Introduction
Malignant glioma is the most common intracranial
brain tumor, characterized by continuous growth, aggressive
behavior, invasiveness and high genetic heterogeneity. The highly
infiltrative and migrating behavior of glioma leads to diffuse
growth and frequent recurrence of the tumor (1). The mean survival of patients with
glioma is ~14 months after diagnosis, despite aggressive treatment
with neurosurgery, radiotherapy and chemotherapy (2). Therefore, better therapeutic
strategies, such as new molecular targets, are urgently required to
effectively prevent glioma cell invasion.
Recent research revealed the important roles of
microRNA-10b (miR-10b) processing in glioma cell growth, migration
and invasion (3). miR-10b belongs to
the miR-10 family and is located on chromosome 2q31.1, in the
middle of the homeobox D (HOXD) cluster and upstream of HOXD4
(4). miR-10b has been reported to be
highly and specifically expressed in glioblastoma tissues and
glioma cell lines (5,6). High expression of miR-10b has been
associated with high-grade glioma and poor prognosis (7), whereas miR-10b downregulation
suppressed the growth of subcutaneous and intracranial glioblastoma
xenografts (8). In glioma, miR-10b
downregulated several key cell cycle inhibitors and proapoptotic
genes, such as Bim, TFAP2C, p16 and p21, indicating its function in
promoting the cell cycle (9). In
addition, miR-10b has been demonstrated to suppress TP53, PAX6,
NOTCH1 and HOXD10 gene expression, which may coordinately regulate
cancer invasion (10). Our previous
study reported that miR-10b directly targeted Apaf-1 to regulate
glioma cell apoptosis (11).
Although several targets of miR-10b have been identified in glioma,
the exact mechanism underlying the induction of glioma cell
invasion by miR-10b remains unclear. Thus, the current study
attempted to follow up on the role of miR-10b-5p in glioma,
exploring its expression and functions on cell proliferation and
invasion.
Based on sequence alignment with the miRBase
database (www.mirbase.org/), miR-10b may target
human HOXB3. HOXB3 is a transcription factor located on chromosome
17, which acts as a metastasis suppressor gene, and participates in
cell apoptosis, proliferation, migration, invasion and
epithelial-to-mesenchymal transition (12). Chen et al (13) reported that miR-10b targeted HOXB3 in
endometrial cancer, which inhibited apoptosis and promoted cell
proliferation, migration and invasion. Yang et al (14) also found that the upregulation of
HOXB3 inhibited pancreatic cancer cell proliferation, migration and
chemosensitivity. Therefore, it may be hypothesized that HOXB3
plays a functional role in glioma cells.
In the present study, to explore the effect of
miR-10b on glioma cell proliferation and invasion, miR-10b-5p mimic
or inhibitor was transfected into U87 and U251 cells. Reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
analysis was applied to assess the mRNA expression level of HOXB3
in glioma cells. Furthermore, the effect of miR-10b-5p on the
expression of invasion-associated proteins in glioma cells was
investigated using various methods, including western blotting and
zymography. The results of the present study may provide an insight
into the molecular mechanisms underlying the effect of miR-10b-5p
in glioma cells.
Materials and methods
Cell lines and cell culture
The human glioma cell lines U251 (cat. no. TCHu 58)
and U87-MG (glioblastoma of human origin, cat. no. TCHu138; Chinese
Academy of sciences) were obtained from the National Infrastructure
of Cell Line Resource (Shanghai, China) and grown in Dulbecco's
modified Eagle's medium and RPMI-1640 medium (Gibco; Thermo Fisher
Scientific, Inc., Waltham, MA, USA), respectively. Complete medium
was supplemented with 10% fetal bovine serum (FBS), 100 U/ml
penicillin and 100 mg/ml streptomycin (Gibco; Thermo Fisher
Scientific, Inc.). The cell lines were cultured in an incubator at
37°C with a 5% CO2 atmosphere. The cell lines were
identified by the Genetic Testing Biotechnology Corporation
(Suzhou, China) and represented a 94% match with the cell lines of
the DSMZ Reference Database.
Transient transfection
miR-10b-5p mimic, inhibitor and corresponding
negative control were chemically synthesized by RiboBio Co., Ltd.
(Guangzhou, China). The mimic and inhibitor sequences were as
follows: miR-10b-5p mimic, 5′-UACCCUGUAGAACCGAAUUUGUG-3′; and
control mimic, 5′-UUUGUACUACACAAAAGUACUG-3′; miR-10b-5p inhibitor,
5′-UACCCUGUAGAACCGAAUUUGUG-3′, and control inhibitor,
5′-UCACAACCUCCUAGAAAGAGUAGA-3′. Prior to transfection, cells were
plated at 70–80% confluence, and then transfection of
oligonucleotides was performed using the Lipofectamine™ 2000
reagent (Invitrogen; Thermo Fisher Scientific, Inc.). A total of
100 nM mimic, 200 nM inhibitor or corresponding control miRNA was
added to each well. The cells were incubated for 48 h after
transfection and then subjected to various assays.
Cell viability assay
U87 and U251 cells (5×103 cells/well)
were seeded in triplicate into 96-well plates in 100 µl complete
medium. Cells were then transfected with miR-10b-5p mimic,
miR-10b-5p inhibitor or control miRNA. After 48 h of incubation,
cell viability was evaluated using an MTT assay. Approximately 20
µl of 5 mg/ml MTT solution (Thermo Fisher Scientific, Inc.) was
added to each well, and the samples were incubated for 4 h at 37°C.
Subsequently, the supernatant was carefully removed, and 150 µl
DMSO was added to dissolve the cells. The optical density at 570 nm
was measured using a microplate reader (Thermo Fisher Scientific,
Inc.).
Cell cycle analysis
U87 and U251 cells (1×105 cells/well)
were seeded into 24-well plates and allowed to grow for 48 h,
followed by transfection with miR-10b-5p mimic or inhibitor. Cells
were harvested by trypsinization, and then cell pellets were
collected, washed twice with phosphate-buffered saline (PBS) and
fixed with 70% ethanol for 3 h at −20°C. The fixed cells were
washed once with PBS, mixed with an equal volume of Muse cell cycle
reagent (Merck KGaA, Darmstadt, Germany) and then incubated for 30
min at room temperature in the dark. Then cells were analyzed by
flow cytometry. The distribution of cells at each cell cycle phase
was determined by Muse® Cell Analyzer using the
Muse® Cell Cycle Assay kit (Merck KGaA), analyzing a
mean number of 10,000 cells.
