Introduction
Human airways are subjected to constant exposure to
Aspergillus fumigatus, resulting in the inhalation of
several hundred spores per day (1).
However, components of the innate immune system, particularly
macrophages, efficiently remove A. fumigatus once the spores
enter alveoli within the lungs, therefore, adverse effects are rare
(2–4). In immunocompromised individuals, A.
fumigatus has been shown to be a causative agent of multiple
forms of aspergillosis, including saprophytic aspergillosis,
allergic aspergillosis and invasive aspergillosis, a
life-threatening systemic fungal infection (5–7). In
addition, A. fumigatus is currently the most prevalent
airborne fungal pathogen, responsible for ~90% of systemic
infections (8–10).
The inhalation of A. fumigatus spores
initiates an antifungal immune response by pattern recognition
receptors (PRRs). In particular, the toll-like receptors (TLRs) are
a class of PRRs that recognize pathogen-associated molecular
patterns and activate the innate immune response to eliminate
invading pathogens and mediate the adaptive immune response. As a
member of the TLR family, TLR2 serves an important role in
recognizing various Aspergillus cell wall components and
modulating host defense responses, including autophagy (11–13).
Furthermore, TLR2 is critical to activating autophagy against fungi
in macrophages (14).
Autophagy maintains cellular homeostasis through
targeted degradation. Multiple roles for autophagy have been
reported in the inflammatory response and the defense against
infections; a recent study suggested an important role in the
clearance of bacterial, viral and parasitic pathogens (15). Regarding the response to fungal
infection, autophagy has been linked to several processes important
to immunity, including pathogen recognition, phagocytosis,
microbial killing, cytokine release, antigen presentation and
inflammation regulation. Several proteins are involved in
autophagy, including light chain 3 (LC3), Beclin-1 and
autophagy-related protein 5 (ATG5) (16–18).
Autophagy can also be regulated by microRNAs
(miRNAs), a class of endogenous short, non-coding RNAs involved in
the regulation of gene expression (19). Our previous study demonstrated that
miR-344b-1-3p is an effective direct modulator of TLR2 (20). However, any correlation between the
levels of miRNA-344b-1-3p and autophagy remain to be
elucidated.
Therefore, in the present study, the expression of
miRNA-344a-1-3p was observed during A. fumigatus infection,
and its role and potential mechanism in regulating
infection-induced autophagy by macrophages was investigated to
provide a better understanding of host defense mechanisms following
Aspergillus infection.
Materials and methods
Cell culture
A rat alveolar macrophage cell line, NR8383, was
purchased from the Shanghai Institute of Biochemistry and Cell
Biology. The cells were maintained in Ham's F-12K medium
(Sigma-Aldrich; Merck KGaA) containing 15% fetal bovine serum
(Gibco; Thermo Fisher Scientific, Inc.) at 37°C and 5%
CO2.
Microbes
A. fumigatus was prepared using the air from
a moisture box and cultured on 1.5% malt extract agar plates
(Biokar Diagnostics; Beuvais, France) placed in the dark at 25°C
for 7 days. Thee spores were harvested using a sterile loop, which
were then suspended in Hank's balanced salt solution containing
0.0001% Triton X-100. The spores were counted under a light
microscope using a Bürker chamber to calculate their concentration
and their strain was identified using an identification service
(Central Bureau of Schimmelcultures, Utrecht, Netherlands). The
collected spores were stored at −20°C until use.
Exposure to A. fumigatus
The NR8383 cells were seeded in triplicate into
6-well plates (5×104 cells per well) and incubated at
37°C in an atmosphere containing 5% CO2. Following
overnight culture, the cells were exposed to A. fumigatus
(107 spores per well) for the indicated durations (0,
30, 45, 60, 90 or 120 min) at 37°C. Following treatment, the cells
were harvested, following which total RNA was extracted and stored
at −80°C until examination.
Cell transfection
The miR-344b-1-3p mimics, negative control (NC), and
inhibitor were purchased from Guangzhou RiboBio Co., Ltd.
(Guangzhou, China). Lipofectamine 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.) was used for transfection, according to the
manufacturer's protocol. The NR8383 cells were seeded into 24-well
plates (1×104 cells per well) and transfected with the
miR-344b-1-3p mimics, NC or inhibitor to a final concentration of
25 nM. Tlr2 cDNA was cloned into the BamHI and
AscI sites of the pLenti6/V5-DEST vector (Invitrogen; Thermo
Fisher Scientific, Inc.). Then, 2 µl solution was added into 100 µl
competence XL-BLUE Escherichia coli (Invitrogen; Thermo
Fisher Scientific, Inc.) and absorbed for 15 min on ice. After
being placed in a water bath for 50 sec at 42°C and rapidly cooled
for 2 min on ice, the cell solution was pipetted into 1 ml LB
culture medium (Merck KGaA), oscillated for 1 h at 37°C and 180 rpm
and then centrifuged for 3 min 2,012.4 × g and 37°C. The pellet was
mixed with 200 ml LB culture medium, coated onto the LB culture
plates containing 100 µg/ml ampicillin and cultured overnight at
37°C. A total of 10 clones were picked out and inoculated in 3 ml
LB culture medium. After culturing overnight at 37°C on a shaker at
250 rpm, the vectors were extracted and then verified by nucleotide
sequencing by GenScript Corporation (Nanjing, China). Subsequently,
the NR8383 cells were stably transfected with the vector using
Lipofectamine 2000 and incubated in Ham's F-12K medium containing
0.1 mg/ml blasticidin (Invitrogen; Thermo Fisher Scientific, Inc.).
