Technology
Program of Qingdao (grant no. 15-9-2-81-nsh).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
CW wrote the manuscript and directed/performed the
experiments. JC performed MSC culture and blood examination. KC, XM
and MQ performed the animal experiments and the histological
examination of the sciatic nerves. ZW and YW designed the
experiments, revised and proved the manuscript. All authors read
and approved the final manuscript.
Ethics approval and consent to
participate
The experiments comply with the current laws of the
country in which they were performed. The present study was
approved by the Institutional Animal Ethical Committee (Qingdao,
China) and the Ethics Committee of the Affiliated Hospital of
Qingdao University (Qingdao, China). Informed consent was obtained
for the present study.
Patient consent for publication
The patient who donated her umbillical cord provided
written informed consent for the publication of any associated data
and accompanying images.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Ogurtsova K, da Rocha Fernandes JD, Huang
Y, Linnenkamp U, Guariguata L, Cho NH, Cavan D, Shaw JE and
Makaroff LE: IDF diabetes atlas: Global estimates for the
prevalence of diabetes for 2015 and 2040. Diabetes Res Clin Pract.
128:40–50. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Chung YC, Lim JH, Oh HM, Kim HW, Kim MY,
Kim EN, Kim Y, Chang YS, Kim HW and Park CW: Calcimimetic restores
diabetic peripheral neuropathy by ameliorating apoptosis and
improving autophagy. Cell Death Dis. 9:11632018. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Lima KCA, Borges LDS, Hatanaka E, Rolim LC
and de Freitas PB: Grip force control and hand dexterity are
impaired in individuals with diabetic peripheral neuropathy.
Neurosci Lett. 659:54–59. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Datta I, Bhadri N, Shahani P, Majumdar D,
Sowmithra S, Razdan R and Bhonde R: Functional recovery upon human
dental pulp stem cell transplantation in a diabetic neuropathy rat
model. Cytotherapy. 19:1208–1224. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Román-Pintos LM, Villegas-Rivera G,
Rodríguez-Carrizalez AD, Miranda-Díaz AG and Cardona-Muñoz EG:
Diabetic polyneuropathy in type 2 diabetes mellitus: Inflammation,
oxidative stress, and mitochondrial function. J Diabetes Res.
2016:34256172016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Erbas O, Taşkıran D, Oltulu F, Yavaşoğlu
A, Bora S, Bilge O, Çınar BP and Peker G: Oxytocin provides
protection against diabetic polyneuropathy in rats. Neurol Res.
39:45–53. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Martí-Centelles R, Murga J, Falomira E,
Carda M and Marco JA: Synthesis and biological evaluation of imines
structurally related to resveratrol as dual inhibitors of VEGF
protein secretion and hTERT gene expression. Nat Prod Commun.
12:699–703. 2017.PubMed/NCBI
|
|
8
|
Cao L, Chen X, Xiao X, Ma Q and Li W:
Resveratrol inhibits hyperglycemia-driven ROS-induced invasion and
migration of pancreatic cancer cells via suppression of the ERK and
p38 MAPK signaling pathways. Int J Oncol. 49:735–743. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Soufi FG, Vardyani M, Sheervalilou R,
Mohammadi M and Somi MH: Long-term treatment with resveratrol
attenuates oxidative stress pro-inflammatory mediators and
apoptosis in streptozotocin-nicotinamide-induced diabetic rats. Gen
Physiol Biophys. 31:431–438. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ding S, Jiang J, Zhang G, Bu Y, Zhang G
and Zhao X: Resveratrol and caloric restriction prevent hepatic
steatosis by regulating SIRT1-autophagy pathway and alleviating
endoplasmic reticulum stress in high-fat diet-fed rats. PLoS One.
12:e01835412017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
He Q, Li Z, Wang Y, Hou Y, Li L and Zhao
J: Resveratrol alleviates cerebral ischemia/reperfusion injury in
rats by inhibiting NLRP3 inflammasome activation through
Sirt1-dependent autophagy induction. Int Immunopharmacol.