Wound healing assay
U87 and U251 cells (1×106 cells/well)
were seeded into 6-well plates. After reaching 70% confluence, the
cells were transfected with miR-10b-5p mimic or inhibitor, and
incubated for 48 h. Using a 200-µl sterile pipette tip, three
scratches were created in each well (15). Next, the cells were washed twice with
PBS and incubated in medium supplemented with 2% FBS for 24 h. In
order to assess the cell migration, images of the wells were
captured at 0 and 24 h in three randomly selected microscopic
fields (Olympus Corporation, Tokyo, Japan). The distance over which
the cells had migrated into the denuded area was determined by
measuring the wound width at 0 and 24 h using ImageJ software
(National Institutes of Health, Bethesda, MD, USA) to calculate the
healing rate.
In vitro migration and invasion
assays
Cell migration and invasion were assayed using 8-µm
pore polycarbonate membrane Transwell chambers (Corning Costar,
Corning, NY, USA). Briefly, U87 and U251 cells seeded in 6-well
plates (1×105 cells/well) transfected with miR-10b-5p
mimic or inhibitor for 48 h were transferred onto the upper
Transwell chamber that was filled with 200 µl serum-free medium for
24 h. Complete medium (600 µl) was added to the lower chamber of
each well. For the Transwell invasion assay, the plate inserts used
were precoated with Matrigel (BD Biosciences, San Jose, CA, USA).
After 24 h, the cells were washed three times with PBS, and
non-migratory or non-invasive cells in the upper side of the
chambers were removed with cotton swabs. The cells that had
migrated or invaded were fixed in 4% paraformaldehyde and then
stained with 0.5% crystal violet solution for 30 min. Following
washing twice with PBS, images of the cells were obtained at a
magnification of ×200. Finally, the chambers were eluted by 33%
acetic acid, and the optical density at 570 nm was measured.
Gelatin zymography
The activity of matrix metalloproteinase 2 (MMP2) in
the miR-10b-5p mimic or inhibitor groups was analyzed in order to
assess changes in the invasive ability of glioma cells. Briefly,
supernatant fractions of cells cultured in serum-free medium for 24
h were concentrated using 10 kDa Vivaspin 500 (Sartorius AG,
Göttingen, Germany). Equal amounts of proteins were separated via
8% SDS-PAGE with 0.1% gelatin under non-reducing conditions.
Following the completion of electrophoresis, the gels were washed
twice with renaturing buffer (containing 50 mM Tris-HCl, pH 7.6, 5
mM CaCl2, 1 µM ZnCl2 and 2.5% Triton X-100)
within 30 min at room temperature. The gels were subsequently
incubated in developing buffer (containing 50 mM Tris-HCl, pH 7.6,
5 mM CaCl2, 1 µM ZnCl2, 200 mM NaCl and 0.02%
Brij-35) at 37°C for 24 h, washed in water, stained with Coomassie
Brilliant Blue R-250 for 1 h, and then destained with 40% methanol
and 5% acetic acid until the bands were visible. Band intensities
were quantitatively determined by ImageJ software.
miR-10b-5p target gene
identification
Using bioinformatics software (TargetScan7.1;
www.targetscan.org/vert_71/; and
miRanda database; www.mirbase.org/), gene binding sites between
miR-10b-5p and HOXB3 were detected.
RT-qPCR analysis
Total RNA was isolated from the cultured cells using
TRIzol reagent (Invitrogen; Thermo Fisher Scientific, Inc.). To
quantify the relative content of miR-10b-5p and HOXB3 mRNA, ~1 µg
of RNA from each sample was reverse-transcribed using the Advantage
RT-for-PCR kit (Takara Biotechnology Co., Ltd., Dalian, China). RT
was conducted with oligo d(T)18 primers (Takara Biotechnology Co.,
Ltd.) and the miR-10b-5p RT primer,
5′-CTCAACTGGTGTCGTGGAGTCGGCAATTCAGTTGAGCACAAATT-3 (Genscript
Biotech, Piscataway, NJ, USA). Next, qPCR was run using SYBR Green
PCR Master Mix (Invitrogen; Thermo Fisher Scientific, Inc.) and
specific primers for miR-10b-5p, U6, HOXB3 and β-actin on an iQ5
Sequence Detector (Bio-Rad Laboratories, Inc., Hercules, CA, USA).
The primer sequences were as follows: miR-10b-5p,
5′-GGGTACCCTGTAGAACCG-3′ (forward) and 5′-AACTGGTGTCGTGGAGTCGGC-3′
(reverse); U6, 5′-CTCGCTTCGGCAGCACA-3′ (forward) and
5′-AACGCTTCACGAATTTGCGT-3′ (reverse); HOXB3,
5′-ACCTTTCCCATCACCCTT-3′ (forward) and 5′-CGCTTCTTGGATTCTACC-3′
(reverse); β-actin, 5′-ACACTGTGCCCATCTACG-3′ (forward) and
5′-TGTCACGCACGATTTCC-3′ (reverse). Subsequently, the comparative
2−ΔΔCq method (16) was
used to quantify the gene expression levels. The relative
expression of miR-10b-5p and HOXB3 mRNA was normalized to that of
U6 and β-actin, respectively.
Western blotting
Radioimmunoprecipitation assay lysis buffer (Boster
Biological Technology, Ltd., Wuhan, China) with freshly added
protease inhibitor cocktail and phenylmethylsulfonyl fluoride was
used to isolate the total protein from the cells. The protein
concentration was then quantified using a BCA Protein Assay kit
(Boster Biological Technology, Ltd.). Next, 30 µg protein was
loaded onto 10% SDS-PAGE and transferred onto polyvinylidene
difluoride membranes (Merck Millipore, Billerica, MA, USA).
Following blocking with 5% (w/v) milk in tris-buffered saline
containing 0.1% (v/v) Tween 20 (TBST) for 1 h at room temperature,
the blots were probed with the following primary antibodies
overnight at 4°C: HOXB3 (cat. no. abs139097-100 µg; Absin
Bioscience, Inc., Shanghai, China; dilution, 1:500), Ras homolog
family member C (RhoC; cat. no. 3430S; Cell Signaling Technology
Inc., Danvers, MA, USA; dilution, 1:1,000), high mobility group box
1 (HMGB1; cat. no. ab92310-100 µl; Abcam, Cambridge, MA, USA;
dilution, 1:1,000) and β-actin (cat. no. 4970S Cell Signaling
Technology, Inc.; dilution, 1:1,000). Subsequently, the samples
were incubated with the appropriate secondary antibodies
(horseradish peroxidase-conjugated anti-rabbit or anti-mouse IgG;
cat. no. BA1054 and BA1050; Boster Biological Technology, Ltd.,
China; 1:3,000) for 1 h at room temperature. The bands were
visualized with ECL substrate (Thermo Fisher Scientific, Inc.)
using the Syngene G:BOX system (Gene Company, Ltd., Hong Kong,
China). Each band was quantified by densitometry using Gen Tools
software program (Gene Company, Ltd.), with β-actin used for
normalization.