The transfected cells were then cultured for 24 h prior to
treatment with A. fumigatus (107 spores per well)
for 60 min at 37°C, and then harvested for examination.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) analysis
Total RNA was extracted using TRIzol reagent
(Invitrogen; Thermo Fisher Scientific, Inc.) according to the
manufacturer's protocol. The samples (500 ng) were
reverse-transcribed into cDNA using a RevertAid First Strand cDNA
Synthesis kit (Thermo Fisher Scientific, Inc.) through incubating
for 5 min at 25°C followed by 60 min at 42°C. qPCR was performed
using the SYBR Green PCR Master Mix kit (Takara Biotechnology Co.,
Ltd.), with the following steps: 95°C for 5 min, followed by 40
cycles of 95°C for 10 sec and 60°C for 40 sec. The primers for
miR-344b-1-3p, Tlr2, glyceraldehyde-3-phosphate
dehydrogenase (Gapdh) and U6 were designed and
obtained from Sangon Biotech Co., Ltd. (Shanghai, China). The
primer sequences are listed in Table
I. The expression levels of the Tlr2 mRNA and
miR-344b-1-3p were normalized to Gapdh and U6,
respectively, using the 2−ΔΔCt method (21).
 | Table I.Primer pairs for the reverse
transcription-quantitative polymerase chain reaction analysis. |
Table I.
Primer pairs for the reverse
transcription-quantitative polymerase chain reaction analysis.
| Name | Forward primer
(5′-3′) | Reverse primer
(5′-3′) |
|---|
| GAPDH |
AATGCATCCTGCACCACCAA |
GATGGCATGGACTGTGGTCA |
| U6 |
CGCTTCACGAATTTGCGTGTCAT |
GCTTCGGCAGCACATATACTAAAAT |
| miR-344b-1-3p |
GGGGTGATATAACCAAAGCC |
GTGCGTGTCGTGGAGTCG |
| TLR2 |
TGCAGGGACCTTTGCTATGATG |
ACAAAGTCCCGCTTGTGGGA |
Western blot assay
Western blot analysis was performed using samples
from whole-cell lysates processed using a Total Protein Extraction
kit (Beyotime Institute of Biotechnology; Beijing, China) and
quantified with a BCA Protein Assay kit. Following heat
denaturation at 95°C for 5 min, the samples of total protein (20
µg) were loaded onto sodium dodecyl sulfate-polyacrylamide gel
electrophoresis gels (5%), electrophoresed and transferred onto
polyvinylidene difluoride membranes (EMD Millipore; Billerica, MA,
USA). The membranes were blocked in 5% nonfat milk for 2 h at room
temperature, and were then incubated overnight at 4°C with one of
the following primary antibodies: TLR2 (cat. no. EPR20303; Abcam,
Cambridge, MA, USA), ATG5 (cat. no. 12994), Beclin-1 (cat. no.
3738), LC3 (cat. no. 4108) and GAPDH (cat. no. 5174) from Cell
Signaling Technology, Inc. (Danvers, MA, USA). The primary
antibodies were used in a 1:1,500 dilution. Following washing three
times with TBST, the membranes were incubated with secondary
antibody in a 1:10,000 dilution (cat. no. A1949; Sigma-Aldrich;
Merck KGaA) for 1 h at room temperature. Following a final wash,
signal was detected using enhanced chemiluminescence reagents (GE
Healthcare Life Sciences). The band intensities were quantified and
normalized to GAPDH using ImageJ 1.47 software (National Institutes
of Health).
Examination of autophagy by GFP-LC3
detection
The NR8383 cells were transfected in the manner
described above with the inclusion of GFP-LC3 (Invitrogen; Thermo
Fisher Scientific, Inc.). The cells transfected only with GFP-LC3
were incubated with A. fumigatus (107 spores per
well) for the indicated duration (0, 30, 45, 60, 90 or 120 min).
The co-transfected cells were incubated with A. fumigatus
spores for 60 min. The cells were then incubated with DAPI (5
µg/ml; Beyotime Institute of Technology) for 10 min. Following
fixing of the cells in 4% paraformaldehyde for 30 min at room
temperature, the cells were visualized by confocal microscopy.
Statistical analysis
Statistical analyses were performed using SPSS
(version 17.0; SPSS, Inc., Chicago, IL, USA). All data are
expressed as the mean ± SEM, with each assay performed in
triplicate. The statistical significance was estimated by using
one-way analysis of variance followed by Bonferroni's post hoc
test. P<0.05 was considered to indicate a statistically
significant difference.
Results
A. fumigatus spores induce a reduction
in miR-344b-1-3p and an increase in TLR2, LC-I, LC-II, beclin-1 and
ATG-5
To confirm the association between miR-344b-1-3p and
TLR2, their expression at the transcriptional level was determined
by RT-qPCR analysis. As shown in Fig.
1A, the levels of miRNA-344b-1-3p decreased in a time-dependent
manner following exposure to the spores, reaching a plateau after
90 min of treatment. By contrast, a corresponding time-dependent
increase was observed in the mRNA expression of Tlr2
following exposure to the spores, reaching a plateau after 90 min
of treatment (Fig. 1B). A similar
effect on the protein expression of TLR2 was observed (Fig. 1C and D). These data show that the
inverse correlation between the mRNA levels of Tlr2 and
miR-344b-1-3p may indicate that TLR2 is a target of
miR-344b-1-3p.