50:208–215. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ko HR, Ahn SY, Chang YS, Hwang I, Yun T,
Sung DK, Sung SI, Park WS and Ahn JY: Human UCB-MSCs treatment upon
intraventricular hemorrhage contributes to attenuate hippocampal
neuron loss and circuit damage through BDNF-CREB signaling. Stem
Cell Res Ther. 9:3262018. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Goudarzi F, Tayebinia H, Karimi J,
Habibitabar E and Khodadadi I: Calcium: A novel and efficient
inducer of differentiation of adipose-derived stem cells into
neuron-like cells. J Cell Physiol. 233:8940–8951. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Hu J, Fu Z, Chen Y, Tang N, Wang L, Wang
F, Sun R and Yan S: Effects of autologous adipose-derived stem cell
infusion on type 2 diabetic rats. Endocr J. 62:339–352. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Hu J, Wang Y, Wang F, Wang L, Yu X, Sun R,
Wang Z, Wang L, Gao H, Fu Z, et al: Effect and mechanisms of human
Wharton's jelly-derived mesenchymal stem cells on type 1 diabetes
in NOD model. Endocrine. 48:124–134. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Hu J, Wang F, Sun R, Wang Z, Yu X, Wang L,
Gao H, Zhao W, Yan S and Wang Y: Effect of combined therapy of
human Wharton's jelly-derived mesenchymal stem cells from umbilical
cord with sitagliptin in type 2 diabetic rats. Endocrine.
45:279–287. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Hu J, Wang Y, Gong H, Yu C, Guo C, Wang F,
Yan S and Xu H: Long term effect and safety of Wharton's
jelly-derived mesenchymal stem cells on type 2 diabetes. Exp Ther
Med. 12:1857–1866. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Larocca TF, Macêdo CT, Souza BSF,
Andrade-Souza YM, Villarreal CF, Matos AC, Silva DN, da Silva KN,
de Souza CLEM, Paixão DDS, et al: Image-guided percutaneous
intralesional administration of mesenchymal stromal cells in
subjects with chronic complete spinal cord injury: A pilot study.
Cytotherapy. 19:1189–1196. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Stewart AN, Kendziorski G, Deak ZM, Brown
DJ, Fini MN, Copely KL, Rossignol J and Dunbar GL:
Co-transplantation of mesenchymal and neural stem cells and
overexpressing stromal-derived factor-1 for treating spinal cord
injury. Brain Res. 1672:91–105. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kim BJ, Jin HK and Bae JS: Bone
marrow-derived mesenchymal stem cells improve the functioning of
neurotrophic factors in a mouse model of diabetic neuropathy. Lab
Anim Res. 27:171–176. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Yun YC, Jeong SG, Kim SH and Cho GW:
Reduced sirtuin 1/adenosine monophosphate-activated protein kinase
in amyotrophic lateral sclerosis patient-derived mesenchymal stem
cells can be restored by resveratrol. J Tissue Eng Regen Med.
13:110–115. 2019.PubMed/NCBI
|
|
22
|
Guo L, Wang L, Wang L, Yun-Peng S, Zhou
JJ, Zhao Z and Li DP: Resveratrol induces differentiation of human
umbilical cord mesenchymal stem cells into neuron-like cells. Stem
Cells Int. 2017:16513252017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Cilibrasi C, Riva G, Romano G, Cadamuro M,
Bazzoni R, Butta V, Paoletta L, Dalprà L, Strazzabosco M, Lavitrano
M, et al: Resveratrol impairs glioma stem cells proliferation and
motility by modulating the wnt signaling pathway. PLoS One.
12:e01698542017. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Peng Y, Wen D, Lin F and Mahato RI:
Co-delivery of siAlox15 and sunitinib for reversing the new-onset
of type 1 diabetes in non-obese diabetic mice. J Control Release.
292:1–12. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Mathews CE, Xue S, Posgai A, Lightfoot YL,
Li X, Lin A, Wasserfall C, Haller MJ, Schatz D and Atkinson MA:
Acute versus progressive onset of diabetes in NOD mice: Potential
implications for therapeutic interventions in type 1 diabetes.