Statistical analysis
Statistical analysis for all the experiments was
performed using Student's two-tailed t-test for comparison of two
independent groups (experimental group and corresponding control
group) with GraphPad Prism 5 software (GraphPad Software, Inc., La
Jolla, CA, USA). Data from three independent experiments are
presented as the mean ± standard error of the mean. A P-value of
<0.05 was considered to denote a statistically significant
difference.
Results
miR-10b-5p downregulation inhibits the
proliferation of glioma cells
As reported previously, miR-10b is significantly
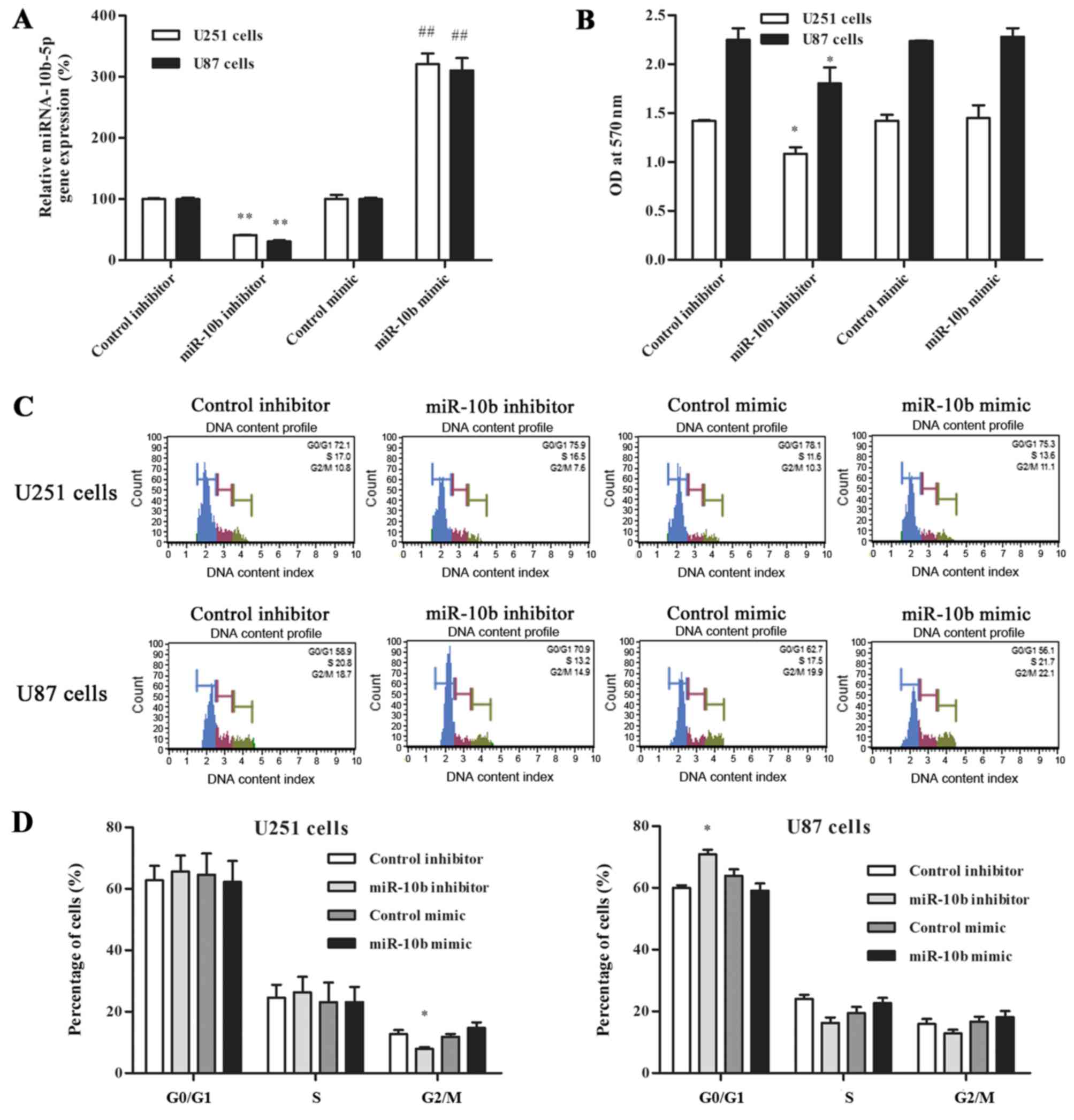
overexpressed in glioma cell lines (6). In the present study, RT-qPCR analysis
was performed to examining the efficiency of mimic/inhibitor
transfection and cell viability in glioma cells (Fig. 1A). Compared with the corresponding
control group, the expression of miR-10b-5p was significantly
increased in the mimic group, while that of the inhibitor group was
decreased. A cell viability assay was also performed to investigate
the effect of miR-10b-5p expression on the proliferation of glioma
cells. In the miR-10b-5p inhibitor group, the cell viability was
significantly lower at 48 h after transfection, reduced to 76.2±4.7
and 80.2±7.1% of the control inhibitor group in U251 and U87 cells,
respectively (P<0.05; Fig. 1B).
By contrast, no statistically significant difference in cell
viability was detected between the miR-10b-5p mimic and control
mimic groups.
To further explore how miR-10b-5p affects the growth
of glioma cells, the cell cycle distribution of glioma cells
transfected with miR-10b-5p mimic or inhibitor was analyzed. The
data shown in Fig. 1C and D
demonstrated that the fraction of U251 cells in G2/M phase was
significantly decreased following inhibitor transfection (7.9±0.5%)
as compared with that in the control group (12.7±1.3%; P<0.05).