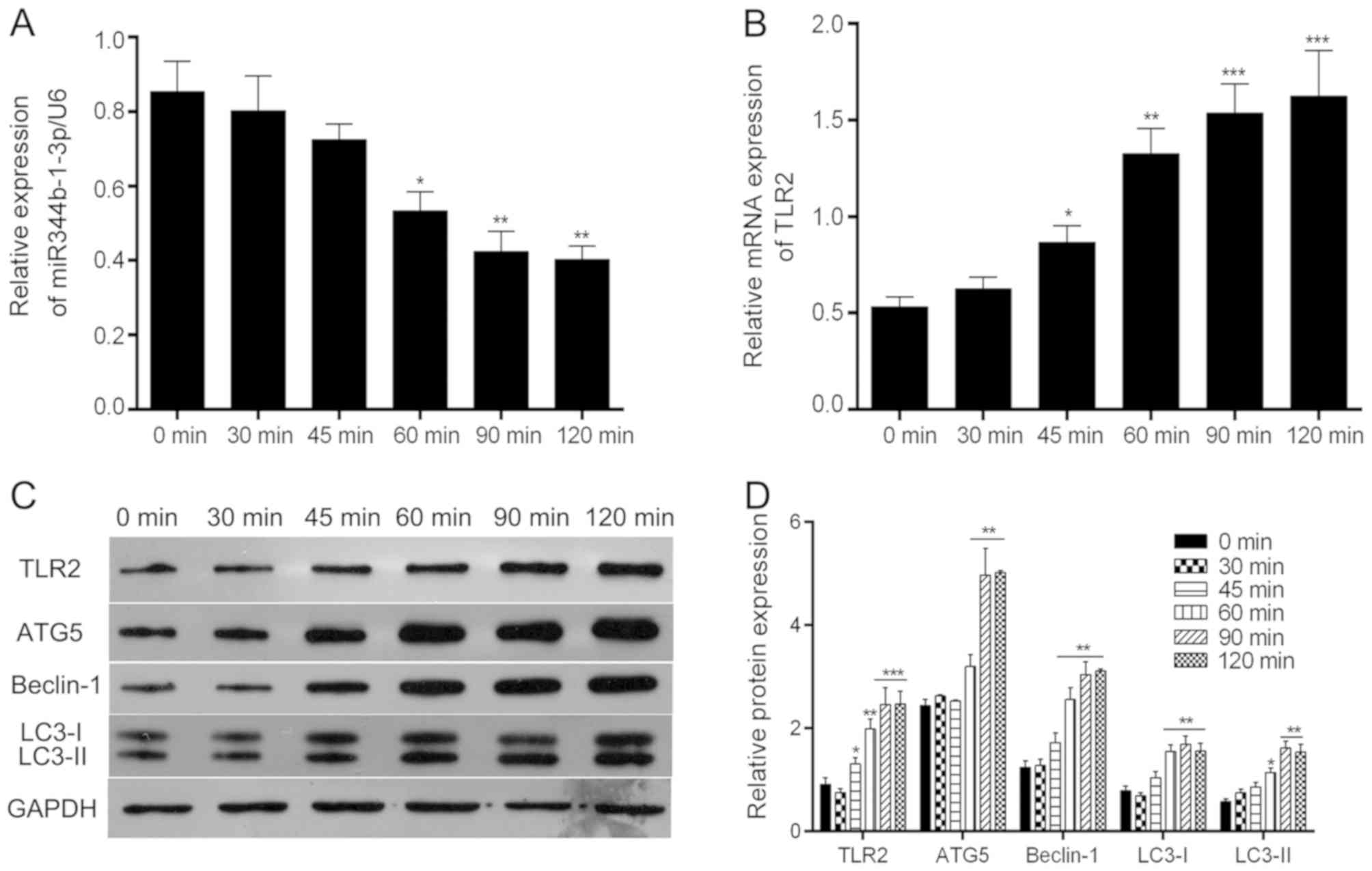 | Figure 1.Effect of A. fumigatus
infection on the expression of miR-344a-1-3p and TLR2. Following
exposure to A. fumigatus for the indicated times, NR8383
cells were harvested. Total RNA was extracted, and (A)
miR-344p-1-3a and (B) Tlr2 mRNA were detected by reverse
transcription-quantitative polymerase chain reaction analysis. (C)
