Introduction
Lung cancer is one of the primary causes of cancer
associated mortalities worldwide and >80% of lung cancer cases
are non-small cell lung cancer (NSCLC). In 2008, >1.6 million
people were diagnosed with lung cancer, accounting for 13% of all
newly diagnosed cancer cases and 1.4 million succumbed to lung
cancer, which accounted for 18% of all cancer associated
mortalities (1).
Apoptosis is a process of programmed cell death,
which maintains a healthy survival/death balance in metazoan cells.
Apoptosis is a key regulator of tissue homeostasis and is tightly
regulated by the interactions of activating and inhibitory pathways
(2). Aberrant inhibition of cellular
apoptosis may result in various diseases, including lung cancer
(3). The mechanism of apoptosis is
complicated and is regulated at many levels. The signals of
carcinogenesis regulate the central control points of the apoptotic
pathways, including inhibitor of apoptosis (IAP) proteins.
Inhibition of apoptosis has an important role in the development of
lung cancer (4). It has been
reported that X-linked IAP (XIAP)-associated factor 1 (XAF1) is
able to inhibit proliferation and induce apoptosis in tumor cells
when combined with XIAP directly (5). Furthermore, our previous study
indicated that XAF1 induced apoptosis in lung cancer cells
(6). Tumor necrosis factor-related
apoptosis-inducing ligand (TRAIL) specifically induces apoptosis of
tumor cells, while no toxicity effect on normal cells has been
demonstrated (7–9). In this study, recombinant adenoviral
vectors were transiently transfected into lung adenocarcinoma cells
to recover the expression of XAF1. Apoptotic effects of XAF1 and
TRAIL on A549 lung adenocarcinoma cell lines were investigated,
which may provide an experimental basis for the application of this
treatment in patients with lung cancer.
Materials and methods
Reagents
Recombinant virus Ad5/F35-XAF1 and controlled virus
Ad5/F35-Null were constructed by Shanghai R&S Biotechnology
Co., Ltd. (Shanghai, China) and stored in a laboratory (Ruijin
Hospital, Shanghai Jiaotong University School of Medicine,
Shanghai, China). rhTRAIL was bought from Peprotech, Inc. (Rocky
Hill, NJ, USA). Reverse transcription system was provided by
Promega Corp. (Madison, WI, USA). Primers of XAF1 and β-actin and
2X Tap PCR MasterMix were from Sangon Biotech Company (Shanghai,
China). BCA protein assay kit was bought from Pierce (Thermo Fisher
Scientific, Inc., Waltham, MA, USA). Rat anti-human XAF1 primary
antibody was supplied by Abcam (Cambridge, UK). Poly ADP-ribose
polymerase (PARP) and capase-3 antibodies were purchased from Cell
Signaling Technology, Inc., (Danvers, MA, USA). Mouse anti-rat
secondary antibody was bought from Santa Cruz Biotechnology, Inc.,
(Dallas, TX, USA). MTT and DMSO were purchased from Sigma-Aldrich
(Merck KGaA, Darmstadt, Germany). Annexin V-fluorescein
isothiocyanate (FITC)/propidium iodide (PI) Apoptosis Detection Kit
and flow cytometry kits were supplied by BD Company (Franklin
Lakes, NJ, USA).
Experimental animals
A total of 20 four-week old female BALB/c nude mice
were provided by Animal Experimental Centre of Shanghai Institutes
for Sciences (Shanghai, China). All animal experiments were
approved by and conducted according to the ethical guidelines of
Medicine Laboratory Animal Ethics Committee of Shanghai Jiaotong
School of Medicine (Shanghai, China).
Cell transfection
A549 cells were cultured in RPMI 1640 complete
medium (Gibco; Thermo Fisher Scientific, Inc.) supplemented with
10% FBS for 24 h. Adenovirus vectors carrying XAF1 and null genes,
respectively, were transfected into lung adenocarcinoma A549 cells
(NOR cells) using Lipofectamine 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.). Cells were divided into different groups based
on transfection with or without TRAIL: XAF1 group, XAF1 + TRAIL
group, Null group, and Null + TRAIL group, NOR group, and Null +
TRAIL group. All transfected cells were cultured in RPMI 1640
serum-free medium with 100 PFU/ml multiplicity of infection (MOI).
The blank normal control group was cultured without any lentiviral
vector transfection. Differing concentrations of TRAIL (25, 50 and
100 ng/ml, respectively) were added to the associated groups.
Following incubation for 4 h, the medium as replenished with RPMI
1640 complete medium supplemented with 10% FBS. Cells were
collected after culturing for 48 h at 37°C (5% CO2)
until collection.
Reverse transcription-polymerase chain
reaction (RT-PCR) analysis
To detect the expression levels of XAF1, total RNA
(10 µg) was extracted from cells using TRIzol, according to the
manufacturer's instructions (Invitrogen; Thermo Fisher Scientific,
Inc.). The cDNA was synthesized with a SensiMix™ SYBR-Green
One-Step kit (Quantace; Bioline Reagents, London, UK), according to
the manufacturer's protocols. The cDNA was treated with polymerase
inhibitor and stored in fluid nitrogen cryopreservation to prevent
degradation. The RT reaction was performed at 42°C for 30 min.