Diabetes. 64:3885–3890. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Duan Y, Wang L, Han L, Wang B, Sun H, Chen
L, Zhu L and Luo Y: Exposure to phthalates in patients with
diabetes and its association with oxidative stress, adiponectin,
and inflammatory cytokines. Environ Int. 109:53–63. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Hsu JD, Wu CC, Hung CN, Wang CJ and Huang
HP: Myrciaria cauliflora extract improves diabetic nephropathy via
suppression of oxidative stress and inflammation in
streptozotocin-nicotinamide mice. J Food Drug Anal. 24:730–737.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Pektas MB, Koca HB, Sadi G and Akar F:
Dietary fructose activates insulin signaling and inflammation in
adipose tissue: Modulatory role of resveratrol. Biomed Res Int.
2016:80142522016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Sadi G, Pektaş MB, Koca HB, Tosun M and
Koca T: Resveratrol improves hepatic insulin signaling and reduces
the inflammatory response in streptozotocin-induced diabetes. Gene.
570:213–220. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
González-Rodríguez Á, Santamaría B,
Mas-Gutierrez JA, Rada P, Fernández-Millán E, Pardo V, Álvarez C,
Cuadrado A, Ros M, Serrano M and Valverde ÁM: Resveratrol treatment
restores peripheral insulin sensitivity in diabetic mice in a
sirt1-independent manner. Mol Nutr Food Res. 59:1431–1442. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Yamakita S, Matsuda M, Yamaguchi Y, Sawa T
and Amaya F: Dexmedetomidine prolongs levobupivacaine analgesia via
inhibition of inflammation and p38 MAPK phosphorylation in rat
dorsal root ganglion. Neuroscience. 361:58–68. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Ostrovskaya RU, Yagubova SS, Gudasheva TA
and Seredenin SB: Low-molecular-weight NGF mimetic corrects the
cognitive deficit and depression-like behavior in experimental
diabetes. Acta Naturae. 9:94–102. 2017.PubMed/NCBI
|
|
33
|
Bastani A, Rajabi S and Kianimarkani F:
The effects of fasting during ramadan on the concentration of
serotonin, dopamine, brain-derived neurotrophic factor and nerve
growth factor. Neurol Int. 9:70432017. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Seow SLS, Hong SL, Lee GS, Malek SNA and
Sabaratnam V: 6-shogaol, a neuroactive compound of ginger (jahe
gajah) induced neuritogenic activity via NGF responsive pathways in
PC-12 cells. BMC Complement Altern Med. 17:3342017. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Kumar A, Mishra HK, Dwivedi P and
Subramaniam JR: Secreted trophic factors of Human umbilical cord
stromal cells induce differentiation and neurite extension through
PI3K and independent of cAMP pathway. Ann Neurosci. 22:97–106.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Mead B, Logan A, Berry M, Leadbeater W and
Scheven BA: Paracrine-mediated neuroprotection and neuritogenesis
of axotomised retinal ganglion cells by human dental pulp stem
cells: Comparison with human bone marrow and adipose-derived
mesenchymal stem cells. PLoS One. 9:e1093052014. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Domènech-Estévez E, Baloui H, Meng X,
Zhang Y, Deinhardt K, Dupree JL, Einheber S, Chrast R and Salzer
JL: Akt regulates axon wrapping and myelin sheath thickness in the
PNS. J Neurosci. 36:4506–4521. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Hou B, Ye Z, Ji W, Cai M, Ling C, Chen C
and Guo Y: Comparison of the effects of BMSC-derived schwann cells
and autologous schwann cells on remyelination using a rat sciatic
nerve defect model. Int J Biol Sci. 14:1910–1922. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Arteaga O, Revuelta M, Urigüen L, Álvarez
A, Montalvo H and Hilario E: Pretreatment with resveratrol prevents
neuronal injury and cognitive deficits induced by perinatal
hypoxia-ischemia in rats. PLoS One. 10:e01424242015. View Article : Google Scholar : PubMed/NCBI
|