By contrast, U87 cells accumulated in the
G0/G1 phase of the cell cycle following
transfection with miR-10b-5p inhibitor (70.9±1.4%), as compared
with the corresponding control group (60±0.8%; P<0.05). These
data clearly indicated that miR-10b-5p downregulation affects cell
cycle progression mainly by prolonging the G2/M or
G0/G1 phase. By contrast, transfection with
miR-10b-5p mimic exerted no marked effect on the cell cycle
distribution (Fig. 1C and D).
miR-10b-5p upregulation promotes
glioma cell migration and invasion
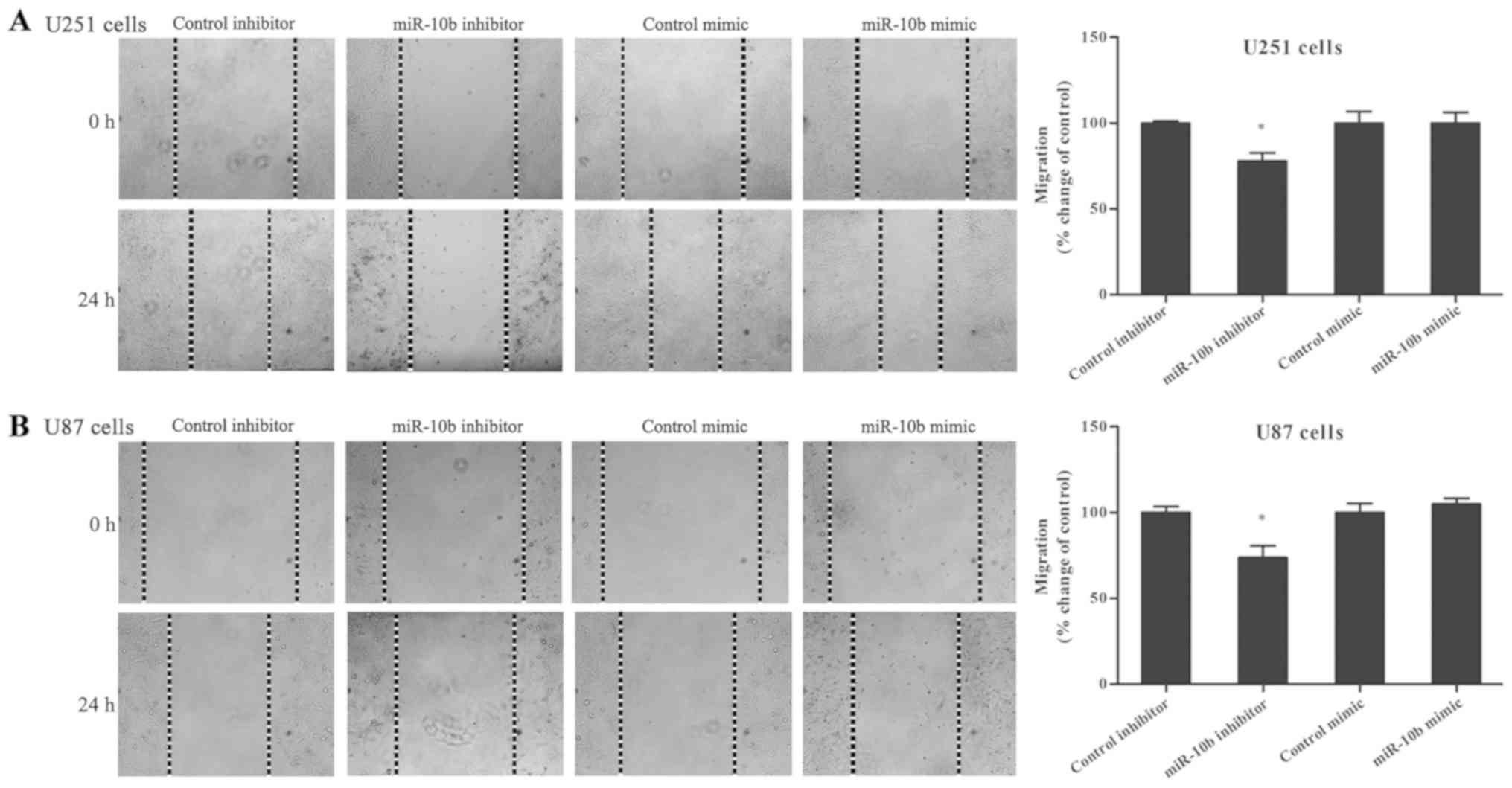
The role of miR-10b-5p on glioma cell migration was
further investigated. Glioma cells transfected with the miR-10b-5p
mimic or inhibitor were used in the wound healing assay. The
results indicated that miR-10b-5p inhibition markedly suppressed
the migration ability of U251 and U87 cells (Fig. 2A and B), with a reduction of
22.1±4.7% observed in U251 cells and 26.2±6.7% in U87 cells
compared with the control inhibitor (P<0.05). By contrast, the
migration ability in the miR-10b-5p mimic-transfected group was not
evidently affected.
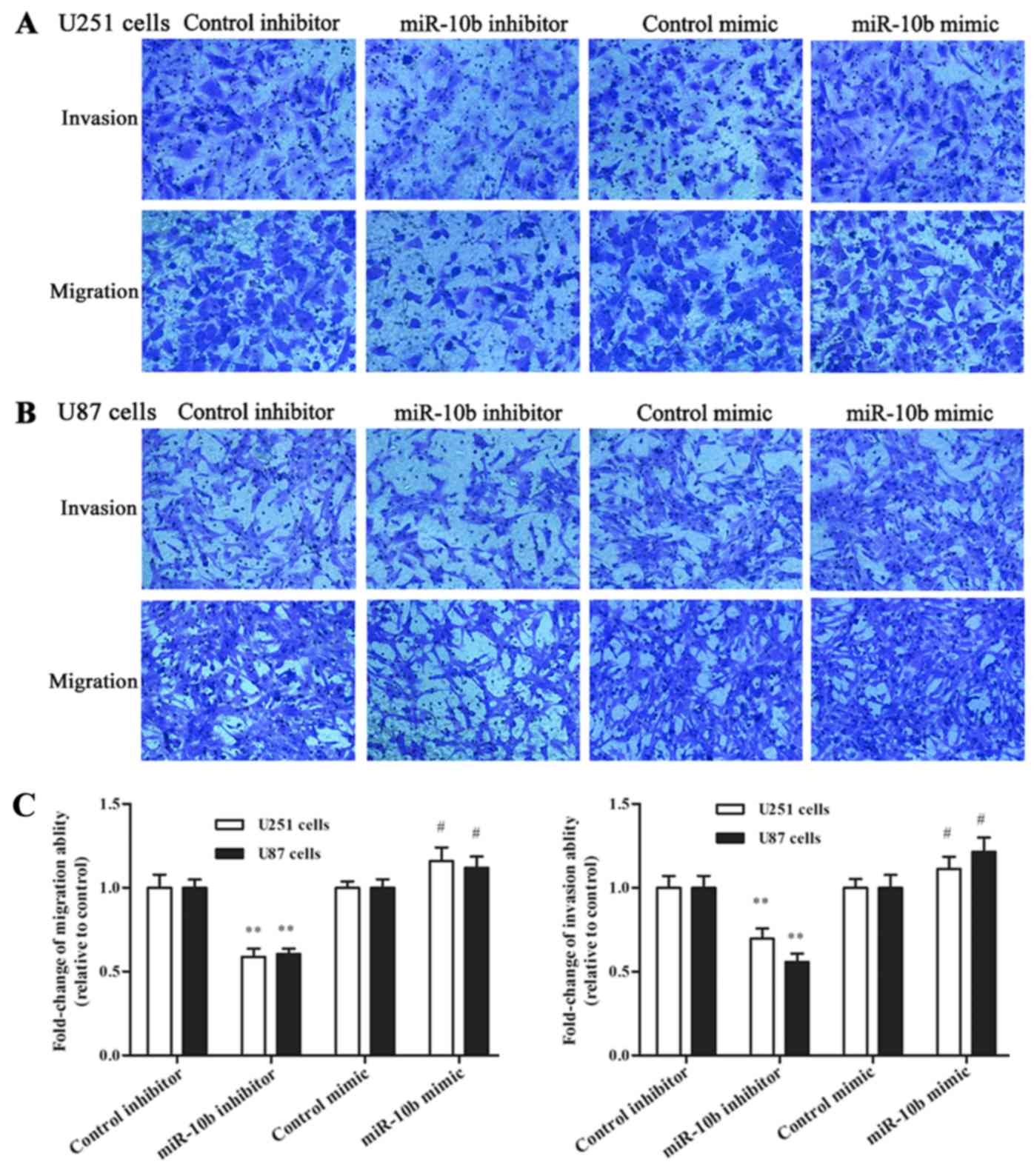
In addition, Transwell assay was performed to test
the effect of miR-10b-5p on cell migration and invasion. The
experiments demonstrated that the number of glioma cells migrating
through non-coated and Matrigel-coated Transwell membranes was
significantly reduced following inhibition of miR-10b-5p
expression. As presented in Fig.