2×Taq PCR MasterMix was used for PCR and the total reaction volume
was 20 µl. Primers of XAF1 were:
5′-TCCGCAATTCATGCTCCACGAGTCCTACTG-3′ (forward) and
5′-ACGCGTCGACAAACTCTGAGTCTGGACAAC-3′ (reverse). Primers of β-actin
were: 5′-ATCTGGCACCACACCTTCTACAATGAGCTGC-3′ (forward) and
5′-CGTCATACTCCTGCTTGCTGATCCACATCTGC-3′ (reverse) (Shanghai R&S
Biotechnology Co., Ltd.). Cycling conditions of PCR were: 95°C for
3 min, followed by 32 cycles at 95°C for 45 sec, 57°C for 45 sec,
and 72°C for 45 sec, and final extension at 72°C for 8 min. PCR
products were detected by 2% agarose gel electrophoresis and
analyzed via the Odyssey Infrared Imaging System (Li-Cor
Biosciences, Lincoln, NE, USA).
Western blot analysis
Total proteins were extracted using
radioimmunoprecipitation assay lysis buffer and
phenylmethylsulfonyl fluoride at a ratio of 200:2, and the
concentration was quantified by BCA kit. Proteins were separated by
10% SDS-PAGE, transferred to membrane, and subsequently blocked
with 5% skim milk. Blots were incubated with specific primary
antibodies against β-actin (cat no. ab8227), XAF1 (cat no.
ab217178), caspase-3 (cat no. 9662) and PARP (cat no. 9542; all
1:1,000), at 4°C overnight. Following washing three times by PBS,
the secondary antibodies were added and shaken for 2 h at room
temperature. The membrane was developed and exposed by adding
electrochemiluminescence reagent, and the images were analyzed
using the gel imaging system. Relative expression was analyzed
using Image J 2.0 software (National Institutes of Health,
Bethesda, MD, USA).
MTT assay
Cells were seeded at the density of 1×104
cells/well. After 48 h incubation at 37°C, MTT (5 mg/ml in DMSO)
was added to the wells (20 µl/well) after discarding the
supernatant liquid and incubated for an additional 4 h.
DMSO-treated cellswere used as control. Following this, MTT was
replaced by DMSO (100 µl/well). Absorbance (A value) was measured
at the wavelength of 570 nm after shaking for 15 min in the dark.
Cell proliferation was calculated as follows: Cell proliferation
ratio=(experiment A value-control A value)/control A value ×100%.
Each group was exposed to different concentrations and replicated
three times to calculate an average.
Flow cytometry
Transfected A549 lung adenocarcinoma cells
(1×105 cells/well) were cultured and divided into
different groups, as outlined. Annexin V-FITC and PI staining fluid
were added after incubation at 37°C for 48 h. Flow cytometry was
used to detect apoptosis of cells in each group. Annexin
V-positivity indicates the apoptotic cells are in early stage,
while PI positive means the cells are necrotic. Annexin V and
PI-double positive cells are late apoptotic cells.
Xenograft mice model
A total of 20 four-week old female BALB/c nude mice
were divided into four groups (XAF1, XAF1 + TRAIL, Null and Null +
TRAIL), as outlined. A549 cell lines (with 1×106/0.1 ml
PBS) were injected into the right side of the back ribs of each
mouse. When the tumor could be observed by naked eyes, tumor size
was measured using a caliper every 7 days. Tumor volume (V) was
calculated according to the following formula: V=V=4/3πxL/2×(w/2)
2; where L is a relatively shorter diameter and w is a relatively
longer diameter. Animals were sacrificed on day 30 after injection,
and their tumors were weighed and harvested. Images were captured
of the tumor specimens, which were subsequently fixed using neutral
formaldehyde and paraffin-embedded for further immunohistochemical
analysis.
Immunohistochemical (IHC)
analysis
Tissue sections were kept at 80°C for 30 min and
subsequently de-paraffinized and rehydrated to retrieve antigen and
block peroxidase. Following blocking with non-immune sheep serum,
rat anti-human XAF1 primary antibody (1:800; cat no. ab217178;
Abcam) was added and incubated at 4°C overnight. Following this,
tumor sections were incubated with horseradish peroxide-conjugated
goat anti-rabbit immunoglobulin G (1:1,000; cat no. cw0105; Beijing
Cowin Biotech Co., Ltd., Beijing, China) and enough peroxidase
substrate was added. A light microscope was used to control
staining and PBS was used to terminate the reaction. Nuclei were
stained with hematoxylin. Following dehydration and coating with
resinene resin, XAF1 IHC slices were observed under a light
microscope. Cells stained brown were deemed as positive.
Statistical analysis
Data are expressed as the mean ± standard deviation.
SPSS 16.0 statistical software (SPSS, Inc., Chicago, IL, USA) was
used for statistical analysis. Analysis of variance (ANOVA) with
Tukey's post hoc test was used for comparisons among multiple
groups. P<0.05 was considered to indicate a statistically
significant difference.