Total protein was isolated, and TLR2, ATG5, Beclin-1, LC3-I and
LC-II were analyzed by western blot analysis and normalized to
GAPDH. (D) Quantification of protein expression. *P<0.05,
**P<0.01 and ***P<0.001 compared with the uninfected group.
A. fumigatus, Aspergillus fumigatus. miR, microRNA; TLR2,
toll-like receptor 2; ATG5, autophagy-related protein 5; LC3, light
chain 3. |
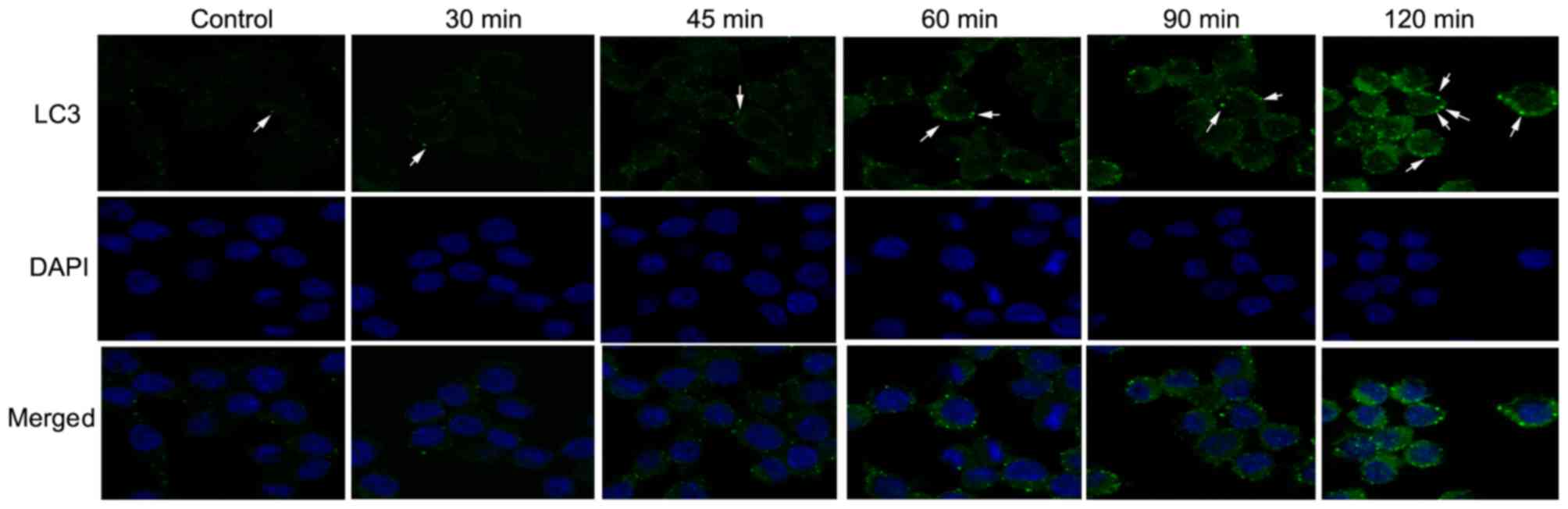
The immunofluorescence analysis showed that the
number of GFP-LC3 puncta increased in a time-dependent manner
following infection (Fig. 2),
indicating that the presence of A. fumigatus spores induced
autophagy time-dependently within 120 min of exposure. Furthermore,
the expression levels of each of the above-mentioned proteins were
negatively and positively associated with the levels of
miR-344b-1-3p and TLR2, respectively. These data indicate that the
expression of autophagy-related proteins and autophagy itself may
be mediated by miR-344b-1-3p and TLR2.
miR-344b-1-3p significantly suppresses
the autophagy induced by A. fumigatus spores
Following transfection with the NC mimic, inhibitor
NC, mimics or inhibitor, the cells were exposed to A.
fumigatus for 60 min to observe the effect of miR-344b-1-3p on
spore-induced autophagy. The 60-min exposure time was selected to
allow time for autophagy to initiate and also avoid the saturation
response. To demonstrate the transfection effects of miR-344b-1-3p
inhibitor or mimics, the expression of miR-344b-1-3p was measured
by RT-qPCR assay. The results showed that the expression of
miR-344b-1-3p was significantly increased in the miR-344b-1-3p
mimics group, and was significantly decreased in the miR-344b-1-3p
inhibitor group, compared with that in the NC groups, which
indicates high transfection efficiency (Fig. 3A). In addition, the expression levels
of ATG5, Beclin-1, LC3-1 and LC3-II were determined by western
blotting. As shown in Fig. 3B, the
expression levels significantly decreased following mimics
treatment and significantly increased following inhibitor