3A-C, a 41.1±5.6% reduction in the migration of U251 cells and
a 39.4±3.4% reduction in U87 cells were observed compared with the
control inhibitor group (P<0.01). Similarly, miR-10b-5p
inhibitor led to a 30.3±3.3% reduction in invasion in U251 cells
and a 44.2±2.0% reduction in U87 cells, as compared with the
control inhibitor group (P<0.01). By contrast, miR-10b-5p mimic
transfection significantly increased the migration and invasion of
glioma cells in comparison with those in the control mimic
group.
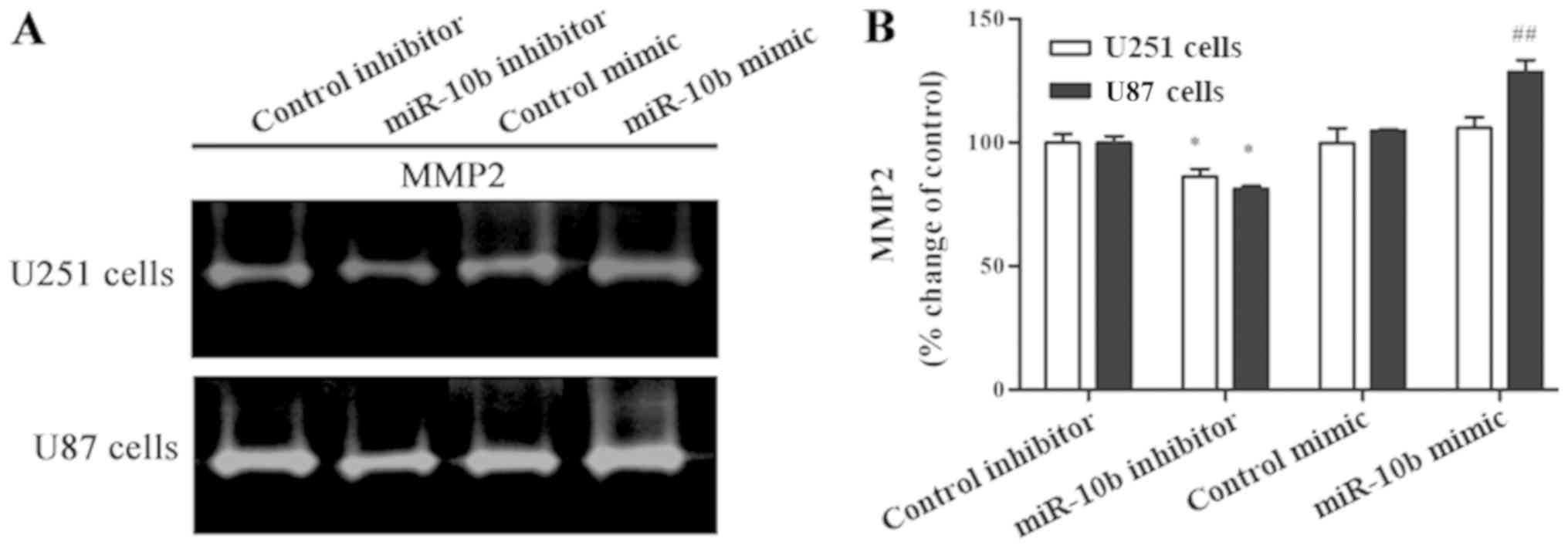
Overexpression of MMP2 and its extracellular
activation, as well as its pro-invasion function, have been widely
reported in glioma (17). To
investigate the effect of miR-10b-5p on the gelatinase activity of
MMP2 in glioma cells, gelatin zymography was performed. As shown by
zymography analysis (Fig. 4A and B),
MMP2 secretion was significantly reduced in the miR-10b-5p
inhibitor group when compared with the control inhibitor group in
glioma cells (P<0.05). In the miR-10b-5p mimic group, a marked
increase in MMP2 secretion was only observed in U87 cells, but not
in U251 cells. Collectively, the results indicate that
downregulated miR-10b-5p inhibits the invasion of glioma cells by
downregulating the expression of MMP-2.
miR-10b-5p modulates HOXB3 and
invasion-associated gene expression in glioma cells
A previous study identified HOXB3 as a target gene
of miR-10b (14). In the current
study, the TargetScan prediction tool was used, and the analysis
revealed that HOXB3 contained potential binding sites for
miR-10b-5p (Fig. 5A). To identify
whether miR-10b-5p can regulate HOXB3 expression in glioma cell
lines, the mRNA level of HOXB3 in miR-10b-5p mimic- or
inhibitor-transfected glioma cells was first examined. In U251
cells, miR-10b-5p overexpression led to a marked decrease in HOXB3
mRNA level, while miR-10b-5p downregulation significantly increased
HOXB3 mRNA (Fig. 5B). Western
blotting further revealed that the HOXB3 protein expression level
was significantly downregulated following miR-10b-5p mimic
transfection in both U251 and U87 cells (P<0.05; Fig. 5C and D). By contrast, the miR-10b-5p
inhibitor transfection effectively increased the HOXB3 protein
expression level (Fig. 5C and D).