Results
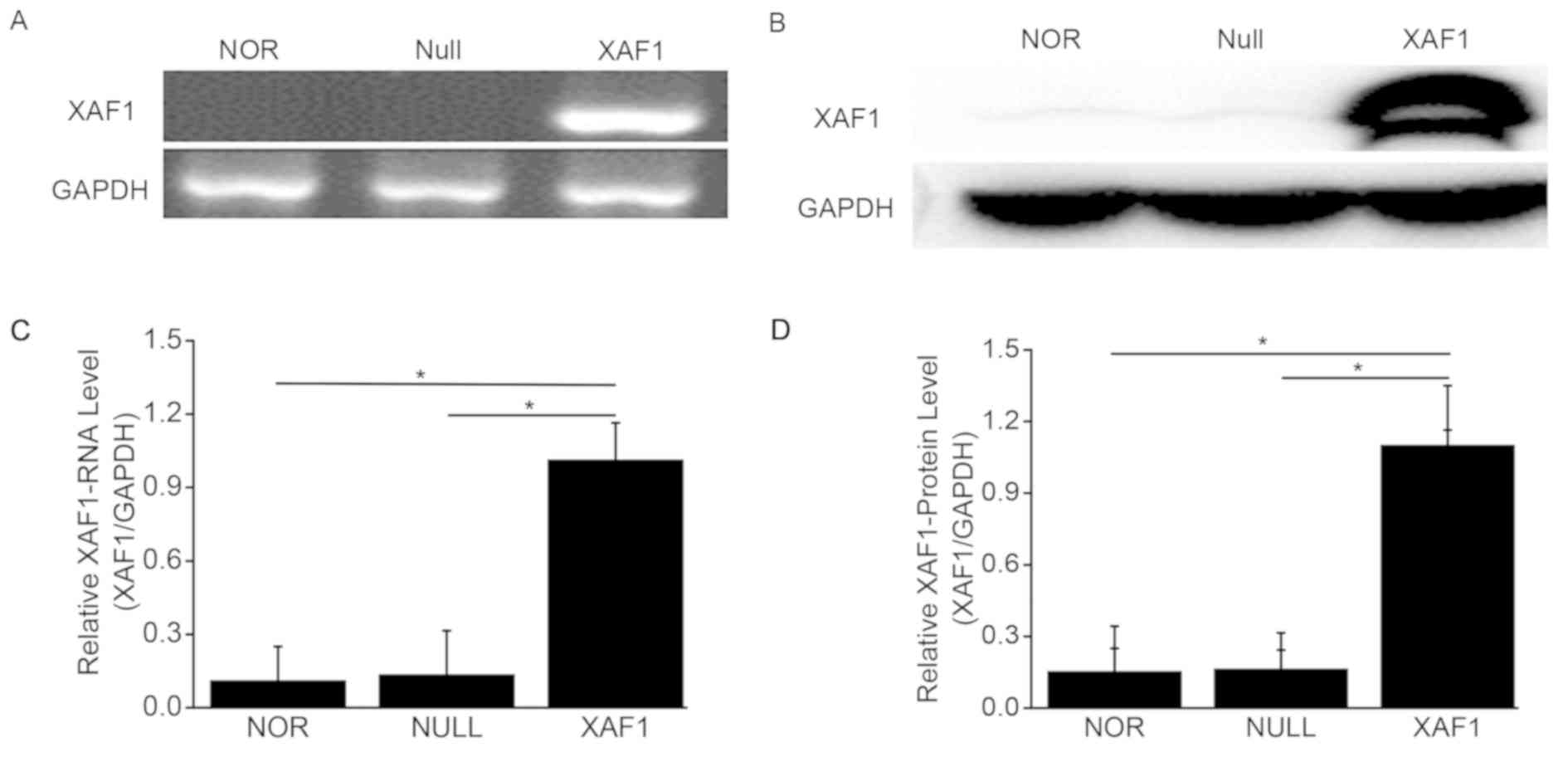
mRNA and protein expression of XAF1 in
A549 cells
To explore whether the expression of XAF1 changed in
transfected cells, RT-PCR and western blot analysis were employed
to detect XAF1 in the different groups of A549 cells. As shown in
Fig. 1A, XAF1 mRNA was markedly
increased in A549 cells after 48 h of XAF1 group, and XAF1 protein
was also upregulated in the XAF1 group when compared with the NOR
and Null groups (Fig. 1B).
Quantitative analysis of gene and the western blot data were
presented respectively (Fig. 1C and
D). These results showed that XAF1 was significantly increased
after transfection with Ad5/F35-XAF1 vector in A549 cells
(P<0.05).
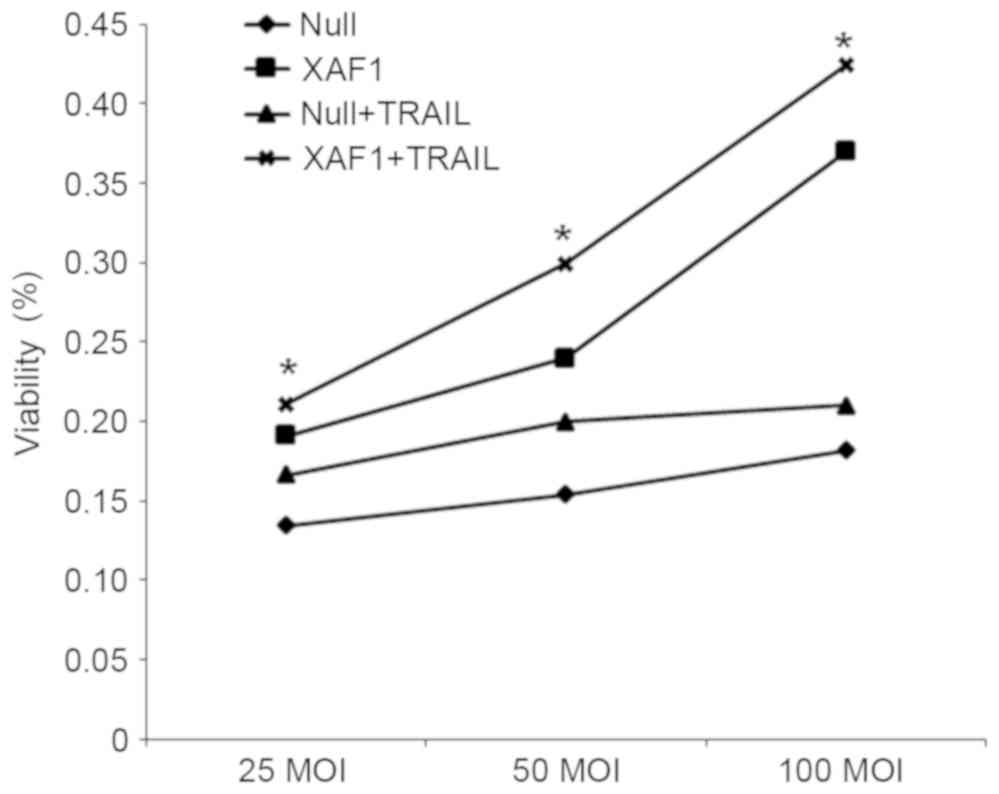
Cell proliferation
To investigate the effect of XAF1 and TRAIL on cell
proliferation, MTT assay was applied to detect the rate of
proliferation in cells of each group. As shown in Fig. 2, the inhibiting effect on the
proliferation of A549 cells demonstrated a positive correlation
with the concentration of XAF1 in the XAF1 + TRAIL group. Among the
Null, XAF1 and Null groups, the inhibition ratio of proliferation
was significantly higher in XAF1 group than in the other groups at
the same concentration of TRAIL (P<0.05). These findings
indicated that TRAIL and different concentrations of XAF1 were able
to inhibit the proliferation of A549 cells in a dose-dependent
manner.