treatment. Similarly, the density of GFP-LC3 puncta markedly
decreased and increased following treatment with mimics and
inhibitor, respectively (Fig. 3C).
These results indicate that miR-344b-1-3p mimics treatment
significantly decreased spore-induced autophagy and that the
inhibition of miR-344b-1-3p significantly increased autophagy.
Consequently, miR-344b-1-3p may inhibit A. fumigatus
autophagy by regulating the levels of ATG5, Beclin-1 and LC3.
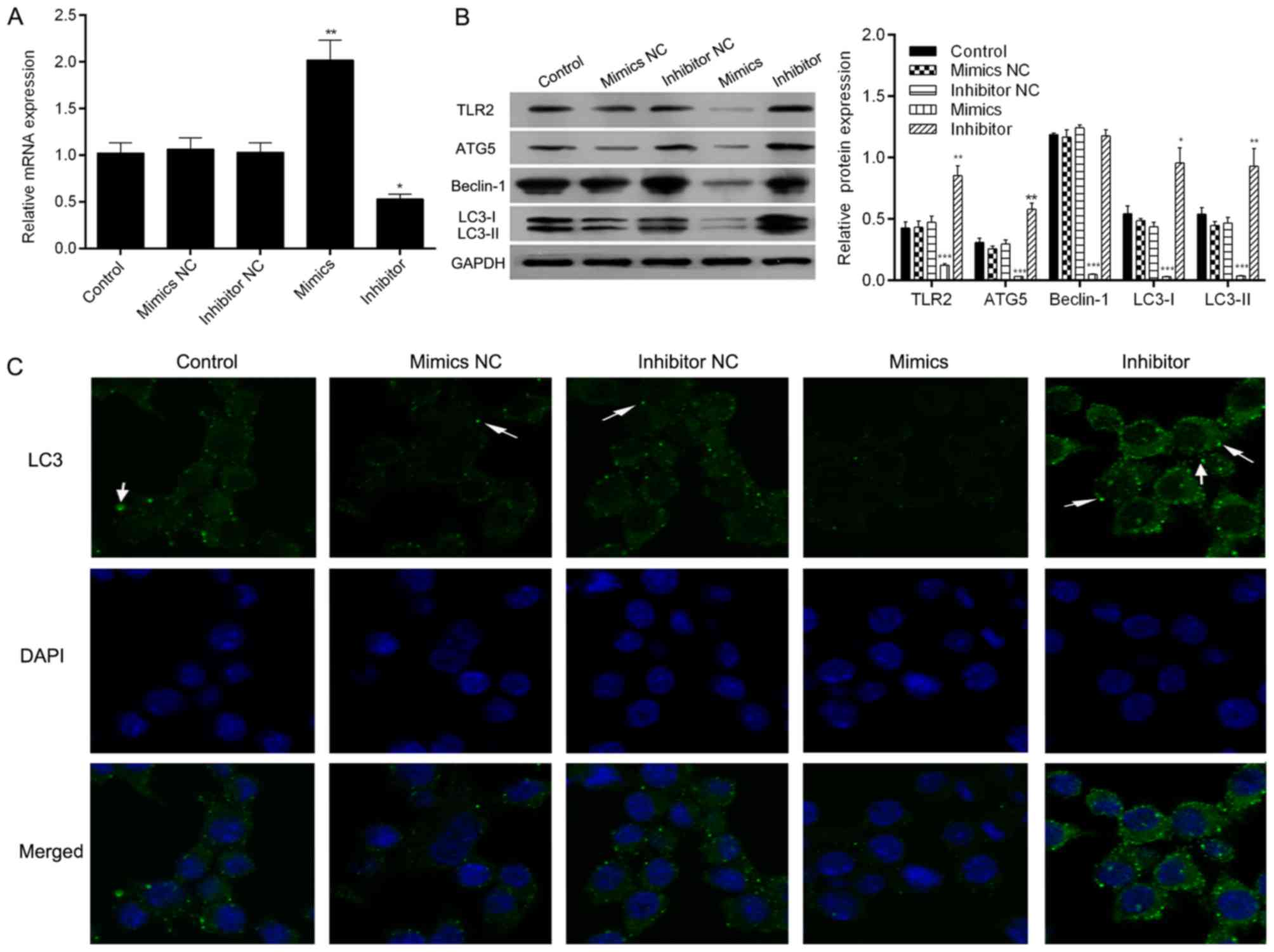 | Figure 3.Effect of miR-344p-1-3p knockdown on
the autophagy of NR8383 cells induced by A. fumigatus
infection. Cells were transfected with miR-344p-1-3a mimics NC,
inhibitor NC, mimics, or inhibitor, and then infected with A.
fumigatus for 60 min. (A) Expression of miR-344p-1-3p was
evaluated via reverse transcription-quantitative polymerase chain
reaction assay in each group. (B) Levels of ATG5, Beclin-1, LC3-I,
LC-II and TLR2 were analyzed by western blot analysis and
normalized to those of GAPDH. *P<0.05, **P<0.01 and
***P<0.001 compared with the uninfected group. (C) Cells were
visualized by detecting GFP-LC3 (green). Nuclei were stained with
DAPI (blue) and analyzed by confocal microscopy. The control group
received the same A. fumigatus treatment as the other groups
(magnification, ×400; arrows indicate GFP-LC3 puncta). A.
fumigatus, Aspergillus fumigatus; miR, microRNA; NC, negative
control; TLR2, toll-like receptor 2; ATG5, autophagy-related
protein 5; LC3, light chain 3. |
Additionally, the levels of TLR2 were determined by
western blot analysis; the levels were markedly decreased following
mimics exposure and markedly increased following inhibitor exposure
(Fig. 4A and B), indicating that
TLR2 is a target of miR-344b-1-3p.
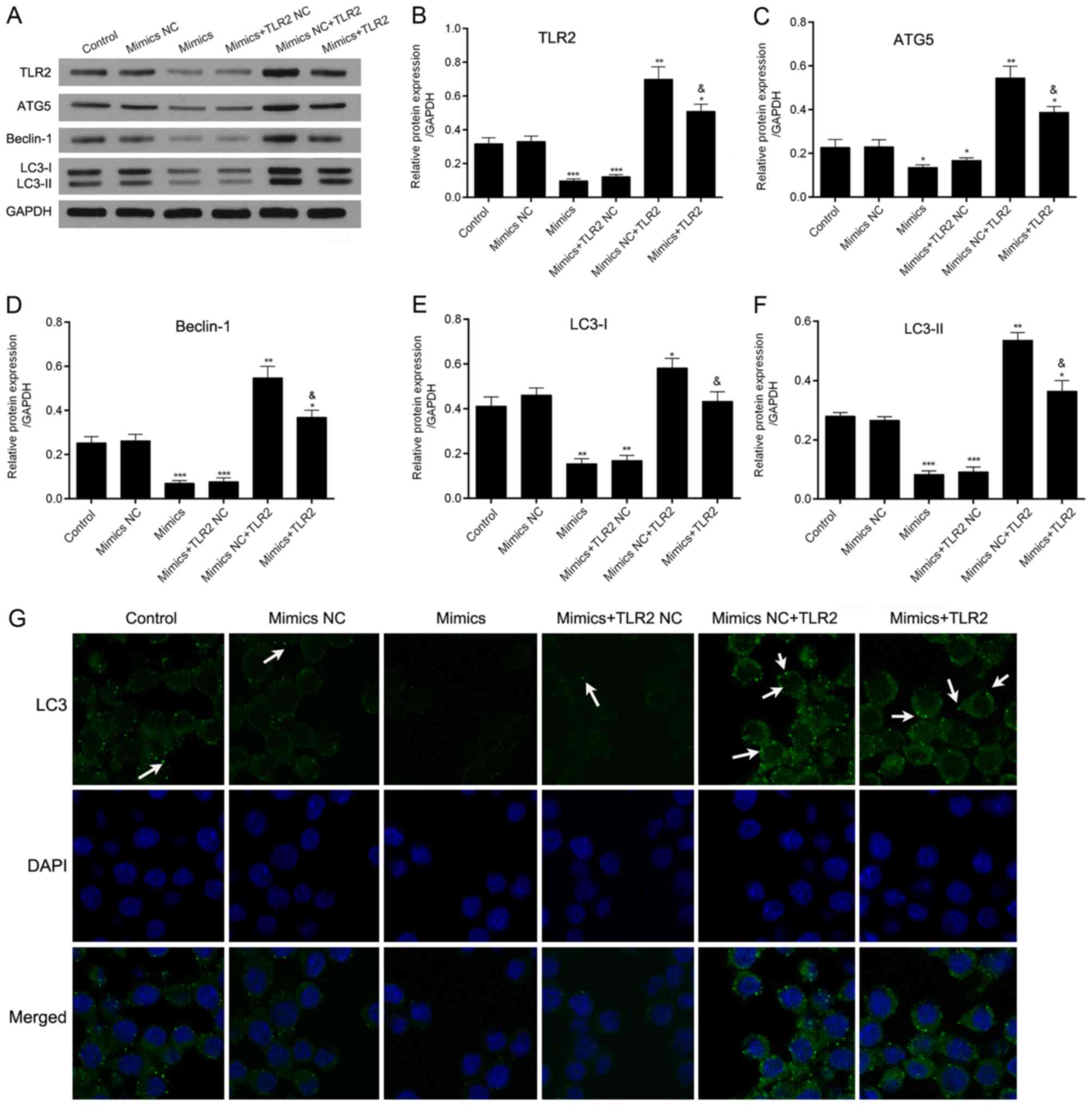 | Figure 4.Effect of the overexpression of
miR-344b-1-3p on the autophagy of A. fumigatus-infected
NR8383 cells by TLR2. Cells were transfected with miR-344b-1-3p
mimics and/or TLR2. The cells were then infected with A.