These results indicated that the HOXB3 gene may be a target of
miR-10b-5p.
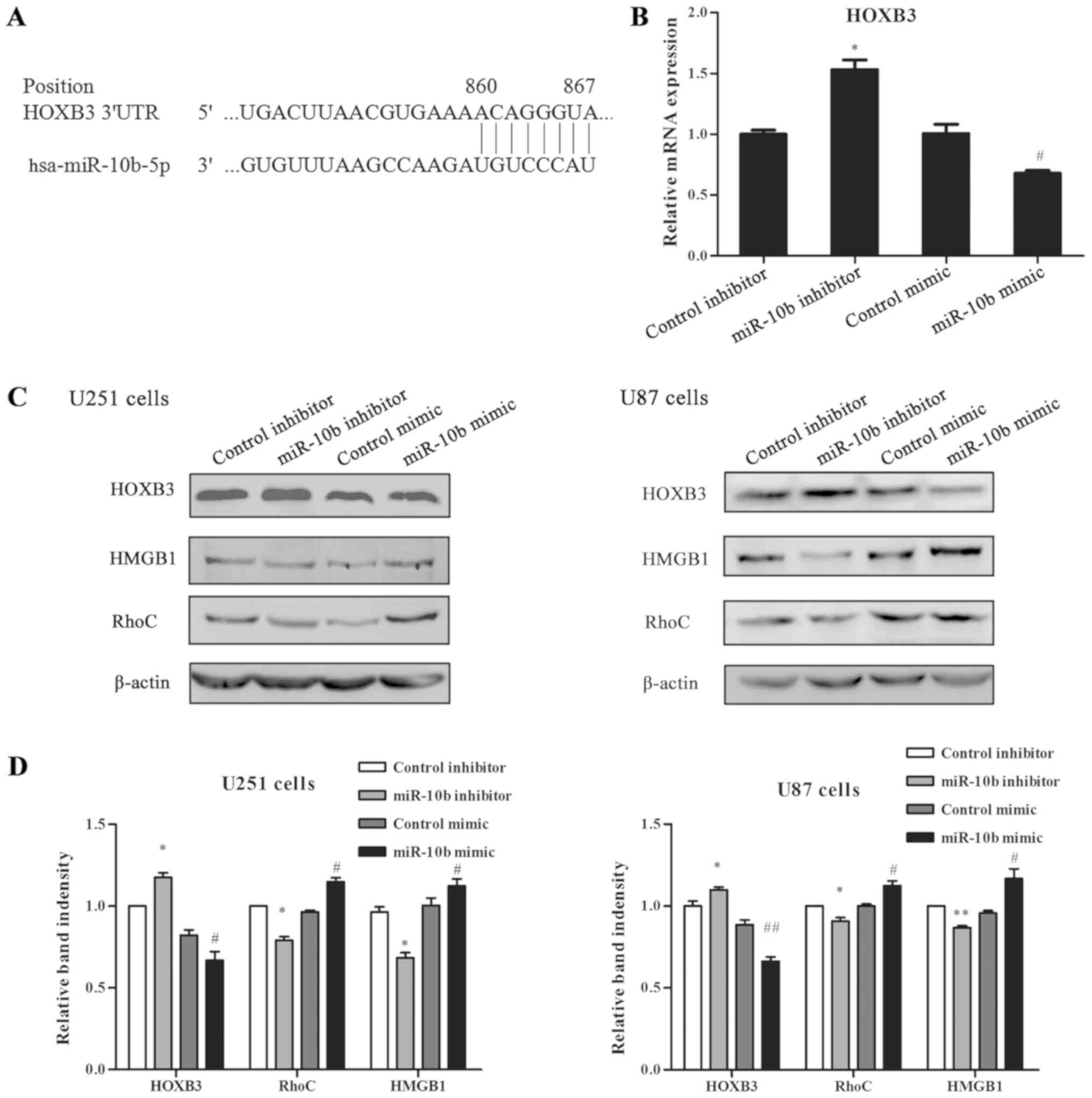 | Figure 5.Effect of miR-10b-5p on the expression
levels of HOXB3, HMGB1 and RhoC in glioma cells. (A) miR-10b-5p
binding sites in the 3′UTR of HOXB3 mRNA, predicted by TargetScan.
(B) Reverse transcription-quantitative polymerase chain reaction
analysis for HOXB3 in glioma cells transfected with miR-10b-5p
mimic, inhibitor or control, indicating that miR-10b-5p regulated
the mRNA level of HOXB3. (C) Western blots and (D) quantified
results of the expression levels of HOXB3, HMGB1 and RhoC in human
glioma cells treated with miR-10b-5p mimic, miR-10b-5p inhibitor or
control. The expression of β-actin served as an endogenous control.
miR-10b-5p inhibitor significantly enhanced HOXB3 protein
expression, and decreased the downstream RhoC and HMGB1 protein
expression levels in U251 and U87 cells. miR-10b-5p mimic exerted
the opposite effect. Data were analyzed by Student's t-test.
*P<0.05 and **P<0.01 vs. control inhibitor group;
#P<0.05 and ##P<0.01 vs. control mimic
group. miR, microRNA; 3′UTR, 3′-untranslated region; HOXB3,
homeobox B3; RhoC, Ras homolog family member C; HMGB1, high
mobility group box 1. |
It has been reported that RhoC, a member of the Ras
homolog gene family, is critical for invasion and metastasis, and
is specifically upregulated in glioma (18). In order to investigate whether RhoC
is regulated by miR-10b-5p, its protein expression was detected
with western blotting. Consistently, the results demonstrated that
RhoC was significantly downregulated by miR-10b-5p in both U251 and
U87 cells (P<0.05; Fig. 5C and
D). Furthermore, the protein expression of HMGB1 was evaluated,
which has been reported to be involved in the malignant phenotype
of glioma cells (19). As shown in
Fig. 5C and D, miR-10b-5p inhibition
significantly reduced the protein expression of HMGB1, while
upregulation of this miRNA markedly increased HMGB1 protein
(P<0.05).