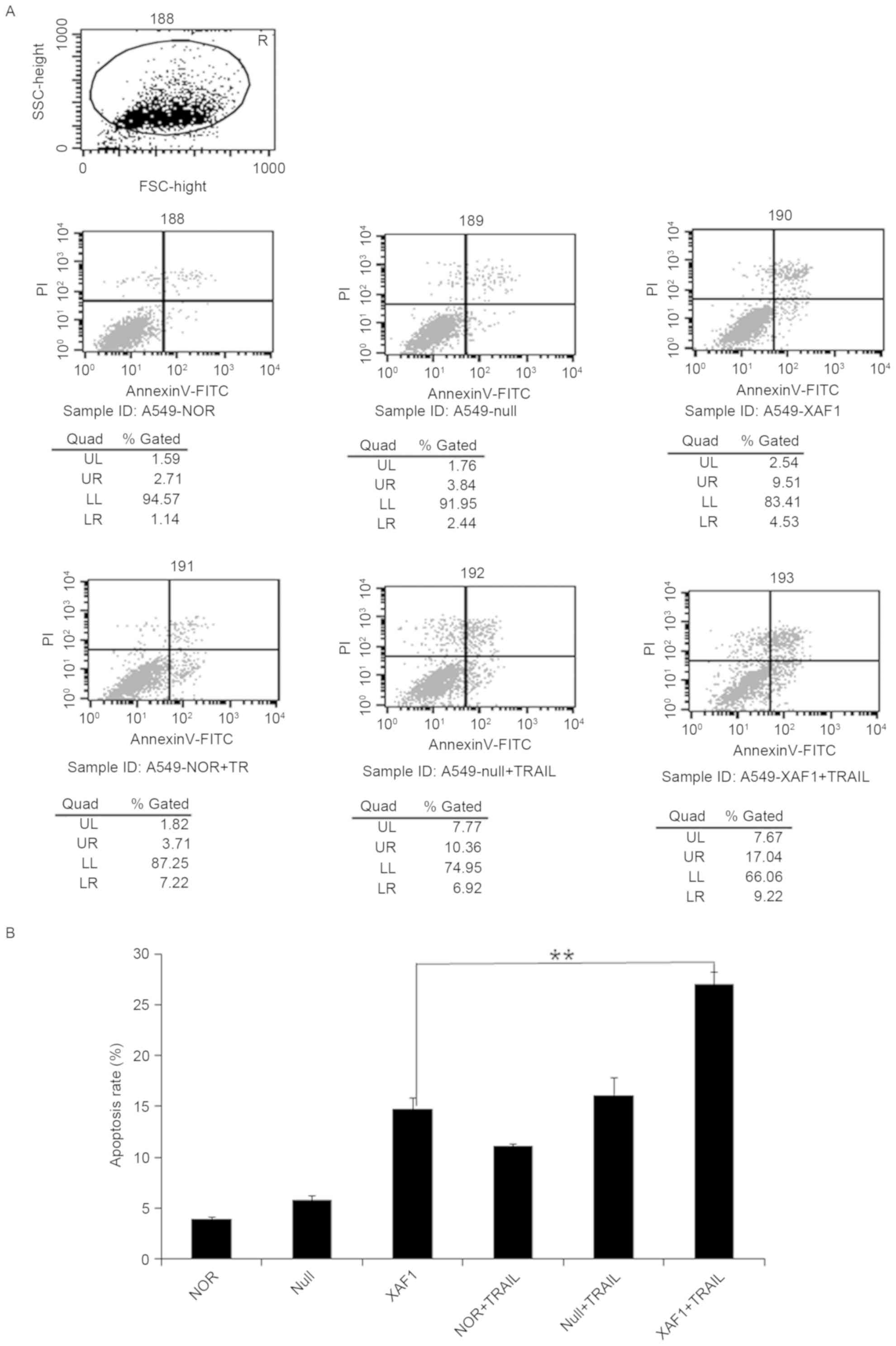
Cell apoptosis by XAF1
To detect how XAF1 regulated apoptosis in A549
cells, the Annexin V-FITC/PI double staining method was used to
assess the different groups. Among the six experimental groups,
compared with the XAFl and Null groups, the apoptosis rate in the
XAF1 + TRAIL group was significantly higher (P<0.05), whereas no
statistically difference was observed between the Null + TRAIL and
NOR groups (Fig. 3). These results
demonstrated that TRAIL was able to promote the apoptotic effect of
Ad5/F35-XAF1 on A549 cells. These findings suggested that a
combination of TRAIL and XAF1 may induce cell apoptosis
coordinately, with TRAIL enhancing the sensitivity of XAF1 to
induce A549 cell apoptosis.