fumigatus for 60 min. (A) Levels of TLR2, ATG5, Beclin-1, LC3-I
and LC-II were analyzed by western blot analysis, GAPDH was used as
loading control. Relative protein expression levels of (B) TLR2,
(C) ATG5, (D) Beclin-1, (E) LC3-I (F) and LC-II were analyzed based
on the protein gray values. *P<0.05, **P<0.01 or
***P<0.001 compared with the control group;
&P<0.05 compared with the mimics NC + TLR2 group.
(G) Cells were evaluated for autophagy by the detection of GFP-LC3
(green). Nuclei were stained with DAPI (blue) and analyzed by
immunofluorescence microscopy (magnification, ×400; arrows indicate
GFP-LC3 puncta). A. fumigatus, Aspergillus fumigatus; miR,
microRNA; NC, negative control; TLR2, toll-like receptor 2; ATG5,
autophagy-related protein 5; LC3, light chain 3. |
miR-344b-1-3p inhibits the autophagy
induced by A. fumigatus spores by regulating TLR2
To investigate the roles of miR-344b-1-3p and TLR2
in the autophagy induced by A. fumigatus spores, cells were
transfected with one of the following: Control, mimics NC, mimics,
mimics + TLR2 NC, mimics NC + TLR2 and mimics + TLR2. The results
from the western blot assay revealed that the expression levels of
TLR2, Beclin-1, ATG5, LC3-I and LC3-II were significantly decreased
in cells transfected with miR-344b-1-3p mimics compared with those
in the NC group. The expression levels of TLR2, Beclin-1, ATG5,
LC3-I and LC3-II were significantly increased in cells transfected
with mimics NC + TLR2 compared with those in cells in the mimics NC
group. The expression levels of TLR2, Beclin-1, ATG5, LC3-I and
LC3-II were markedly increased in the cells transfected with mimics
+ TLR2 compared with those in the mimics group (Fig. 4A-F). Similarly, the number of GFP-LC3
puncta in the cells was significantly decreased following
miR-344b-1-3p mimics transfection, whereas the overexpression of
TLR2 markedly attenuated the inhibitory effect on autophagy by
miR-344b-1-3p (Fig. 4G). Therefore,
these data indicated that miR-344b-1-3p may serve an important role
in Aspergillus spore-induced autophagy by regulating
TLR2.
Discussion
miRNAs are critical to multiple facets of immune
system function, including pathogen recognition, inflammation
activation and resolution. TLR2, an important pathogen recognition
receptor, is targeted and regulated by miR-344b-1-3p, however, its
role in regulation of immune function remains to be elucidated. To
the best of our knowledge, miR-344b-1-3p was first associated with
A. fumigatus-induced autophagy in macrophages, and may act
by mediating recognition of the fungus.
Accumulating evidence suggests that autophagy is
closely associated with immunity. A. fumigatus infection can
elicit autophagy in several types of inflammatory cell, including
monocytes and macrophages, and regulate their physiological
activity (22,23). Studies have shown that autophagy
directly affects the production of certain cytokines by mediating
transcription, processing and secretion (24). These cytokines, which include
interleukin-1β (IL-1β), IFN-γ, and tumor necrosis factor (TNF)-α,
in turn dampen pro-inflammatory responses (25,26).
Autophagy also accelerates macrophage aging, resulting in various
functional changes, including impaired maturation, decreased
antigen presentation capacity and reduced innate responses,
alongside increased basal production of inflammatory cytokines
(27). Additionally, autophagy
triggers the death of activated macrophages through a
caspase-independent pathway (28).
Autophagy exerts dual functionality in macrophages,
pro-inflammation and anti-inflammation, to achieve host protection
and intracellular pathogen elimination.
Autophagy is both provoked and regulated. In its
recognition of A. fumigatus, TLR2 serves a critical role in
autophagy-induction in infected macrophages (14). In RAW264.7 cells, this induction is
reached by activating multiple signaling pathways, including the
JNK, ERK, and PI3K signaling pathways (29–31).