Discussion
Several microRNAs have recently been identified as
either promoters or suppressors of glioma cell invasion and
metastasis. miR-10b is known to be induced by the
metastasis-promoting transcription factor Twist (20). The present study data from MTT and
cell cycle assays demonstrated that downregulated miR-10b-5p
inhibited U251 and U87 cell proliferation, and caused cell cycle
arrest. These findings are consistent with those of a previous
study by Gabriely et al (6),
which reported that miR-10b inhibition led to reduction of glioma
cell growth and caused cell cycle arrest at G2 phase in U251 cells,
whereas it induced accumulation of U87 cells in the G1 phase.
Further wound healing, Transwell and gelatin zymography assays
conducted in the present study demonstrated that miR-10b-5p
inhibition prevented glioma cell migration and invasion, whereas
miR-10b-5p mimic promoted the migration and invasion of both U87
and U251 cells. Previous research revealed that U87 cells lost
their invasion potential upon miR-10b inhibition, as well as
exhibited reduced MMP2 protein expression, while the invasive
ability of U251 cells was enhanced by miR-10b mimic transfection
(21,22). Taken together, the current study
results combined with previous data suggest an important role for
miR-10b-5p in the regulation of glioma cell growth.
HOX genes are known to be master genes encoding a
subset of highly conserved transcription factors that serve key
roles in the morphogenesis of tissues and organ development in
embryos, and their dysregulation is implicated in various types of
cancer, including glioma (23). As a
member of the HOX gene family, HOXB3 is known as a regulator of
gene transcription during development. In fact, HOXB3 has been
reported to serve critical roles in cancer initiation and
progression, which was confirmed in breast (24), prostate (25), pancreatic (14) and endometrial cancer (13). However, the role of HOXB3 in glioma
remains unclear. The findings of the present study indicated a
significant association between HOXB3 and miR-10b-5p. HOXB3
expression was upregulated at the mRNA and protein levels in glioma
cells with inhibition of miR-10b-5p. These results indicated that
HOXB3 may be a downstream target gene of miR-10b-5p in glioma.
In the present study, RhoC expression was regulated
by transfection with miR-10b-5p mimic and inhibitor, and its
alteration was inversely associated with that of HOXB3 expression.
Overexpression of miR-10b-5p upregulated RhoC expression and
downregulated HOXB3 expression. Previous correlative studies
indicated that miR-10b targeted HOXD10 in glioma cells, and HOXD10
was reported to suppress the expression of genes involved in tumor
metastasis, including RhoC (9,22).
However, it has yet to be determined whether HOXD10 signaling
exerts a cross-link effect on HOXB3 regulation in glioma cells,
since miR-10b functions as a powerful oncogenic miRNA directly
targeting different genes. Future experiments may help elucidate
the different miR-10b-associated molecular pathways regulating
glioma development.
HMGB1, as a member of the HMGB family, has been
found to be overexpressed in various tumors, including human glioma
(19). It has been reported that
HMGB1 was significantly upregulated in glioma cells, whereas
inhibition of HMGB1 suppressed the proliferation and migration of
glioma cells, and promoted apoptosis (19,26).
HMGB1 participates in the crosstalk with multiple miRNAs, and
promotes metastasis and invasion in glioma. HMGB1 is target gene of
miR-218 in glioma cells, which directly suppresses the HMGB1-RAGE
pathway (27). miR-129-2 targets
HMGB1, thereby suppressing glioma cell growth, migration and
invasion, and promoting cell apoptosis (28). In the present study, the regulation
of HMGB1 protein expression by miR-10b-5p was similar with that of
RhoC expression, with inhibition of miR-10b-5p triggering a
significant reduction in the protein expression of HMGB1. It has
been reported that HMGB1 induces glioma cell growth and migration
through its intracellular signaling pathways, including nuclear
factor-κB, mitogen-activated protein kinase and extracellular
signal-regulated kinase (29,30),
suggesting that the mechanism of action of miR-10b may be involved
in glioma invasion. Further studies are needed to elucidate the
underlying molecular mechanism. To the best of our knowledge, the
data reported in the present study are the first to demonstrate an
association of the expression level of miR-10b-5p with HOXB3 and
HMGB1 in glioma.
A limitation of the present study is that the
results illustrated the effect of miR-10b-5p on the invasion
ability of glioma cells in vitro. In the future, animal
models will also be applied to our study to comprehensively
evaluate the effect of miR-10b-5p in the glioma. Furthermore,
although the current data demonstrated that there was an inverse
correlation between expression of the miR-10b-5p and that of HOXB3
mRNA and protein, it is important to consolidate this finding using
dual-luciferase reporter assay, as well as rescue experiments. It
is thus recommended that the exact correlation between miR-10b and
HOXB3 in the invasion of glioma cells is investigated in future
studies.
In conclusion, miR-10b-5p mimic and inhibitor were
demonstrated to serve key roles in the regulation of U87 and U251
cell proliferation and invasion capacities in the present study.
Transfection with miR-10b-5p inhibitor suppressed glioma cell
growth, while miR-10b-5p mimic promoted the migration and invasion
of glioma cells. Downregulation of miR-10b-5p increased the
expression of HOXB3, resulting in a marked decrease in HMGB1 and
RhoC protein levels. These results expanded the list of
miR-10b-moderated genes to include HOXB3 in glioma. In addition,
the findings provided an insight into the possible mechanism
underlying the regulation of glioma by miR-10b-5p, indicating that
miR-10b-5p may represent a promising target in glioma
treatment.
Acknowledgements
Not applicable.
Funding
This study was supported by the Natural Science
Foundation of China (grant no. 81302203).
Availability of data and materials
All data generated or analyzed during the present
study are included in this published article.
Authors' contributions
LL and QR performed the research. QR designed the
study. LL and QR analyzed the data. LL wrote the paper. XT provided
technical assistance. CL and QX revised the manuscript. All authors
have read and approved the final version of this manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Brandes AA, Tosoni A, Franceschi E, Sotti
G, Frezza G, Amistà P, Morandi L, Spagnolli F and Ermani M:
Recurrence pattern after temozolomide concomitant with and adjuvant
to radiotherapy in newly diagnosed patients with glioblastoma:
Correlation with MGMT promoter methylation status. J Clin Oncol.
27:1275–1279. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ru Q, Tian X, Wu YX, Wu RH, Pi MS and Li
CY: Voltage-gated and ATP-sensitive K+ channels are associated with
cell proliferation and tumorigenesis of human glioma. Oncol Rep.
31:842–848. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Mathupala SP, Mittal S, Guthikonda M and
Sloan AE: MicroRNA and brain tumors: A cause and a cure? DNA Cell
Biol. 26:301–310. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ma L, Teruya-Feldstein J and Weinberg RA:
Tumour invasion and metastasis initiated by microRNA-10b in breast
cancer. Nature. 449:682–688. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ciafrè SA, Galardi S, Mangiola A, Ferracin
M, Liu CG, Sabatino G, Negrini M, Maira G, Croce CM and Farace MG:
Extensive modulation of a set of microRNAs in primary glioblastoma.