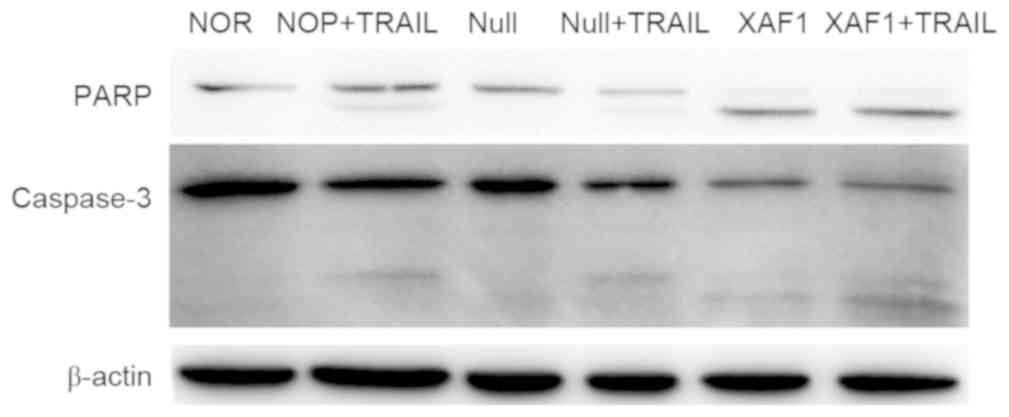
Expression of apoptosis-related
proteins
To detect changes in the expression of
apoptosis-related proteins, western blot analysis was used to
detect the expression levels of PARP, caspase-3 and XAF1 in A549
cells after transfection with Ad5/F35-XAF1 (MOI 150) for 4 h. As
shown in Fig. 4, the results showed
that the XAF1 + TRAIL induced marked PARP and caspase-3 cleavage
compared with other groups (Fig. 4).
No notable difference was observed between the NOR and Null
groups.
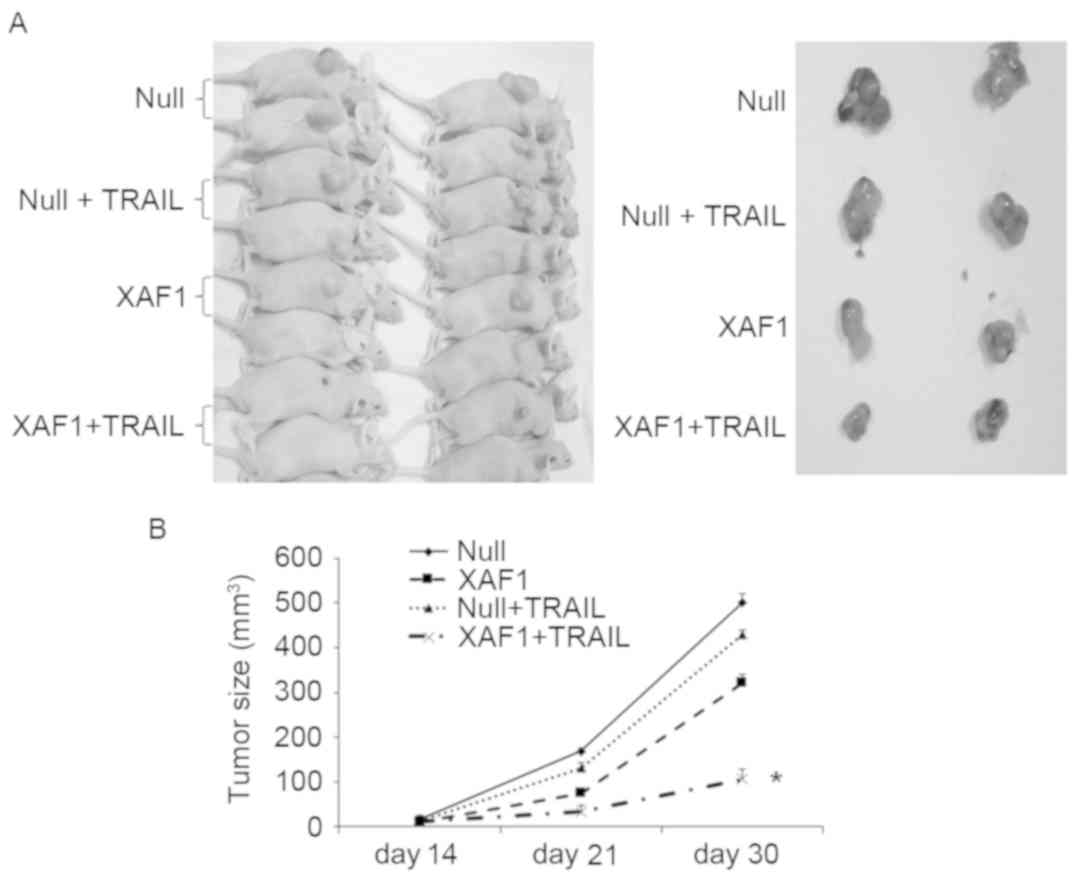
Transplanted tumor model of nude
mice
To determine the impact of XAF1 following combined
treatment with TRAIL in vivo, a transplanted tumor model of
nude mice was generated. Visible tumor tissue was formed 14 days
after the injections in the four groups of nude mice (XAF1, XAF1 +
TRAIL, Null and Null + TRAIL). Subcutaneously transplanted tumors
were stripped from nude mice 30 days after injection (Fig. 5A). Growth rates of subcutaneously
transplanted tumors in the XAF1 and XAF1 + TRAIL groups were
significantly lower than those of the Null and Null + TRAIL groups
(P<0.05). Growth rates of subcutaneously transplanted tumors in
the XAF1 + TRAIL group were significantly lower than those of the
XAF1 group (P<0.05) and no significant difference was observed
between the Null and Null + TRAIL groups (Fig. 5B). These results indicate that XAF1 +
TRAIL inhibited the growth of tumor cells in the murine xenograft
model.