Although the fungus used in these studies was not A.
fumigatus, the same pathways were suggested to contribute to
the autophagy in macrophages induced by A. fumigatus. Among
the multiple functions of autophagy-induction, increased levels of
pro-inflammatory cytokines have been reported. The cytokines,
including IL-1β and TNF-α, can in turn upregulate autophagy,
forming a positive feedback loop (24).
The ability of TLR2 to detect Aspergillus
spores and induce autophagy is associated with its expression
levels. Wu et al reported that induced expression of TLR2 by
the fungus was noted in human corneal epithelial cells and mice
macrophages (32,33). Consistent with these findings, in the
present study, A. fumigatus exposure significantly increased
the expression of TLR2 in rat alveolar cells, which was accompanied
by the induction of autophagy and a decrease in the levels of
miR-344b-1-3p. In addition, treatment with miRNA mimics in the
present study attenuated the induction of TLR2 and autophagy.
miR-344b-1-3p has been demonstrated to directly target TLR2
(20), suggesting a mechanism by
which the miRNA may mediate autophagy. Additionally, targeting TLR2
with miR-344b-1-3p inhibited the expression of downstream genes,
including TNF-α and IL-1β (20). In
addition to the regulatory role of cytokines in autophagy,
miR-344b-1-3p may also regulate autophagy in macrophages exposed to
fungi by inhibiting cytokines. In addition to miR-344b-1-3p, miR-19
and miR-105 have been demonstrated to regulate the expression of
TLR2 (34,35), indicating they may be associated with
the activation of autophagy in macrophages. However, this requires
confirmation by further investigations.
Taken together, the results of the present study
demonstrate that the levels of miR-344b-1-3p decreased following
challenge by A. fumigatus, which enhanced the activation of
autophagy in infected macrophages by targeting TLR2 to protect
against infection by intracellular mycobacteria. The present study
reveals the importance of an miRNA in the regulation of autophagy
and elimination of A. fumigatus. At present, miR-344b-1-3p
has only been examined in rats. There is evidence to show that
miR-344b-1-3p is an effective modulator of the TLR2 gene in COPD
rats (1). It was also found that
miR-344b-1-3p cannot be detected in Homo sapiens. In the
future, there may be potential therapeutic interventions for
saprophytic aspergillosis, allergic aspergillosis and invasive
aspergillosis by artificially synthesizing miR-344b-1-3p and the
targeted drugs.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Natural
Science Foundation of Guangdong Province (grant no
2014A030313597).
Availability of data and materials
The analyzed datasets generated during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
All authors designed the study. YW, HX, YL, DH
performed the experiments. YW, HX, YL, LC, YH and LL collected and
analyzed the data. YW wrote the manuscript. DZ and WH provided
advice on the experimental design and critically revised the
manuscript. All authors read and approved the final version of the
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Morton CO, Bouzani M, Loeffler J and
Rogers TR: Direct interaction studies between Aspergillus
fumigatus and human immune cells; what have we learned about
pathogenicity and host immunity? Front Microbiol. 3:4132012.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Roilides E, Sein T, Schaufele R, Chanock
SJ and Walsh TJ: Increased serum concentrations of interleukin-10
in patients with hepatosplenic candidiasis. J Infect Dis.
178:589–592. 1998. View
Article : Google Scholar : PubMed/NCBI
|
|
3
|
Chotirmall SH, Al-Alawi M, Mirkovic B,
Lavelle G, Logan PM, Greene CM and McElvaney NG:
Aspergillus-associated airway disease, inflammation, and the innate
immune response. Biomed Res Int. 2013:7231292013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Schneemann M and Schaffner A: Host defense
mechanism in Aspergillus fumigatus infections. Contrib
Microbiol. 2:57–68. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Segal BH and Walsh TJ: Current approaches
to diagnosis and treatment of invasive aspergillosis. Am J Respir
Crit Care Med. 173:707–717. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Weinberger M, Elattar I, Marshall D,
Steinberg SM, Redner RL, Young NS and Pizzo PA: Patterns of
infection in patients with aplastic anemia and the emergence of
Aspergillus as a major cause of death. Medicine (Baltimore).
71:24–43. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Duong M, Ouellet N, Simard M, Bergeron Y,
Olivier M and Bergeron MG: Kinetic study of host defense and
inflammatory response to Aspergillus fumigatus in
steroid-induced immunosuppressed mice. J Infect Dis. 178:1472–1482.
1998. View
Article : Google Scholar : PubMed/NCBI
|
|
8
|
Bodey GP and Vartivarian S: Aspergillosis.
Eur J Clin Microbiol Infect Dis. 8:413–437. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kurup VP and Kumar A: Immunodiagnosis of
aspergillosis. Clin Microbiol Rev. 4:439–456. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Latgé JP: Aspergillus fumigatus and
aspergillosis. Clin Microbiol Rev. 12:310–350. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Bellocchio S, Montagnoli C, Bozza S,
Gaziano R, Rossi G, Mambula SS, Vecchi A, Mantovani A, Levitz SM
and Romani L: The contribution of the Toll-like/IL-1 receptor
superfamily to innate and adaptive immunity to fungal pathogens in
vivo. J Immunol. 172:3059–3069. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Mambula SS, Sau K, Henneke P, Golenbock DT
and Levitz SM: Toll-like receptor (TLR) signaling in response to
Aspergillus fumigatus. J Biol Chem. 277:39320–39326. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Chai LY, Kullberg BJ, Vonk AG, Warris A,
Cambi A, Latgé JP, Joosten LA, van der Meer JW and Netea MG:
Modulation of Toll-like receptor 2 (TLR2) and TLR4 responses by
Aspergillus fumigatus. Infect Immun. 77:2184–2192. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Anand PK, Tait SW, Lamkanfi M, Amer AO,
Nunez G, Pagès G, Pouysségur J, McGargill MA, Green DR and
Kanneganti TD: TLR2 and RIP2 pathways mediate autophagy of Listeria
monocytogenes via extracellular signal-regulated kinase (ERK)
activation. J Biol Chem. 286:42981–42991. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kanayama M and Shinohara ML: Roles of
autophagy and autophagy-related proteins in antifungal immunity.