Biochem Biophys Res Commun. 334:1351–1358. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Gabriely G, Yi M, Narayan RS, Niers JM,
Wurdinger T, Imitola J, Ligon KL, Kesari S, Esau C, Stephens RM, et
al: Human glioma growth is controlled by microRNA-10b. Cancer Res.
71:3563–3572. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ji Y, Wei Y, Wang J, Gong K, Zhang Y and
Zuo H: Correlation of microRNA-10b upregulation and poor prognosis
in human gliomas. Tumour Biol. 36:6249–6254. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Guessous F, Alvarado-Velez M,
Marcinkiewicz L, Zhang Y, Kim J, Heister S, Kefas B, Godlewski J,
Schiff D, Purow B and Abounader R: Oncogenic effects of miR-10b in
glioblastoma stem cells. J Neurooncol. 112:153–163. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Sasayama T, Nishihara M, Kondoh T, Hosoda
K and Kohmura E: MicroRNA-10b is overexpressed in malignant glioma
and associated with tumor invasive factors, uPR and RhoC. Int J
Cancer. 125:1407–1413. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lin J, Teo S, Lam DH, Jeyaseelan K and
Wang S: MicroRNA-10b pleiotropically regulates invasion,
angiogenicity and apoptosis of tumor cells resembling mesenchymal
subtype of glioblastoma multiforme. Cell Death Dis. 3:e3982012.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ru Q, Li WL, Xiong Q, Chen L, Tian X and
Li CY: Voltage-gated potassium channel blocker 4-aminopyridine
induces glioma cell apoptosis by reducing expression of
microRNA-10b-5p. Mol Biol Cell. 29:1125–1136. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Weiss FU, Marques IJ, Woltering JM,
Vlecken DH, Aghdassi A, Partecke LI, Heidecke CD, Lerch MM and
Bagowski CP: Retinoic acid receptor antagonists inhibit miR-10a
expression and block metastatic behavior of pancreatic cancer.
Gastroenterology. 137:2136–2145. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Chen H, Fan Y, Xu W, Chen J, Xu C, Wei X,
Fang D and Feng Y: miR-10b inhibits apoptosis and promotes
proliferation and invasion of endometrial cancer cells via
targeting HOXB3. Cancer Biother Radiopharm. 31:225–231. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Yang D, Yan R and Zhang X, Zhu Z, Wang C,
Liang C and Zhang X: Deregulation of MicroRNA-375 inhibits cancer
proliferation migration and chemosensitivity in pancreatic cancer
through the association of HOXB3. Am J Transl Res. 8:1551–1559.
2016.PubMed/NCBI
|
|
15
|
Hussain R, Umer HM, Björkqvist M and
Roomans GM: ENaC, iNOS, mucins, and wound healing in cystic
fibrosis airway epithelial and submucosal cells. Cell Biol Int Rep.
21:25–38. 2014.
|
|
16
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Mayes DA, Hu Y, Teng Y, Siegel E, Wu X,
Panda K, Tan F, Yung WK and Zhou YH: PAX6 suppresses the
invasiveness of glioblastoma cells and the expression of the matrix
metalloproteinase-2 gene. Cancer Res. 66:9809–9817. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Huang H, Okamoto Y, Yokoo H, Heppner FL,
Vital A, Fevre-Montange M, Jouvet A, Yonekawa Y, Lazaridis EN,
Kleihues P and Ohgaki H: Gene expression profiling and subgroup
identification of oligodendrogliomas. Oncogene. 23:6012–6022. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Angelopoulou E, Piperi C, Adamopoulos C
and Papavassiliou AG: Pivotal role of high-mobility group box 1
(HMGB1) signaling pathways in glioma development and progression. J
Mol Med (Berl). 94:867–874. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Gabriely G, Teplyuk NM and Krichevsky AM:
Context effect: microRNA-10b in cancer cell proliferation, spread
and death. Autophagy. 7:1384–1386. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Sun L, Yan W, Wang Y, Sun G, Luo H, Zhang
J, Wang X, You Y, Yang Z and Liu N: MicroRNA-10b induces glioma
cell invasion by modulating MMP-14 and uPAR expression via HOXD10.
Brain Res. 1389:9–18. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Dong CG, Wu WK, Feng SY, Wang XJ, Shao JF
and Qiao J: Co-inhibition of microRNA-10b and microRNA-21 exerts
synergistic inhibition on the proliferation and invasion of human
glioma cells. Int J Oncol. 41:1005–1012. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Shah N and Sukumar S: The Hox genes and
their roles in oncogenesis. Nat Rev Cancer. 10:361–371. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Fu H, Fu L, Xie C, Zuo WS, Liu YS, Zheng
MZ and Yu JM: miR-375 inhibits cancer stem cell phenotype and
tamoxifen resistance by degrading HOXB3 in human ER-positive breast
cancer. Oncol Rep. 37:1093–1099. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Chen J, Zhu S, Jiang N, Shang Z, Quan C
and Niu Y: HoxB3 promotes prostate cancer cell progression by
transactivating CDCA3. Cancer Lett. 330:217–224. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhang J, Liu C and Hou R: Knockdown of
HMGB1 improves apoptosis and suppresses proliferation and invasion
of glioma cells. Chin J Cancer Res. 26:658–668. 2014.PubMed/NCBI
|
|
27
|
Gu J, Xu R, Li Y, Zhang J and Wang S:
MicroRNA-218 modulates activities of glioma cells by targeting
HMGB1. Am J Transl Res. 8:3780–3790. 2016.PubMed/NCBI
|
|
28
|
Yang Y, Huang JQ, Zhang X and Shen LF:
MiR-129-2 functions as a tumor suppressor in glioma cells by
targeting HMGB1 and is down-regulated by DNA methylation. Mol Cell
Biochem. 404:229–239. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Fiuza C, Bustin M, Talwar S, Tropea M,
Gerstenberger E, Shelhamer JH and Suffredini AF:
Inflammation-promoting activity of HMGB1 on human microvascular
endothelial cells. Blood. 101:2652–2660. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Treutiger CJ, Mullins GE, Johansson AS,
Rouhiainen A, Rauvala HM, Erlandsson-Harris H, Andersson U, Yang H,
Tracey KJ, Andersson J and Palmblad JE: High mobility group 1 B-box
mediates activation of human endothelium. J Intern Med.
254:375–385. 2003. View Article : Google Scholar : PubMed/NCBI
|