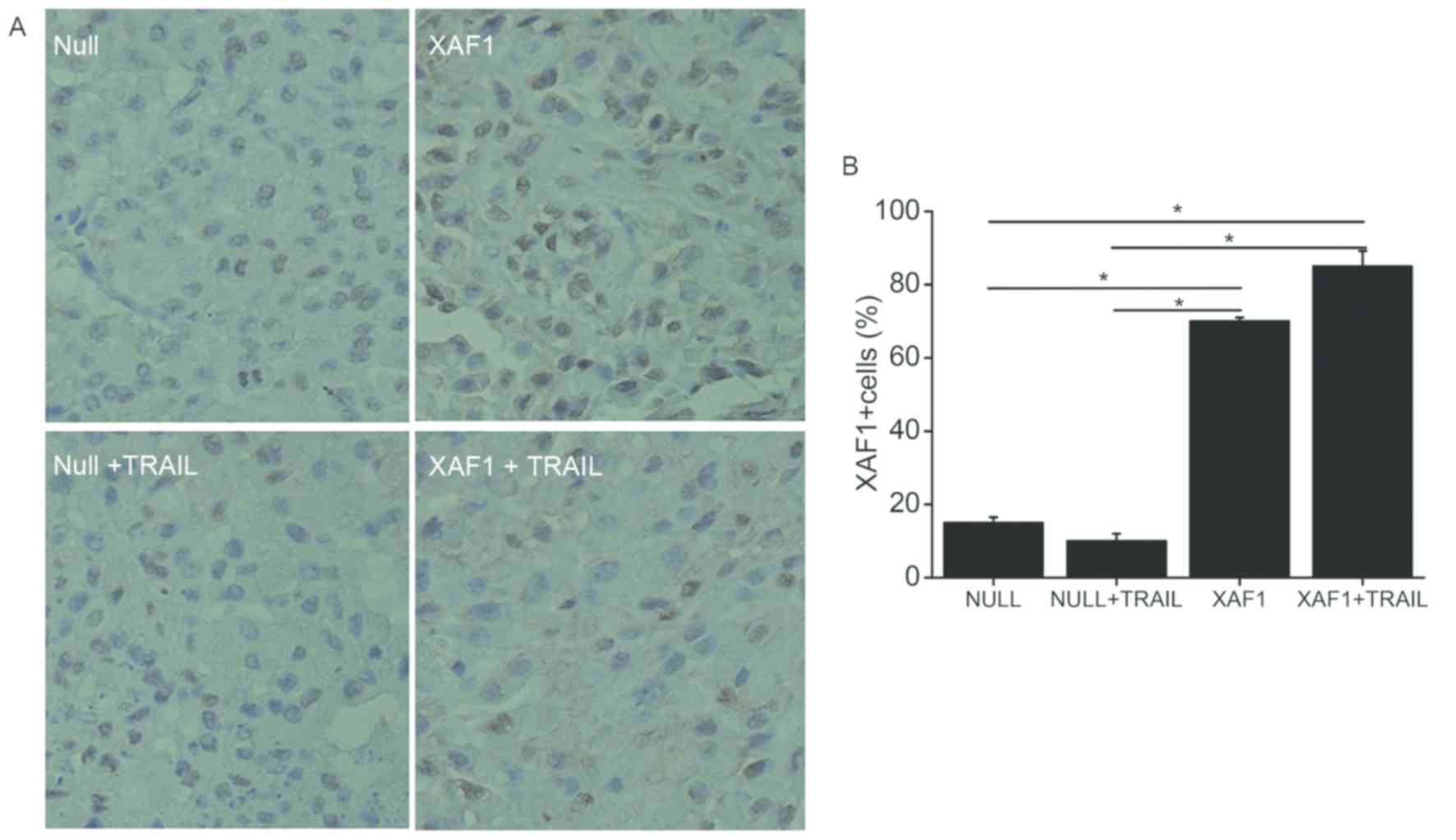
Expression of XAF1 protein on
transplanted tumor tissue biopsies
To determine the expression of XAF1 protein in
vivo, tumor tissue biopsies were performed. According to IHC
analysis, the expression of XAF1 protein was significantly higher
in the XAF1 and XAF1 + TRAIL groups when compared with the Null and
Null + TRAIL groups (P<0.05; Fig.
6). The results indicate that XAF1 may have important roles in
inhibiting the growth of xenograft.
Discussion
Lung cancer, which is one of the most common
malignant tumors in China, has become the major cause of
cancer-associated mortality due to its gradually increasing
morbidity and mortality rates (10).