Front Immunol. 7:472016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Tanida I, Ueno T and Kominami E: LC3 and
autophagy. Methods Mol Biol. 445:77–88. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kang R, Zeh HJ, Lotze MT and Tang D: The
Beclin 1 network regulates autophagy and apoptosis. Cell Death
Differ. 18:571–580. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Mihalache CC and Simon HU: Autophagy
regulation in macrophages and neutrophils. Exp Cell Res.
318:1187–1192. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhu H, Wu H, Liu X, Li B, Chen Y, Ren X,
Liu CG and Yang JM: Regulation of autophagy by a beclin 1-targeted
microRNA, miR-30a, in cancer cells. Autophagy. 5:816–823. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Xu H, Wu Y, Li L, Yuan W, Zhang D, Yan Q,
Guo Z and Huang W: MiR-344b-1-3p targets TLR2 and negatively
regulates TLR2 signaling pathway. Int J Chron Obstruct Pulmon Dis.
12:627–638. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Xu H, Fei D, Zong S and Fan Z:
MicroRNA-154 inhibits growth and invasion of breast cancer cells
through targeting E2F5. Am J Transl Res. 8:2620–2630.
2016.PubMed/NCBI
|
|
22
|
Kyrmizi I, Gresnigt MS, Akoumianaki T,
Samonis G, Sidiropoulos P, Boumpas D, Netea MG, van de Veerdonk FL,
Kontoyiannis DP and Chamilos G: Corticosteroids block autophagy
protein recruitment in Aspergillus fumigatus phagosomes via
targeting dectin-1/Syk kinase signaling. J Immunol. 191:1287–1299.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
de Luca A, Smeekens SP, Casagrande A,
Iannitti R, Conway KL, Gresnigt MS, Begun J, Plantinga TS, Joosten
LA, van der Meer JW, et al: IL-1 receptor blockade restores
autophagy and reduces inflammation in chronic granulomatous disease
in mice and in humans. Proc Natl Acad Sci USA. 111:3526–3531. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Harris J: Autophagy and cytokines.
Cytokine. 56:140–144. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Harris J, Hartman M, Roche C, Zeng SG,
O'Shea A, Sharp FA, Lambe EM, Creagh EM, Golenbock DT, Tschopp J,
et al: Autophagy controls IL-1beta secretion by targeting
pro-IL-1beta for degradation. J Biol Chem. 286:9587–9597. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Crisan TO, Plantinga TS, van de Veerdonk
FL, Farcaş MF, Stoffels M, Kullberg BJ, van der Meer JW, Joosten LA
and Netea MG: Inflammasome-independent modulation of cytokine
response by autophagy in human cells. PLoS One. 6:e186662011.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Stranks AJ, Hansen AL, Panse I, Mortensen
M, Ferguson DJ, Puleston DJ, Shenderov K, Watson AS, Veldhoen M,
Phadwal K, et al: Autophagy controls acquisition of aging features
in macrophages. J Innate Immun. 7:375–391. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Xu Y, Kim SO, Li Y and Han J: Autophagy
contributes to caspase-independent macrophage cell death. J Biol
Chem. 281:19179–19187. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Fang L, Wu HM, Ding PS and Liu RY: TLR2
mediates phagocytosis and autophagy through JNK signaling pathway
in Staphylococcus aureus-stimulated RAW264.7 cells. Cell Signal.
26:806–814. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Lu Z, Xie D, Chen Y, Tian E, Muhammad I,
Chen X, Miao Y, Hu W, Wu Z, Ni H, et al: TLR2 mediates autophagy
through ERK signaling pathway in Mycoplasma gallisepticum-infected
RAW264.7 cells. Mol Immunol. 87:161–170. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Shen Y, Kawamura I, Nomura T, Tsuchiya K,
Hara H, Dewamitta SR, Sakai S, Qu H, Daim S, Yamamoto T and
Mitsuyama M: Toll-like receptor 2- and MyD88-dependent
phosphatidylinositol 3-kinase and Rac1 activation facilitates the
phagocytosis of Listeria monocytogenes by murine macrophages.
Infect Immun. 78:2857–2867. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Wu J, Zhang Y, Xin Z and Wu X: The
crosstalk between TLR2 and NOD2 in Aspergillus fumigatus
keratitis. Mol Immunol. 64:235–243. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Reza KA, Noushin S, Zuhair H, Mehdi M, Ali
AA, Majid T, Hojjatollah S and Hoseinali EM: Evaluation of the
expression of TLR-2, Dectin-1 and TNF-α level in invasive
aspergillosis in cancer mice. Comparative Clin Pathol. 19:601–605.
2010. View Article : Google Scholar
|
|
34
|
Philippe L, Alsaleh G, Suffert G, Meyer A,
Georgel P, Sibilia J, Wachsmann D and Pfeffer S: TLR2 expression is
regulated by microRNA miR-19 in rheumatoid fibroblast-like
synoviocytes. J Immunol. 188:454–461. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Benakanakere MR, Li Q, Eskan MA, Singh AV,
Zhao J, Galicia JC, Stathopoulou P, Knudsen TB and Kinane DF:
Modulation of TLR2 protein expression by miR-105 in human oral
keratinocytes. J Biol Chem. 284:23107–23115. 2009. View Article : Google Scholar : PubMed/NCBI
|