Multiple studies have confirmed that inhibition of cell apoptosis
is common and dysregulation of cell apoptosis has an important role
in tumorigenesis and tumor progression (3,11). XAF1
is a newly discovered type of XIAP antagonistic protein that was
via a yeast two-hybrid system (12).
XAF1 is able to directly degrade XAP1 via mitochondrial pathways
(13). XAF1 is a tumor suppressor
gene that has been demonstrated to decrease multiple tumor cells
and tissues in humans (14).
Numerous patients with terminal lung cancer who are unsuitable for
surgery are administered with conventional chemotherapeutics, which
kill tumor cells and normal tissue cells indistinctively, with
serious side effects. Therefore, the present study aimed to
investigate the combined effects of XAF1 and TRAIL, with the hope
that this therapy would kill tumor cells but not normal cells in
the A549 lung adenocarcinoma cell line. Whether XAF1 and TRAIL was
able to kill lung tumor cells and induce cell apoptosis
collaboratively or not was investigated, and the mechanism may
provide a novel type of gene therapy in lung cancer.
TRAIL is a tumor-targeted therapy and its specific
effect on killing tumor cells has been well-documented (15–17). At
present, rhTRAIL alone or combined with chemotherapeutics for the
treatment of various human tumors has progressed to phase III
clinical trials and no obvious side effects has been observed. This
indicates that TRAIL, as a tumor targeted therapy, possesses wide
prospects for clinical application (18–20).
XAF1 is able to significantly inhibit the growth of gastric cancer
and enhance the apoptosis of gastric cancer cells, which prolonged
the survival time of the mice under investigation when combined
with TRAIL (21,22). In this study, cell proliferation in
the XAF1 + TRAIL group was significantly lower than in XAF1 group
at the same concentration, while no difference was observed when
compared with Null group. This indicates that TRAIL enhanced the
killing effect of XAF1 on A549 lung adenocarcinoma cells.
Furthermore, XAF1 alone was able to induce lung cancer cell
apoptosis and promote apoptosis rates significantly when combined
with TRAIL. There was no significant difference between the Null
group and Null + TRAIL group, which indicates that XAF1 and TRAIL
induce lung cancer cell apoptosis synergistically.
Apoptosis-related proteins have important roles in
the progression of programmed cell death. Caspase-3, which is a
member of the caspase family of 13 aspartate-specific cysteine
proteases that have a central role in the execution of apoptotic
mechanisms (23–25) is primarily responsible for the
cleavage of PARP during cell death (26). PARP was suggested to contribute to
cell death by depleting cells of NAD and ATP (27), as it is activated by binding to DNA
ends or strand breaks. The present study investigated the effect of
XAF1 and TRAIL on the growth of lung tumors in vivo and
in vitro, and western blotting results confirmed that XAF1
and TRAIL led to activation of caspase-3 and PARP. This result
indicates that the mechanism of XAF1 and TRAIL in inhibiting
proliferation and inducing apoptosis in A549 lung adenocarcinoma
cells is related to caspase-associated apoptosis signaling
pathways.
In conclusion, XAF1 combined with TRAIL was able to
significantly inhibit proliferation and induce apoptosis
synergistically in lung cancer cells in vivo and in
vitro. As TRAIL has been progressed to phase III clinical
trials with preferably security, XAF1 + TRAIL may be a potential
clinical therapy strategy for the treatment of lung cancer.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Jiangsu
Provincial Commission of Health and Family Planning (grant no.
H201521), the Natural Science Foundation of Jiangsu Province (grant
no. BK20161224), Science and Technology Research Foundation of
Suzhou Municipality (grant no. SYS2018063), and the Youth Science
and Technology Project of Suzhou Health and Family Planning
Commission (grant no. KJXW2016016).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
All authors contributed to designing the study,
performing experiments, collecting and analyzing data and preparing
the manuscript. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
All animal experiments were approved by and
conducted according to the ethical guidelines of Medicine
Laboratory Animal Ethics Committee of Shanghai Jiaotong School of
Medicine (Shanghai, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lockshin RA and Williams CM: Programmed
cell death-I. Cytology of degeneration in the intersegmental
muscles of the Pernyi silkmoth. J Insect Physiol. 11:123–133. 1965.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Hanahan D and Weinberg RA: The hallmarks
of cancer. Cell. 100:57–70. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Yan F, Gou L, Yang J, Chen L, Tong A, Tang
M, Yuan Z, Yao S, Zhang P and Wei Y: A novel pro-apoptosis gene
PNAS4 that induces apoptosis in A549 human lung adenocarcinoma
cells and inhibits tumor growth in mice. Biochimie. 91:502–507.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Yang WT, Chen DL, Zhang FQ, Xia YC, Zhu
RY, Zhou DS and Chen YB: Experimental study on inhibition effects
of the XAF1 gene against lung cancer cell proliferation. Asian Pac
J Cancer Prev. 15:7825–7829. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Yang WT, Chen DL, Zhang FQ, Xia YC, Zhu
RY, Zhou DS and Chen YB: Experimental study on inhibition effects
of the XAF1 gene against lung cancer cell proliferation. Asian Pac
J Cancer Prev. 15:7825–7829. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ashkenazi A, Pai RC, Fong S, Leung S,
Lawrence DA, Marsters SA, Blackie C, Chang L, McMurtrey AE, Hebert
A, et al: Safety and antitumor activity of recombinant soluble Apo2
ligand. J Clin Invest. 104:155–162. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hao C, Song JH, Hsi B, Lewis J, Song DK,
Petruk KC, Tyrrell DL and Kneteman NM: TRAIL inhibits tumor growth
but is nontoxic to human hepatocytes in chimeric mice. Cancer Res.
64:8502–8506. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lawrence D, Shahrokh Z, Marsters S,
Achilles K, Shih D, Mounho B, Hillan K, Totpal K, DeForge L, Schow
P, et al: Differential hepatocyte toxicity of recombinant
Apo2L/TRAIL versions. Nat Med. 7:383–385. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Arrieta O, Saavedra-Perez D, Kuri R,
Aviles-Salas A, Martinez L, Mendoza-Posada D, Castillo P, Astorga
A, Guzman E and De la Garza J: Brain metastasis development and
poor survival associated with carcinoembryonic antigen (CEA) level
in advanced non-small cell lung cancer: A prospective analysis. BMC
Cancer. 9:1192009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Brown JM and Attardi LD: The role of
apoptosis in cancer development and treatment response. Nat Rev
Cancer. 5:231–237. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
12
|
Liston P, Fong WG and Korneluk RG: The
inhibitors of apoptosis: There is more to life than Bcl2. Oncogene.
22:8568–8580. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Straszewski-Chavez SL, Visintin IP,
Karassina N, Los G, Liston P, Halaban R, Fadiel A and Mor G: XAF1
mediates tumor necrosis factor-alpha-induced apoptosis and X-linked
inhibitor of apoptosis cleavage by acting through the mitochondrial
pathway. J BiolChem. 282:13059–13072. 2007.
|
|
14
|
Fong WG, Liston P, Rajcan-Separovic E, St
Jean M, Craig C and Korneluk RG: Expression and genetic analysis of
XIAP-associated factor 1 (XAF1) in cancer cell lines. Genomics.
70:113–122. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Finnberg N and El-Deiry WS: Selective
TRAIL-induced apoptosis in dysplastic neoplasia of the colon may
lead to new neoadjuvant or adjuvant therapies. Clin Cancer Res.
12:4132–4136. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Jalving M, de Jong S, Koornstra JJ,
Boersma-van Ek W, Zwart N, Wesseling J, de Vries EG and Kleibeuker
JH: TRAIL induces apoptosis in human colorectal adenoma cell lines
and human colorectal adenomas. Clin Cancer Res. 12:4350–4356. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhang L and Fang B: Mechanisms of
resistance to TRAIL-induced apoptosis in cancer. Cancer Gene Ther.
12:228–237. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Herbst RS, Mendolson DS, Ebbinghaus S,
Gordon MS, O'Dwyer P, Lieberman G, Ing J, Kurzrock R, Novotny W and
Eckhardt G: A phase I safety and pharmacokinetic (PK) study of
recombinant Apo2L/TRAIL, an apoptosis-inducing protein in patients
with advanced cancer. J ClinOncol. 24:1242006.
|
|
19
|
Soria JC, Mark Z, Zatloukal P, Szima B,
Albert I, Juhász E, Pujol JL, Kozielski J, Baker N, Smethurst D, et
al: Randomized phase II study of dulanermin in combination with
paclitaxel, carboplatin, and bevacizumab in advanced non-small-cell
lung cancer. J Clin Oncol. 29:4442–4451. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Yee L, Fanale M, Dimick K, Calvert S,
Robins C, Ing J, Ling J, Novotny W, Ashkenazi A and Burris H: A
phase IB safety and pharmacokinetic (PK) study of recombinant human
Apo2L/TRAIL in combination with rituximab in patients with
low-grade non-Hodgkin lymphoma. J ClinOncol. 25:80782007.
|
|
21
|
Tu SP, Liston P, Cui JT, Lin MC, Jiang XH,
Yang Y, Gu Q, Jiang SH, Lum CT, Kung HF, et al: Restoration of XAF1
expression induces apoptosis and inhibits tumor growth in gastric
cancer. Int J Cancer. 125:688–697. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Tu SP, Sun YW, Cui JT, Zou B, Lin MC, Gu
Q, Jiang SH, Kung HF, Korneluk RG and Wong BC: Tumor suppressor
XIAP-associated factor 1 (XAF1) cooperates with tumor necrosis
factor-related apoptosis-inducing ligand to suppress colon cancer
growth and trigger tumor regression. Cancer. 116:1252–1263. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Alnemri ES, Livingston DJ, Nicholson DW,
Salvesen G, Thornberry NA, Wong WW and Yuan J: Human ICE/CED-3
protease nomenclature. Cell. 87:1711996. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Cohen GM: Caspases: The executioners of
apoptosis. Biochem J. 326:1–16. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wang J and Lenardo MJ: Roles of caspases
in apoptosis, development, and cytokine maturation revealed by
homozygous gene deficiencies. J Cell Sci. 113:753–757.
2000.PubMed/NCBI
|
|
26
|
Tewari M, Quan LT, O'Rourke K, Desnoyers
S, Zeng Z, Beidler DR, Poirier GG, Salvesen GS and Dixit VM:
Yama/CPP32beta, a mammalian homolog of CED-3, is a CrmA-inhibitable
protease that cleaves the death substrate poly(ADP-ribose)
polymerase. Cell. 81:801–809. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Virág L, Robaszkiewicz A, Rodriguez-Vargas
JM and Oliver FJ: Poly(ADP-ribose) signaling in cell death. Mol
Aspects Med. 34:1153–1167. 2013. View Article : Google Scholar : PubMed/NCBI
|